
Abstract
Small extracellular vesicles (sEVs) are cell-derived, nanometer-sized particles enclosed by a lipid bilayer. All kinds of biological molecules, including proteins, DNA fragments, RNA, lipids, and metabolites, can be selectively loaded into sEVs and transmitted to recipient cells that are near and distant. Growing shreds of evidence show the significant biological function and the clinical significance of sEVs in cancers. Numerous recent studies have validated that sEVs play an important role in tumor progression and can be utilized to diagnose, stage, grading, and monitor early tumors. In addition, sEVs have also served as drug delivery nanocarriers and cancer vaccines. Although it is still infancy, the field of basic and translational research based on sEVs has grown rapidly. In this review, we summarize the latest research on sEVs in gliomas, including their role in the malignant biological function of gliomas, and the potential of sEVs in non-invasive diagnostic and therapeutic approaches, i.e., as nanocarriers for drug or gene delivery and cancer vaccines.
Similar content being viewed by others
Introduction
Glioma, the most common primary tumor of the central nervous system (CNS), is classified into four grades. Among them, the grade IV glioma, also called glioblastoma (GBM), is the most malignant and characterized by high invasiveness. Worldwide, there are about 100,000 people diagnosed with glioma every year, and GBM accounts for 70–75% of all diffuse glioma diagnoses [1, 2]. However, the prognosis of GBM patients is unsatisfactory, and the average survival is only 12–15 months. Moreover, in cancer patients between 15 and 34 years, GBM is the third most common cause of death [3, 4]. At present, the standard treatment of GBM is surgery and subsequent chemoradiotherapy. However, due to high heterogeneity and invasiveness, the outcome of GBM patients is still dismal [5,6,7]. Thus, in order to improve the survival rate and quality of life of glioma patients, more effective treatments and highly accurate and noninvasive tumor biomarkers which can identify the glioma even in its early stages are urgently needed.
sEVs released by most cells are essential to intercellular connections and pathophysiology. In recent years, research on sEVs has grown due to their unique structure and function. Previous studies showed that sEVs can transfer proteins and nucleic acids from the original cell to the target cells and then have biological effects on them. Regarding tumors themselves, growing evidence suggests that sEVs deriving from tumors participate in many physiological and pathological processes, including tumor microenvironment (TME), angiogenesis, epithelial-to-mesenchymal transition (EMT), immune regulation, metastasis, and therapeutic resistance [8,9,10].
Recently, many studies have verified that sEVs can be utilized for early diagnosis, staging, grading, and treatment monitoring of cancers [11, 12]. In addition, sEVs show potential as nanocarriers for cancer vaccines and drug delivery systems. sEVs have the advantages of specificity, safety, stability, and ability to penetrate the blood–brain barrier. These findings raise important clinical implications and highlight the need to further explore sEVs-based diagnostic and therapeutic approaches, especially in gliomas.
In this review, we primarily discuss the effects of sEVs on glioma progression, angiogenesis, metastasis, TME, resistance to treatment, liquid biopsy, and treatment. We hope to inspire readers to further explore the role of sEVs in glioma and that leveraging these properties may open new avenues to address the complexities of glioma diagnosis and treatment.
The origin and characteristics of extracellular vesicle (EVs)
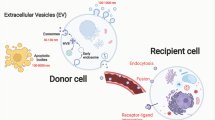
In 1983, John Stone et al. researched the transformation of sheep reticulocytes to mature erythrocytes and observed that the erythrocytes release transferrin metabolites by secreting small vesicles that were later called exosomes [13]. The secretion pattern is shown in Fig. 1. First, the plasma membrane buds inward and generates early endosomes, which are then processed into multivesicular bodies (MVBs) [14]. MVBs involve in endocytosis and transporting intracellular substances. There are two endings of MVBs. The lysosome degrades one. The other is fusing with the cell membrane and then releasing outside the cell in the form of exosomes [15]. Regarding exosome budding, the most accepted hypothesis is the endosomal sorting complex required for transport (ESCRT) family catalytic [16]. However, exosome biogenesis is not substantial reduction when the activity of the ESCRT family is inhibited [17], and researchers have discovered ESCRT-independent mechanisms affecting the production of exosomes, such as heterogeneous nuclear ribonucleoprotein-dependent pathway, neutral sphingomyelinase 2-dependent pathway, and a recently discovered novel pathway marked by RAB31 [18, 19]. ESCRT-dependent and independent secretory pathways are shown in Figure S1 [20]. With the deepening understanding of exosome biogenesis, other mechanisms may be discovered in the future.

The biogenesis and cargo loading of sEVs. Cells mainly secrete three types of EVs, including microvesicles, apoptotic bodies and exsomes. Exsomes are formed in the endosomal compartment of the cell by inward budding of limiting endosome membrane into exosomal precursors, intraluminal vesicles (ILVs). Endosomal multivesicular bodies (MVBs) containing accumulated ILVs can then be degraded by fusing with lysosomes or autophagosomes or released to extracellular matrix as exosomes via ESCRT-independent or ESCRT-dependent mechanisms. The main contents of exosomes include miRNA, non-coding RNA, protein, etc
Exosomes are lipid bilayer membrane vesicles ~ 30–150 nm in diameter and 1.13–1.21 g/ml in density [21]. Almost all human cells can secrete exosomes which widely exist in various body fluids, including blood, urine, semen, cerebrospinal fluid, tears, saliva, breast milk, bile, ascites, lymph, and amniotic fluid [22]. Other EVs mainly include microvesicles and apoptotic bodies. Microvesicles also referred to as exosomes, derive from the plasma membrane and are 100–1000 nm in size. Studies have shown that microvesicles have largely similar functions to exosomes [23]. Apoptotic bodies, also known as apoptosomes, are vesicles with a 500–5000 nm diameter and are released by cells undergoing apoptosis [24]. The function of EVs mainly relies on their loaded cargo, including proteins and mRNAs. The remaining components consist of long noncoding RNAs (lncRNAs), microRNAs (miRNAs), DNAs, and circular RNAs (circRNAs) [25,26,27,28,29,30].
At present, it is hard to put forward specific and widely recognized markers of "exosomes" derived from MVB compared to other small EVs due to the lack of an optimal isolation approach and specific markers for EVs from different sources. In general, the term "exosomes" is used to refer to a heterogenous mixture of EVs that are less than 200 nm in diameter. Thus, it contains exosomal and non-exosomal particles [30,31,32]. According to the MISEV2018 guidelines, we substitute "small extracellular vesicles" for "exosomes" [9].
Separation, detection and storage of sEVs
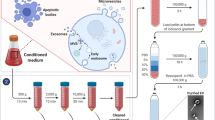
The premise of sEVs research is to isolate sEVs at high purity and in sufficient quantities. Whereas various separation methods have been systematically introduced in other studies [33, 34], we briefly summarize the methods in Table 1 which utilize density, size, membrane proteins, solubility, surface charge, and lipid membrane of sEVs and point out their merit and demerit in this review. Density Ultracentrifugation (UC) is the most widely used method for isolating sEVs with minimal reagents and can obtain large volumes. However, the heavy protein contamination and comparatively low throughput negatively impact further studies of these sEVs [35,36,37,38]. Density gradient ultracentrifugation (dgUC) is considered to be a gold standard, which can provide the highest purity of sEVs and enables researchers to isolate specific subpopulations. Despite this, the requirement of expensive equipment and rare recovery efficiency are significant disadvantages [39,40,41]. Acoustofluidics can maintain EVs integrity and require a minimal sample, but they may cause protein contamination [39, 42]. Solubility precipitation preserves sEVs integrity and has commercially available kits. However, the lowest purity has negative impacts on the downstream functional analyses [43,44,45]. Size ultrafiltration (UF) is easily integrated with other EVs isolation methods, but it is not as effective at producing pure samples as UC [46,47,48]. Immunoaffinity antibodies can provide very pure samples, but it is hard to harvest intact EVs from antibodies when downstream analyses require intact vesicles [49,50,51]. Moreover, charge dielectrophoresis allows for separating specific EVs subpopulations with good purity, but some disadvantages, including low yield and damaging EVs membranes, negatively impact the downstream analyses [52,53,54]. Size exclusion chromatography (SEC) isolates EVs via utilizing size, and the purity of separated EVs is equivalent to that of dgUC. However, SEC is not able to utilize to concentrate EVs samples. Aiming at this problem, researchers obviously improve the purification efficiency of EVs by combining SEC with UF [55, 56]. In addition, electrostatic interaction and lipid nanoprobes (LNP) utilize surface charges and their lipid bilayer to isolate EVs [57, 58]. These approaches can obtain EVs within a short time without ponderous equipment and allow to perform of downstream analyses and functional studies. However, protein contamination may influence the accuracy of downstream analysis [59]. In brief, it is impractical to isolate sEVs at present, but the combined application of various methods is still recommended.
Besides, it is also required to develop more efficient and economical methods to detect sEVs. Rapid advances in on-chip detection technologies make sEVs easier to discover. We briefly summarize the sEVs detection methods in Table 2 and point out their advantages and disadvantages. The ELISA-based on-chip detection method is compassionate and provides noticeable color variations to indicate the presence of the target with 2–4 h to finish detection [60, 61]. The lateral flow immunoassays (LFIA) method, Au-conjugated anti-CD9 is used as the capture line. Meanwhile, Au-conjugated anti-CD63 is used for the detection/control line. It is less sensitive than ELISA and takes about 2 h [62, 63]. In addition, several biosensors with fast and sensitive detection performance have been examined, including plasmon resonance and quantum dots (QDs), the readouts of which have compatibility with clinical settings [64,65,66,67]. Surface plasmon resonance (SPR), relying on light to excite electrons and then producing resonant oscillation currents, can be used to detect nanoparticles, including sEVs [68]. In 2018, Liu et al. developed a small SPR system to detect sEVs from lung cancer [65]. Moreover, as inorganic colloid tracers which are utilized in signal transduction labeling, QDs provide a sensitive way to detect sEVs [66]. In the future, sEVs may replace the current diagnostics, but before that, much work, including high specificity, low cost, and fast detection, needs to be fulfilled.
Until now, there has been no consensus on the storage of sEVs. In the study of Rosario Maroto et al., it was pointed out that repeated freezing-thawed processes are extremely destructive to sEVs structure and physical properties, and that sEVs stored at + 4 °C and -80 °C are different, and their analysis found that 756 (89%) proteins did not change in abundance due to changes in storage temperature. However, after storage at + 4 °C, 61 proteins were depleted; In contrast, 31 proteins were more abundant in exosomes stored at + 4 °C than in sEVs stored at -80 °C, suggesting that small amounts of sEVs proteins are more sensitive to storage temperature. Moreover, according to the different storage conditions, a small proteome appeared in the supernatant, 67 proteins were enriched in the sEVs supernatant stored at -80° C, and 78 unique proteins were enriched in the sEVs supernatant stored at + 4 °C. These proteins include cytokines (CXCL15, CC10) and serine proteases (Serpina1c and -1d). Another finding of the study was that storage conditions affect the proteomic content of airway sEVs. Analysis of sEVs in cell culture showed that storage at + 4 °C had a significant effect on CD63 and Hsp70 content, a finding that confirmed the relative depletion of CD63 in stored samples [69].
Biofunctions of sEVs in glioma
Numerous shreds of evidence have shown that sEVs mediate the occurrence and progression of various tumors by promoting intercellular communication, pro-inflammatory responses, and regulating the TME. At the same time, the function of sEVs in glioma has not been systematically reviewed. Here, we comprehensively show the biological function of sEVs in glioma (Fig. 2).

sEVs play vital roles in mediating glioma proliferation/invasion, angiogenesis, metastasis, immunosuppressive microenvironment, and treatment resistance
sEVs and cell proliferation/invasion in glioma
The most essential characteristics of malignant glioma are rapid proliferation and extensive invasion, which involve complex molecular regulation and dynamic crosstalk between the tumor and microenvironment. This leads to difficulty in complete surgical resection and postoperative recurrence [70]. Therefore, the identification of critical molecules and related mechanisms involved in the proliferation and invasion of tumors is necessary for the development of new therapeutic strategies. Glioma-derived sEVs have been reported to play an essential role in tumor proliferation and invasion by regulating intercellular communication in local and distant microenvironments. Bian et al. demonstrated that sEVs derived from glioma cells and transporting lncRNA-ATB can activate astrocytes via inhibition of miR-204-3p, leading to promotion of glioma cell invasion by the activated astrocytes [71]. LncRNA AHIF, the natural antisense transcript of hypoxia-inducible factor 1α (HIF 1α), has been reported to induce glioma cell proliferation, invasion, and radioresistance through sEVs, suggesting its potential as a therapeutic target [72,73,74]. Furthermore, Chai and colleagues have found that glioma tissues exhibit upregulated expression of lncRNA ROR1-AS1 in comparison to normal tissue, with high levels of lncRNA ROR1-AS1 indicating a poor prognosis. Mechanistic investigations have revealed that sEVs carrying lncRNA ROR1-AS1 derived from glioma cells can facilitate glioma progression by suppressing miR-4686 [75]. Ma et al. revealed that LINC00470 was up-regulated in serum sEVs from glioma patients and correlated with disease progression and postoperative survival of glioma patients, and sEVs-LINC00470 in GBM can bind to miR-580-3p in glioma cells to regulate WEE1 expression and activate the PI3K/AKT/mTOR pathway, thereby enhancing the proliferation of glioma cells [76].
In addition to lncRNAs, sEVs loading miRNAs or proteins are also crucial contributors to glioma progression. Xu and colleagues reported that sEVs-miR-375 derived from glioma cells was able to activate the CTGF-EGFR oncogenic pathway to promote glioma proliferation and invasion [77]. Research by Yang et al. found that sEVs-miR-221 induced tumor proliferation and migration in glioma via targeting DNM3 [78]. The latest research have shown that glioma stem cells (GSCs)-derived sEVs contribute importantly to the plasticity, heterogeneity, and aggressiveness of glioma [79]. GSCs-derived sEVs-miR-155-5p slao play a key role in enhancing the invasiveness of glioma cells by targeting ACOT12 and promoting mesenchymal transformation [80]. In the study of Li et al., sEVs- miR-3065-5p derived from GSCs induced astrocyte transformation through the miR-3065-5p/DLG2 signal transduction axis, further promoting the tumorigenesis of GSCs [81]. Immunoglobulin κ J region recombinant signal binding protein (RBP-J) is a transcriptional regulator indicating that sEVs-circBTG2 secreted by RBP-J overexpressed macrophages inhibits the proliferation and invasion of glioma cells through the circBTG2/miR-25-3p/PTEN pathway [82]. L1-cell adhesion molecule (L1CAM), the autocrine/paracrine of which is one of the factors that promote glioma cell proliferation, migration, and invasiveness [83, 84]. In another study, researchers isolated tumor cell sEVs packaging L1CAM and found these sEVs promoted glioma cell migration and proliferation. Furthermore, the results of in vivo experiment, which employed the white leghorn chicken embryo, were in accordance with in vitro experiment [85].
sEVs and glioma angiogenesis
Continuous growth is one of the main characteristics of malignant tumors. Angiogenesis being able to provide nutrition and oxygen, is an important factor in the sustained growth of most solid tumors [86]. Similarly, GBM, which has the worst prognosis, has been demonstrated to be the most vascularized tumor [87, 88]. Recently, many studies indicated that sEVs are involved in glioma angiogenesis. Lang and colleagues found that sEVs derived from glioma cells transferred lncRNA CCAT2 to endothelial cells. The angiogenesis-related genes, including VEGF, TGFβ, FGF, and KDR, were then activated to promote angiogenesis [89]. Another study reported that sEVs delivering lncRNA POU3F3 could promote glioma angiogenesis [90]. In addition, the glioma cells derived sEVs which packaged lncRNA HOTAIR, and miR-9, can be transferred to endothelial cells, both leading to increased angiogenesis [91, 92]. Other reports indicate that M2 polarized GBM-related microglia-derived sEV-circKIF18A participate in GBM angiogenesis by targeting FOXC 2 and that glioma secretion of circGLIS3-loaded sEVs can also induce endothelial cell angiogenesis [93, 94]. As a member of the scaffolding adaptor protein family, ERBB receptor feedback inhibitor 1 (ERRFI1) plays a vital role in the epidermal growth factor receptor (EGFR) signaling pathway [95]. Wang et al. reported that sEVs-miR-148a-3p secreted by glioma cells stimulated glioma angiogenesis via activating the EGFR/MAPK signaling pathway by ERRFI1 inhibition [96]. In GBM, increased miR-21 has the function of upregulating VEGF expression. Sun and colleagues found that GSCs-derived sEVs promoted the angiogenesis of endothelial cells via miR-21/VEGF/VEGFR2 signaling [97]. Moreover, sEVs-miR-26a, which were derived from GSCs, could also stimulate the angiogenesis of microvessel endothelial cells in glioma by targeting PTEN and further activating the PI3K/Akt signaling pathway [98]. In addition, GSCs-derived sEVs-miR-944 inhibited glioma progression and angiogenesis by inhibition of VEGFC expression and inhibition of AKT/ERK signaling pathway [99]. Unlike the above, Zhang et al. reported that sEVs-miR-29a-3p derived from engineered human mesenchymal stem cells suppressed glioma angiogenesis [100].
Wang and colleagues found a unique form of VEGF with a length of 120-kDa (VEGF-120 kDa) existing in sEVs which were derived from GBM cells. The further study verified that the VEGF-120 kDa was a specific isoform of VEGF-C. In addition, by binding to VEGF receptor 2 (VEGFR2) and then inhibiting the Hippo pathway, the VEGF-120 kDa derived from GBM cells strongly stimulated the expression of tafazzin (TAZ) in endothelial cells, which eventually promoted angiogenesis [101]. Another study showed that hypoxia increased connexin 43 (Cx43) levels in sEVs secreted by glioma cells and acted on vascular endothelial cells to promote glioma angiogenesis [102].
sEVs and glioma metastasis
In many cancer-related deaths, metastasis is an important issue [103]. As one of the most dangerous cancers, GBM rarely occurs in distant metastasis but mainly spreads through local invasion of the brain. However, GBM can spread to the CNS, which makes it hard to completely remove the tumor by surgery [104]. Emerging evidence suggests that advanced cancer cells can secrete sEVs that facilitate tumor progression [105, 106]. Recently, a study revealed that miR-5096 was able to stimulate the formation of filopodia filamentous pseudopodia and promote glioma cell invasion via regulating the K+ channel Kir 4.1. Further studies showed that miR-5096 could also promote the secretion of sEVs leading to GBM metastasis [107]. Cell adhesion molecule 1 (CADM1), a neural tissue-specific protein, plays an important role in cell–cell adhesion and is able to suppress the activation of STAT3 signaling, which is usually activated in GBM [108,109,110]. In addition, suppressing the phosphorylation of STAT3 can obviously decrease metastasis [111], and it has been reported that miR-148a could facilitate GBM progression via increasing CADM1/STAT3 signaling [112, 113]. Cai and colleagues found that miR-148a loaded in sEVs could promote GBM progression and metastasis by activating STAT3 signaling via CADM1, and the level of which in body fluids of GBM patients was higher than that of healthy individuals. These suggest that sEVs-miR-148a may serve as an effective diagnostic biomarker for GBM [114]. As a member of the adhesion molecules family, epithelial cell adhesion molecule (EpCAM) is a single transmembrane protein encoded by the tumor-associated calcium signal transduction gene 1 (TACSTD1) [115]. The research of Gu and colleagues showed that sEVs-EpCAM promoted glioma metastasis via targeting CD44 signaling molecules which are on the surface of glioma cells [116].
sEVs in the TME of glioma
In gliomas, TME consists of non-tumor cells, including microglia, resident astrocytes, endothelial cells, extracellular matrix components, tumor-associated macrophages (TAMs), proteins, and secreted molecules. All these components play important roles in intercellular communication with tumor cells, thereby regulating disease progression [117]. GBM establishes a highly immunosuppressive microenvironment, promotes tumor progression, and has a typical characteristic of inflammatory response with an accumulation of macrophages via communicating with normal brain cells [118, 119]. GBM can recruit immune cells from the bloodstream, with monocytes comprising the predominant subset. Monocytes are highly malleable and classified into M1 macrophages with the function of pro-inflammatory and M2 macrophages with the function of anti-inflammatory [120, 121]. It has been reported that microglia/macrophages account for up to one-third of the tumor mass. Moreover, glioma-associated microglia and macrophages have been identified as key players in the resistance to immunoregulatory therapy [122, 123]. Therefore, the targeting of these immune cells and related molecules offers a novel and promising therapeutic strategy for glioma patients.
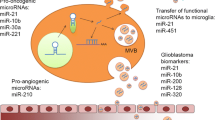
Gabrusiewicz et al. demonstrated that macrophages convert to an immunosuppressed M2 phenotype after uptake by GSCs-derived sEVs, which may be able to act as an effective regulator of the immunosuppressive tumor microenvironment [124]. Similarly, Juliana Azambuja et al. also found the transformation of the macrophage phenotype into type M2 after being exposed to sEVs derived from GBM cell lines in culture, and further experiments in mice yielded the same results [118]. Van der Vos et al. investigated the relationship between sEVs-miRNA secreted by glioma cells and microglia. They found that miR-451/miR-21 in sEVs derived from glioma cells were transported to microglia, resulting in increased proliferation of microglia and the transfer of cytokines profile to immunosuppression [125]. Glioma cells have been shown to secrete sEVs loaded with miR-3591-3p and target TAMs to promote the establishment of an immunosuppressive microenvironment [126]. In addition, research by Li et al. found that sEVs-miR1246 derived from hypoxic glioma could regulate NF-κB and STAT3 pathways by targeting TERF2IP to induce polarization of M2 macrophages and promote the proliferation and metastasis of gliomas [127]. Similarly, glioma-derived sEVs-miR-3184 were able to polarize macrophages to an M2-like phenotype and exacerbate tumor progression [128]. In the latest study, Guo et al. found that hypoxia drove GSCs to produce higher levels of glutamate, which activated local neurons. Neuronal activity promoted GBM progression by facilitating microglial M2 polarization through enriching miR-200c-3p in neuron-derived sEVs [129].
Myeloid-derived suppressor cells (MDSCs) were reported to play a pivotal role in regulating the formation of immunosuppressive environments that enable gliomas to evade host immune responses. However, the exact mechanism is not yet clear. Guo et al. observed that hypoxia-stimulated glioma-derived sEVs-miR-10a and miR-21 mediated differentiation and activation of MDSC, which made a stronger ability to induce MDSCs than normoxia-stimulated glioma-derived sEVs [130]. Another research by the same authors showed that glioma-derived sEVs-miR-29a/miR-92a could also promote the generation of the immunosuppressive microenvironment by stimulating the proliferation and differentiation of functional MDSCs [131]. In addition, sEVs-miR-1298-5p in cerebrospinal fluid (CSF) can promote the immunosuppressive effect of MDSCs, and then promote the development of glioma [132].
sEVs and resistance to treatment
Maximum surgical resection followed by chemotherapy and radiotherapy is the standard treatment for high-grade gliomas. However, GBM, the highest glioma grade, is often resistant to multiple treatments [133, 134]. The resistance of glioma to radiotherapy and chemotherapy is the main factor affecting the treatment effect and leading to poor prognosis. Therefore, it is necessary to figure out the underlying mechanism and explore new treatments to reverse the resistance of GBM to radiation and cytotoxic drugs. In recent years, accumulating shreds of evidence reveals that sEVs, playing a vital role in cell–cell communication, may cause horizontal transmission of resistance capacity between cancer cells [28, 135].
Research by Yue et al. showed that sEVs-miR-301a derived from hypoxic GBM cells could be transported to corresponding normoxia-cultured cells, leading to radiation resistance via directly targeting TCEAL7 genes. This may provide a new target to reverse the resistance of glioma cells to radiotherapy [136]. In Guo et al. 's study, neuronal activation led to increased miR-184-3p content in sEVs, which were transmitted to GSCs and decreased N6-methyladenosine (m6A) level in GSCs by inhibiting the expression of RBM15. RBM15 deficiency decreased m6A modification of DLG3 mRNA and subsequently induced GSC proneural-to-mesenchymal transition by activating the STAT3 pathway to support glioblastoma progression and radioresistance [137]. Zhang et al. reported an upregulation of Circ_0012381 expression in radiation-treated GBM cells, and Circ_0012381 entered microglia through sEVs and induced M2 type microglia to increase ARG 1 expression, and further promoted the growth of GBM cells after radiation treatment. Therefore, inhibition of sEVs secretion may represent a promising approach to improve the therapeutic outcome of radiotherapy in GBM patients [138]. Temozolomide (TMZ), a monofunctional DNA-alkylating agent, acts as the first-line chemotherapy drug for the treatment of glioma [139]. It was reported that sEVs-miR-106a-5p derived from hypoxic glioma cells decreased the sensitivity of glioma cells to TMZ chemotherapy through the downregulation of PTEN [140]. Yang and colleagues revealed that sEVs-miR-221 induced TMZ resistance in glioma via targeting DNM3 [78]. TMZ resistance greatly reduces the effectiveness of treatment. Moreover, related studies have shown that sEVs secreted by TMZ-resistant glioma cells can spread TMZ chemoresistance to TMZ-sensitive glioma cells. Yin et al. revealed that TMZ-resistant glioma cells secreted sEVs containing bioactive miR-1238, which could be absorbed by TMZ-sensitive cells, then acquire TMZ resistance. Thus, sEVs-miR-1238 may be an effective biomarker for assessing the effect of chemotherapy [141]. Zeng et al. reported that overexpression of miR-151a made GBM cells sensitive to TMZ by inhibiting the X-ray repair cross-complementing 4 (XRCC4), which can trigger the DNA repair. They further incubated GBM receptor cells with sEVs derived from TMZ-resistant or TMZ-sensitive cells. The results showed that GBM receptor cells co-cultured with sEVs secreted by TMZ-resistant glioma cells were more resistant to TMZ. However, when they restored miR-151a in TMZ-resistant sEVs, the TMZ resistance in GBM receptor cells was significantly reduced [142].
In addition to the miRNAs, the circWDR62 delivery mediated by sEVs can promote TMZ resistance and malignant progression in vitro and in vivo by targeting the glioma miR-37–30-3p/MGMT axis. In addition, sEVs-circWDR62 derived from serum may act as an effective prognostic marker for glioma [143]. The study of Ding et al. showed that sEVs-circ_0072083 level was enhanced in TMZ-resistant patients and indicated a lower overall survival in glioma. Mechanically, sEVs-circ_0072083 promoted TMZ resistance via increasing NANOG via regulating miR-1252-5p mediated degradation and demethylation in glioma [144]. Other studies have shown that TMZ-resistant GBM cell-derived sEVs-circ_0043949 promotes TMZ resistance through upregulation of ITGA1 expression, providing a potential therapeutic target for TMZ-resistant GBM [145]. Zhang et al. revealed that sEVs could deliver lncRNA SBF2-AS1 from TMZ-resistant glioma cells to TMZ-sensitive glioma cells to spread TMZ resistance in glioma. The mechanism is that lncRNA SBF2-AS1 functioned as a ceRNA and sponged miR-151a-3p to regulate the expression of XRCC4, thus accelerating the repairment of TMZ-induced DNA damage [146].
Potential clinical applications of sEVs in glioma
sEVs in glioma diagnosis: promising candidates for liquid biopsy
In the field of neuro-oncology, the diagnosis and monitoring of gliomas are still challenging [147]. The main methods currently include neuroimaging and histological analysis of brain biopsy samples. The imaging method is a non-invasive examination, which does little harm to patients but is not sensitive enough, especially in the early stage of the tumor. Tumor tissue biopsy is able to accurately diagnose and evaluate the development of tumors. However, it cannot be performed in large quantities and repeatedly due to the characteristic of invasion. In addition, biopsy specimens may not completely represent the entire tumor. Thus, it is necessary to develop a non-invasive and accurate method of evaluating tumors to improve the quality of life belonging to GBM patients. Liquid biopsy referring to a new method for assessing the progress of GBM tumors and monitoring the effects of treatment, is attracting more and more attention.
Although liquid biopsy has been proven to be promising, the development of clinically validated biomarkers for tumor detection remains a considerable challenge, especially for gliomas. Currently, the most researched biomarkers mainly include circulating tumor cells (CTCs), circulating tumor DNA (ctDNA), and EVs [148]. The advantage of CTCs is that they can analyze the entire tumor genome, but CTCs are difficult to detect due to their small quantity and can only present a single cell type of the heterogeneous tumor. In addition, although ctDNA seems to contain more mutations belonging to tumors than CTCs, it can still not reflect the heterogeneity of tumors. In comparison, sEVs containing many kinds of functional biomolecules can reveal the complicated heterogeneity of the entire tumor. More importantly, they are very stable and readily available in almost all types of human biological fluids [149]. In addition, due to nano-sized characteristics, sEVs can cross the intact blood–brain barrier (BBB), which has the function of protecting the central nervous system from toxins and infectious pathogens [150,151,152]. These factors make sEVs the most promising biomarker suitable for early diagnosis and genotyping of tumors, regardless of the stage.
As early as 2008, Johan Skog et al. successfully detected the status of EGFRvIII in GBM by using microvesicles extracted from peripheral blood. The researchers summarized that longitudinal blood sampling provided a novel approach to monitoring the genetic dynamics of tumors [153]. Fraser et al. found that the amount of sEVs-protein is correlated with the invasion of glioma, which reveals the potential of this measurement in glioma diagnosis [154]. In 2018, Manda et al. used sEVs as biomarkers to detect high-grade gliomas which were EGFR-positive [155]. In fact, 90% of GBM patients aberrantly express at least one of the four markers at the exosomal level, which are EGFR, EGRRvIII, podoplanin, and IDH1 [156]. So far, in the field of sEVs for tumor diagnosis, protein-loaded sEVs are the most frequently studied [157]. Nevertheless, other components that sEVs contain, such as miRNAs, have also attracted increasing attention.
Research by Yang et al. revealed that the expression level of miR-221 was elevated in high-grade glioma tissues, and sEVs-miR-221 derived from the serum of glioma patients had a higher level than that of the control group by further study. What's more, the level of sEVs-miR-221 in serum increases with the elevation of glioma grade. This research suggested that sEVs-miR-221 derived from serum may potentially serve as a valuable biomarker for glioma diagnosis [78]. More promising data came from Manterola et al. Firstly, they isolated sEVs from the serum of 30 GBM patients and 30 healthy people. Then, they found that two miRNA (miR-564-3p and miR-320) and one small noncoding RNA (RUN6–1) presented the greatest difference in expression via miRNA chip technology, and the further study found that their expression level was significantly correlated with the diagnosis of GBM patients. Moreover, RNU6-1 was consistently an independent predictor of GBM diagnosis [158]. In addition, a study by Lan et al. revealed that the level of sEVs-miR-301a derived from serum was significantly increased in glioma patients than that in healthy controls. Further research showed that heightened levels of serum sEVs-miR-301a were associated with increasing pathological grades of glioma. Notably, the levels of serum sEVs-miR-301a were obviously decreased after the primary tumor was surgically removed but subsequently increased during GBM recurrence. These findings revealed that serum sEVs-miR-301a might act as a valuable Diagnostic biomarker for glioma [159].
Cerebrospinal fluid (CSF) examination has been widely utilized in the clinical monitoring of CNS diseases, but it is rarely used in glioma so far. MiR-21, the expression level of which was up-regulated in glioma and associated with the histological grade of glioma [160, 161]. Shi and colleagues found that the levels of sEVs-miR-21 isolated from CSF of glioma patients were obviously higher compared to healthy subjects. In contrast, there was no difference in the expression of sEVs-miR-21 isolated from serum. Furthermore, the CSF-derived sEVs-miR-21 levels were associated with tumor spinal/ventricle metastasis and recurrence [160]. This suggested sEVs-miR-21 in CSF might be a promising diagnostic and prognostic biomarker for glioma patients.
There are some other reports of sEVs serving as promising diagnostic biomarkers [80, 114, 143, 146, 162,163,164,165]. We summarize in Table 3.
The potential application of sEVs in anti-glioma therapy
In recent years, the function of sEVs in intercellular communication has become widely known. They can reveal much information about the parental cells, leading to in-depth studies on the diagnostic application of sEVs. In fact, sEVs also have great application potential in the therapy of cancer, although the current research is relatively backward. Current biological therapeutics, such as short interfering RNA and recombinant proteins, have many deficiencies, including easy degradation, restricted membrane permeability, and triggering undesirable immune reactions. However, sEVs have the following advantages as glioma treatment. First, sEVs can transport their contents to specific targets through their surface molecules and homing characteristics, which is good specificity. Second, the double-layer phospholipid membrane structure of sEVs protects its contents from the decomposition of protease and RNAase, which is highly stable. Third, self-derived sEVs have a high degree of histocompatibility and do not induce an adverse immune response, which is safe. Fourth, the clearance from the mononuclear phagocyte system is reduced due to their nanoscale size. Fifth, sEVs can cross the BBB [166].
There are two ways to load sEVs with cargo, including exogenous loading and endogenous loading. In endogenous loading, the modifications occur during the formation of the sEVs. In exogenous loading, sEVs are first isolated and then modified by freeze–thaw cycles, incubation, sonication, electroporation, and extrusion [167]. Researchers use sEVs to deliver tumor-suppressing ncRNAs for research. For example, in the rat model of GBM, Hamideh, and colleagues showed that the administration of sEVs loaded with miR-21-sponge construct could significantly reduce the tumor volume [168]. Katakowski et al. found that sEVs derived from MSCs overexpressing miR-146b decreased the growth of glioma xenograft in a rat model of primary brain tumor via intra-tumor injection [169]. Similar research was performed by Fareh and colleagues. They engineered primary glioma cells to stably express the miR-302–367, which has the function of inhibiting tumors and was mainly packaged in sEVs. These sEVs were taken up by neighboring GBM cells leading to antitumor effects both in vivo and in vitro [170]. Moreover, Munoz et al. revealed that anti-miR-9-delivering sEVs improved the expression level of multidrug transporters and the sensitivity to TMZ in drug-resistant GBM cells, resulting in higher cell mortality and caspase activity. This may be an effective way to overcome the resistance of gliomas to chemotherapy [171]. Qian et al. used sEVs derived from neural stem cells to deliver miR-124-3p to glioma and significantly inhibited the malignant biological behavior of glioma cells [172]. Another approach is to package the drug itself into sEVs. It has been reported that paclitaxel-loaded sEVs resulted in a nearly 50-fold increase in cytotoxicity in multidrug-resistant tumors compared to paclitaxel without exosomes [173]. In addition, the researchers developed a nano delivery system based on functionalized macrophage sEVs targeted with heliostat and PPM1D-siRNA for PPM1D mutant diffuse intrinsic pontine glioma, with higher administration efficiency and better therapeutic efficacy than free drugs [174]. Other reports on the use of dual receptor-specific sEVs as carriers loaded with TMZ and O6-benzyguanine for the eradication of TMZ-resistant GBM showed that the sEVs had good proliferation inhibition in vitro and prolonged the median survival of U87MG tumor-bearing mice without causing adverse effects [175]. The latest approach is achieved by using a microfluidic electroporation approach in which a combination of nano- and milli-second pulses produces large amounts of IFN-γ mRNA-loaded sEVs with CD64 overexpressed on their surface. The CD64 molecule serves as an adaptor to dock targeting ligands, such as anti-CD71 and anti-programmed cell death-ligand 1 (PD-L1) antibodies. The resulting immunogenic sEVs preferentially target glioma cells and generate potent antitumour activities in vivo, including against tumours intrinsically resistant to immunotherapy [176]. These studies demonstrate the potential value of sEVs in the treatment of gliomas.
Conclusions and prospects
Gliomas present poor prognosis due to invasion and resistance to multiple treatments. Up to now, the complex pathogenesis of glioma is still not fully understood. sEVs can transport a variety of biomolecules and mediate communication between tumor cells and TME via the horizontal transfer of information. In gliomas, extensive studies have shown that sEVs are selectively packaged, secreted, and transferred between cells, thus regulating multiple biological characteristics, including proliferation, invasion, angiogenesis, immune escape, and treatment resistance. Moreover, the ability to easily cross various biological barriers (such as BBB) provides a broad prospect for clinical applications of sEVs in the diagnosis and treatment of gliomas.
Nevertheless, there are still many challenges in the view of clinical applications. Firstly, the separation and purification of sEVs, which is the premise for large-scale use of sEVs, have not been standardized. Secondly, more reliable biomarkers should be identified. In the future, a diagnostic panel will combine multiple biomarkers rather than single biomarkers to provide information on early diagnosis and prognosis. Then, how to maximize the use, standardization, and quantification of sEVs still needs further research due to its limited genetic information. Although the therapeutic applications of sEVs have made great progress in vitro and animal experiments, before applying sEVs to the clinical treatment of glioma, sufficient safety, targeting efficacy, and avoidance of adverse reactions should be considered ensured. In addition, recent studies have also found that neuronal sEVs play a key role in the malignant biological behaviors of glioma such as radiotherapy resistance and immunosuppression, which is worthy of further in-depth and extensive study. Finally, more structured studies in different laboratories around the world and achieving a consensus on sEVs terminology are needed to advance the field of sEVs.
We believe that with the improvement of technology, sEVs will be widely used in the clinical diagnosis, treatment, and prognosis of glioma in the near future.
Data availability
No datasets were generated or analysed during the current study.
References
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Molinaro AM, Taylor JW, Wiencke JK, Wrensch MR. Genetic and molecular epidemiology of adult diffuse glioma. Nat Rev Neurol. 2019;15(7):405–17.
Hanif F, Muzaffar K, Perveen K, Malhi SM, Simjee ShU. Glioblastoma multiforme: a review of its epidemiology and pathogenesis through clinical presentation and treatment. Asian Pac J Cancer Prev. 2017;18(1):3–9.
Witthayanuwat S, Pesee M, Supaadirek C, Supakalin N, Thamronganantasakul K, Krusun S. Survival analysis of glioblastoma multiforme. Asian Pac J Cancer Prev. 2018;19(9):2613–7.
Jain KK. A critical overview of targeted therapies for glioblastoma. Front Oncol. 2018;8:419.
Touat M, Idbaih A, Sanson M, Ligon KL. Glioblastoma targeted therapy: updated approaches from recent biological insights. Ann Oncol. 2017;28(7):1457–72.
Bradshaw A, Wickremsekera A, Tan ST, Peng L, Davis PF, Itinteang T. Cancer stem cell hierarchy in glioblastoma multiforme. Front Surg. 2016;3:21.
Zhang L, Yu D. Exosomes in cancer development, metastasis, and immunity. Biochimica et Biophysica Acta (BBA). 2019;1871(2):455–68.
Thery C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F, Atkin-Smith GK, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750.
Jabbari N, Akbariazar E, Feqhhi M, Rahbarghazi R, Rezaie J. Breast cancer-derived exosomes: tumor progression and therapeutic agents. J Cell Physiol. 2020;235(10):6345–56.
Fu F, Jiang W, Zhou L, Chen Z. Circulating Exosomal miR-17-5p and miR-92a-3p Predict Pathologic Stage and Grade of Colorectal Cancer. Transl Oncol. 2018;11(2):221–32.
Elsharkawi F, Elsabah M, Shabayek M, Khaled H. Urine and serum exosomes as novel biomarkers in detection of bladder cancer. Asian Pac J Cancer Prev. 2019;20(7):2219–24.
Pan BT, Johnstone RM. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor. Cell. 1983;33(3):967–78.
Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019;8:7.
Borges FT, Reis LA, Schor N. Extracellular vesicles: structure, function, and potential clinical uses in renal diseases. Braz J Med Biol Res. 2013;46(10):824–30.
Gould SJ, Booth AM, Hildreth JEK. The Trojan exosome hypothesis. Proc Natl Acad Sci USA. 2003;100(19):10592–7.
Fang Y, Wu N, Gan X, Yan W, Morrell JC, Gould SJ. Higher-order oligomerization targets plasma membrane proteins and HIV gag to exosomes. PLoS Biol. 2007;5(6): e158.
Tian X, Shen H, Li Z, Wang T, Wang S. Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor microenvironment. J Hematol Oncol. 2019;12(1):84.
Wei D, Zhan W, Gao Y, Huang L, Gong R, Wang W, Zhang R, Wu Y, Gao S, Kang T. RAB31 marks and controls an ESCRT-independent exosome pathway. Cell Res. 2021;31(2):157–77.
Cheng J, Meng J, Zhu L, Peng Y. Exosomal noncoding RNAs in Glioma: biological functions and potential clinical applications. Mol Cancer. 2020;19(1):66.
Frydrychowicz M, Kolecka-Bednarczyk A, Madejczyk M, Yasar S, Dworacki G. Exosomes - structure, biogenesis and biological role in non-small-cell lung cancer. Scand J Immunol. 2015;81(1):2–10.
Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88:487–514.
Cocucci E, Meldolesi J. Ectosomes and exosomes: shedding the confusion between extracellular vesicles. Trends Cell Biol. 2015;25(6):364–72.
Atkin-Smith GK, Tixeira R, Paone S, Mathivanan S, Collins C, Liem M, Goodall KJ, Ravichandran KS, Hulett MD, Poon IK. A novel mechanism of generating extracellular vesicles during apoptosis via a beads-on-a-string membrane structure. Nat Commun. 2015;6:7439.
Li Y, Zhao J, Yu S, Wang Z, He X, Su Y, Guo T, Sheng H, Chen J, Zheng Q, et al. Extracellular vesicles long RNA sequencing reveals abundant mRNA, circRNA, and lncRNA in human blood as potential biomarkers for cancer diagnosis. Clin Chem. 2019;65(6):798–808.
Makarova J, Turchinovich A, Shkurnikov M, Tonevitsky A. Extracellular miRNAs and cell-cell communication: problems and prospects. Trends Biochem Sci. 2021;46(8):640–51.
Wang S, Zhang K, Tan S, Xin J, Yuan Q, Xu H, Xu X, Liang Q, Christiani DC, Wang M, et al. Circular RNAs in body fluids as cancer biomarkers: the new frontier of liquid biopsies. Mol Cancer. 2021;20(1):13.
Sun Z, Yang S, Zhou Q, Wang G, Song J, Li Z, Zhang Z, Xu J, Xia K, Chang Y, et al. Emerging role of exosome-derived long non-coding RNAs in tumor microenvironment. Mol Cancer. 2018;17(1):82.
Campos-Carrillo A, Weitzel JN, Sahoo P, Rockne R, Mokhnatkin JV, Murtaza M, Gray SW, Goetz L, Goel A, Schork N, et al. Circulating tumor DNA as an early cancer detection tool. Pharmacol Ther. 2020;207: 107458.
Wu H, Fu M, Liu J, Chong W, Fang Z, Du F, Liu Y, Shang L, Li L. The role and application of small extracellular vesicles in gastric cancer. Mol Cancer. 2021;20(1):71.
Trinidad CV, Tetlow AL, Bantis LE, Godwin AK. Reducing ovarian cancer mortality through early detection: approaches using circulating biomarkers. Cancer Prev Res (Phila). 2020;13(3):241–52.
Mathieu M, Martin-Jaular L, Lavieu G, Thery C. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17.
Hood JL. Natural melanoma-derived extracellular vesicles. Semin Cancer Biol. 2019;59:251–65.
Bordas M, Genard G, Ohl S, Nessling M, Richter K, Roider T, Dietrich S, Maass KK, Seiffert M. Optimized protocol for isolation of small extracellular vesicles from human and murine lymphoid tissues. Int J Mol Sci. 2020;21:15.
Van Deun J, Mestdagh P, Sormunen R, Cocquyt V, Vermaelen K, Vandesompele J, Bracke M, De Wever O, Hendrix A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J Extracell Vesicles. 2014;3:1.
Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation, density-gradient separation, and immunoaffinity capture methods. Methods Mol Biol. 2015;1295:179–209.
Carnino JM, Lee H, Jin Y. Isolation and characterization of extracellular vesicles from Broncho-alveolar lavage fluid: a review and comparison of different methods. Respir Res. 2019;20(1):240.
Abhange K, Makler A, Wen Y, Ramnauth N, Mao W, Asghar W, Wan Y. Small extracellular vesicles in cancer. Bioact Mater. 2021;6(11):3705–43.
Wu M, Ozcelik A, Rufo J, Wang Z, Fang R, Jun Huang T. Acoustofluidic separation of cells and particles. Microsyst Nanoeng. 2019;5:32.
Iwai K, Minamisawa T, Suga K, Yajima Y, Shiba K. Isolation of human salivary extracellular vesicles by iodixanol density gradient ultracentrifugation and their characterizations. J Extracell Vesicles. 2016;5:30829.
Monguió-Tortajada M, Gálvez-Montón C, Bayes-Genis A, Roura S, Borràs FE. Extracellular vesicle isolation methods: rising impact of size-exclusion chromatography. Cell Mol Life Sci. 2019;76(12):2369–82.
Guo SC, Tao SC, Dawn H. Microfluidics-based on-a-chip systems for isolating and analysing extracellular vesicles. J Extracell Vesicles. 2018;7(1):1508271.
Szatanek R, Baran J, Siedlar M, Baj-Krzyworzeka M. Isolation of extracellular vesicles: Determining the correct approach (Review). Int J Mol Med. 2015;36(1):11–7.
Niu Z, Pang RTK, Liu W, Li Q, Cheng R, Yeung WSB. Polymer-based precipitation preserves biological activities of extracellular vesicles from an endometrial cell line. PLoS ONE. 2017;12(10): e0186534.
Abramowicz A, Widlak P, Pietrowska M. Proteomic analysis of exosomal cargo: the challenge of high purity vesicle isolation. Mol Biosyst. 2016;12(5):1407–19.
Liang LG, Kong MQ, Zhou S, Sheng YF, Wang P, Yu T, Inci F, Kuo WP, Li LJ, Demirci U, et al. An integrated double-filtration microfluidic device for isolation, enrichment and quantification of urinary extracellular vesicles for detection of bladder cancer. Sci Rep. 2017;7:46224.
Woo HK, Sunkara V, Park J, Kim TH, Han JR, Kim CJ, Choi HI, Kim YK, Cho YK. Exodisc for rapid, size-selective, and efficient isolation and analysis of nanoscale extracellular vesicles from biological samples. ACS Nano. 2017;11(2):1360–70.
Sunkara V, Kim CJ, Park J, Woo HK, Kim D, Ha HK, Kim MH, Son Y, Kim JR, Cho YK. Fully automated, label-free isolation of extracellular vesicles from whole blood for cancer diagnosis and monitoring. Theranostics. 2019;9(7):1851–63.
Brett SI, Lucien F, Guo C, Williams KC, Kim Y, Durfee PN, Brinker CJ, Chin JI, Yang J, Leong HS. Immunoaffinity based methods are superior to kits for purification of prostate derived extracellular vesicles from plasma samples. Prostate. 2017;77(13):1335–43.
Sharma P, Ludwig S, Muller L, Hong CS, Kirkwood JM, Ferrone S, Whiteside TL. Immunoaffinity-based isolation of melanoma cell-derived exosomes from plasma of patients with melanoma. J Extracell Vesicles. 2018;7(1):1435138.
Lo TW, Zhu Z, Purcell E, Watza D, Wang J, Kang YT, Jolly S, Nagrath D, Nagrath S. Microfluidic device for high-throughput affinity-based isolation of extracellular vesicles. Lab Chip. 2020;20(10):1762–70.
Abd Rahman N, Ibrahim F, Yafouz B. Dielectrophoresis for biomedical sciences applications: a review. Sensors (Basel). 2017;17:3.
Ibsen SD, Wright J, Lewis JM, Kim S, Ko SY, Ong J, Manouchehri S, Vyas A, Akers J, Chen CC, et al. Rapid isolation and detection of exosomes and associated biomarkers from plasma. ACS Nano. 2017;11(7):6641–51.
Lewis J, Alattar AA, Akers J, Carter BS, Heller M, Chen CC. A pilot proof-of-principle analysis demonstrating dielectrophoresis (DEP) as a glioblastoma biomarker platform. Sci Rep. 2019;9(1):10279.
Shu S, Yang Y, Allen CL, Hurley E, Tung KH, Minderman H, Wu Y, Ernstoff MS. Purity and yield of melanoma exosomes are dependent on isolation method. J Extracell Vesicles. 2020;9(1):1692401.
Oeyen E, Van Mol K, Baggerman G, Willems H, Boonen K, Rolfo C, Pauwels P, Jacobs A, Schildermans K, Cho WC, et al. Ultrafiltration and size exclusion chromatography combined with asymmetrical-flow field-flow fractionation for the isolation and characterisation of extracellular vesicles from urine. J Extracell Vesicles. 2018;7(1):1490143.
Wan Y, Cheng G, Liu X, Hao SJ, Nisic M, Zhu CD, Xia YQ, Li WQ, Wang ZG, Zhang WL, et al. Rapid magnetic isolation of extracellular vesicles via lipid-based nanoprobes. Nat Biomed Eng. 2017;1:1.
Ludwig AK, De Miroschedji K, Doeppner TR, Borger V, Ruesing J, Rebmann V, Durst S, Jansen S, Bremer M, Behrmann E, et al. Precipitation with polyethylene glycol followed by washing and pelleting by ultracentrifugation enriches extracellular vesicles from tissue culture supernatants in small and large scales. J Extracell Vesicles. 2018;7(1):1528109.
Mao W, Wen Y, Lei H, Lu R, Wang S, Wang Y, Chen R, Gu Y, Zhu L, Abhange KK, et al. Isolation and retrieval of extracellular vesicles for liquid biopsy of malignant ground-glass opacity. Anal Chem. 2019;91(21):13729–36.
Boriachek K, Masud MK, Palma C, Phan HP, Yamauchi Y, Hossain MSA, Nguyen NT, Salomon C, Shiddiky MJA. Avoiding pre-isolation step in exosome analysis: direct isolation and sensitive detection of exosomes using gold-loaded nanoporous ferric oxide nanozymes. Anal Chem. 2019;91(6):3827–34.
Liu C, Xu X, Li B, Situ B, Pan W, Hu Y, An T, Yao S, Zheng L. Single-exosome-counting immunoassays for cancer diagnostics. Nano Lett. 2018;18(7):4226–32.
Lopez-Cobo S, Campos-Silva C, Moyano A, Oliveira-Rodriguez M, Paschen A, Yanez-Mo M, Blanco-Lopez MC, Vales-Gomez M. Immunoassays for scarce tumour-antigens in exosomes: detection of the human NKG2D-Ligand, MICA, in tetraspanin-containing nanovesicles from melanoma. J Nanobiotechnology. 2018;16(1):47.
Oliveira-Rodriguez M, Serrano-Pertierra E, Garcia AC, Lopez-Martin S, Yanez-Mo M, Cernuda-Morollon E, Blanco-Lopez MC. Point-of-care detection of extracellular vesicles: Sensitivity optimization and multiple-target detection. Biosens Bioelectron. 2017;87:38–45.
Liang K, Liu F, Fan J, Sun D, Liu C, Lyon CJ, Bernard DW, Li Y, Yokoi K, Katz MH, et al. Nanoplasmonic Quantification of Tumor-derived Extracellular Vesicles in Plasma Microsamples for Diagnosis and Treatment Monitoring. Nat Biomed Eng. 2017;1:1.
Liu C, Zeng X, An Z, Yang Y, Eisenbaum M, Gu X, Jornet JM, Dy GK, Reid ME, Gan Q, et al. Sensitive detection of exosomal proteins via a compact surface plasmon resonance biosensor for cancer diagnosis. ACS Sens. 2018;3(8):1471–9.
Boriachek K, Islam MN, Gopalan V, Lam AK, Nguyen NT, Shiddiky MJA. Quantum dot-based sensitive detection of disease specific exosome in serum. Analyst. 2017;142(12):2211–9.
Bai Y, Lu Y, Wang K, Cheng Z, Qu Y, Qiu S, Zhou L, Wu Z, Liu H, Zhao J, et al. Rapid isolation and multiplexed detection of exosome tumor markers via queued beads combined with quantum dots in a microarray. Nanomicro Lett. 2019;11(1):59.
Bellassai N, D’Agata R, Jungbluth V, Spoto G. Surface plasmon resonance for biomarker detection: advances in non-invasive cancer diagnosis. Front Chem. 2019;7:570.
Maroto R, Zhao Y, Jamaluddin M, Popov VL, Wang H, Kalubowilage M, Zhang Y, Luisi J, Sun H, Culbertson CT, et al. Effects of storage temperature on airway exosome integrity for diagnostic and functional analyses. J Extracell Vesicles. 2017;6(1):1359478.
Xie Q, Mittal S, Berens ME. Targeting adaptive glioblastoma: an overview of proliferation and invasion. Neuro Oncol. 2014;16(12):1575–84.
Bian EB, Chen EF, Xu YD, Yang ZH, Tang F, Ma CC, Wang HL, Zhao B. Exosomal lncRNA-ATB activates astrocytes that promote glioma cell invasion. Int J Oncol. 2019;54(2):713–21.
Cayre A, Rossignol F, Clottes E, Penault-Llorca F. aHIF but not HIF-1alpha transcript is a poor prognostic marker in human breast cancer. Breast Cancer Res. 2003;5(6):R223-230.
Thrash-Bingham CA, Tartof KD. aHIF: a natural antisense transcript overexpressed in human renal cancer and during hypoxia. J Natl Cancer Inst. 1999;91(2):143–51.
Dai X, Liao K, Zhuang Z, Chen B, Zhou Z, Zhou S, Lin G, Zhang F, Lin Y, Miao Y, et al. AHIF promotes glioblastoma progression and radioresistance via exosomes. Int J Oncol. 2019;54(1):261–70.
Chai Y, Wu HT, Liang CD, You CY, Xie MX, Xiao SW. Exosomal lncRNA ROR1-AS1 derived from tumor cells promotes glioma progression via regulating miR-4686. Int J Nanomedicine. 2020;15:8863–72.
Ma W, Zhou Y, Liu M, Qin Q, Cui Y. Long non-coding RNA LINC00470 in serum derived exosome: a critical regulator for proliferation and autophagy in glioma cells. Cancer Cell Int. 2021;21(1):149.
Xu X, Liu Y, Li Y, Chen H, Zhang Y, Liu J, Deng S, Zheng Y, Sun X, Wang J, et al. Selective exosome exclusion of miR-375 by glioma cells promotes glioma progression by activating the CTGF-EGFR pathway. J Exp Clin Cancer Res. 2021;40(1):16.
Yang JK, Yang JP, Tong J, Jing SY, Fan B, Wang F, Sun GZ, Jiao BH. Exosomal miR-221 targets DNM3 to induce tumor progression and temozolomide resistance in glioma. J Neurooncol. 2017;131(2):255–65.
Lokumcu T, Iskar M, Schneider M, Helm D, Klinke G, Schlicker L, Bethke F, Muller G, Richter K, Poschet G, et al. Proteomic, metabolomic, and fatty acid profiling of small extracellular vesicles from glioblastoma stem-like cells and their role in tumor heterogeneity. ACS Nano. 2024;18(3):2500–19.
Bao Z, Zhang N, Niu W, Mu M, Zhang X, Hu S, Niu C. Exosomal miR-155-5p derived from glioma stem-like cells promotes mesenchymal transition via targeting ACOT12. Cell Death Dis. 2022;13(8):725.
Li H, Zhu J, Liu X, Liu L, Huang S, Wu A, Xu Z, Zhang X, Li Z, Ni F, et al. Glioma stem cell-derived exosomes induce the transformation of astrocytes via the miR-3065-5p/DLG2 signaling axis. Glia. 2024;72(5):857–71.
Shi L, Cao Y, Yuan W, Guo J, Sun G. Exosomal circRNA BTG2 derived from RBP-J overexpressed-macrophages inhibits glioma progression via miR-25-3p/PTEN. Cell Death Dis. 2022;13(5):506.
Bao S, Wu Q, Li Z, Sathornsumetee S, Wang H, McLendon RE, Hjelmeland AB, Rich JN. Targeting cancer stem cells through L1CAM suppresses glioma growth. Cancer Res. 2008;68(15):6043–8.
Raveh S, Gavert N, Ben-Ze’ev A. L1 cell adhesion molecule (L1CAM) in invasive tumors. Cancer Lett. 2009;282(2):137–45.
Pace KR, Dutt R, Galileo DS. Exosomal L1CAM Stimulates Glioblastoma Cell Motility, Proliferation, and Invasiveness. Int J Mol Sci. 2019;20:16.
Hida K, Maishi N, Torii C, Hida Y. Tumor angiogenesis–characteristics of tumor endothelial cells. Int J Clin Oncol. 2016;21(2):206–12.
Rahman R, Smith S, Rahman C, Grundy R. Antiangiogenic therapy and mechanisms of tumor resistance in malignant glioma. J Oncol. 2010;2010: 251231.
Onishi M, Ichikawa T, Kurozumi K, Date I. Angiogenesis and invasion in glioma. Brain Tumor Pathol. 2011;28(1):13–24.
Lang H-L, Hu G-W, Zhang B, Kuang W, Chen Y, Wu L, Xu G-H. Glioma cells enhance angiogenesis and inhibit endothelial cell apoptosis through the release of exosomes that contain long non-coding RNA CCAT2. Oncol Rep. 2017;38(2):785–98.
Lang HL, Hu GW, Chen Y, Liu Y, Tu W, Lu YM, Wu L, Xu GH. Glioma cells promote angiogenesis through the release of exosomes containing long non-coding RNA POU3F3. Eur Rev Med Pharmacol Sci. 2017;21(5):959–72.
Chen X, Yang F, Zhang T, Wang W, Xi W, Li Y, Zhang D, Huo Y, Zhang J, Yang A, et al. MiR-9 promotes tumorigenesis and angiogenesis and is activated by MYC and OCT4 in human glioma. J Exp Clin Cancer Res. 2019;38(1):99.
Wu K, Liu F, Wu W, Chen Y, Wu H, Zhang W. Long non-coding RNA HOX transcript antisense RNA (HOTAIR) suppresses the angiogenesis of human placentation by inhibiting vascular endothelial growth factor A expression. Reprod Fertil Dev. 2019;31(2):377–85.
Jiang Y, Zhao J, Xu J, Zhang H, Zhou J, Li H, Zhang G, Xu K, Jing Z. Glioblastoma-associated microglia-derived exosomal circKIF18A promotes angiogenesis by targeting FOXC2. Oncogene. 2022;41(26):3461–73.
Li Y, Chen J, Chen Z, Xu X, Weng J, Zhang Y, Mo Y, Liu Y, Wang J, Ke Y. CircGLIS3 Promotes High-Grade Glioma Invasion via Modulating Ezrin Phosphorylation. Front Cell Dev Biol. 2021;9: 663207.
Ahn SE, Jeong W, Kim JH, Lim W, Kim J, Bazer FW, Han JY, Song G. ERBB receptor feedback inhibitor 1: identification and regulation by estrogen in chickens. Gen Comp Endocrinol. 2012;175(1):194–205.
Wang M, Zhao Y, Yu ZY, Zhang RD, Li SA, Zhang P, Shan TK, Liu XY, Wang ZM, Zhao PC, et al. Glioma exosomal microRNA-148a-3p promotes tumor angiogenesis through activating the EGFR/MAPK signaling pathway via inhibiting ERRFI1. Cancer Cell Int. 2020;20:518.
Sun X, Ma X, Wang J, Zhao Y, Wang Y, Bihl JC, Chen Y, Jiang C. Glioma stem cells-derived exosomes promote the angiogenic ability of endothelial cells through miR-21/VEGF signal. Oncotarget. 2017;8(22):36137–48.
Wang ZF, Liao F, Wu H, Dai J. Glioma stem cells-derived exosomal miR-26a promotes angiogenesis of microvessel endothelial cells in glioma. J Exp Clin Cancer Res. 2019;38(1):201.
Jiang J, Lu J, Wang X, Sun B, Liu X, Ding Y, Gao G. Glioma stem cell-derived exosomal miR-944 reduces glioma growth and angiogenesis by inhibiting AKT/ERK signaling. Aging (Albany NY). 2021;13(15):19243–59.
Zhang Z, Guo X, Guo X, Yu R, Qian M, Wang S, Gao X, Qiu W, Guo Q, Xu J, et al. MicroRNA-29a-3p delivery via exosomes derived from engineered human mesenchymal stem cells exerts tumour suppressive effects by inhibiting migration and vasculogenic mimicry in glioma. Aging (Albany NY). 2021;13(4):5055–68.
Wang Z, Yuan Y, Ji X, Xiao X, Li Z, Yi X, Zhu Y, Guo T, Wang Y, Chen L, et al. The Hippo-TAZ axis mediates vascular endothelial growth factor C in glioblastoma-derived exosomes to promote angiogenesis. Cancer Lett. 2021;513:1–13.
Yang ZJ, Bi QC, Gan LJ, Zhang LL, Wei MJ, Hong T, Liu R, Qiu CL, Han XJ, Jiang LP. Exosomes Derived from Glioma Cells under Hypoxia Promote Angiogenesis through Up-regulated Exosomal Connexin 43. Int J Med Sci. 2022;19(7):1205–15.
Eccles SA, Welch DR. Metastasis: recent discoveries and novel treatment strategies. Lancet. 2007;369(9574):1742–57.
Lee SJ, Kang WY, Yoon Y, Jin JY, Song HJ, Her JH, Kang SM, Hwang YK, Kang KJ, Joo KM, et al. Natural killer (NK) cells inhibit systemic metastasis of glioblastoma cells and have therapeutic effects against glioblastomas in the brain. BMC Cancer. 2015;15:1011.
Xue M, Chen W, Xiang A, Wang R, Chen H, Pan J, Pang H, An H, Wang X, Hou H, et al. Hypoxic exosomes facilitate bladder tumor growth and development through transferring long non-coding RNA-UCA1. Mol Cancer. 2017;16(1):143.
Jung T, Castellana D, Klingbeil P, Cuesta Hernandez I, Vitacolonna M, Orlicky DJ, Roffler SR, Brodt P, Zoller M. CD44v6 dependence of premetastatic niche preparation by exosomes. Neoplasia. 2009;11(10):1093–105.
Thuringer D, Chanteloup G, Boucher J, Pernet N, Boudesco C, Jego G, Chatelier A, Bois P, Gobbo J, Cronier L, et al. Modulation of the inwardly rectifying potassium channel Kir41 by the pro-invasive miR-5096 in glioblastoma cells. Oncotarget. 2017;8(23):37681–93.
Nakahata S, Morishita K. CADM1/TSLC1 is a novel cell surface marker for adult T-cell leukemia/lymphoma. J Clin Exp Hematop. 2012;52(1):17–22.
Chen F, Xu Y, Luo Y, Zheng D, Song Y, Yu K, Li H, Zhang L, Zhong W, Ji Y. Down-regulation of Stat3 decreases invasion activity and induces apoptosis of human glioma cells. J Mol Neurosci. 2010;40(3):353–9.
Vallath S, Sage EK, Kolluri KK, Lourenco SN, Teixeira VS, Chimalapati S, George PJ, Janes SM, Giangreco A. CADM1 inhibits squamous cell carcinoma progression by reducing STAT3 activity. Sci Rep. 2016;6:24006.
Kim JE, Patel M, Ruzevick J, Jackson CM, Lim M. STAT3 activation in glioblastoma: biochemical and therapeutic implications. Cancers (Basel). 2014;6(1):376–95.
Attarha S, Roy A, Westermark B, Tchougounova E. Mast cells modulate proliferation, migration and stemness of glioma cells through downregulation of GSK3beta expression and inhibition of STAT3 activation. Cell Signal. 2017;37:81–92.
Zhu Y, Zhang X, Wang L, Ji Z, Xie M, Zhou X, Liu Z, Shi H, Yu R. Loss of SH3GL2 promotes the migration and invasion behaviours of glioblastoma cells through activating the STAT3/MMP2 signalling. J Cell Mol Med. 2017;21(11):2685–94.
Cai Q, Zhu A, Gong L. Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1. Bull Cancer. 2018;105(7–8):643–51.
Kurtz J-E, Dufour P. Adecatumumab: an anti-EpCAM monoclonal antibody, from the bench to the bedside. Expert Opin Biol Ther. 2010;10(6):951–8.
Gu Q, Chen X, Zhou L, Liu X. Exosome EpCAM promotes the metastasis of glioma by targeting the CD44 signaling molecule on the surface of glioma cells. Adv Clin Exp Med. 2020;29(11):1277–82.
Lathia JD, Heddleston JM, Venere M, Rich JN. Deadly teamwork: neural cancer stem cells and the tumor microenvironment. Cell Stem Cell. 2011;8(5):482–5.
Azambuja JH, Ludwig N, Yerneni S, Rao A, Braganhol E, Whiteside TL. Molecular profiles and immunomodulatory activities of glioblastoma-derived exosomes. Neurooncol Adv. 2020;2(1):056.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74.
Mantovani A, Biswas SK, Galdiero MR, Sica A, Locati M. Macrophage plasticity and polarization in tissue repair and remodelling. J Pathol. 2013;229(2):176–85.
Gordon S, Plüddemann A, Martinez Estrada F. Macrophage heterogeneity in tissues: phenotypic diversity and functions. Immunol Rev. 2014;262(1):36–55.
Raes G, Van den Bergh R, De Baetselier P, Ghassabeh GH, Scotton C, Locati M, Mantovani A, Sozzani S: Arginase-1 and Ym1 are markers for murine, but not human, alternatively activated myeloid cells. Journal of Immunology (Baltimore, Md : 1950) 2005, 174(11):6561; author reply 6561–6561; author reply 6562.
Locarno CV, Simonelli M, Carenza C, Capucetti A, Stanzani E, Lorenzi E, Persico P, Della Bella S, Passoni L, Mavilio D, et al. Role of myeloid cells in the immunosuppressive microenvironment in gliomas. Immunobiology. 2020;225(1): 151853.
Gabrusiewicz K, Li X, Wei J, Hashimoto Y, Marisetty AL, Ott M, Wang F, Hawke D, Yu J, Healy LM, et al. Glioblastoma stem cell-derived exosomes induce M2 macrophages and PD-L1 expression on human monocytes. Oncoimmunology. 2018;7(4): e1412909.
van der Vos KE, Abels ER, Zhang X, Lai C, Carrizosa E, Oakley D, Prabhakar S, Mardini O, Crommentuijn MH, Skog J, et al. Directly visualized glioblastoma-derived extracellular vesicles transfer RNA to microglia/macrophages in the brain. Neuro Oncol. 2016;18(1):58–69.
Li M, Xu H, Qi Y, Pan Z, Li B, Gao Z, Zhao R, Xue H, Li G. Tumor-derived exosomes deliver the tumor suppressor miR-3591-3p to induce M2 macrophage polarization and promote glioma progression. Oncogene. 2022;41(41):4618–32.
Qian M, Wang S, Guo X, Wang J, Zhang Z, Qiu W, Gao X, Chen Z, Xu J, Zhao R, et al. Hypoxic glioma-derived exosomes deliver microRNA-1246 to induce M2 macrophage polarization by targeting TERF2IP via the STAT3 and NF-kappaB pathways. Oncogene. 2020;39(2):428–42.
Xu H, Li M, Pan Z, Zhang Z, Gao Z, Zhao R, Li B, Qi Y, Qiu W, Guo Q, et al. miR-3184-3p enriched in cerebrospinal fluid exosomes contributes to progression of glioma and promotes M2-like macrophage polarization. Cancer Sci. 2022;113(8):2668–80.
Guo X, Qiu W, Li B, Qi Y, Wang S, Zhao R, Cheng B, Han X, Du H, Pan Z, et al. Hypoxia-induced neuronal activity in glioma patients polarizes microglia by potentiating RNA m6A demethylation. Clin Cancer Res. 2024;30(6):1160–74.
Guo X, Qiu W, Liu Q, Qian M, Wang S, Zhang Z, Gao X, Chen Z, Xue H, Li G. Immunosuppressive effects of hypoxia-induced glioma exosomes through myeloid-derived suppressor cells via the miR-10a/Rora and miR-21/Pten Pathways. Oncogene. 2018;37(31):4239–59.
Guo X, Qiu W, Wang J, Liu Q, Qian M, Wang S, Zhang Z, Gao X, Chen Z, Guo Q, et al. Glioma exosomes mediate the expansion and function of myeloid-derived suppressor cells through microRNA-29a/Hbp1 and microRNA-92a/Prkar1a pathways. Int J Cancer. 2019;144(12):3111–26.
Qi Y, Jin C, Qiu W, Zhao R, Wang S, Li B, Zhang Z, Guo Q, Zhang S, Gao Z, et al. The dual role of glioma exosomal microRNAs: glioma eliminates tumor suppressor miR-1298-5p via exosomes to promote immunosuppressive effects of MDSCs. Cell Death Dis. 2022;13(5):426.
Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJB, Belanger K, Brandes AA, Marosi C, Bogdahn U, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352(10):987–96.
Qazi MA, Vora P, Venugopal C, Sidhu SS, Moffat J, Swanton C, Singh SK. Intratumoral heterogeneity: pathways to treatment resistance and relapse in human glioblastoma. Ann Oncol. 2017;28(7):1448–56.
Xie Y, Dang W, Zhang S, Yue W, Yang L, Zhai X, Yan Q, Lu J. The role of exosomal noncoding RNAs in cancer. Mol Cancer. 2019;18(1):37.
Yue X, Lan F, Xia T. Hypoxic Glioma Cell-Secreted Exosomal miR-301a Activates Wnt/beta-catenin signaling and promotes radiation resistance by targeting TCEAL7. Mol Ther. 2019;27(11):1939–49.
Guo X, Qiu W, Wang C, Qi Y, Li B, Wang S, Zhao R, Cheng B, Han X, Du H, et al. Neuronal activity promotes glioma progression by inducing proneural-to-mesenchymal transition in glioma stem cells. Can Res. 2024;84(3):372–87.
Zhang C, Zhou Y, Gao Y, Zhu Z, Zeng X, Liang W, Sun S, Chen X, Wang H. Radiated glioblastoma cell-derived exosomal circ_0012381 induce M2 polarization of microglia to promote the growth of glioblastoma by CCL2/CCR2 axis. J Transl Med. 2022;20(1):388.
Yoshimoto K, Mizoguchi M, Hata N, Murata H, Hatae R, Amano T, Nakamizo A, Sasaki T. Complex DNA repair pathways as possible therapeutic targets to overcome temozolomide resistance in glioblastoma. Front Oncol. 2012;2:186.
Wu P, Guo J, Yang H, Yuan D, Wang C, Wang Z. Exosomes derived from hypoxic glioma cells reduce the sensitivity of glioma cells to temozolomide through carrying miR-106a-5p. Drug Des Devel Ther. 2022;16:3589–98.
Yin J, Zeng A, Zhang Z, Shi Z, Yan W, You Y. Exosomal transfer of miR-1238 contributes to temozolomide-resistance in glioblastoma. EBioMedicine. 2019;42:238–51.
Zeng A, Wei Z, Yan W, Yin J, Huang X, Zhou X, Li R, Shen F, Wu W, Wang X, et al. Exosomal transfer of miR-151a enhances chemosensitivity to temozolomide in drug-resistant glioblastoma. Cancer Lett. 2018;436:10–21.
Geng X, Zhang Y, Lin X, Zeng Z, Hu J, Hao L, Xu J, Wang X, Wang H, Li Q. Exosomal circWDR62 promotes temozolomide resistance and malignant progression through regulation of the miR-370-3p/MGMT axis in glioma. Cell Death Dis. 2022;13(7):596.
Ding C, Yi X, Chen X, Wu Z, You H, Chen X, Zhang G, Sun Y, Bu X, Wu X et al: Warburg effect-promoted exosomal circ_0072083 releasing up-regulates NANGO expression through multiple pathways and enhances temozolomide resistance in glioma. Journal of Experimental & Clinical Cancer Research 2021, 40(1).
Li X, Wang N, Leng H, Yuan H, Xu L. Hsa_circ_0043949 reinforces temozolomide resistance via upregulating oncogene ITGA1 axis in glioblastoma. Metab Brain Dis. 2022;37(8):2979–93.
Zhang Z, Yin J, Lu C, Wei Y, Zeng A, You Y. Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J Exp Clin Cancer Res. 2019;38(1):166.
Ellingson BM, Wen PY, Cloughesy TF. Modified criteria for radiographic response assessment in glioblastoma clinical trials. Neurotherapeutics. 2017;14(2):307–20.
Westphal M, Lamszus K. Circulating biomarkers for gliomas. Nat Rev Neurol. 2015;11(10):556–66.
Bullock MD, Silva AM, Kanlikilicer-Unaldi P, Filant J, Rashed MH, Sood AK, Lopez-Berestein G, Calin GA. Exosomal non-coding RNAs: diagnostic, prognostic and therapeutic applications in cancer. Noncoding RNA. 2015;1(1):53–68.
Wolburg H, Noell S, Fallier-Becker P, Mack AF, Wolburg-Buchholz K. The disturbed blood-brain barrier in human glioblastoma. Mol Aspects Med. 2012;33(5–6):579–89.
Saint-Pol J, Gosselet F, Duban-Deweer S, Pottiez G, Karamanos Y: Targeting and Crossing the Blood-Brain Barrier with Extracellular Vesicles. Cells 2020, 9(4).
García-Romero N, Carrión-Navarro J, Esteban-Rubio S, Lázaro-Ibáñez E, Peris-Celda M, Alonso MM, Guzmán-De-Villoria J, Fernández-Carballal C, de Mendivil AO, García-Duque S, et al. DNA sequences within glioma-derived extracellular vesicles can cross the intact blood-brain barrier and be detected in peripheral blood of patients. Oncotarget. 2017;8(1):1416–28.
Skog J, Wurdinger T, van Rijn S, Meijer DH, Gainche L, Sena-Esteves M, Curry WT Jr, Carter BS, Krichevsky AM, Breakefield XO. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10(12):1470–6.
Fraser K, Jo A, Giedt J, Vinegoni C, Yang KS, Peruzzi P, Chiocca EA, Breakefield XO, Lee H, Weissleder R. Characterization of single microvesicles in plasma from glioblastoma patients. Neuro Oncol. 2019;21(5):606–15.
Manda SV, Kataria Y, Tatireddy BR, Ramakrishnan B, Ratnam BG, Lath R, Ranjan A, Ray A. Exosomes as a biomarker platform for detecting epidermal growth factor receptor-positive high-grade gliomas. J Neurosurg. 2018;128(4):1091–101.
Shao H, Chung J, Balaj L, Charest A, Bigner DD, Carter BS, Hochberg FH, Breakefield XO, Weissleder R, Lee H. Protein typing of circulating microvesicles allows real-time monitoring of glioblastoma therapy. Nat Med. 2012;18(12):1835–40.
Zhou B, Xu K, Zheng X, Chen T, Wang J, Song Y, Shao Y, Zheng S. Application of exosomes as liquid biopsy in clinical diagnosis. Signal Transduct Target Ther. 2020;5(1):144.
Manterola L, Guruceaga E, Gallego Perez-Larraya J, Gonzalez-Huarriz M, Jauregui P, Tejada S, Diez-Valle R, Segura V, Sampron N, Barrena C, et al. A small noncoding RNA signature found in exosomes of GBM patient serum as a diagnostic tool. Neuro Oncol. 2014;16(4):520–7.
Lan F, Qing Q, Pan Q, Hu M, Yu H, Yue X. Serum exosomal miR-301a as a potential diagnostic and prognostic biomarker for human glioma. Cell Oncol (Dordr). 2018;41(1):25–33.
Shi R, Wang P-Y, Li X-Y, Chen J-X, Li Y, Zhang X-Z, Zhang C-G, Jiang T, Li W-B, Ding W, et al. Exosomal levels of miRNA-21 from cerebrospinal fluids associated with poor prognosis and tumor recurrence of glioma patients. Oncotarget. 2015;6(29):26971–81.
Chan JA, Krichevsky AM, Kosik KS. MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells. Cancer Res. 2005;65(14):6029–33.
Huang K, Fang C, Yi K, Liu X, Qi H, Tan Y, Zhou J, Li Y, Liu M, Zhang Y, et al. The role of PTRF/Cavin1 as a biomarker in both glioma and serum exosomes. Theranostics. 2018;8(6):1540–57.
Tan SK, Pastori C, Penas C, Komotar RJ, Ivan ME, Wahlestedt C, Ayad NG. Serum long noncoding RNA HOTAIR as a novel diagnostic and prognostic biomarker in glioblastoma multiforme. Mol Cancer. 2018;17(1):74.
Santangelo A, Imbruce P, Gardenghi B, Belli L, Agushi R, Tamanini A, Munari S, Bossi AM, Scambi I, Benati D, et al. A microRNA signature from serum exosomes of patients with glioma as complementary diagnostic biomarker. J Neurooncol. 2018;136(1):51–62.
Shao N, Xue L, Wang R, Luo K, Zhi F, Lan Q. miR-454-3p Is an Exosomal Biomarker and Functions as a Tumor Suppressor in Glioma. Mol Cancer Ther. 2019;18(2):459–69.
Kooijmans SA, Vader P, van Dommelen SM, van Solinge WW, Schiffelers RM. Exosome mimetics: a novel class of drug delivery systems. Int J Nanomedicine. 2012;7:1525–41.
Benecke L, Coray M, Umbricht S, Chiang D, Figueiro F, Muller L. Exosomes: Small EVs with Large Immunomodulatory Effect in Glioblastoma. Int J Mol Sci. 2021;22:7.
Monfared H, Jahangard Y, Nikkhah M, Mirnajafi-Zadeh J, Mowla SJ. Potential Therapeutic Effects of Exosomes Packed With a miR-21-Sponge Construct in a Rat Model of Glioblastoma. Front Oncol. 2019;9:782.
Katakowski M, Buller B, Zheng X, Lu Y, Rogers T, Osobamiro O, Shu W, Jiang F, Chopp M. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013;335(1):201–4.
Fareh M, Almairac F, Turchi L, Burel-Vandenbos F, Paquis P, Fontaine D, Lacas-Gervais S, Junier MP, Chneiweiss H, Virolle T. Cell-based therapy using miR-302-367 expressing cells represses glioblastoma growth. Cell Death Dis. 2017;8(3): e2713.
Munoz JL, Bliss SA, Greco SJ, Ramkissoon SH, Ligon KL, Rameshwar P. Delivery of functional anti-mir-9 by mesenchymal stem cell-derived exosomes to glioblastoma multiforme cells conferred chemosensitivity. Mol Ther Nucleic Acids. 2013;2(10): e126.
Qian C, Wang Y, Ji Y, Chen D, Wang C, Zhang G, Wang Y. Neural stem cell-derived exosomes transfer miR-124-3p into cells to inhibit glioma growth by targeting FLOT2. Int J Oncol. 2022;61:4.
Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, Klyachko NL, Inskoe E, Piroyan A, Sokolsky M, Okolie O, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–64.
Shan S, Chen J, Sun Y, Wang Y, Xia B, Tan H, Pan C, Gu G, Zhong J, Qing G, et al. Functionalized Macrophage Exosomes with Panobinostat and PPM1D-siRNA for Diffuse Intrinsic Pontine Gliomas Therapy. Adv Sci (Weinh). 2022;9(21): e2200353.
Liang S, Xu H, Ye BC. Membrane-decorated exosomes for combination drug delivery and improved glioma therapy. Langmuir. 2022;38(1):299–308.
Dong S, Liu X, Bi Y, Wang Y, Antony A, Lee D, Huntoon K, Jeong S, Ma Y, Li X, et al. Adaptive design of mRNA-loaded extracellular vesicles for targeted immunotherapy of cancer. Nat Commun. 2023;14(1):6610.
Acknowledgements
Thanks to the support of this research by the Natural Science Foundation of Anhui Province (2208085MH220), Anhui Medical University Foundation (2023xkj034).
Author information
Authors and Affiliations
Contributions
ZY, EB and BZ contributed to the study design. ZY, HW and ZW consulted literature and wrote the manuscript. All authors made substantial contributions to the study and provided the approval for the version to be submitted.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent to publication
All of author are consent for publication.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1
. The ESCRT- dependent and independent pathway are implicated in controlling the cargos sorting of exosomes. [20] Copyright 2020, Molecular Cancer.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yang, Z., Wu, H., Wang, Z. et al. The role and application of small extracellular vesicles in glioma. Cancer Cell Int 24, 229 (2024). https://doi.org/10.1186/s12935-024-03389-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03389-z