
Abstract
Background
Artificial induction of polyploidy is the most common and effective way to improve the biological properties of Populus and develop new varieties of this tree. In this study, in order to confirm and expand earlier findings, we established a protocol using colchicine and based on an efficient shoot regeneration system of leaf blades to induce tetraploidy in vitro in three genotypes from diploid Populus hopeiensis. The stomatal characteristics, leaf blade size, and growth were evaluated for diploids and tetraploids of three genotypes.
Results
We found that genotype, preculture duration, colchicine concentration, and colchicine exposure time had highly significant effects on the tetraploid induction rate. The optimal protocol for inducing tetraploidy in P. hopeiensis was to preculture leaf blades for 7 days and then treat them for 4 days with 40 mg/L colchicine. The tetraploid induction rates of genotypes BT1, BT3, and BT8 were 21.2, 11.4 and 16.7%, respectively. A total of 136 tetraploids were identified by flow cytometry analysis and somatic chromosome counting. The stomatal length, width, and density of leaf blades significantly differed between diploid and tetraploid plants. Compared with their diploid counterparts, the tetraploids produced larger leaf blades and had a slower growth rate. Our findings further document the modified morphological characteristics of P. hopeiensis following whole-genome duplication (e.g., induced tetraploidy).
Conclusions
We established a protocol for in vitro induction of tetraploidy from diploid leaf blades treated with colchicine, which can be applied to different genotypes of P. hopeiensis.
Similar content being viewed by others


Background
Triploid breeding is one of the most effective approaches for Populus genetic improvement. For example, growth rates [1, 2], fiber properties [3, 4], timber quality [5, 6], and biomass production [7,8,9] have been enhanced using a triploid breeding program of Populus. The improved performance of triploids is due to the combined effect of higher ploidy levels and heterosis [10]. The most direct method to achieve new triploid germplasms is to cross diploids with tetraploids [11,12,13]. However, the application of this method has been limited due to the lack of natural tetraploids. Therefore, artificial tetraploids induced by colchicine are useful as bridge parents to produce the triploids in Populus.
In vascular plants, somatic chromosome doubling has already been successfully implemented to generate new germplasms with higher ploidy levels. Woody perennials such as octaploid Ziziphus jujuba [14], Neolamarckia cadamba [15], Jatropha curcas [16], and Panicum virgatum L. [17, 18]; and hexaploid Populus [19, 20], Pennisetum [21, 22], and Prunus pseudocerasus [23], as well as tetraploid Populus [24,25,26], Ziziphus jujuba [27, 28], Robinia pseudoacacia [29, 30], Platanus acerifolia [31], Pogostemon cablin [32], Liquidambar styraciflua [33], and Lagerstroemia indica [34] are examples of successful colchicine induction to achieve somatic chromosome doubling. Different explant types in vascular plants have been induced to obtain somatic chromosome doubling: the explant types include seeds [35], zygotes [36,37,38], shoot tips [16, 30, 39], nodal segments [15, 20], leaf petioles [33, 40], and leaf blades [24, 28]. Hence, somatic chromosome doubling is a promising method to produce new polyploids.
Increase in the chromosome number of tetraploid plants is actually the result of whole-genome duplication events. It is worth mentioning that Populous in general (while currently diploid) has evidence of a whole genome duplication event [41, 42]. One of the most interesting features of whole-genome duplication is the increased dosage of all genes, which alters gene expression resulting in increased cell and organelle sizes, and ultimately, variation in the traits of tetraploid plants [43, 44]. Furthermore, whole-genome duplication events often confer tetraploid plants with superior traits compared to their isogenic diploid progenitor. For example, faster growth [28], larger leaf blades [24, 26, 32], thicker leaf blades [29, 31, 45], thicker stems [46, 47], larger flowers [27, 48, 49], and greater numbers of seeds [50] are often found in synthetic tetraploid plants. Additionally, several studies have shown that tetraploidization improves physiological tolerance to stress in several plant species, such as Ziziphus jujuba [51], Escallonia [49], Stevia rebaudiana [52], Dioscorea zingiberensis [53], Dendranthema nankingense [54]. Thus, tetraploidy is thought to play an important role in plant diversification and evolution.
Here, we focused on the model woody plant Populus (Populus hopeiensis) to investigate the phenotypic consequences of gene dosage effects. For this, a protocol for in vitro tetraploid induction of P. hopeiensis by colchicine treatment was established based on an efficient shoot regeneration system using leaf blades. To compare the different ploidy levels of the three genotypes, the responses of stomatal characteristics, leaf blade size, and growth traits were evaluated. Our data showed that tetraploids of P. hopeiensis were distinguished from their isogenic diploid progenitor by having larger stomata, larger leaf blades, and a slower growth rate.
Results
Shoot regeneration of leaf blades
Nine treatments were used to evaluate the effects of 6-BA, TDZ, and IAA on shoot regeneration from P. hopeiensis leaf blades. Adventitious shoots formed from leaf blades after 25 days of culture on solid shoot regeneration medium (Fig. 1A), and healthy adventitious shoots successfully regenerated from leaf blades after 45 days (Fig. 1B). The roots of regenerated plants were formed after 30 days of culture on rooting medium (Fig. 1C). The shoot regeneration rates from nine treatments are shown in Table 1. The shoot regeneration rates ranged from 16.7 to 96.7%. The univariate generalized linear model (GLM) analysis showed that genotype (F = 7.190, P = 0.001), 6-BA (F = 84.451, P = 0.000), TDZ (F = 21.486, P = 0.000), and IAA (F = 25.062, P = 0.000) had highly significant effects on the shoot regeneration rates. Least significant difference (LSD) multiple comparison tests revealed that the shoot regeneration rate of genotype BT1 was significantly higher than that of genotypes BT3 or BT8 (P < 0.05) (Table 2). The shoot regeneration rate was significantly higher on the medium containing 0.4 mg/L 6-BA than those on medium containing 0.3 or 0.2 mg/L 6-BA. The shoot regeneration rate was also significantly higher on medium containing 0.015 mg/L TDZ than those on medium containing 0.01 or 0.005 mg/L TDZ. However, the shoot regeneration rate was significantly lower on media containing 0.2 mg/L IAA than those on medium containing 0.1 or 0.05 mg/L IAA.

The regeneration of plants from leaf blades of P. hopeiensis. A Shoots formed from leaf blades after culture for 25 days. B Adventitious shoots regenerated from leaf blades after culture for 45 days. C Successfully rooted healthy regenerated plants
The number of shoots per explant from nine treatments are presented in Table 1. The number of shoots per explant varied from 1.2 to 11.7. The number of shoots per explant significantly varied among different genotypes (F = 9.268, P = 0.000) and among the hormone treatments—6-BA (F = 148.470, P = 0.000), TDZ (F = 34.747, P = 0.000), and IAA (F = 72.259, P = 0.000)—according to univariate GLM analysis. The LSD multiple comparison tests showed that the number of shoots per explant of genotypes BT1 or BT3 was significantly higher than that of genotype BT8 (P < 0.05) (Table 2). The number of shoots per explant was significantly higher on the medium containing 0.4 mg/L 6-BA than those on medium containing 0.3 or 0.2 mg/L 6-BA. The number of shoots per explant was also significantly higher on medium containing 0.015 mg/L TDZ than those on medium containing 0.01 or 0.005 mg/L TDZ. However, the number of shoots per explant was significantly lower on medium containing 0.2 or 0.05 mg/L IAA than those on medium containing 0.1 mg/L IAA. Based on these data, Murashige and Skoog (MS) medium containing 0.4 mg/L 6-BA, 0.015 mg/L TDZ, and 0.01 mg/L IAA was determined to be suitable for shoot regeneration from leaf blades of P. hopeiensis.
Tetraploid induction of leaf blades treated with colchicine
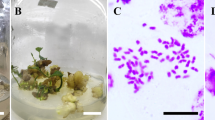
A total of 4,736 regenerated plants of the three genotypes were obtained from 27 treatments after leaf blades were treated with colchicine (Table 3). Individual regenerated plants were classified as diploid (Fig. 2A) or tetraploid (Fig. 2B) based on the peaks obtained by flow cytometry analysis. A total of 136 putative tetraploids were preliminarily determined. Somatic chromosome counting showed that the chromosome number of diploids was 2n = 2x = 38 (Fig. 2C), and all the putative tetraploids were ultimately confirmed to be real tetraploids (2n = 4x = 76, Fig. 2D).

Determination of the ploidy levels of regenerated P. hopeiensis plants. A Flow cytometry histogram of the diploids. B Flow cytometry histogram of the tetraploids. C Somatic chromosome number of the diploids (2n = 2x = 38). D Somatic chromosome number of the tetraploids (2n = 4x = 76)
The effects of genotype, preculture duration, colchicine concentration, and colchicine exposure times on the tetraploid induction rates of P. hopeiensis leaf blades were investigated (Table 3). The tetraploid induction rates ranged from 0 to 21.2%. The univariate GLM analysis revealed that genotype (F = 30.379, P = 0.000), preculture duration (F = 22.869, P = 0.000), colchicine concentration (F = 17.068, P = 0.000), and colchicine exposure time (F = 65.220, P = 0.000) had highly significant effects on the tetraploid induction rate. LSD multiple comparison tests showed that the tetraploid induction rate of genotype BT1 or BT3 was significantly lower than that of genotype BT8 (P < 0.05) (Table 4). The tetraploid induction rate following a preculture of 7 or 6 days was significantly higher than that following a preculture of 5 days. The tetraploid induction rate following treatment with 40 mg/L colchicine was significantly higher than that following treatment with 20 or 30 mg/L colchicine. The tetraploid induction rate following treatment with colchicine for 4 days was significantly higher than that following treatment for 2 or 3 days. Hence, the optimal protocol for inducing tetraploid P. hopeiensis was preculture of the leaf blades for 7 days, followed by treatment with 40 mg/L colchicine for 4 days.
Variations in stomata and morphological features in diploid and tetraploid plants
To assess the effect of ploidy level on leaf stomata, the lower epidermis of the leaf blades from 30-day-old sterile-rooted plantlets were used for stomata characteristic analysis. As shown in Fig. 3, the leaf stomata size was larger in tetraploids than in their diploid counterparts. The stomatal length, width, and density of diploid and tetraploid plants were measured (Fig. 4). The mean stomatal length and width in tetraploid plants of the three genotypes were significantly higher than that of diploids (Fig. 4A, B) (t-test, P < 0.01). The stomatal length and width of tetraploid plants were approximately twice that of diploid plants. However, the stomatal density in tetraploids of the three genotypes was significantly lower than that of diploids (Fig. 4C) (t-test, P < 0.01). The stomatal density of tetraploid plants was about half that of diploid plants. The results imply that stomatal characteristics are useful for distinguishing plant ploidy levels.

Stomata of the leaf blade in diploid (A) and tetraploid (B) plants of P. hopeiensis

The stomatal length (A), stomatal width (B), and stomatal density (C) in diploid and tetraploid plants of P. hopeiensis. The vertical bars show the standard error; the asterisk indicates significant differences between diploid and tetraploid plants (** p < 0.01)
To analyze the effect of ploidy level on phenotypic variation, 45 diploids and 45 tetraploids for each genotype were transplanted into plastic pots and transferred to greenhouse cultures for 6 months. Changes in the phenotypic traits of diploids and tetraploids of the three genotypes were characterized. The size of the leaf blades was much larger in tetraploid plants than in diploid plants (Fig. 5A). However, due to their slower growth rate, average plant height was lower in the six-month-old tetraploids than in the diploids (Fig. 5B). Pronounced differences in growth were observed among the three genotypes; the growth of the ramets of genotype BT1 was the fastest.

Morphological features in diploid and tetraploid plants of P. hopeiensis. A Fully expanded leaf blades of diploids and tetraploids. B Transplants of regenerated diploids and tetraploids. D1, D3, and D8 represent the diploids of genotypes BT1, BT3, and BT8, respectively. T1, T3, and T8 represent the tetraploids of genotypes BT1, BT3, and BT8, respectively
The leaf length, leaf width, basal diameter, and plant height of diploid and tetraploid plants were further analyzed (Fig. 6). The mean leaf length and width in tetraploid plants of the three genotypes were significantly larger than that of diploids (Fig. 6A, B) (t-test, P < 0.01). However, the mean basal diameter and plant height in tetraploid plants of the three genotypes were significantly lower than that of diploids (Fig. 6C, D) (t-test, P < 0.01). The basal diameter of genotypes BT1, BT3, and BT8 decreased by 17.1, 17.2, and 22.5%, respectively. The plant height of genotypes BT1, BT3, and BT8 decreased by 38.6, 36.8, and 28.9%, respectively. These results indicate that the whole-genome duplication leads to tetraploids exhibiting a dwarf phenotype.

The leaf length A, leaf width B, basal diameter C, and plant height D in diploid and tetraploid plants of P. hopeiensis. The vertical bars show the standard error; the asterisk indicates significant differences between diploid and tetraploid plants (** p < 0.01)
Discussion
Effects of genotype, 6-BA, TDZ, and IAA on shoot regeneration from leaf blades
Combining adventitious shoot regeneration from leaf blades with somatic chromosome doubling is a credible approach for creating new polyploids [24, 28, 55]. The shoot regeneration rate and the number of newly-regenerated shoots play a key role in affecting the polyploidization efficiency [24, 26]. Several studies have shown that adventitious shoot regeneration from leaf blades is affected by both the explant genotypes and the culture conditions, including the medium composition and addition of plant growth regulators [25, 56,57,58]. Li et al. [59] observed a significant correlation between genotype and leaf differentiation rate. Xu et al. [60] reported significant differences in the patterns of adventitious shoot initiation and callus development among different genotypes. In this study, we also found that the genotype had highly significant effects on the shoot regeneration rates and the number of shoots per explant, suggesting that the callus development of leaf blades among genotypes was inconsistent. These observations corroborate the findings of previous studies [60, 61].
In Populus, 6-BA, TDZ and IAA are widely used for adventitious shoot regeneration from leaf blades [56,57,58]. 6-BA and TDZ are the primary cytokinins that efficiently induce shoot regeneration and multiplication from leaf explants [24, 56]. However, high concentrations of 6-BA and TDZ may lead to the vitrification of the regenerated shoots. Previous studies documented that TDZ combined with 6-BA had a greater effect on the induction of adventitious shoots from leaf blades than 6-BA or TDZ used alone [26, 33, 56]. In the present study, 6-BA and TDZ had highly significant effects on shoot regeneration rates and the number of shoots produced per explant. The highest shoot regeneration rate and the number of shoots per explant were observed when MS medium contained 0.4 mg/L 6-BA and 0.015 mg/L TDZ, and no vitrification of regenerated shoots was observed under these conditions.
IAA is considered as a primary endogenous auxin. It plays an important role in regulating adventitious shoot regeneration from leaf blades [57]. Several previous studies have reported that the concentration and ratio of cytokinin and auxin are the main factors inducing shoot regeneration from leaf explants [56,57,58]. In general, low IAA concentrations can promote adventitious shoot elongation, thereby increasing the number of shoots per explant. However, high IAA concentrations may inhibit the induction and multiplication of adventitious shoots, leading to a reduced shoot regeneration rate [56,57,58]. Our results indicated that the shoot regeneration rates and the number of shoots per explant from P. hopeiensis leaf blades were obviously higher than those reported for Populus deltoides [57, 58], Populus tremula [62], and Populus pseudo-simonii [24].
Effects of genotype, preculture duration, and colchicine treatment on tetraploid induction
Most research on tetraploid induction technology aims to establish an efficient procedure for inducing tetraploids. However, the most suitable treatment conditions may change with the nuances of the explant genotype, which may result in a lower rate of tetraploid production [63, 64]. Zhang et al. [33] and Xu et al. [25] found that leaf blades of different genotypes responded differently to colchicine treatment during tetraploid production. In our study, highly significant effects of genotype on the tetraploid induction rate were also observed. This might be due to variations in sensitivity to colchicine among the genotypes. Similar results have been reported in previous studies [33, 60, 63].
Colchicine is an antimitotic chemical mutagen that has been commonly used to generate new germplasm with higher ploidy levels in vascular plants [26, 28]. It can inhibit the polymerization of microtubules by binding to tubulin. Microtubules are a fundamental component of the cytoskeleton and involved in the formation of the mitotic spindle. Therefore, inhibiting polymerization inhibits chromosome separation during cell division and leads to the formation of a polyploid cell [63, 64]. Colchicine has been widely used to generate new tetraploids by somatic chromosome doubling in some tree species, such as Populus [25, 26, 65], Platanus acerifolia [31], Pogostemon cablin [32], Robinia pseudoacacia [29, 30], Ziziphus jujuba [27, 28], Liquidambar styraciflua [33], and Lagerstroemia indica [34]. In the present study, colchicine was applied to induce chromosome doubling by treating leaf blades, and 136 tetraploids of P. hopeiensis were successfully obtained. Hence, colchicine treatment of leaf blades is a promising way for creating new polyploids.
The preculture duration of leaf blades before colchicine treatment is an important factor affecting the formation of polyploids [32, 66, 67]. Cai and Kang [24] reported that no tetraploid plants were produced from leaf explants of Populus pseudo-simonii Kitag when there was no preculture before colchicine treatment. Cui et al. [28]. showed that the most effective preculture duration for inducing the tetraploids of leaf blades in diploid Ziziphus jujuba by colchicine treatment was 10 days. Zhang et al. [33] documented that the most effective preculture duration for inducing tetraploids with leaf blades via colchicine was 8 days for diploid Liquidambar styraciflua. However, in our study, the most suitable preculture duration for colchicine-induced tetraploidy of leaf blades in P. hopeiensis was 7 days. Our finding was inconsistent with the reported results in previous studies [24, 28, 33]. It is likely that the most suitable preculture duration depends on the species. Variations may also be due to the fact cell responses vary with cell cycle stages during treatment [68].
Polyploid formation was also highly correlated with the colchicine concentration and exposure time. In general, low colchicine concentrations or short exposure times are not effective for inducing polyploidy [27, 32]; high colchicine concentrations or long exposure time can result in leaf blade mortality due to the toxicity of colchicine [24, 55]. In our study, the highest tetraploid induction rate (21.2%) was observed when the leaf blades of P. hopeiensis were treated with 40 mg/L colchicine for 4 days, which is slightly higher than what has been reported in many other studies [24, 28]. This suggests that using a suitable colchicine concentration and exposure time will help improve the polyploid induction rate.
Changes in ploidy-related morphological characteristics
Previous studies documented that the stomatal characteristics, such as stomatal length, stomatal width, and stomatal density, may be used as simple and efficient indicators to distinguish plant ploidy levels [29, 32, 54]. Furthermore, the stomata-based method is simple, virtually nondestructive, and does not require expensive instruments [28]. Here, we found that the stomatal length, width, and density of leaf explants in the induced tetraploids significantly differed from those observed in diploids. The stomatal length and width in tetraploids of the three genotypes were approximately twice that of diploid plants. However, the stomatal density in the tetraploids of the three genotypes was significantly lower than that of the diploids. Our results were consistent with the findings of previous studies [19, 55, 65, 69].
Polyploidy, or whole-genome duplication, provides genomic plasticity for functional divergence of duplicated genes, genomic and/or chromosomal recombination, transcriptome changes, and gene dosage effects, hence contributing to evolution [70,71,72]. An increase in DNA content and dosage effect of each gene, as is the case in polyploids, usually leads to an increase in organ size. Tsukaya [68] showed that the organs/bodies of the tetraploids in Arabidopsis were larger than the organs/bodies of diploids. Li et al. [29] documented that the leaf length and width were significantly larger in the tetraploids than in the diploids of Robinia pseudoacacia. In our study, tetraploids of P. hopeiensis produced larger leaf blades and had modified leaf blade morphology compared with diploids. However, the growth rate of the induced tetraploids in P. hopeiensis was also slower than that of the diploids. This suggests that gene dosage effects might increase the size of cells and organelles by altering gene expression, and this is generally accompanied by significant growth inhibition. Besides, the trend of trait variation after whole-genome duplication was consistent for all three genotypes of diploid P. hopeiensis. Hence, significant differences in the morphological characteristics between the diploids and the tetraploids may be a promising and stable tool for plant improvement. Additionally, tetraploid plants can be used as bridge parents to provide variation in chromosome number for the polyploid breeding program of Populus [11,12,13]. In future studies, we will focus on evaluating the growth and fertility of tetraploids and conduct interploid hybridation between the diploid and tetraploid P. hopeiensis individuals.
Conclusions
Genotype, 6-BA, TDZ, and IAA had highly significant effects on the shoot regeneration rates and the number of shoots per explant. The suitable shoot regeneration medium from leaf blades of P. hopeiensis was MS medium supplemented with 0.4 mg/L 6-BA, 0.015 mg/L TDZ, and 0.1 mg/L IAA. The highest shoot regeneration rate and the number of shoots per explant were 96.7% and 11.7, respectively. The tetraploid induction rate varied highly significantly among different genotype, preculture duration, colchicine concentration, and colchicine exposure time. The optimal protocol for inducing tetraploids of different genotypes P. hopeiensis was leaf blades precultured for 7 days and then treated for 4 days by 40 mg/L colchicine, which can be applied to different genotypes of P. hopeiensis. The tetraploid induction rates of genotypes BT1, BT3, and BT8 were 21.2, 11.4, and 16.7%, respectively. A total of 136 tetraploids were determined by flow cytometry analysis and somatic chromosome counting. The stomatal length, width, and density of leaf blades significantly differed between diploid and tetraploid plants of the three genotypes. After six months of culture, the length and width of tetraploid leaf blades of the three genotypes were significantly larger than those of the diploids. However, the basal diameter and plant height were lower in tetraploids of the three genotypes than that of the diploids. In summary, the morphological features of tetraploid plants differed obviously and significantly from those of diploid plants.
Methods
Plant materials
The State Key Laboratory of Efficient Production of Forest Tree Resource, Beijing Forestry University, China, provided tissue culture sterile-rooted plantlets of the diploid P. hopeiensis genotypes (BT1, BT3, and BT8). These materials were transferred into rooting medium containing 0.2 mg/L IBA, 0.65% (w/v) agar, 3% (w/v) sucrose, and half-strength Murashige and Skoog (MS) medium [73] for rooting. All media were adjusted to pH 5.8–6.2, and subsequently autoclaved at 121℃ for 15 min. All cultures were illuminated by fluorescent tubes of 2,000 lx with a 14 h photoperiod in a growth chamber at 25 °C.
Shoot regeneration of leaf blades
Fully expanded leaf blades from 30-day-old sterile-rooted plantlets were partially transected in two locations crossing the main vein so the leaf remained intact; they were then transferred to MS basal medium containing different concentrations of 6-BA (0.2, 0.3, and 0.4 mg/L), TDZ (0.005, 0.01, and 0.015 mg/L) and IAA (0.05, 0.1, and 0.2 mg/L). The effects of 6-BA, TDZ, and IAA on shoot regeneration were studied utilizing an orthogonal experimental design (Table 1). After 45 days in culture, the number of regenerated leaf blades and regenerated shoots per treatment was recorded to determine the shoot regeneration rates and the number of shoots per explant. The experiments were replicated three times with 10 explants per treatment for each genotype.
Tetraploid induction by colchicine treatment
The leaf blades were partially transected in two locations crossing the main vein, so the leaf remained intact. The leaf blades were then transferred to MS shoot regeneration medium containing 0.4 mg/L 6-BA, 0.015 mg/L TDZ, and 0.01 mg/L IAA, 0.65% (w/v) agar, and 3% (w/v) sucrose for 5, 6, and 7 days. Explants were then immersed in the same liquid shoot regeneration medium with filter-sterilized colchicine at concentrations of 20, 30, and 40 mg/L and treated in the dark for 2, 3, and 4 days, respectively. The 27 treatments are listed in Table 3. Following colchicine treatment, the explants were transferred to fresh solid shoot regeneration medium without colchicine after being rinsed three times with sterile distilled water. The experiments were replicated three times with 10 explants per treatment for each genotype.
After culture for 45 days, single shoots regenerated from the injuries of leaf blades were excised and transferred to the rooting medium. The number of regenerated shoots and tetraploids per explant were recorded to determine tetraploid induction rates. Subsequently, every 30 days, apical buds excised from the sterile-rooted plantlets were transferred to fresh solid rooting medium for culture.
Ploidy level determination
The ploidy level of all regenerated plantlets arising from leaf blades by colchicine treatment was preliminarily determined by flow cytometry analysis using the same procedures, reagents, and cytometers as those used by Wu et al. [26]. The standard peak of leaf blades from untreated diploid plants was set to appear at channel 50 relative fluorescent intensity. Therefore, the regenerated plantlets were considered as tetraploids when their peaks appeared at channel 100 relative fluorescent intensity.
The putative tetraploid plants were further confirmed by chromosome counting. Stem tips were harvested from 25-day-old sterile-rooted plantlets and pretreated in a saturated solution of paradichlorobenzene for 2–4 h. Then, they were washed three times with distilled water and fixed in fresh Carnoy’s solution (acetic acid: ethanol, 1:3) for 24 h at 4℃. The fixed samples were dissociated in 38% HCl for 25 min and washed with distilled water three times for 15 min. Subsequently, the dissociated samples were chopped on a glass slide in a drop of carbol-fuchsin solution and covered with a coverslip. The samples were observed and photographed under a 100 × oil lens using an Olympus BX 51 microscope. The ploidy number was verified from 10 randomly sampled cells of each sample.
Analysis of stomatal and morphological characteristics
Mature leaf blades from 30-day-old sterile-rooted plantlets were used for the analysis of stomatal characteristics. Six tetraploid and diploid plantlets (three each) were sampled for observation of stomata. Stomatal length, width, and density from tetraploid and diploid leaf blades were measured following previously described methods [55].
After 30 days of rooting culture, 45 diploid and 45 tetraploid rooted plantlets with prolific root systems from each genotype were randomly sampled, subsequently transplanted into plastic pots that were filled with potting media (turf soil: vermiculite: perlite, 2:1:1), and placed in a greenhouse. All the plants for the growth experiment were randomly planted and placed to reduce environmental influence. After six months of culture, the leaf length, leaf width, basal diameter, and plant height of diploid and tetraploid plants of the three genotypes were measured. Plants tested in the greenhouse were 15 replicates per genotype and ploidy level, and these plants were randomly selected.
Statistical analysis
Statistical analyses were implemented using IBM SPSS Statistics 20.0 software (IBM Inc., New York, USA). Before analysis of variance, percentage data were converted to proportions (p/100), and then arcsine square root-transformed to satisfy the heteroscedasticity assumptions. The univariate GLM analyses were used to analyze variation in the shoot regeneration rates, the number of shoots per explant, and tetraploid induction rates. The LSD multiple comparison tests were performed to assess the significance of differences between treatments; the threshold for statistical significance in these tests was P < 0.05. Two-sample t-tests were conducted to evaluate whether diploid and tetraploid plants significantly differed in stomatal length, stomatal width, stomatal density, leaf length, leaf width, basal diameter, and plant height.
Availability of data and materials
All data generated or analysed during this study are included in this published article.
Abbreviations
- MS:
-
Murashige and Skoog basal medium
- 6-BA:
-
6-Benzylaminopurine
- TDZ:
-
Thidiazuron
- IAA:
-
Indole-3-acetic acid
- IBA:
-
Indole-3-butyric acid
- GLM:
-
General linear model
- LSD:
-
Least significant difference
References
Zhu ZT, Lin HB, Kang XY. Studies on allotriploid breeding of Populus tomentosa B301 clones. Sci Silvae Sin. 1995;31:499–505.
Zhu ZT, Kang XY, Zhang ZY. Studies on selection of natural triploid of Populus tomentosa. Sci Silvae Sin. 1998;34:22–31.
Zhang PD, Wu F, Kang XY. Genetic control of fiber properties and growth in triploid hybrid clones of Populus tomentosa. Scand J Forest Res. 2013;28(7):621–30.
Wu J, Zhou Q, Sang YR, Kang XY, Zhang PD. Genotype-environment interaction and stability of fiber properties and growth traits in triploid hybrid clones of Populus tomentosa. BMC Plant Biol. 2021;21(1):405.
Zhang PD, Wu F, Kang XY. Chemical properties of wood are under stronger genetic control than growth traits in Populus tomentosa Carr. Ann for Sci. 2014;72(1):89–97.
Zhang PD, Wu F, Kang XY. Genotypic variation in wood properties and growth traits of triploid hybrid clones of Populus tomentosa at three clonal trials. Tree Genet Genomes. 2012;8(5):1041–50.
Zhang PD, Wu F, Kang XY, Zhao CG, Li YJ. Genotypic variations of biomass feedstock properties for energy in triploid hybrid clones of Populus tomentosa. Bioenerg Res. 2015;8(4):1705–13.
Hennig A, Kleinschmit JRG, Schoneberg S, Löffler S, Janßen A, Polle A. Water consumption and biomass production of protoplast fusion lines of poplar hybrids under drought stress. Front Plant Sci. 2015;6:330.
Guo LQ, Zhang JG, Liu XX, Rao GD. Polyploidy-related differential gene expression between diploid and synthesized allotriploid and allotetraploid hybrids of Populus. Mol Breeding. 2019;39(5):717871.
Qin J, Mo R, Li H, Ni Z, Sun Q, Liu Z. The transcriptional and splicing changes caused by hybridization can be globally recovered by genome doubling during allopolyploidization. Mol Biol Evol. 2021;38(6):2513–9.
Johnsson H. The triploid progeny of the cross diploid × tetraploid Populus tremula. Hereditas. 1945;31:411–40.
Lu N, Dai L, Wu B, Zhang Y, Luo ZJ, Xun SH, Sun YH, Li Y. A preliminary study on the crossability in Robinia pseudoacacia L. Euphytica. 2015;206(3):555–66.
Yan FF, Wang LH, Zheng XJ, Luo Z, Wang JR, Liu MJ. Acquisition of triploid germplasms by controlled hybridisation between diploid and tetraploid in Chinese jujube. J Hortic Sci Biotech. 2018;94(1):123–9.
Shi QH, Liu P, Liu MJ, Wang JR, Xu J. A novel method for rapid in vivo induction of homogeneous polyploids via calluses in a woody fruit tree (Ziziphus jujuba Mill). Plant Cell Tiss Org. 2015;121(2):423–33.
Eng WH, Ho WS, Ling KH. In vitro induction and identification of polyploid Neolamarckia cadamba plants by colchicine treatment. PeerJ. 2021;9:e12399.
Niu LJ, Tao YB, Chen MS, Fu QT, Dong YL, He HY, Xu ZF. Identification and characterization of tetraploid and octoploid Jatropha curcas induced by colchicine. Caryologia. 2016;69(1):58–66.
Yang ZY, Shen ZX, Tetreault H, Johnson L, Friebe B, Frazier T, Huang LK, Burklew C, Zhang XQ, Zhao BY. Production of autopolyploid lowland switchgrass lines through in vitro chromosome doubling. Bioenerg Res. 2013;7(1):232–42.
Yue YS, Zhu Y, Fan XF, Hou XC, Zhao CQ, Zhang S, Wu JY. Generation of octoploid switchgrass in three cultivars by colchicine treatment. Ind Crop Prod. 2017;107:20–1.
Liu WT, Zheng YF, Song SY, Huo BB, Li DL, Wang J. In vitro induction of allohexaploid and resulting phenotypic variation in Populus. Plant Cell Tiss Org. 2018;134(2):183–92.
Zeng QQ, Liu Z, Du K, Kang XY. Oryzalin-induced chromosome doubling in triploid Populus and its effect on plant morphology and anatomy. Plant Cell Tiss Org. 2019;138(3):571–81.
Campos JMS, Davide LC, Salgado CC, Santos FC, Costa PN, Silva PS, Alves CCS, Viccini LF, Pereira AV. In vitro induction of hexaploid plants from triploid hybrids of Pennisetum purpureum and Pennisetum glaucum. Plant Breeding. 2009;128:101–4.
Yue YS, Fan XF, Hu YF, Han C, Li H, Teng WJ, Zhang H, Teng K, Wen HF, Yang XJ, et al. In vitro induction and characterization of hexaploid Pennisetum × advena, an ornamental grass. Plant Cell Tiss Org. 2020;142(2):221–8.
James DJ, Mackenzie KAD, Malhotra SB. The induction of hexaploidy in cherry rootstocks using in vitro regeneration techniques. Theor Appl Genet. 1987;73:589–94.
Cai X, Kang XY. In vitro tetraploid induction from leaf explants of Populus pseudo-simonii Kitag. Plant Cell Rep. 2011;30(9):1771–8.
Xu CP, Huang Z, Liao T, Li Y, Kang XY. In vitro tetraploid plants regeneration from leaf explants of multiple genotypes in Populus. Plant Cell Tiss Org. 2016;125(1):1–9.
Wu J, Sang YR, Zhou Q, Zhang PD. Colchicine in vitro tetraploid induction of Populus hopeiensis from leaf blades. Plant Cell Tiss Org. 2020;141(2):339–49.
Gu XF, Yang AF, Meng H, Zhang JR. In vitro induction of tetraploid plants from diploid Zizyphus Jujuba Mill. Cv. Zhanhua. Plant Cell Rep. 2005;24(11):671–6.
Cui YH, Hou L, Li X, Huang FY, Pang XM, Li YY. In vitro induction of tetraploid Ziziphus jujuba Mill. Var. Spinosa plants from leaf explants. Plant Cell Tiss Org. 2017;131(1):175–82.
Li XY, Zhang ZJ, Ren YH, Feng Y, Guo Q, Dong L, Sun YH, Li Y. Induction and early identification of tetraploid black Locust by hypocotyl in vitro. In Vitro Cell Dev-Pl. 2021;57(3):372–9.
Ewald D, Ulrich K, Naujoks G, Schröder MB. Induction of tetraploid poplar and black Locust plants using colchicine: chloroplast number as an early marker for selecting polyploids in vitro. Plant Cell Tiss Org. 2009;99(3):353–7.
Liu GF, Li ZN, Bao MZ. Colchicine-induced chromosome doubling in Platanus acerifolia and its effect on plant morphology. Euphytica. 2007;157(1–2):145–54.
Widoretno W. In vitro induction and characterization of tetraploid patchouli (Pogostemon cablin Benth.) Plant. Plant Cell Tiss Org. 2016;125(2):261–7.
Zhang Y, Wang ZW, Qi SZ, Wang XQ, Zhao J, Zhang JF, Li BL, Zhang YD, Liu XZ, Yuan W. In vitro tetraploid induction from leaf and petiole explants of hybrid sweetgum (Liquidambar styraciflua × Liquidambar formosana). Forests. 2017;8(8):264.
Zhang QY, Luo FX, Liu L, Guo FC. In vitro induction of tetraploids in crape myrtle (Lagerstroemia indica L). Plant Cell Tiss Org. 2010;101(1):41–7.
Mo L, Chen JH, Chen F, Xu QW, Tong ZK, Huang HH, Dong RH, Lou XZ, Lin EP. Induction and characterization of polyploids from seeds of Rhododendron Fortunei Lindl. J Integr Agr. 2020;19(8):2016–26.
Wang J, Shi L, Song SY, Tian J, Kang XY. Tetraploid production through zygotic chromosome doubling in Populus. Silva Fennica. 2013;47(2):12.
Lu M, Zhang PD, Wang J, Kang XY, Wu JY, Wang XJ, Chen Y. Induction of tetraploidy using high temperature exposure during the first zygote division in Populus Adenopoda Maxim. Plant Growth Regul. 2013;72(3):279–87.
Guo LQ, Xu WT, Zhang Y, Zhang JF, Wei ZZ. Inducing triploids and tetraploids with high temperatures in Populus sect. Tacamahaca. Plant Cell Rep. 2017;36(2):313–26.
Yan XF, Zhang J, Zhang H. Induction and characterization of tetraploids in poplar. Plant Cell Tiss Org. 2021;146(1):185–9.
Nilanthi D, Chen XL, Zhao FC, Yang YS, Wu H. Induction of tetraploids from petiole explants through colchicine treatments in Echinacea purpurea L. J Biomed Biotechnol. 2009;2009:7.
Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U, Putnam N, Ralph S, Rombauts S, Salamov A, et al. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science. 2006;313(5793):1596–604.
Dai X, Hu Q, Cai Q, Feng K, Ye N, Tuskan GA, Milne R, Chen Y, Wan Z, Wang Z, et al. The willow genome and divergent evolution from poplar after the common genome duplication. Cell Res. 2014;24(10):1274–7.
Shi XL, Zhang CQ, Ko DK, Chen ZJ. Genome-wide dosage-dependent and -independent regulation contributes to gene expression and evolutionary novelty in plant polyploids. Mol Biol Evol. 2015;32(9):2351–66.
Zhang J, Liu Y, Xia EH, Yao QY, Liu XD, Gao LZ. Autotetraploid rice methylome analysis reveals methylation variation of transposable elements and their effects on gene expression. Proc Natl Acad Sci USA. 2015;112(50):7022–9.
Wu YX, Li WY, Dong J, Yang N, Zhao XM, Yang WD. Tetraploid induction and cytogenetic characterization for Clematis heracleifolia. Caryologia. 2013;66(3):215–20.
Głowacka K, Jeżowski S, Kaczmarek Z. In vitro induction of polyploidy by colchicine treatment of shoots and preliminary characterisation of induced polyploids in two Miscanthus species. Ind Crop Prod. 2010;32(2):88–96.
Tavan M, Mirjalili MH, Karimzadeh G. In vitro polyploidy induction: changes in morphological, anatomical and phytochemical characteristics of Thymus persicus (Lamiaceae). Plant Cell Tiss Org. 2015;122(3):573–83.
Allum JF, Bringloe DH, Roberts AV. Chromosome doubling in a Rosa rugosa Thunb. Hybrid by exposure of in vitro nodes to oryzalin: the effects of node length, oryzalin concentration and exposure time. Plant Cell Rep. 2007;26(11):1977–84.
Denaeghel HER, Van Laere K, Leus L, Lootens P, Van Huylenbroeck J, Van Labeke MC. The variable effect of polyploidization on the phenotype in Escallonia. Front Plant Sci. 2018;9:354.
Xue H, Zhang B, Tian JR, Chen MM, Zhang YY, Zhang ZH, Ma Y. Comparison of the morphology, growth and development of diploid and autotetraploid ‘Hanfu’ apple trees. Sci Hort. 2017;225:277–85.
Li M, Zhang C, Hou L, Yang W, Liu S, Pang X, Li Y. Multiple responses contribute to the enhanced drought tolerance of the autotetraploid Ziziphus jujuba Mill. Var. Spinosa. Cell Biosci. 2021;11(1):119.
Xiang ZX, Tang XL, Liu WH, Song CN. A comparative morphological and transcriptomic study on autotetraploid Stevia rebaudiana (bertoni) and its diploid. Plant Physiol Bioch. 2019;143:154–64.
Zhang XY, Hu CG, Yao JL. Tetraploidization of diploid Dioscorea results in activation of the antioxidant defense system and increased heat tolerance. J Plant Physiol. 2010;167(2):88–94.
Liu S, Chen S, Chen Y, Guan Z, Yin D, Chen F. In vitro induced tetraploid of Dendranthema nankingense (Nakai) Tzvel. Shows an improved level of abiotic stress tolerance. Sci Hort. 2011;127(3):411–9.
Wu J, Cheng X, Kong B, Zhou Q, Sang Y, Zhang P. In vitro octaploid induction of Populus hopeiensis with colchicine. BMC Plant Biol. 2022;22(1):176.
Zeng QQ, Han ZQ, Kang XY. Adventitious shoot regeneration from leaf, petiole and root explants in triploid (Populus alba × P. glandulosa)× P. tomentosa. Plant Cell Tiss Org. 2019;138(1):121–30.
Yadav R, Arora P, Kumar D, Katyal D, Dilbaghi N, Chaudhury A. High frequency direct plant regeneration from leaf, internode, and root segments of Eastern Cottonwood (Populus deltoides). Plant Biotechnol Rep. 2009;3(3):175–82.
Thakur AK, Saraswat A, Srivastava DK. In vitro plant regeneration through direct organogenesis in Populus deltoides clone G48 from petiole explants. J Plant Biochem Biotechnol. 2011;21(1):23–9.
Li Y, Zhen H, Wang PQ, Li Y, Kang XY. Establishment of leafregeneration system for hybrids of Populus Cathayana Rehd. J Nucl Agric Sci. 2014;28:0978–84.
Xu CP, Zhang Y, Huang Z, Yao PQ, Li Y, Kang XY. Impact of the leaf cut callus development stages of Populus on the tetraploid production rate by colchicine treatment. J Plant Growth Regul. 2017;37(2):635–44.
Coleman G, Ernst S. Shoot induction competence and callus determination in Populus deltoids. Plant Sci. 1990;71:83–92.
Huang D, Dai W. Direct regeneration from in vitro leaf and petiole tissues of Populus tremula ‘Erecta.’ Plant Cell Tiss Org. 2011;107(1):169–74.
Dhooghe E, Van Laere K, Eeckhaut T, Leus L, Van Huylenbroeck J. Mitotic chromosome doubling of plant tissues in vitro. Plant Cell Tiss Org. 2011;104(3):359–73.
Khosravi P, Kermani MJ, Nematzadeh GA, Bihamta MR, Yokoya K. Role of mitotic inhibitors and genotype on chromosome doubling of Rosa. Euphytica. 2007;160(2):267–75.
Ren YY, Jing YC, Kang XY. In vitro induction of tetraploid and resulting trait variation in Populus alba × Populus glandulosa clone 84 K. Plant Cell Tiss Org. 2021;146(2):285–96.
Manzoor A, Ahmad T, Bashir MA, Hafiz IA, Silvestri C. Studies on Colchicine Induced chromosome doubling for enhancement of Quality traits in Ornamental plants. Plants (Basel). 2019;8(7):194.
Luo Z, Iaffaldano BJ, Cornish K. Colchicine-induced polyploidy has the potential to improve rubber yield in Taraxacum kok-saghyz. Ind Crop Prod. 2018;112:75–81.
Tsukaya H. Controlling size in multicellular organs: focus on the leaf. Plos Biol. 2008;6(7):e174.
Zhou Q, Wu J, Sang YR, Zhao ZY, Zhang PD, Liu MQ. Effects of colchicine on Populus canescens ectexine structure and 2n pollen production. Front Plant Sci. 2020;11:295.
Leitch AR, Leitch IJ. Genomic plasticity and the diversity of polyploid plants. Science. 2008;320(5875):481–3.
Van de Peer Y, Mizrachi E, Marchal K. The evolutionary significance of polyploidy. Nat Rev Genet. 2017;18(7):411–24.
Wu J, Kong B, Zhou Q, Sun Q, Sang Y, Zhao Y, Yuan T, Zhang P. SCL14 inhibits the functions of the NAC043-MYB61 Signaling Cascade to reduce the Lignin Content in Autotetraploid Populus hopeiensis. Int J Mol Sci. 2023;24(6): 5809.
Murashige T, Skoog F. A revised medium for rapid growth and bio assays with Tobacco tissue cultures. Physiol Plant. 1962;15(3):473–97.
Acknowledgements
We thank Mary Beth A., a senior editor at University of Rhode Island, for her language editing.
Funding
This study was supported by the National Science Foundation of China (31570646).
Author information
Authors and Affiliations
Contributions
PZ designed the research and revised the manuscript. JW, QZ, YS, YZ, BK, LL, JD, LM, and ML performed the experiments. JW analyzed the data and wrote the main manuscript. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
In this study, tissue culture plantlets of Populus hopeiensis were provided from the State Key Laboratory of Efficient Production of Forest Tree Resource, Beijing Forestry University, China. As one of the most widely distributed and commonly cultivated species of poplar in China, Populus hopeiensis is not an endangered species. Thus, no specific permits are required for sample collection of Populus hopeiensis. All methods in this study were performed in accordance with relevant institutional, national, and international guidelines and legislation.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wu, J., Zhou, Q., Sang, Y. et al. In vitro induction of tetraploidy and its effects on phenotypic variations in Populus hopeiensis. BMC Plant Biol 23, 557 (2023). https://doi.org/10.1186/s12870-023-04578-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04578-0