
Abstract
Background
The importance of uridine 5′-diphosphate glucose (UDP-G) synthesis and degradation on carbon (C) partitioning has been indicated in several studies of plant systems, whereby the kinetic properties and abundance of involved enzymes had a significant effect upon the volume of C moving into the hemicellulose, cellulose and sucrose pools. In this study, the expression of 136 genes belonging to 32 gene families related to UDP-G metabolism was studied in 3 major sugarcane organs (including leaf, internode and root) at 6 different developmental stages in 2 commercial genotypes.
Results
Analysis of the genes associated with UDP-G metabolism in leaves indicated low expression of sucrose synthase, but relatively high expression of invertase genes, specifically cell-wall invertase 4 and neutral acid invertase 1–1 and 3 genes. Further, organs that are primarily responsible for sucrose synthesis or bioaccumulation, i.e., in source organs (mature leaves) and storage sink organs (mature internodes), had very low expression of sucrose, cellulose and hemicellulose synthesis genes, specifically sucrose synthase 1 and 2, UDP-G dehydrogenase 5 and several cellulose synthase subunit genes. Gene expression was mostly very low in both leaf and mature internode samples; however, leaves did have a comparatively heightened invertase and sucrose phosphate synthase expression. Major differences were observed in the transcription of several genes between immature sink organs (roots and immature internodes). Gene transcription favoured utilisation of UDP-G toward insoluble and respiratory pools in roots. Whereas, there was comparatively higher expression of sucrose synthetic genes, sucrose phosphate synthase 1 and 4, and comparatively lower expression of many genes associated with C flow to insoluble and respiratory pools including myo-Inositol oxygenase, UDP-G dehydrogenase 4, vacuolar invertase 1, and several cell-wall invertases in immature internodes.
Conclusion
This study represents the first effort to quantify the expression of gene families associated with UDP-G metabolism in sugarcane. Transcriptional analysis displayed the likelihood that C partitioning in sugarcane is closely related to the transcription of genes associated with the UDP-G metabolism. The data presented may provide an accurate genetic reference for future efforts in altering UDP-G metabolism and in turn C partitioning in sugarcane.
Similar content being viewed by others

Introduction
In plants, uridine 5′-diphosphate glucose (UDP-G) is a nucleotide sugar that is consumed irreversibly to produce the major components of the cell wall, cellulose and hemicellulose [1,2,3,4]. UDP-G can also be degraded into sucrose then resynthesized [5,6,7]. As UDP-G is directly synthesised into/or degraded from sucrose, it is likely that this nucleotide sugar’s consumption is also linked to lignin production, which occurs via the hydrolysis of sucrose leading into the glycolytic and oxidative pentose phosphate pathways [8]. Other roles for UDP-G in plants include its role as a glucose residue donor in callose formation, glycoproteins, glycolipids and sulpholipids, and its essential role in the glycosylation of steroids, flavonoids, betalains, glucosinolates and terpenoids [9]. Further, several studies have suggested a central role for UDP-G as a signal molecule regulating growth and development, biomass accumulation, and programmed cell death [8, 10,11,12]. The synthesis and degradation of the UDP-G metabolite is central to the amount of carbon (C) moving into the major pools within the sugarcane plant, specifically sucrose, cellulose and hemicellulose pools [7]. The importance of UDP-G control and its effect on C partitioning has been indicated in several studies of plant systems, whereby the kinetic properties and abundance of enzymes involved in UDP-G synthesis/utilisation had a significant effect upon the volume of C moving into the insoluble and soluble pools [8, 13,14,15,16,17,18,19], see Fig. 1 for a summary.

Metabolism associated with the synthesis/degradation of the UDP-glucose metabolite. Abbreviations, CWI: Cell Wall Invertase; VINV: Vacuolar Invertase; CINV: Cytosolic Invertase; ANINV: Alkaline/Neutral Invertase; FK: Fructokinase; MIPS: myo-Inositol phosphate synthase; MIP: myo-Inositol phosphatase; MIOX: myo-Inositol oxygenase; GluK: Glucuronokinase; PGM: Phosphoglucomutase; UGPase: Uridine Diphosphate Glucose pyrophosphorylase; UGD: UDP-Glucose Dehydrogenase; AGP: Adenosine Diphosphate Glucose pyrophosphorylase; CesA: Cellulose Synthase; CSL: Cellulose Synthase-Like; G6PI: Glucose-6-Phosphate Isomerase; SPP: Sucrose Phosphate Phosphatase; SPS: Sucrose Phosphate Synthase; SuSy: Sucrose Synthase; RHM: Uridine Diphosphate Glucose 4,6-dehydratase; UAXS: Uridine Diphosphate Glucose Apiose/Xylose Synthase; GALE: Uridine Diphosphate Glucose 4-Epimerase; UGE: Uridine Diphosphate Glucuronic Acid Epimerase; UXS: Uridine Diphosphate Xylose Synthase; UXE: Uridine Diphosphate Xylose Epimerase; HXK: Hexokinase
A gene family is a set of functionally similar genes formed via whole genome duplication and derived originally from a single ancestral gene [20]. The value for plant systems having developed multiple gene isoforms coding for the same enzyme is that often small alterations to enzyme structure can confer differing affinities to substrates and feedback inhibitory molecules, meaning the flux through this point in the pathway can continue in a variety of cellular conditions [4, 21, 22]. The value in understanding the full array of gene family isoforms was shown in an attempt to downregulate an important consumer of UDP-G into the hemicellulosic precursor UDP-glucuronic acid, via the activity of UDP-G dehydrogenase (UGD) in sugarcane. A sugarcane genetic knockout mutant of a single UGD gene was created in [23]. Cessation of UGD activity was expected to have a reductive effect on the hemicellulose fraction, however, no differences in composition were observed. Upregulation of the myo-inositol pathway, which provides an alternative pathway for C to enter hemicellulose synthesis, was detected in the mutant, which was used to explain the lack of compositional difference in comparison to the control [23]. Also, based on previous characterisations of the UGD gene family in other plants [4, 21], it is highly unlikely that there is a single UGD gene in the sugarcane genome, which may also have contributed to the reasons why no difference in cell wall composition was recorded in the UGD single mutant. Therefore, in sugarcane, successful alterations of UDP-glucuronic acid synthesis is likely contingent on changes to genes associated with the myo-Inositol pathway, but also the identification and alteration of additional UGD genes. In a related study within the Arabidopsis genome [14], of the 4 UGD genes, only double genetic knockout mutants of UGD 2 and UGD 3 genes combined led to reduced UDP-glucuronic acid synthesis. Suppression of UDP-glucuronic acid synthesis caused a significant decrease in the hemicelluloses arabinose, xylose, apiose and galacturonic acid, in turn causing dwarfism and other developmental defects in the double mutants [14]. Single knockout mutants of UGD 2 or UGD 3 did not affect the cell wall composition, which indicates the activity of only one of these genes is sufficient to make up for the lack of expression in the other, whilst the other isoforms likely had organ-specific functions. In a related study within Arabidopsis UGD 2 and 3 had significantly higher expression than isoforms 1 and 4 in all tissues [21]. This result may suggest that gene family members with significantly higher expression have a larger effect on the pathway than isoforms with lower expression. This is also supported by the finding that each UDG isoform had differing affinity for the substrate. UGD 3 had a far higher affinity and catalytic constant, suggesting it is responsible for the bulk of C assimilation into the hemicellulose pool [21], in turn explaining the adverse effects of knockouts. Studies of multi-gene families related to the UDP-G metabolism have reported similar results, whereby knockout mutants of 1, 2 or even 3 isoforms were found to have a disproportionate effect on the physiology of the mutant, relating to alterations in the cell wall [15, 24,25,26,27,28,29,30].
Modern high throughput technologies such as next-generation sequencing (NGS) allow a platform to look broadly at differences in gene expression in different tissues and organs [31]. In this study, to study the organ-specific expression of genes associated with the UDP-G metabolism in the sugarcane plant, we used mature sugarcane leaves (dedicated source organ), young internodes (sink organ where the bulk of UDP-G and hexose phosphates are utilised in cellulose, hemicellulose, protein synthesis and respiration), maturing and mature internodes (sink organ where respiration is limited and UDP-G is utilised in sucrose storage) and root (a meristematic sink that has similar C requirements to immature internodes) [32]. These differing metabolic requirements suggest the action of different mechanisms at the transcriptomic level in related metabolic pathways. As the metabolism of UDP-G is central to key sources of C deposition, it stands to reason that the expression of related genes would have differed expression as related to how C is utilised in that tissue. Additionally, most of these genes are in multi-gene families, and many multi-gene families likely contain isoforms that overall have a larger effect on metabolism than others. By determining which members are expressed and in which tissue, gene candidates for altering UDP-G metabolism and by proxy C metabolism could be identified.
Materials and methods
Plant material collection, data generation and transcript profiling
The plant material used in this experiment is derived from two commercial sugarcane genotypes, KQ228 and Q208 provided by Sugar Research Australia (SRA). Two sugarcane genotypes were analysed as an additional validatory mechanism. KQ228 and Q208 share common ancestry and agronomic traits, so they were expected to have comparable transcriptomic profiles [33]. Agronomic details of both genotypes can be found in Table S1, within Supplementary Data File 1. Leaf and internode samples of both genotypes were taken from 9-month old commercial stands. Root samples were obtained from 3-month old research plants in ‘soft’ above-ground pots. For sampling, schematic see [32]. Briefly, a total of 6 samples were collected at different developmental stages from three major sugarcane organs, including internodes, leaves and root. Hereafter, these samples are referred to as TI (Top Internode), MI (Middle Internode), BI (Bottom Internode), L1 (1st Visible Dewlap Leaf), L5 (5th Visible Dewlap Leaf) and R (Root). All samples were collected in triplicate and ‘snap frozen’ in liquid nitrogen within a minute of excision, put on dry ice in transit and kept in a − 80 °C freezer before preparation. Before RNA extraction samples were homogenized in cryogenic conditions. RNA extractions were conducted using the combined Trizol kit and RNeasy Plant minikit, as described in [34]. Samples were prepared with Illumina TruSeq™ stranded total RNA library prep kit, using 1000 ng of the submitted RNA, by QBI (Queensland Brain Institute, University of Queensland, St Lucia QLD, 4072). Each sample was sequenced in 3 lanes using an Illumina HiSeq2000 instrument to obtain 125 bp paired end reads. Read data was assessed using FastQC [35], to determine the quality and adapter sequences. A full-length sugarcane Iso-seq transcriptome database (SUGIT) developed by Hoang et al. 2017, was utilised as the reference sequence [36], in the assessment of transcript specific gene expression. The pipeline used to profile the transcriptome of the multiple samples in this experiment is based on the protocol ‘Expression Analysis using RNA-seq’ by QIAGEN bioinformatics [37] in the CLC Genomics Workbench version 12 (CLC-GWB, CLC Bio-QIAGEN, Aarhus, Denmark). The CLC-GWB analyses were conducted on a CLC Genomics Server, the CLC server, nodes and CLC-clients which are part of the Robert Henry Bioinformatics infrastructure at QAAFI, The University of Queensland, Australia. All CLC infrastructure was hosted by the Research Computing Centre (RCC), based at the University of Queensland, Australia [38]. For further details of plant collection and data generation, details see [39].
Extraction of transcripts of interest and subsequent matching to gene family isoforms
A search for all transcripts associated with gene families related to the consumption/synthesis of the UDP-G metabolite was conducted in the sugarcane SUGIT transcriptome. Using CLC-GWB, relevant search terms, i.e. “sucrose synthase”, were queried against the SUGIT transcript annotations, to derive lists of relevant transcripts, which were extracted as FASTA files. Additional mapping was undertaken using the large-gap mapping tool in CLC-GWB (length fraction of 0.8 and similarity fraction of 0.5), whereby genes of interest from the Sorghum bicolor and Zea mays genomes were mapped against the SUGIT transcriptome, to ensure any mislabeled transcripts were included in downstream analysis. All extracted transcripts were blasted against the S. bicolor genome (accession number NC_012875.2) using NCBI’s nucleotide Basic Local Alignment Search Tool (BLAST) [40], with default settings in blastn, to group transcripts into their respective gene family homologues. The top BLAST hit based on query coverage and per cent identity was used to define the transcripts gene identity.
Genomic location and gene length determination using the sugarcane STP monoploid genome
The sugarcane monoploid genome, a 382-Mb high-quality single-tiling-path (STP) sequence [41], enabled the genomic location and gene length to be determined in most of the genes of interest. The sequences of the genes of interest were also extracted and mapped to the extracted SUGIT transcripts (in CLC-GWB) to ensure paralogous genes (derived from recent genome duplications) were assigned the correct transcripts. To find the genes of interest within the sugarcane monoploid genome, the coding region of equivalent sorghum genes were blasted against the STP sequence. Sequences with high degrees of similarity were extracted then blasted against the sorghum genome to ensure the manual annotation was correct. Enzyme concession numbers for each gene family was defined by blasting the corresponding sorghum sequence in the UniProt database [42].
Collapsing transcripts into a single expression value
The large degree of alternative splices and redundancy within the SUGIT reference transcriptome could reduce the accuracy of expression comparisons between the sugarcane organ types included in this study. Collapsing the normalised read counts (normalised using transcript per million (TPM) calculation) of all transcripts related to a gene, into a single value was deemed to be a far better option for determining the importance of specific gene family homologues, as the more isoforms a specific gene has in a reference transcriptome the less accurate differential gene expression comparisons will be [43]. The RNA-seq application within CLC-GWB assigns reads exclusively to the reference, which means in the case of a read sharing the same identity with two reference sequences it will be assigned to one of the references randomly. Due to this, quantifying the expression of a gene that has multiple isoforms, was simply done by adding the TPM values together to form a single TPM value (Fig. S1).
Data processing of gene family expression values
The statistical significance of the mean of the biological replicates was calculated utilising the one-way ANOVA function, with the additional Tukey’s post-hoc t-test (honestly significant difference, HSD test) within IBM SPSS Statistics v27 (SPSS Inc., Chicago, IL, USA). The null hypothesis was accepted at a p-value of > 0.05. Minimum and maximum outliers (two-sided) were removed from some datasets using the modified Thompson Tau test [44]. For one-way ANOVA and Tukey t-test values, see Table S1 and Figs. S2, S3, S4, S5, S6, S7, S8, S9 and S10.
Results
Identification of gene families associated with UDP-glucose metabolism in sugarcane
A total of 136 genes from 32 gene families associated with the UDP-G metabolism, were identified within the STP sugarcane monoploid genome (see Fig. 1 for a depiction of the associated pathway). Most of these genes were also represented by 560 transcripts in the long-read SUGIT transcriptome. It must be noted that some of the transcripts represented in the SUGIT transcriptome were not represented in the STP genome, which accounted for 27 genes (Table 1). Comparisons to equivalent genes in the S. bicolor genome identified several likely gene duplications in ADP-glucose pyrophosphorylase (AGP), cellulose synthase-like (CSLE), cytosolic invertase (CINV), sucrose synthase (SuSy), and UDP-G-4-epimerase (GALE) gene families in the sugarcane genome, otherwise known as paralogues [45, 46]. Among the 136 genes, 10 gene homologues/isoforms belonged to the AGP gene family. Within the cellulose synthase (CESA) and cellulose synthase-like (CSL) subfamilies, 10 genes belonged to the CESA subgroup, and 5, 4, 4, 7, 2, 1 and 2 genes in CSL sub groups A, C, D, E, F and H, respectively. Of the four invertase sub-families, we identified 3 cytosolic invertase (CINV) genes, 5 alkaline/neutral invertase (ANINV), 1 vacuolar invertase (VINV) and 8 cell wall invertase (CWI) I. Note that CINV and ANINV are both known as “neutral invertases”, however, they have been annotated as different genes in the S. bicolor genome. Gene families associated with the myo-inositol pathway including glucuronokinase (GluK), myo-inositol phosphatase (MIP) and myo-inositol oxygenase (MIOX) each only had a single gene within the monoploid genome, whilst the MIPS had two gene family members. Gene families associated with hemicellulose synthesis had varying numbers of associated gene family members. A single gene was identified for uridine diphosphate glucose apiose/xylose synthase (UAXS), whilst multiple genes were identified in the UGD, GALE, UDP-xylose synthase (UXS), UDP-xylose epimerase (UXE), UDP-G 4,6-dehydratase (RHM) and uridine diphosphate glucuronic acid epimerase (UGE) gene families, with 3, 5, 6, 3, 3, 4 members, respectively. Single chloroplastic, cytoplasmic and 2 bi-functional phosphoglucomutase (PGM) genes were identified, while 5 UDP-G pyrophosphorylase (UGPase) genes, 3 glucose-6-phosphate isomerase (G6PI), 8 hexokinase (HXK) and 3 fructokinase (FK) genes were also identified. Some genes had large numbers of transcripts represented in the SUGIT transcriptome with MIPS 2, SuSy 1, SuSy 2, SuSy 4, UGD 4, UGD 5, UGPase 2, CesA 5, CSLE 6–2, FK 2, and GALE 1 all had over 15 representative transcripts each.
Gene-specific and gene family specific expression profile throughout the sugarcane plant
Sucrose to UDP-glucose and UDP-glucose/hexose phosphate to sucrose associated gene families
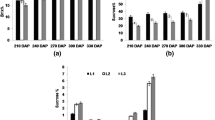
In the SuSy gene family, cumulative expression was significantly higher (p ≤ 0.05) in R and TI compared to other organ samples of both Q208 and KQ228 genotypes (Fig. 2a). The 4 gene isoforms of SuSy displayed differing degrees of expression throughout the sugarcane plant. SuSy 1, 2 and 7 had significantly higher expression in TI organs in comparison to leaf and mature internodal organs. No significant difference was observed between R and TI samples, however, differences between R and all other organs were not significant due to the high variance of triplicate values. SuSy 1 and 2 were the most prominent gene isoforms in terms of expression.

Cumulative and Isoform specific expression of gene families associated with the sucrose to UDP-Glucose, and UDP-Glucose to sucrose. Gene families: Sucrose Synthase (SuSy)in Figure a, Sucrose Phosphate Phosphatase (SPP) in Figure b, and Sucrose Phosphate Synthase (SPS) in Figure c. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was measured separately within each genotype
In the sucrose synthetic sucrose phosphate phosphatase (SPP) gene family, cumulative expression was higher in KQ228 L5 and Q208 L1 leaf samples (Fig. 2b). in comparison to the mature internodal samples, in both genotypes. The SPP 2 isoform had significantly higher expression values in roots in comparison to leaf and mature internodal samples. The SPP 1 isoform had heightened expression in L5 and L1 leaf samples in genotypes KQ228 and Q208, respectively. Among the two SPP isoforms, SPP 1 had the highest volume of expression.
Cumulative sucrose phosphate synthase (SPS) gene expression was not significantly different throughout the KQ228 genotype, whereas a significantly higher degree of expression was observed in the Q208 genotype (Fig. 2c). In the KQ228 genotype, SPS gene isoform-specific expression did not differ between SPS 2, 3, 4 and 5 genes, only SPS 1 displayed significantly higher expression in L5 in comparison to root and internodal organs. Distinctly different isoform expression patterns were observed in the Q208 genotype with SPS 2, 4 and 5 having significantly higher expression in TI sample in comparison to R, all leaf and BI samples. SPS 1 expression was significantly higher in L1 sample in comparison to R, BI and L5 samples. The expression of SPS 1 and 4 was the highest of all isoforms.
Cellulose synthase and cellulose synthase-like gene families
In general, comparing between two genotypes, cumulative expression of gene families related to cellulose biosynthesis including CesA, CSLA, CSLC and CSLD showed a significantly higher expression in TI and R samples compared to other samples used in this study (Fig. 3a, b, c and d, respectively). Cumulative expression of the CesA gene family was significantly higher in TI samples compared to other samples, while roots had significantly higher CesA expression than leaves. Isoform-specific expression for CesA 1–1, 1–2, 2, 3, 5, 7 and 8 was also significantly higher in TI sample, whereas, CesA 6 expression was significantly higher in root sample compared to other samples. The volume of CesA 1–1, 3, 6 and 8 expression was the highest of all isoforms.

Cumulative and Isoform specific expression of gene families associated with the cellulose synthase (CesA) and cellulose synthase-like (CSL) gene families. Gene families: Cellulose Synthase (CesA) in Figure a, and Cellulose Synthase-Like (CSL) in Figures b to h. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was measured separately within each genotype
Cumulative expression of the CSLA gene family was significantly higher in TI samples, followed by R sample which had significantly higher expression than mature internode and leaf samples. Among the isoforms, CSLA 1 and 7 expression was significantly higher in TI sample, and CSLA 1 expression was significantly higher in R sample, in comparison to leaf and mature internodal samples. CSLC cumulative expression was significantly higher in TI sample than all other samples in both genotypes, followed by R sample which was significantly higher than mature internodal and leaf samples. Isoform-specific expression displayed enhanced expression of CSLC 2 and 7 in TI samples, followed by R sample. In the KQ228 genotype, CSLC 9 expression was significantly higher in R sample, whereas in Q208 there was no significant difference between TI and R sample, both of which had significantly higher expression values than the other samples.
Cumulative expression in the CSLD gene family was significantly higher in R samples. Isoform-specific expression of CSLD 1 and 2 was significantly higher in root sample. CSLD 1 expression was significantly higher than CSLD 2 expression. The sum of CSLE expression was significantly higher in leaf samples (Fig. 3e). Isoform-specific expression of CSLE 6–1 was significantly higher in leaf samples, whilst CSLE 6–2 was significantly higher in leaf sample only in the Q208 genotype. CSLE 6–2 had the most pronounced expression of the CSLE isoforms. Cumulative expression of the CSLF gene family was significantly higher in R and TI samples (Fig. 3f). Expression of the CSLF 3 isoform was significantly higher in R samples, whilst expression for both CSLF 6 and 8 was significantly higher in R and TI samples. CSLF 6 had the most pronounced expression of the CSLF isoforms. Expression in the single CSLG gene was significantly higher in L1 sample in the Q208 genotype (Fig. 3g).
CSLG expression values tended to be higher in leaf samples, however due to the large variance in expression between replicates the differences were not significant, except L1 sample of Q208 genotype. Cumulative expression in the CSLH gene family did not differ between samples of the KQ228 genotype (Fig. 3h), however, in the Q208 genotype expression was significantly higher in leaf samples. Isoform-specific expression of CSLH 1–1 and 1–2 was significantly higher in leaf sample within the Q208 genotype. CSLH 1–2 expression was significantly higher in root sample in comparison to internodal samples. CSLH 1–1 had the most pronounced expression of the CSLH isoforms.
Hemicellulose synthesis associated gene families
The UDP-G consuming UGD gene family displayed significantly higher cumulative expression rates in R and TI samples (Fig. 4a). Within the two UGD isoforms, UGD 5 displayed significantly higher rates of expression in TI sample in comparison to all other samples. Root organ also had significant rates of UGD 5 expression, which were significantly higher than leaf and mature internodal organs. UGD 4 expression rates were significantly higher in R samples, closely followed by TI samples. Cumulative expression rates of the UGE gene family were significantly higher in R and TI samples across both genotypes. Isoform-specific expression of UGE 1–1 and UGE 6–2 was significantly higher in both R and TI samples (Fig. 4b). UGE 1–1 had the most pronounced expression of the UGE isoforms. Within the UXE gene family, cumulative expression was significantly higher in R and TI samples. Expression of the UXE 1 and 3 isoforms was significantly higher in TI samples (Fig. 4c), whereas UXE 2 had significantly higher expression in R samples, followed by TI sample. UXE 2 had the most pronounced expression of the UXE isoforms.

Cumulative and Isoform specific expression of gene families associated with hemicellulose synthesis. Gene families: Uridine Diphosphate Glucose Dehydrogenase (UGD) in Figure a, Uridine Diphosphate Glucuronic Acid Epimerase (UGE) in Figure b, Uridine Diphosphate Xylose Epimerase (UXE) in Figure c, Uridine Diphosphate Glucose 4,6-dehydratase (RHM) in Figure d, Uridine Diphosphate Xylose Synthase (UXS) in Figure e, Uridine Diphosphate Glucose 4-Epimerase (GALE) in Figure f, and Uridine Diphosphate Glucose Apiose/Xylose Synthase (UAXS) in Figure g. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was measured separately within each genotype
Cumulative expression of the RHM gene family was significantly higher in root samples, in comparison to all other samples (Fig. 4d). Isoform-specific expression of RHM 1–1 was significantly higher in root samples. Expression of RHM 1–2 was not significantly different between all samples in the KQ228 genotype, whereas in the Q208 genotype expression was significantly higher in R samples. The UXS gene family had significantly higher expression in TI samples, closely followed by R samples which had significantly higher expression than mature internode and leaf samples (Fig. 4e). Isoform-specific expression of the UXS gene family was as follows: UXS 4 had significantly higher expression in the R samples, whereas UXS 6 expression was significantly higher in TI samples. Expression rates of the other UXS isoforms tended to be higher in TI samples, although in some cases the differences were not significant between all samples. UXS 6 had the most pronounced expression of the UXS isoforms. Cumulative expression of the GALE gene family did not show a clear expression pattern between samples (Fig. 4f), with all leaf, R and TI samples having pronounced expression, although expressional differences were not significant between all samples. Isoform-specific expression of GALE 2 was significantly higher in TI sample, whereas GALE 1 expression tended to be higher in R and both leaf samples, although the differences between these samples and the internodal samples were not consistent across both genotypes. The single gene with UAXS functionality had significantly higher expression in TI samples (Fig. 4g), followed by R sample which had significantly higher expression than leaf and mature internodal samples.
Myo-inositol pathway associated genes
Figure 5 displays the cumulative and gene isoform-specific expression values of genes associated with the myo-Inositol pathway. Only single genes were identified at each point in the myo-Inositol pathway. MIPS gene expression displayed distinctly different expression patterns between the two genotypes with larger rates of expression being observed in the KQ228 genotype (Fig. 5a). Whilst no significant differences were observed between samples in the Q208 genotype, within the KQ228 genotype, TI sample had significantly higher expression than R and mature internodal samples. GluK and MIP gene expression were significantly higher in TI samples in both genotypes (Fig. 5b and c). MIOX gene expression was significantly higher in root samples (Fig. 5d).

Expression of genes associated with the myo-Inositol pathway. Gene families: myo-Inositol Phosphate Synthase (MIPS) in Figure a, Glucuronokinase (GluK) in Figure b, myo-Inositol Phosphatase (MIP) in Figure c, and myo-Inositol Oxygenase (MIOX) in Figure d. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was measured separately within each genotype
Invertase gene families
Figure 6 displays the cumulative and gene isoform-specific expression values of invertase associated gene families. Within the ANINV gene family, cumulative and isoform-specific expression did not display clear expression patterns between the sample types (Fig. 6a). Cumulative expression patterns of the CINV gene family were higher in R samples in both genotypes, although differences were only significant in the KQ228 genotype (Fig. 6b). Isoform-specific expression of CINV 1 and 2 was significantly higher in root sample in the KQ228 genotype, however, in the Q208 genotype these differences were not significant. Expression of the CINV 1 isoform was higher than that of CINV 2. Cumulative expression of the CWI gene family was significantly higher in R samples in the KQ228 genotype (Fig. 6c), whereas R and both leaf samples had tended to have higher expression in the Q208 genotype, although only L5 sample had significantly higher expression than internodal samples. CWI isoforms 1, 2, 5 and 7 had significantly higher expression in R across both genotypes. CWI 4 expression tended to be higher in leaf sample, with significantly higher expression being observed in the Q208 genotype. In the KQ228 genotype, only L5 sample had significantly higher expression than that of internodal samples. CWI isoforms 1 and 4 had the most pronounced expression of all CWI isoforms. VINV expression was significantly higher in roots in both genotypes (Fig. 6d).

Cumulative and Isoform specific expression of gene families associated with Invertase activity. Gene families: Alkaline/Neutral Invertase (ANINV) in Figure a, Cytosolic Invertase (CINV) in Figure b, Cell Wall Invertase (CWI) in Figure c, and Vacuolar Invertase (VINV) in Figure d. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was measured separately within each genotype
Phosphorylated sugar synthesis /degradation associated gene families
Figure 7 displays the cumulative and gene isoform-specific expression values of genes associated with the synthesis and degradation of phosphorylated sugars. Cumulative UGPase expression was significantly higher in TI samples in comparison to leaf and mature internodal samples in the KQ228 genotype (Fig. 7a). In the Q208 genotype whilst expression was observed to be higher in TI samples, this result was not significant. Isoform-specific expression of the UGPase gene family did not display any conclusive evidence of significant differences between samples in both genotypes. The UGPase 2 isoform had pronounced expression in comparison to the 3 other UGPase isoforms. Cumulative AGP expression tended to be higher in leaf samples, closely followed by TI sample, however, this trend was only significant in the KQ228 genotype (Fig. 7b). Isoform-specific expression of AGP 4 and 6 was significantly higher in leaf samples. Cumulative expression of the PGM gene family did not display any consistent expressional trends between samples in both genotypes (Fig. 7c). Isoforms AGP 1, 2 and 6 had far more expression than other AGP isoforms. The chloroplastic PGM 1 isoform had significantly higher expression in leaf sample, whilst bi-functional PGM 1 and cytoplasmic PGM 2 had significantly higher expression in TI sample. A significant amount of bi-functional PGM 2 expression was observed in both L1 and L5 samples of the Q208 genotype, however, this trend was not observed in the KQ228 genotype.

Cumulative and Isoform specific expression of gene families associated with Phosphorylated sugar synthesis and degradation. Gene families: Uridine Diphosphate Glucose Pyrophosphorylase (UGPase) in Figure a, Adenosine Diphosphate Glucose Pyrophosphorylase (AGP) in Figure b, Phosphoglucomutase (PGM) in Figure c, Glucose-6-Phosphate Isomerase (G6PI) in Figure d, Hexokinase (HXK) in Figure e and Fructokinase (FK) in Figure f. Abbreviations, TI: Top Internode; MI: Middle Internode; BI: Bottom Internode; 1st Visible Dewlap Leaf: L1; 5th Visible Dewlap Leaf: L5; R: Root. The bar chart displays the cumulative expression of all genes within a gene family. Letters above each bar indicate the presence of a significant difference between values within the same genotype. Error bars +/− 1 S.D. from biological triplicates. The heat map plots the Log2 TPM values for each individual gene family member. Green displays higher expression values, yellow for mid-range values and red for lower range expression values. Letters within the heatmap indicate the presence of a significant difference between values within an individual gene, within the same genotype. Significance was calculated via one way ANOVA, with the post-hoc Tukey’s T-test to separate statistically dissimilar groups. Statistical analysis was easured separately within each genotype
G6PI cumulative and isoform-specific expression did not display any consistently significant expressional trends between samples across both genotypes (Fig. 7d). Cumulative expression of the FK gene family was significantly higher in TI and R samples across both genotypes (Fig. 7f). Expression of the FK 1 isoform was significantly higher in R sample, whereas expression for FK 2 and 6 was significantly higher in TI sample. FK 1 and 2 had pronounced expressional values in comparison to the FK 3 isoform. Cumulative expression in the HXK gene family was significantly higher in TI and R samples (Fig. 7e). Expression of HXK 6 and 8 isoforms was significantly higher in TI samples across both genotypes. There was a significantly higher expression of HXK 7 isoform in R samples. HXK 3 expression was significantly higher in both R and TI samples.
Discussion
The importance of UDP-G control and its effect on C partitioning has been indicated in several studies of plant systems, whereby the kinetic properties and abundance of enzymes involved in UDP-G synthesis/utilisation had a significant effect upon the volume of C moving into the hemicellulose, cellulose and sucrose pools [13,14,15,16,17,18,19]. UDP-G metabolism is central to key sources of C deposition in the sugarcane plant, therefore is likely that the expression of genes involved in UDP-G metabolism would have differed expression in relation to how C is utilised in a given tissue/organ. Additionally, most of these genes are known to be in multi-gene families, and many of these multi-gene families likely contain isoforms that overall have a larger effect on metabolism than others. By determining tissue-specific isoforms, gene candidates for altering UDP-G metabolism and by proxy C metabolism could be identified, in order to define the strategies to modify plant biomass. Here, to provide a broad overview of differences in organ-specific expression of genes associated with the UDP-G metabolism, we analysed their expression in 3 major organs of the sugarcane plant including leaves (mature source organ), internodes (young and mature sink organ) and root (meristematic sink organ).
Expression of genes associated with the UDP-G metabolism in internodes
Sucrose synthesis and degradation in internodal samples by the SPP, SPS and SuSy enzymes (Fig. 2), is closely linked to sugarcane internodal maturity, whereby sucrose cleavage and hydrolysis prevails in immature internodes and sucrose synthesis and lack of sucrose utilization into respiration and insoluble components prevails in mature internodes [47]. As indicated in this study and previous studies of SuSy genes, expression and enzymatic activity are significantly higher in immature internodes [47, 48], likely providing C for hemicellulose and cellulose synthesis from UDP-G. Interestingly, within the SuSy gene family, the highest expression was limited to 2 of the 4 SuSy genes (SuSy 1 and 2) indicating that they may code for enzymes that have heightened importance in the cleavage of sucrose into UDP-G (Fig. 2a), as has been reported in Arabidopsis and Sugarcane [30, 48, 49]. As expected, enhanced expression of SuSy in immature internodes, also coincided with higher expression in cell wall synthetic related gene families, including CesA, CSL (A, C, D and F families, see Fig. 3), GALE, UAXS, UGE, UXE and UGD (Fig. 4). Some of the aforementioned genes code for enzymes that directly consume UDP-G, including UGD, CesA and some CSL gene families [50,51,52], in essence having direct competition with sucrose synthetic enzymes. The key genes coding for enzymes that are responsible for the bulk of UDP-G into cellulose and hemicellulose pools are likely coded for by UGD 5 (Fig. 4a), and CesA subunit genes 1–1, 1–2, 3, 5, 7 and 8 (Fig. 2a). In a related study in the UGD gene family in Arabidopsis, an enzyme isoform with high affinity for UDP-Glucose, also had high affinity for the downstream product UDP-xylose, which acts as a feedback inhibitor [21], The associated genes also had higher expression, suggesting they code for enzymes that hold a primary role in the assimilation of C into the hemicellulose pool in immature tissues. Other UGD gene isoforms with lower volumes of expression likely code for enzymes with low substrate affinity and operate in the background throughout the sugarcane plants lifecycle. This concept could credibly be applied to most gene families in this study.
In conjunction with the heightened sucrose cleavage to UDP-G, cumulative invertase expression in CINV and CWI gene families was significantly higher in immature internodes in comparison to mature internode samples (Fig. 4b and c, respectively). This was expected as sucrose hydrolysis into reducing sugars characterizes the first step of C movement into several pathways, leading into cell wall, protein, respiratory and other secondary metabolite pools [53]. The heightened levels of reducing sugars in immature internodes [32], indicates the activity of invertases hydrolysing imported sucrose. The heightened expression of the CWI 1 and 7 genes in the immature internodes (Fig. 4c), suggests prominent activity of C fixation into parenchyma tissue from conducting tissues, and the apoplastic/symplastic transfer of sucrose [54, 55]. Lower expression of CWI in mature internodes may suggest that C importation facilitated by the activity of invertases is less pronounced. The heightened expression of genes encoding phosphorylating enzymes, FK and HXK in immature internodes (Fig. 7e and f), particularly genes HX 3 and 5 and FK 2, further supports the notion of high C movement likely toward pentose phosphate pathway and glycolysis, as reviewed by [56]. The indication of enhanced C flux into these pathways that may be mediated by gene expression is important, as results from [39] suggested very little difference in expression of transcripts associated with these pathways, which may suggest associated enzymes operate during favourable metabolic conditions, i.e. when metabolites are available. This indicates the potential importance of invertases and phosphorylating enzymes in releasing C to the pentose phosphate and glycolytic pathways.
Unexpectedly, in the sucrose synthetic gene families, SPS and SPP, between the mature and immature internodes, expression was higher in immature internodes (Fig. 2b and c), which is in contradiction to several studies that have found an opposite trend in enzymatic activity of SPS [47, 57]. It must be noted that enzymatic activity and gene expression are not necessarily correlated [58]. Further, another study has found similar results, whereby enzymatic activity was higher in immature internodes. The results from this study may indicate that the absence of competition for C in mature internodes by sucrose degradative and downstream enzymes, is the key to sucrose accumulation in mature internodes, as has been indicated in several studies [47, 59]. In a previous study by Botha and Black (2000), it was intimated that there could be an additional kinetic form of SPS that allows for heightened enzymatic activity, which explains the heightened activity of SPS in mature internodes [47], however, there is no evidence of increased transcription in any of the 5 SPS isoforms in mature internodes in our study. These results suggest enzymatic activity of SPS is determined beyond transcription.
Expression of genes associated with the UDP-G metabolism in roots
Root and immature internodes are both meristematic sinks, moving a large proportion of fixed C into the cell wall, protein and respiratory pools. Despite this, there were large expressional differences between root and immature internode samples likely indicating spatial regulation of specific gene family isoforms. Some of the transcriptional differences between the two meristematic sinks is likely a result of the difference in age the samples were taken from the sugarcane plant (roots from a 3-month-old plant and immature internodes from a 9-month-old plant). However, previous compositional analyses of sugarcane roots display a differing requirement for fixed C than immature internodes [32, 60]. Compositional analyses of 3-month old sugarcane roots indicated differences in simple sugar and hemicellulosic monomer content in comparison to immature internodes [32]. Also, in immature internodes, there is an underlying trend for the accumulation of sucrose that does not exist in roots [60]. Concerning the simple sugar content, i.e. fructose, glucose and sucrose, it was postulated that the reduced levels in the root sample indicate the efficient breakdown and movement of fixed C into the cell wall, protein, organic acid and respiratory pools in roots. This is made clear by the high expression of VINV in roots (Fig. 4d), which has been implicated as a key enzyme negatively affecting the accumulation of sucrose in several plant systems, as reviewed by [61]; and the heightened expression of SPS 1 and 4 genes (only significant in Q208 genotype, see Fig. 2c). In this study, low VINV expression in all internodes and leaf samples, indicating low VINV activity (Fig. 4d), likely allows the accumulation of sucrose for storage or transportation. Higher observed expression of VINV in roots suggests a lower requirement for sucrose bioaccumulation. Additionally, the high expression of two other invertase families in roots in comparison to immature internodes, including, CINV and CWI may also likely indicate a low inclination for sucrose bioaccumulation in roots (Fig. 4b and c). The higher expression of CWI isoforms 1, 2, 5 and 7 in root samples in comparison to immature internodes suggest a heightened role for CWI in roots, may enable an increase in hydrolysis of apoplastic sucrose, which in turn ensures a steep concentration gradient enhancing sucrose delivery to roots from mature leaves [62]. Interestingly, in sugarcane, the activity of CWI in internodes is correlated with higher sucrose levels [63, 64]. Enhanced invertase activity is counterintuitive to enhanced sucrose levels, however, the sucrose cleavage and resynthesis model as proposed by Glasziou and Gayler [65], may explain this. Higher expression of CWI genes in roots suggests the intercellular sucrose cleavage and intracellular resynthesis model to not be relevant in the root sample, as there is no evidence for large degrees of sucrose resynthesis to be occurring [32]. This notion is further supported by the lower expression of SPS observed in roots (only significantly different in Q208 genotype, Fig. 2c) in comparison to immature internodes and leaf samples.
Corresponding expression of SuSy genes between immature internodes and root samples likely indicates a high degree of UDP-G formulation from sucrose (Fig. 2a), followed by C utilisation into cellulose and hemicellulose pools. The expression of genes associated with UDP-G into cell wall polysaccharides differed greatly between immature internodes and root samples. Heightened expression of an additional UGD gene family isoform (UGD 4), and the MIOX gene suggests there may be enhanced enzymatic activity indicating the strong demand of C to be moved into the hemicellulose fraction in roots or an organ specific function (Figs. 4a and 5d, respectively). Evidence of heightened expression in a specific UGD isoform has been reported in Arabidopsis seedlings [21]. Additionally, downstream steps of hemicellulose synthesis displayed a significant difference between these two samples, which could be responsible for the heightened amount of arabinose and galactose mixed linkages in roots [32], particularly GALE 1 and UXE 2 (Fig. 4f and c). Although, as shown in related analyses, there was also higher expression in other hemicellulose related transcripts in roots, that do not result in differences in composition [32, 39]. Interestingly, CesA expression was significantly higher in immature internodes (Fig. 3a). The significant differences in gene expression related to hemicellulose and cellulose synthesis could be related to the presence of specialised cells in both roots and internodes [66,67,68], having different requirements for C, or differing metabolic conditions, i.e. access to substrates or the presence of feedback inhibitory molecules [21]. Notably, compositional analysis of roots and internodes, as presented in [32], displayed no difference in the ratio of hemicellulose and cellulose, indicating differences in expression of related genes may not affect the fixed nature of the cell wall component ratios.
Expression of genes associated with the UDP-G metabolism in leaves
The expression of genes related to the UDP-G metabolism in sugarcane leaves is connected to the status of this organ as a net exporter of C in the form of sucrose. As expected, cell wall-related genes that directly synthesize or consume UDP-G, including SuSy, CesA, CSL and UGD gene families, had insignificant amounts of expression in the leaf samples, indicating a transcriptional regulation as a means of ceasing C flow into the cell wall pool in this organ. Interestingly, some CSL gene family groups (Fig. 3) had significant expression in leaves including CSLE, CSLG and CSLH, specifically CSL6–2, CSLG2 and CSLH1–1 genes (Fig. 3e, g and h, respectively). CSL enzymes are responsible for the transfer of UDP-G to 1–3 and 1–4 β-glucan, or the transfer of other nucleotide sugars to form other β-linked backbones, within the hemicellulose fraction. Most CSL genes had significantly higher expression in both meristematic/immature sink samples, which was expected due to the requirement for hemicellulose synthesis. It is unclear why there was significant expression of some CSL gene family groups in leaf samples. Of the CSL gene groups with significant expression in leaves, CSLE and CSLG have an unknown function (Fig. 3e and g), as reviewed by [69], although it is likely still associated with hemicellulose synthesis, whereas the CSLH group (Fig. 3h) encodes mixed linkage glucan synthases [70]. These CSL groups may have a heightened requirement in leaf sample during development constructing leaf specific structures, with different arrangements of cell wall compounds. The data would also suggest that expression of these genes are retained throughout maturity exclusively in leaf samples, and may also have a role in maintenance.
Unlike most cell wall-related gene expression, high expression of sucrose synthetic genes SPS and SPP (Fig. 2b and c), equivalent to both meristematic samples were observed in leaf sample. Expression of SPP 1 and SPS 1 was most prominent in the leaf (although not significantly different from other samples), which may suggest these specific genes have an enhanced role in leaf sample. This trend has been hypothesised to be due to pronounced role of SPS and SPP in the synthesis of sucrose in source tissues [71, 72]. Relevant to sucrose biosynthesis in leaves is the production of UDP-G which is likely primarily derived from the activity of UGPase which transfers glucose-1-phosphate to UDP-G, whereas in sink samples UDP-G synthesis is derived primarily from sucrose cleavage by SuSy [73]. In support of this notion, transcription of SuSy related genes (Fig. 2a) was significantly higher in meristematic sinks. However, despite the primary the role of UGPase in sucrose biosynthesis in leaves, there was not higher expression in related gene family isoforms (Fig. 7a). This suggests the pronounced role of UGPase in source samples is not determined at the transcriptional level, but at the metabolic level, likely via the availability of hexose phosphates [74]. As indicated by the low expression of SuSy and high expression of SPS and SPP in leaves (Fig. 2c and b), this suggests a bias toward sucrose synthesis in this source organ, which will then be transported to various sink organs. However, the high expression of some invertase genes in the CWI (CWI 4), and ANINV (ANINV 1–1 and 3) gene families suggest sucrose hydrolysis to be a major competing sink for C (Fig. 6c and a, respectively). In a previous experiment of photosynthetic regulation by sugars in sugarcane leaves, fed radiolabelled sucrose was rapidly converted into hexoses, which was stipulated to be due to the activity of SuSy and invertase enzymes [75]. However, based on the low expression of SuSy genes in source organs in this study, the rapid conversion of sucrose may be mostly derived from invertase activity in leaves. The heightened activity of some invertases in sugarcane leaves suggests a major role in the regulation of sugar levels, especially due to the inhibitory nature of high sucrose concentrations on photosynthetic activity [76, 77]. In the case of the CWI gene family, most expression in leaves was contributed by the CWI 4 gene (Fig. 6c), which was also significantly higher than other organ types, which suggests an organ-specific function for this gene, potentially as a regulator of sucrose levels. It must be noted that as leaf organ is highly metabolically active and is a protein-rich organ [32], there is likely still an underlying requirement for C to be moved into respiratory and protein fractions. It is possible that invertase activity is contributing C to these pools.
Conclusions
This study represents the first effort to quantify the expression of gene families associated with UDP-G metabolism in a sugarcane plants. The data presented provides a quality reference for future efforts in altering UDP-G metabolism and in turn carbon partitioning in sugarcane. Transcriptional analysis displayed the likelihood that carbon partitioning in sugarcane is closely related to the transcription of genes associated with the UDP-G metabolism in the four major modes of carbon partitioning as defined in leaves (source), roots (non-storage immature/meristematic sink), mature internodes (storage sink) and immature internodes (immature sink that will become a storage sink). The data presented may provide an accurate genetic reference for future efforts in altering UDP-G metabolism and in turn C partitioning in sugarcane.
Availability of data and materials
The version of the SUGIT database used in this study can be accessed in Figshare under the https://doi.org/10.6084/m9.figshare.4981655. The RNA-seq data used for expression profiling has been deposited in the NCBI Sequence Read Archive (SRA) under the BioProject ID PRJNA479814, Study Accession Number SRP152893.
Abbreviations
- C:
-
Carbon
- NGS:
-
Next Generation Sequencing
- mRNA:
-
Messenger RNA
- cDNA:
-
Complementary DNA
- BI:
-
Bottom Internode
- MI:
-
Middle Internode
- TI:
-
Top Internode
- L1:
-
1st visible dewlap leaf
- L5:
-
5th visible dewlap leaf
- R:
-
Root
- UDP-G:
-
Uridine Diphosphate Glucose
- CWI:
-
Cell Wall Invertase
- VINV:
-
Vacuolar Invertase
- CINV:
-
Cytosolic Invertase
- ANINV:
-
Alkaline/Neutral Invertase
- FK:
-
Fructokinase
- MIPS:
-
myo-Inositol phosphate synthase
- MIP:
-
myo-Inositol phosphatase
- MIOX:
-
myo-Inositol oxygenase
- GluK:
-
Glucuronokinase
- PGM:
-
Phosphoglucomutase
- UGPase:
-
Uridine Diphosphate Glucose pyrophosphorylase
- UGD:
-
UDP-Glucose Dehydrogenase
- AGP:
-
Adenosine Diphosphate Glucose pyrophosphorylase
- CesA:
-
Cellulose Synthase
- CSL:
-
Cellulose Synthase-Like
- G6PI:
-
Glucose-6-Phosphate Isomerase
- SPP:
-
Sucrose Phosphate Phosphatase
- SPS:
-
Sucrose Phosphate Synthase
- SuSy:
-
Sucrose Synthase
- RHM:
-
Uridine Diphosphate Glucose 4,6-dehydratase
- UAXS:
-
Uridine Diphosphate Glucose Apiose/Xylose Synthase
- GALE:
-
Uridine Diphosphate Glucose 4-Epimerase
- UGE:
-
Uridine Diphosphate Glucuronic Acid Epimerase
- UXS:
-
Uridine Diphosphate Xylose Synthase
- UXE:
-
Uridine Diphosphate Xylose Epimerase
- HXK:
-
Hexokinase
References
Feingold DS, Barber GA. Nucleotide sugars. InMethods in plant biochemistry. Academic Press. 1990;2:39-78.
Gibeaut DM. Nucleotide sugars and glycosyltransferases for synthesis of cell wall matrix polysaccharides. Plant Physiol Biochem. 2000;38(1):69–80.
Richmond TA, Somerville CR. The cellulose synthase superfamily. Plant Physiol. 2000;124(2):495–8.
Kärkönen A, et al. UDP-glucose dehydrogenases of maize: a role in cell wall pentose biosynthesis. Biochem J. 2005;391(2):409–15.
Amor Y, et al. A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc Natl Acad Sci. 1995;92(20):9353–7.
Fung RW, et al. Differential expression within an SPS gene family. Plant Sci. 2003;164(4):459–70.
Kleczkowski LA, Kunz S, Wilczynska M. Mechanisms of UDP-glucose synthesis in plants. Crit Rev Plant Sci. 2010;29(4):191–203.
Wai CM, et al. Cell wall metabolism and hexose allocation contribute to biomass accumulation in high yielding extreme segregants of a Saccharum interspecific F2 population. BMC Genomics. 2017;18(1):773.
Kleczkowski LA. Glucose activation and metabolism through UDP-glucose pyrophosphorylase in plants. Phytochemistry. 1994;37(6):1507–15.
Chivasa S, Tomé DF, Slabas AR. UDP-glucose pyrophosphorylase is a novel plant cell death regulator. J Proteome Res. 2013;12(4):1743–53.
Wang Z, et al. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defence responses in rice. J Exp Bot. 2014;66(3):973–87.
Xiao G, et al. Excessive UDPG resulting from the mutation of UAP1 causes programmed cell death by triggering reactive oxygen species accumulation and caspase-like activity in rice. New Phytol. 2018;217(1):332–43.
Samac DA, et al. Expression of UDP-glucose dehydrogenase reduces cell-wall polysaccharide concentration and increases xylose content in alfalfa stems. Appl Biochem Biotechnol. 2004;116(1–3):1167–82.
Reboul R, et al. Down-regulation of UDP-glucuronic acid biosynthesis leads to swollen plant cell walls and severe developmental defects associated with changes in pectic polysaccharides. J Biol Chem. 2011;286(46):39982–92.
Bahaji A, et al. Characterization of multiple SPS knockout mutants reveals redundant functions of the four Arabidopsis sucrose phosphate synthase isoforms in plant viability, and strongly indicates that enhanced respiration and accelerated starch turnover can alleviate the blockage of sucrose biosynthesis. Plant Sci. 2015;238:135–47.
Kumar M, et al. Exploiting cellulose synthase (CESA) class specificity to probe cellulose microfibril biosynthesis. Plant Physiol. 2018;177(1):151–67.
Chu Z, et al. Knockout of the AtCESA2 gene affects microtubule orientation and causes abnormal cell expansion in Arabidopsis. Plant Physiol. 2007;143(1):213–24.
Harris D, Stork J, Debolt S. Genetic modification in cellulose-synthase reduces crystallinity and improves biochemical conversion to fermentable sugar. GCB Bioenergy. 2009;1(1):51–61.
Angeles-Núnez JG, Tiessen A. Arabidopsis sucrose synthase 2 and 3 modulate metabolic homeostasis and direct carbon towards starch synthesis in developing seeds. Planta. 2010;232(3):701–18.
Henikoff S, et al. Gene families: the taxonomy of protein paralogs and chimeras. Science. 1997;278(5338):609–14.
Klinghammer M, Tenhaken R. Genome-wide analysis of the UDP-glucose dehydrogenase gene family in Arabidopsis, a key enzyme for matrix polysaccharides in cell walls. J Exp Bot. 2007;58(13):3609–21.
Kanter U, et al. The inositol oxygenase gene family of Arabidopsis is involved in the biosynthesis of nucleotide sugar precursors for cell-wall matrix polysaccharides. Planta. 2005;221(2):243–54.
Bekker JP. Genetic manipulation of the Cell Wall composition of sugarcane. Stellenbosch: University of Stellenbosch; 2007. p. 66.
Chourey P, et al. Genetic evidence that the two isozymes of sucrose synthase present in developing maize endosperm are critical, one for cell wall integrity and the other for starch biosynthesis. Mol Gen Genet MGG. 1998;259(1):88–96.
Rösti J, et al. UDP-glucose 4-epimerase isoforms UGE2 and UGE4 cooperate in providing UDP-galactose for cell wall biosynthesis and growth of Arabidopsis thaliana. Plant Cell. 2007;19(5):1565–79.
Craig J, et al. Mutations at the rug4 locus alter the carbon and nitrogen metabolism of pea plants through an effect on sucrose synthase. Plant J. 1999;17(4):353–62.
Gordon AJ, et al. Sucrose synthase in legume nodules is essential for nitrogen fixation. Plant Physiol. 1999;120(3):867–78.
Ruan Y-L, Llewellyn DJ, Furbank RT. Suppression of sucrose synthase gene expression represses cotton fiber cell initiation, elongation, and seed development. Plant Cell. 2003;15(4):952–64.
Park J-Y, et al. Over-expression of an arabidopsis family a sucrose phosphate synthase (SPS) gene alters plant growth and fibre development. Transgenic Res. 2008;17(2):181–92.
Bieniawska Z, et al. Analysis of the sucrose synthase gene family in Arabidopsis. Plant J. 2007;49(5):810–28.
Schuster SC. Next-generation sequencing transforms today's biology. Nat Methods. 2008;5(1):16–8.
Mason PJ, et al. Variation in sugarcane biomass composition and enzymatic saccharification of leaves, internodes and roots. Biotechnol Biofuels. 2020;13(1):1–19.
SRA, QCANESelect. 2016, Sugar Research Australia: https://tools.sugarresearch.com.au/QCANESelect/Default.aspx.
Furtado A. RNA extraction from developing or mature wheat seeds. Cereal Genomics. 2014;1:23–8.
Andrews S. FastQC: a quality control tool for high throughput sequence data; 2010.
Hoang NV, et al. A survey of the complex transcriptome from the highly polyploid sugarcane genome using full-length isoform sequencing and de novo assembly from short read sequencing. BMC Genomics. 2017;18(1):395.
QIAGEN. Expression Analysis using RNA-seq, in Tutorial. Aarhus: QIAGEN Bioinformatics; 2017.
UQ. Research Computing Centre (RCC). St Lucia: University of Queensland; 2019.
Mason PJ, et al. Comparison of the root, leaf and internode transcriptomes in sugarcane (Saccharum spp. hybrids). Curr Res Biotechnol. 2022;4:167–78.
Johnson M, et al. NCBI BLAST: a better web interface. Nucleic Acids Res. 2008;36(suppl_2):W5–9.
Garsmeur O, et al. A mosaic monoploid reference sequence for the highly complex genome of sugarcane. Nat Commun. 2018;9(1):2638.
Consortium, U. UniProt: a worldwide hub of protein knowledge. Nucleic Acids Res. 2019;47(D1):D506–15.
Zhang C, et al. Evaluation and comparison of computational tools for RNA-seq isoform quantification. BMC Genomics. 2017;18(1):583.
Garcia FAA. Tests to identify outliers in data series. Rio de Janeiro: Pontifical Catholic University of Rio de Janeiro, Industrial Engineering Department; 2012.
Altenhoff AM, et al. Standardized benchmarking in the quest for orthologs. Nat Methods. 2016;13(5):425.
Fitch WM. Distinguishing homologous from analogous proteins. Syst Zool. 1970;19(2):99–113.
Botha FC, Black KG. Sucrose phosphate synthase and sucrose synthase activity during maturation of internodal tissue in sugarcane. Funct Plant Biol. 2000;27(1):81–5.
Thirugnanasambandam P, et al. Analysis of the diversity and tissue specificity of sucrose synthase genes in the long read transcriptome of sugarcane. BMC Plant Biol. 2019;19(1):1–4.
Noman M, et al. In Silico dissection and expression analysis of sucrose synthase gene family in sugarcane. Sugar Tech. 2022;24:1–12.
Kaur S, et al. Genome-wide analysis of the cellulose synthase-like (Csl) gene family in bread wheat (Triticum aestivum L.). BMC Plant Biol. 2017;17(1):193.
Turner W, Botha FC. Purification and kinetic properties of UDP-glucose dehydrogenase from sugarcane. Arch Biochem Biophys. 2002;407(2):209–16.
Taylor NG, Laurie S, Turner SR. Multiple cellulose synthase catalytic subunits are required for cellulose synthesis in Arabidopsis. Plant Cell. 2000;12(12):2529–39.
Koch K. Sucrose metabolism: regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr Opin Plant Biol. 2004;7(3):235–46.
Hatch M, Sacher J, Glasziou K. Sugar accumulation cycle in sugar cane. I. Studies on enzymes of the cycle. Plant Physiol. 1963;38(3):338.
Gayler K, Glasziou K. Physiological functions of acid and neutral invertases in growth and sugar storage in sugar cane. Physiol Plant. 1972;27(1):25–31.
Turner JF, Turner DH. The regulation of glycolysis and the pentose phosphate pathway. InMetabolism and Respiration. Academic Press. 1980:279-316.
Verma AK, et al. Functional analysis of sucrose phosphate synthase (SPS) and sucrose synthase (SS) in sugarcane (Saccharum) cultivars. Plant Biol. 2011;13(2):325–32.
Glanemann C, et al. Disparity between changes in mRNA abundance and enzyme activity in Corynebacterium glutamicum: implications for DNA microarray analysis. Appl Microbiol Biotechnol. 2003;61(1):61–8.
Batta SK, Singh R. Sucrose metabolism in sugar cane grown under varying climatic conditions: synthesis and storage of sucrose in relation to the activities of sucrose synthase, sucrose-phosphate synthase and invertase. Phytochemistry. 1986;25(11):2431–7.
De Souza AP, et al. Diurnal variation in gas exchange and nonstructural carbohydrates throughout sugarcane development. Funct Plant Biol. 2018;45(8):865–76.
Huber SC. Biochemical mechanism for regulation of sucrose accumulation in leaves during photosynthesis. Plant Physiol. 1989;91(2):656–62.
Roitsch T, et al. Extracellular invertase: key metabolic enzyme and PR protein. J Exp Bot. 2003;54(382):513–24.
Botha FC, Whittaker A, Vorster DJ, Black KG. Sucrose accumulation rate, carbon partitioning and expression of key enzyme activities in sugarcane stem tissue. In: Wilson DMHJR, Campbell JA, Garside AL, editors. Sugarcane: Research Towards Efficient and Sustainable Production: CSIRO Division of Tropical Crops and Pastures; 1996. p. 98–101.
Lingle S. Effect of transient temperature change on sucrose metabolism in sugarcane internodes. J Am Soc Sugar Cane Technol. 2004;24:132–40.
Glasziou K, Gayler K. Storage of sugars in stalks of sugar cane. Bot Rev. 1972;38(4):471–90.
Steffens B, Rasmussen A. The physiology of adventitious roots. Plant Physiol. 2016;170(2):603–17.
Moore PH, Botha FC. Sugarcane: physiology, biochemistry and functional biology. Oxford: Wiley; 2013.
Walsh KB, Sky RC, Brown SM. The anatomy of the pathway of sucrose unloading within the sugarcane stalk. Funct Plant Biol. 2005;32(4):367–74.
Yin Y, Huang J, Xu Y. The cellulose synthase superfamily in fully sequenced plants and algae. BMC Plant Biol. 2009;9(1):99.
Doblin MS, et al. A barley cellulose synthase-like CSLH gene mediates (1, 3; 1, 4)-β-D-glucan synthesis in transgenic Arabidopsis. Proc Natl Acad Sci. 2009;106(14):5996–6001.
Ma P, et al. Comparative analysis of sucrose phosphate synthase (SPS) gene family between Saccharum officinarum and Saccharum spontaneum. BMC Plant Biol. 2020;20(1):1–15.
Lunn JE. Sucrose-phosphatase gene families in plants. Gene. 2003;303:187–96.
Kotake T, et al. UDP-sugar pyrophosphorylase with broad substrate specificity toward various monosaccharide 1-phosphates from pea sprouts. J Biol Chem. 2004;279(44):45728–36.
Appeldoorn NJ, et al. Developmental changes of enzymes involved in conversion of sucrose to hexose-phosphate during early tuberisation of potato. Planta. 1997;202(2):220–6.
McCormick AJ, Cramer MD, Watt DA. Regulation of photosynthesis by sugars in sugarcane leaves. J Plant Physiol. 2008;165(17):1817–29.
Ribeiro RV, et al. Increased sink strength offsets the inhibitory effect of sucrose on sugarcane photosynthesis. J Plant Physiol. 2017;208:61–9.
McCormick A, Cramer M, Watt D. Sink strength regulates photosynthesis in sugarcane. New Phytol. 2006;171(4):759–70.
Acknowledgements
We thank the staff at the SRA Burdekin branch for their assistance in preparing and sampling the plant material. Research was supported by Sugar Research Australia (SRA), providing access to plant material. Also, the high-performance computing resources required for in silico analysis was provided by the Research Computing Centre (RCC), University of Queensland.
Permission to collect plant material
All collection of plant material for this study was undertaken at Sugar Research Australia site in the Burdekin, QLD, under the supervision and with permission of Sugar Research Australia.
Funding
This research was supported by a Sugar Industry Postgraduate Research Scholarship (SPRS) from Sugar Research Australia to P.J.M.
Author information
Authors and Affiliations
Contributions
Patrick J Mason: Conceptualisation, Methodology, Investigation, Data Curation, Writing-Original Draft, Writing-Review and Editing, Visualisation Nam V Hoang: Conceptualisation, Methodology, Investigation, Data Curation, Writing-Review and Editing, Visualisation Frederick C Botha: Conceptualisation, Methodology, Writing-Review and Editing, Supervision, Resources Agnelo Furtado: Conceptualisation, Methodology, Investigation, Writing-Review and Editing, Supervision Annelie Marquardt: Methodology, Investigation, Writing-Review and Editing Robert J Henry: Conceptualisation, Methodology, Writing-Review and Editing, Supervision, Resources. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
None declared.
Consent for publication
None declared.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Variety factsheet for Q208 and KQ228 from QCANESelect™ variety selection webpage. Fig. S1. Schematic of sugarcane transcript gene family homologue assignment and quantification. Table S2. Numerical Identifier for Tukey t-test results and one-way ANOVA significance values. Fig. S2. One-way ANOVA and Tukey t-test results from SuSy gene family expression comparisons. Fig. S3. One-way ANOVA and Tukey t-test results from SPS gene family expression comparisons. Fig. S4. One-way ANOVA and Tukey t-test results from SPP gene family expression comparisons. Fig. S5. One-way ANOVA and Tukey t-test results from CSLA gene family expression comparisons. Fig. S6. One-way ANOVA and Tukey t-test results from CesA gene family expression comparisons. Fig. S7. One-way ANOVA and Tukey t-test results from CSLC gene family expression comparisons. Fig. S8. One-way ANOVA and Tukey t-test results from CSLD gene family expression comparisons. Fig. S9. One-way ANOVA and Tukey t-test results from CSLE gene family expression comparisons. Fig. S10. One-way ANOVA and Tukey t-test results from CSLF gene family expression comparisons. Fig. S11. One-way ANOVA and Tukey t-test results from CSLG gene family expression comparisons. Fig. S12. One-way ANOVA and Tukey t-test results from CSLH gene family expression comparisons. Fig. S13. One-way ANOVA and Tukey t-test results from UGD gene family expression comparisons. Fig. S14. One-way ANOVA and Tukey t-test results from UGE gene family expression comparisons. Fig. S15. One-way ANOVA and Tukey t-test results from UXE gene family expression comparisons. Fig. S16. One-way ANOVA and Tukey t-test results from RHM gene family expression comparisons. Fig. S17. One-way ANOVA and Tukey t-test results from GALE gene family expression comparisons. Fig. S18. One-way ANOVA and Tukey t-test results from UXS gene family expression comparisons. Fig. S19. One-way ANOVA and Tukey t-test results from UAXS gene family expression comparisons. Fig. S20. One-way ANOVA and Tukey t-test results from MIPS gene family expression comparisons. Fig. S21. One-way ANOVA and Tukey t-test results from GluK gene family expression comparisons. Fig. S22. One-way ANOVA and Tukey t-test results from MIP gene family expression comparisons. Fig. S23. One-way ANOVA and Tukey t-test results from MIOX gene family expression comparisons. Fig. S24. One-way ANOVA and Tukey t-test results from Invertase gene family ANINV. Fig. S25. One-way ANOVA and Tukey t-test results from Invertase gene family CINV. Fig. S26. One-way ANOVA and Tukey t-test results from Invertase gene family CWI. Fig. S27. One-way ANOVA and Tukey t-test results from Invertase gene family VINV. Fig. S28. One-way ANOVA and Tukey t-test results from UGPase gene family expression comparisons. Fig. S29. One-way ANOVA and Tukey t-test results from AGP gene family expression comparisons. Fig. S30. One-way ANOVA and Tukey t-test results from PGM gene family expression comparisons. Fig. S31. One-way ANOVA and Tukey t-test results from G6PI gene family expression comparisons. Fig. S32. One-way ANOVA and Tukey t-test results from HXK gene family expression comparisons. Fig. S33. One-way ANOVA and Tukey t-test results from FK gene family expression comparisons.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Mason, P.J., Hoang, N.V., Botha, F.C. et al. Organ-specific expression of genes associated with the UDP-glucose metabolism in sugarcane (Saccharum spp. hybrids). BMC Genomics 24, 18 (2023). https://doi.org/10.1186/s12864-023-09124-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-023-09124-8