
Abstract
Such carbon structures as fullerenes, endofullerenes, nanotubes, nanodiamonds, and graphenes, which were discovered over the past few decades, possess a number of unique properties and could become the basis for the design of a new class of neuroprotective agents; however, despite years of research, this has not happened yet. In the first part of the review, the significance of the functionalization of carbon nanoparticles for their use in biology and medicine is described, and the data on their toxicity are also discussed. The second part presents the works of Russian and foreign scientists demonstrating the neuroprotective properties of carbon nanoparticles and the possibilities of their application in neurobiology and neurology. The proven experience of such experiments is described and the existing problems are indicated.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
CONTENTS
Introduction
1. Neuroprotective activity of carbon nanoparticles
1.1. Anti-amyloidogenic activity
1.2. Antioxidant activity
2. Nanoparticle as a vector for drug delivery through the blood-brain barrier
3. Diagnostics and imaging. Biosensors
4. Platforms and substrates for the growth and differentiation of neural cells
Conclusions
INTRODUCTION
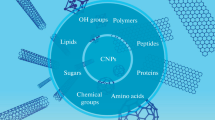
The proposed pattern of the initiation and development of some neurodegenerative diseases (NDDs) includes numerous factors, among which amyloid-protein aggregation and oxidative stress (OS) can be noted. Carbon nanoparticles (CNPs) could become the basis for designing neuroprotective agents since they have unique properties: fullerenes and their derivatives are known as powerful antioxidants [1, 2], nanotubes and nanodiamonds (NDs) are promising materials for the regeneration of damaged neural fibers [3–5], and graphene is a good basis for biosensors [6, 7]. Many nanoparticles (NPs) are able to cross the blood-brain barrier (BBB) and can be used both for direct treatment and for drug delivery to neural cells (Fig. 1). In the previous part of the review, we describe the need for the functionalization of CNPs, taking into account its directionality for successful application in biological and medical research, and analyze works aimed at studying the toxicity of NPs in vitro and in vivo. However, studies of the neuroprotective activity of these structures and the possibility of their application in neurology, neurobiology, and cellular technologies are of no less interest.

Application of carbon nanoparticles in biology and medicine.
1 NEUROPROTECTIVE ACTIVITY OF CARBON NANOPARTICLES
Currently, there are many studies demonstrating the neuroprotective activity of CNPs in various models of NDDs. For example, the effects of an aqueous suspension of fullerene C60, hydrated C60 fullerene (C60HyFn), and NDs on disease symptoms were analyzed in rat models of Alzheimer’s disease (AD). When studying the state of spatial memory in animals and comparing the obtained results with those of using donepezil, an acetylcholinesterase inhibitor used in AD therapy, the effectiveness of fullerene was shown [8]. C60HyFn prevented disturbance of the electroencephalogram spectra and reduced destruction of the cortical layer of the hippocampus in animals. The authors suggested that neuroprotective effects may be associated with the effect of fullerenol on presynaptic dopamine receptors [9]. The use of NDs improved learning and memory in rats, as well as the histological pattern in their brain [10].
The neuroprotective activity of fullerene derivatives (C60–OH and C60–NH2) was demonstrated in a prion-disease cell model in [11]. Polyethylene glycol (PEG)-functionalized single-walled carbon nanotubes (SWCNTs) were tested in a model of Parkinson’s disease (PD) in mice. They were administered intravenously or directly into the lateral ventricle of the brain. Regardless of the method of administration, SWCNTs prevented the death of dopaminergic neurons and behavioral disorders in mice [12]. Moreover, SWCNTs protected the neurons of stroke-induced rats from damage [13, 14]. There are data on the neuroprotective properties of graphene oxide (GO). Thus, it has been shown that GO enhances the differentiation of SH-SY5Y cells, increases the length of neurites and the expression of the neuronal marker MAP2 in human neuroblastoma SH-SY5Y cells [15]. In this case, the main mechanisms ensuring the neuroprotective activity of NPs may be their ability to prevent the aggregation of amyloid proteins and antioxidant activity (Fig. 2).

Neuroprotective activity of carbon nanoparticles.
1.1 Anti-amyloidogenic Activity
The impaired folding of some proteins, which leads to the formation of amyloid aggregates, can cause the development of a number of diseases, including AD, PD, Creutzfeldt–Jakob disease, type 2 diabetes, bovine spongiform encephalopathy, and various forms of systemic amyloidosis [16–18]. A common feature of these diseases is the transformation of proteins from a soluble, usually harmless form into insoluble amyloid aggregates with a high content of β-sheet structure [19, 20]. In addition to amyloid diseases, several other degenerative diseases may be due to intracellular or extracellular aggregate deposition, but their molecular basis is currently unknown [17]. It is believed that the general physico-chemical properties of proteins, such as average hydrophobicity, net charge, and the tendency to form α- and β structures, can affect the tendency to aggregation [21]. The accumulation of amyloid aggregates (oligomers, protofibrils, and fibrils) leads to violation of the regulation of cellular homeostasis, intracellular transport, and membrane integrity [18].
One of the strategies for treating diseases characterized by the deposition of pathological aggregates in organs and tissues is aimed at finding and developing drugs that can slow down or prevent their formation, as well as destroy already formed structures [22]. The use of CNPs as inhibitors of amyloidogenesis is of particular interest because of such physico-chemical properties as a large surface area with a very small size, the possibility of functionalization and modification, and the ability to penetrate through the BBB [23].
At present, a significant number of studies devoted to analysis of the anti-amyloid properties of fullerenes can be found in publications. It is assumed that the LVFF motif (from the 17th to the 20th amino-acid residue) of the amyloid beta peptide (Aβ) is of paramount importance in the generation of an ordered β structure of amyloid fibrils, and the hydrophobic aromatic residue of phenylalanine in position 19, which interacts with fullerenes much stronger than other amino-acid residues, is crucial in their formation and stabilization. At the same time, the phenylalanine–phenylalanine intermolecular interaction necessary for the formation of β layers is significantly weakened, which leads to slowing of the process of Aβ aggregation [24–26]. Using molecular-modeling methods, it has been shown that among C20, C36, C60, C70, and C84 fullerenes, C60 fullerene most effectively destroys fibrils formed by the Aβ17–42 peptide [24]. Using atomic force microscopy (AFM), it was found that C60 and C180 fullerenes inhibit the growth of fibrils formed by the Aβ16–22 peptide; moreover, C180 fullerene exhibits a stronger anti-amyloidogenic potential [25]. Various methods (fluorescence analysis (FA), AFM, and computer simulation) demonstrated the efficiency of C60(OH)16 fullerenol as an inhibitor of the growth of amyloid fibrils formed by the Aβ40 peptide [26]. The ability of water-soluble C60 fullerene derivatives to destroy mature amyloid fibrils formed by Aβ40 and Aβ42 and to inhibit the growth of new ones was found by electron microscopy (EM) and FA [27–29]. It has been shown by AFM that the incubation of Aβ42 with C60(OH)30 fullerenol for 24 h at 37°C reduces Aβ42-peptide aggregation, which indicates its anti-amyloidogenic activity [30]. Hydrophobic interaction plays a crucial role in the interaction between Aβ and NPs, this interaction is weakened with an increase in the number of hydroxyl groups. Thus, using molecular dynamics (MD) simulation, it has been found that an increase in the number of hydroxyl groups carried by C60 leads to slower dynamics and a weaker binding force of NPs to Aβ40/42 protofibrils [31]. In [32], it was concluded that for the development of anti-amyloid agents combining antioxidant and anti-aggregatory effects, an aqueous molecular-colloidal solution of C60 fullerene can be used; this solution prevents Aβ25–35 aggregation in the cytoplasm of neurons and reduces neurodegeneration in an AD model in rats, which was created using microinjections of Aβ25–35. An analysis of the dynamics of the hippocampus state showed that there was degeneration in most pyramidal cells and the Aβ25–35 deposits in the cytoplasm of neurons by day 14. Since OS accompanies these processes, it was assumed that an aqueous molecular-colloidal solution of C60 fullerene would prevent these negative effects, while acting as an antioxidant. The introduction of low concentrations of a C60-fullerene solution prevented neurodegeneration and the accumulation of amyloid in hippocampal neurons [32]. The fact that C60 fullerene and its derivatives efficiently reduce Aβ aggregation has also been reported in other works [33–37].
Fullerene derivatives can inhibit aggregation not only of the Aβ peptide, but also of other amyloid proteins. For example, using MD simulation, AFM, and thioflavin T staining, it has been demonstrated that C60(OH)24 suppresses human islet amyloid polypeptide (IAPP) aggregation, which is associated with type-2 diabetes [38]. С60(OH)30 and С70(OH)30 fullerenols prevented the aggregation of α-synuclein in a model of PD in Drosophila melanogaster [39]. The inhibitory effect of C60 fullerene derivatives on the formation of amyloid fibrils by muscle X protein of the titin family was studied by transmission electron microscopy (TEM). It has been shown that these structures not only destroy mature amyloid fibrils, but also prevent the formation of new ones [40]. Thus, at present, the anti-amyloidogenic activity of fullerenes and their derivatives can be considered a proven fact.
As for CNTs, they are characterized by hydrophobic surfaces with a high specific area [41], and when they enter a biological medium, they adsorb proteins, forming a “protein corona” [42]. Two opposite effects of CNTs on the process of fibril formation are reported in publications, namely, its acceleration and inhibition. It is assumed that the effect of CNTs depends on the affinity of proteins for CNTs, the conformation of proteins in the “protein corona”, and the kinetic barrier of fibril formation [43]. The effect of multiwalled carbon nanotubes (MWCNTs) on the aggregation of β2-microglobulin was studied; it was found that MWCNTs accelerated the process of fibril formation due to shortening of the lag phase [44]. It has been shown that β2-microglobulin can form several layers on the surface of MWCNTs; therefore, it is assumed that an increased concentration of protein on the surface of MWCNTs actually promotes the formation of oligomers. Since no mature fibrils associated with CNTs were found, it is hypothesized that CNTs act as conventional catalysts and accelerate the process of fibril formation [45]. The possibility of MWCNTs acting as artificial molecular chaperones, which help in protein folding and prevent its aggregation, was considered [46]. Using TEM, it has been demonstrated that MWCNTs prevent the 2,2,2-trifluoroethanol-induced aggregation of recombinant human fibroblast growth factor-1 (hFGF-1) containing β sheets. Moreover, the protein can either be adsorbed on the surface of MWCNTs with closed ends or encapsulated in MWCNTs with open ends. The study of circular-dichroism spectra in the ultraviolet (UV) range showed that MWCNTs not only prevented hFGF-1 aggregation in the presence of 2,2,2-trifluoroethanol, but also helped the protein to recover its native state. SWCNTs penetrate into the cytoplasm and nucleus through the lipid bilayer more easily than MWCNTs [47] and do not exhibit cytotoxicity [48, 49]. Therefore, they are more promising for the development of inhibitors of amyloid-protein aggregation. Thus, using the replica exchange molecular dynamics (REMD) method, it has been shown that the interaction of SWCNTs with amorphous Aβ16–22 octamers prevents the formation of β sheets and leads to the formation of disordered aggregates, and the interaction of SWCNTs with the prefibrillar bilayer of the β sheet destabilizes the structure of the β sheet also with the formation of disordered aggregates. Inhibition of the formation of Aβ16–22 β sheets and the destabilization of prefibrillar structures result from the action of the same forces between SWCNTs and the Aβ16–22 peptide: strong hydrophobic and π-stacking interactions [50]. Using MD simulation, it has been demonstrated that two unbonded Aβ25–35 β sheets combining parallel and antiparallel strands can be assembled into β barrels wrapping SWCNTs. This mechanism leads to blocking of the peptide seed for the further addition of monomers or oligomers and a decrease in the number of monomers or oligomers available for fibril growth. Both effects have an inhibitory effect on fibril formation; therefore, taking into account the results obtained, it has been suggested that SWCNTs can be powerful inhibitors of Aβ25–35 fibril formation [51].
The poor dispersibility of CNTs in an aqueous medium significantly hinders their practical application as inhibitors of amyloidogenesis. The functionalization of CNTs by adding hydrophilic groups (–NH2, –OH, –COOH, etc.) can significantly improve their dispersibility and biocompatibility [52]. The study of the inhibitory effect of hydroxylated SWCNTs (SWCNTs–OH) on Aβ42 fibril formation by fluorimetric analysis with thioflavin T and AFM showed that SWCNTs–OH inhibit fibrillogenesis and disaggregate preformed amyloid fibrils. Moreover, the ratio of hydroxyl groups in SWCNTs–OH plays an important role in their ability to block Aβ42 aggregation; an increase in the number of hydroxyl groups significantly improves the inhibitory activity of SWCNTs–OH [53]. The effect of SWCNTs–OH on hIAPP aggregation was studied by the REMD method [54]. It has been shown that SWCNTs–OH due to hydrogen bonds, and van der Waals and π-stacking interactions with hIAPP significantly weaken the bond between proteins, which are crucial for the formation of β sheets, thereby slowing down further aggregation. The data obtained as a result of modeling were confirmed by fluorimetric analysis with thioflavin T, the circular-dichroism method, TEM, and AFM. Experimental results have demonstrated that SWCNTs–OH interact with hIAPP and significantly inhibit hIAPP fibrillation.
Graphene can also interact with amyloids. In this case, an important role in amyloid–graphene interactions belongs to aromatic rings, since the relationship of graphene with amyloids containing aromatic amino acids is stronger than with those without them. Using density functional theory, it has been found that the interaction between graphene and amyloids is stronger than between two amyloid β sheets. This is consistent with experimental observations that graphene and its modifications (GO and graphene quantum dots (GQDs)) strongly interact with aromatic amino acids in amyloid side chains and inhibit the aggregation of amyloid fibrils [55]. Using FA with thioflavin T and TEM, it has been demonstrated that graphene blocks the formation of amyloid aggregates of egg-white lysozyme, presumably through π–π interactions between graphene and aromatic amino acids [56]. It has been shown both experimentally and theoretically that graphene can inhibit Aβ fibrillation and destroy preformed amyloid fibrils. Using MD simulation, it has been found that graphene nanolayers can penetrate into fibrils and extract peptides from them due to dispersion interactions between graphene and peptide molecules, as well as π-stacking interactions between graphene and phenylalanine, which significantly weakens the bond between peptide molecules inside the fibril.
Since graphene is insoluble in water, a promising direction is to study the anti-amyloidogenic properties of water-soluble graphene-based nanoparticles. Thus, it has been shown that GO and GQD sheets act as effective inhibitors of the in-vitro aggregation of full-sized and fragmented forms of Aβ [58–61], α-synuclein [62, 63], and hIAPP [64, 65]. GO and GQD sheets inhibited α-synuclein aggregation at high concentrations; however, at low NP-to-protein ratios, the reverse effect of increased aggregation was observed [63]. Thioflavin S-modified GO can effectively dissociate amyloid deposits when irradiated with a near-infrared (IR) laser in both the buffer and cerebrospinal fluid of mice. In addition, amyloid disaggregation can be controlled by changing the fluorescence of thioflavin-S dye that selectively binds to fibrils, but not to protein monomers [66].
Data that graphene can promote the amyloidogenesis of some proteins are not found in publications; however, graphite being a “stack” of multiple layers of graphene possesses this property [67, 68].
The analysis of publications on the ability of NDs to stimulate or inhibit the process of amyloidosis shows that a small number of research papers have been devoted to this issue. Thus, it has been found that NDs administered intraperitoneally to rats reduced the level of Aβ42 in the hippocampus [10], where pathological changes in AD occur earlier than in other brain areas. NDs appeared to be more efficient when combined with memantine administered orally to rats.
Despite the fact that the anti-amyloidogenic properties of CNPs require further study, the data obtained to date indicate that the development of amyloidosis nanotherapy based on them is certainly justified.
1.2 Antioxidant Activity
Among the factors contributing to the development of many NDDs, the OS plays the most important role. It is known that brain tissues have a high metabolic level and consume a significant amount of oxygen. Neural cells rich in lipids have a predominantly aerobic metabolism and, at the same time, a low activity of enzymes that neutralize reactive oxygen species (ROS) [69]. In addition, some cells, for example, astrocytes and oligodendrocytes, can accumulate iron ions as part of protein complexes and, therefore, are highly sensitive to OS [70]. ROS are capable of causing damage to cell membranes, DNA fragmentation, the inactivation of transport proteins, and inhibition of mitochondrial energy production. At the same time, the regeneration of neural cells is limited. Examples of neurodegenerative disorders associated with an excess of ROS and nitric oxide (NO) radicals include AD, PD, HD, Lou Gehrig’s disease, and amyotrophic lateral sclerosis [69, 71]. The OS makes its negative contribution in cases of brain and spinal-cord injuries, strokes, and ischemia. Various exogenous antioxidants are used to eliminate ROS in the brain and mitigate OS.
Among CNPs, fullerenes and their derivatives are the most promising in this regard. They possess antioxidant properties due to their ability to attach dozens of free radicals and deactivate them [1]. This prevents the harmful effects of OS on proteins, lipids, DNA, etc. Water-soluble derivatives of fullerenes, fullerenols, absorb free radicals more efficiently than traditional antioxidants [2]. Computer simulations have shown that C60 can absorb protons, acquiring a positive charge, and penetrate into mitochondria, separating respiration and phosphorylation, which, in turn, leads to a decrease in ROS production [72]. Currently, there is more and more evidence that fullerenes and their derivatives exhibit antioxidant activity not only in solutions, but also in cell cultures and in experiments in vivo. At the same time, they only reduce, but do not prevent the death of neural cells.
Oxidative stress can be associated with the excessive excitation of glutamate receptors and induce apoptosis through a cascade of events [73]. L-Glutamate is a neurotransmitter involved in learning and memory processes; in elevated concentrations, it can cause the degeneration and death of neurons, thus participating in genesis of the development of NDDs. It has been shown in neural cell culture that fullerenol exhibits a neuroprotective effect by blocking glutamate receptors and reducing the concentration of intracellular calcium [74]. It is assumed that OS promotes both the excitotoxic and apoptotic death of neurons. Two fullerene derivatives, C60(OH)12 and C60(OH)18–20O3–7, were studied; it was found that these preparations reduced OS-induced neuronal death in a culture by 80% [75]. Water-soluble carboxyfullerenes containing malonic-acid fragments turned out to be efficient neuroprotectors. They reduced the apoptotic death of cultured cortical neurons [76]. A similar compound (C3-fullero-tris-methanodicarboxylic acid) prevented apoptosis in cerebellar cells under OS conditions [77]. Soluble carboxyfullerenes DF70 and TF70 protected cells from oxidative damage by penetrating into them and localizing mainly in lysosomes [78]. According to [79], carboxyfullerenes can accumulate in mitochondria, neutralizing free radicals and protecting cells from apoptosis. Using computer molecular modeling, it was shown that the reactivity of carboxyfullerenes towards superoxide depends not only on the number of carboxyl groups, but also on their distribution on the fullerene surface. That is, there is a relationship between the activity of neuroprotection and the structure of the compound [80].
As is known, one of the causes of the death of substantia nigra neurons in PD may be dopamine oxidation and the impairment of dopaminergic neurotransmission. It turned out that carboxyfullerene can protect dopaminergic neurons from the harmful effects of OS caused by neurotoxins [81]. Therefore, it is not surprising that interest has arisen in the analysis of the antioxidant and neuroprotective effects of fullerene derivatives in PD models. Using a PD cell model, it was shown that C60(OH)24 fullerenol prevents mitochondrial dysfunction, reduces the amount of ROS, and increases cell viability [82]. In a study on monkeys, C3 carboxyfullerene was used; it is a functionalized soluble fullerene capable of penetrating into neuronal membranes and also possessing anti-inflammatory properties [83]. The use of C3 improved functional outcomes in a PD model in primates. Moreover, there was no evidence of drug toxicity. In 2018, the pharmacokinetics of C3 in primates and mice was reported [84]. According to the results obtained, the preparation was widely distributed throughout tissues, including the brain, and remained stable for a long time. The half-life of C3 was 8.2 ± 0.2 h. C3 was excreted by the liver and kidneys; there were no signs of renal, hepatic, electrolyte, or hematological abnormalities, as well as disturbances in the electrical activity of the heart. However, there is evidence that another carboxyfullerene, although it attenuated cortical infarction during ischemia in rats treated with it by intracerebroventricular infusion (0.3 mg per rat), caused adverse behavioral changes and even death in them. Intravenous administration of this preparation did not result in protection against cerebral infarction [85]. A study of C60(OH)30 and C70(OH)30 fullerenols in a model of PD in Drosophila melanogaster showed their neuroprotective activity, decreased the death of dopaminergic neurons in the brain of the flies, and reduced OS [39]. The study [86] proposes a new strategy for the treatment of PD by combining the clinical use of levodopa and potent antioxidants. In this work, a water-soluble C60 fullerene derivative prevented the levodopa-induced death of chromaffin cells in a culture, which were supposed to be used as transplants in the case of PD.
Fullerene derivatives proved to be efficient in other NDD models as well. For example, C60(OH)30 fullerenol decreased OS and reduced overall neurodegeneration in the brain of flies in a model of AD in Drosophila melanogaster [30]. The fullerene-based agent ABS-75 was developed for receptor-specific antioxidant therapy. It is a water-soluble fullerene derivative containing adamantyl groups, glutamate-receptor antagonists, and an antioxidant carboxyfullerene component [87]. The preparation combined antioxidant and anti-excitotoxic properties. It stopped oxidative damage in the spinal cord of mice in a chronic model of multiple sclerosis, reduced axonal degeneration and demyelination, and blocked the inflammatory component of the disease. As a result, the development of the disease was significantly slowed down. The treatment was especially efficient during the progressive stage of the disease. The use of hydrated C60 fullerene (C60HyFn) for the treatment of mice with hyperhomocysteinemia, which is also associated with AD, significantly reduced the level of homocysteine in blood serum, as well as the levels of TRPM2 gene expression and apoptosis in neurons. In this case, an increase in the level of TRPM2 expression can be induced by OS [88]. The fullerene derivative FC4S reduced the extent of infarction in focal cerebral ischemia. After the drug was intravenously administered to rats, the NO content in the plasma increased, and the level of lactate dehydrogenase decreased. It is assumed that the positive effects may be associated with its antioxidant properties and increased NO production [89]. At the same time, there is evidence that under certain conditions fullerenol can capture NO synthesized by special synthases, which is known to be an important signaling molecule, but can be cytotoxic in high concentrations. By capturing NO, fullerenol prevents a decrease in the activity of antioxidant enzymes, i.e., it works as a modulator of ROS involvement in cellular processes [90].
It should be noted that not all fullerene derivatives are antioxidants. For example, a water-soluble derivative of C60 fullerene, pyrrolidine with three ethylene glycol chains and three ammonium groups did not show a significant change in the concentration of the superoxide radical anion O2– and, as a result, did not reduce neuronal damage [91]. Thus, fullerene functionalization, in particular, the nature of chemical fragments attached to the fullerene base, the aggregation properties of the resulting product, and other factors can affect the antioxidant activity.
There are much less published data on the antioxidant activity of other CNPs. According to [92], the ability of the studied SWCNTs to absorb oxygen radicals was almost 40 times higher than that of DF-1 fullerene. When SWCNTs were functionalized with hydroxytoluene, a decrease in the antioxidant activity was observed. The OS level and mitochondrial disorders characteristic of NDDs were analyzed in SH-SY5Y human neuroblastoma cells [12]. When 50-μM oxidopamine was added to the culture medium, the amount of ROS in the cells strongly increased. If cultivation was carried out on a surface coated with SWCNTs–PEG, only a slight increase in ROS was observed, and the mitochondrial membrane was not damaged. The ROS level also decreased when the cells were directly treated with SWCNTs–PEG and oxidopamine.
There is information about graphene and its derivatives that, on the contrary, they can cause a dose-dependent OS and even lead to a slight decrease in cell viability at high concentrations [93].
It has been suggested that NDs possess antioxidant activity due to their inherent properties of oxidase, peroxidase, and catalase, which depend on the pH of the medium [94]. It has been hypothesized that the molecular mechanism of their peroxidase-like activity is the acceleration of electron transfer, the source of which is oxygen-containing functional groups on their surface. The antioxidant activity of hydrated carboxylated NDs has been demonstrated [95]; they had the ability to scavenge for free radicals and reduce oxidative damage in erythrocytes after γ irradiation. Carboxylated ND (ND–COOH) and the ND–COOH/eugenol complex also showed an antioxidant effect compared to the negative control. In this case, the preparations were highly stable, improved skin permeability, and did not exhibit significant toxic effects on murine macrophage cells [96]. There is evidence that lactoferrin-conjugated NDs exhibit antioxidant activity in osteoblast-like MC3T3-E1 cells treated with H2O2 [97]. However, no information about the use of NDs as antioxidants in the study of diseases of the nervous system has been found.
2 NANOPARTICLE AS A VECTOR FOR DRUG DELIVERY THROUGH THE BLOOD-BRAIN BARRIER
Recently, much attention has been paid to the design of systems for targeted efficient drug delivery to affected organs and tissues. Liposomes, dendrimers, various NPs, etc. are used for this [98–100]. In the treatment of NDDs, it is also necessary that drugs can cross the BBB. Being a restrictive barrier, the BBB prevents various molecules (including neurotoxic ones) from entering the central nervous system (CNS): the presence of tight contacts between cells and the absence of transport routes between them makes it difficult for solutes to pass through the cerebral endothelium. Important factors affecting the passive transport of particles through the BBB are their molecular weight and lipophilicity. But even substances with the property of lipophilicity can spontaneously return to the bloodstream [101]. If the medicinal agent is not able to cross the BBB, it is delivered directly to the brain tissue or by invasive methods of temporarily disrupting the BBB (osmotic, chemical, physical, and biological), which can lead to significant damage to the brain. It is clear that this approach is not efficient. The intranasal administration of preparations can be considered a promising method, but it must be remembered that even agents entering the CNS can be unselectively distributed in the brain, subjected to opsonization, and have significant side effects [101]. At the same time, most CNPs easily cross the cell membrane, including the membrane of BBB endothelial cells; therefore, their use as vectors for selective drug delivery to the brain is much safer and seems to be extremely promising. Review [102] describes in detail how NPs can pass through biological barriers, including the BBB. It has been noted that neuronal pathways can be used to deliver nanopharmaceuticals and, at the same time, they can provide the unintentional access of NPs to the brain. NPs are able not only to penetrate the BBB, but also, due to functionalization of the surface, release active ingredients in a certain place [103]. An ideal option would be the ability to deliver NPs to certain types of neural cells. There are various strategies for this: the use of ligands with dopamine, coating of NPs with apolipoprotein E, modification of their surface to facilitate penetration into certain groups of cells, etc. [104]. An important role in the distribution of NPs in the brain is played by the surface charge, which can be changed during functionalization. For example, cationic molecules are transported through the BBB more intensively than anionic or neutral ones [105].
Fullerenes due to their spherical shape and lipophilicity can easily penetrate into cells. It has been shown for the first time that fullerenol C60(OH)24 is able to induce the ionic permeability of a bilayer lipid membrane through the formation of ionic pores [106]. The interaction of fullerenes with lipid membranes and passive transport in them was analyzed by MD simulation. It has been shown that the permeability of the lipid membrane for fullerenes is much higher than for other known penetrants [107]. With appropriate modification, these NPs can serve as carriers of drugs, contrast agents, radiopharmaceuticals, and as systems for their targeted delivery. This is facilitated by the possibility of fullerenes to covalently bind to biologically active aromatic molecules [108]. There is evidence that the amino derivative of fullerene functionalized with plasmid vector DNA penetrates cells more efficiently than lipid-based vectors. The absorption of the fullerene/DNA complex by cells occurs by the mechanism of endocytosis; in this case, DNA is protected by fullerene from enzymatic cleavage, which increases the success of transfection [109]. The interaction of C60 fullerene containing porphyrin-like transition metal-N4 clusters (TMN4C55, TM = Fe, Co, and Ni) with the nonsteroidal anti-inflammatory drug ibuprofen was studied [110]; ibuprofen is proposed to be used in the complex treatment of AD and PD, since inflammatory processes are involved in their pathogenesis. The results showed that in such a complex, C60 strongly adsorbs ibuprofen via electrostatic bonding, i.e., it can serve as a drug delivery system. If the molecules of four chemotherapeutic agents for glioblastoma treatment are attached to C60-fullerene molecules, drug delivery to biological systems is facilitated. At the same time, in all cases, C60 did not violate the chemical characteristics of molecules, being a good carrier for their delivery to target cells [111].
The above examples show that the use of fullerenes as vectors has great prospects not only in oncology, where such developments are currently underway, but also in experimental and clinical neurology.
According to [112], CNTs have great potential as one of the components of delivery systems. It is believed that highly purified and functionalized CNTs are practically nontoxic if used at low concentrations and locally [113]. At the same time, they are able to carry large doses of radionuclides and chemotherapeutic agents, increasing the selectivity of pharmaceuticals and reducing side effects. However, there is evidence that inhalation or intravenous contact with them can cause vascular reactions, such as an increase in the number of atherosclerotic plaques, vasomotor dysfunction, as well as changes in blood pressure and cardiac dysrhythmia in laboratory animals [114]. However, the toxicity of CNTs can be reduced by strict control of their physico-chemical properties. For example, the efficacy of SWCNTs functionalized with PEG and lactoferrin to transport and deliver dopamine to the brain has been demonstrated in a model of PD in mice. In these mice, OS level and inflammatory responses were reduced [115].
Graphene as a platform for the delivery of genes and pharmaceuticals attracts researchers because of its large surface area and the presence of free π electrons. Due to these properties, graphene allows the loading of many bioactive compounds at once and their release in a controlled manner [116]. For example, GO has been successfully used to load an anticancer agent by simple physical adsorption via π stacking [117]. A GO-containing biocompatible hydrogel designed for efficient delivery of the vascular endothelial growth factor-165 gene for the treatment of cardiac injury was described. Its efficiency has been demonstrated in vivo in a rat model of myocardial infarction [118]. According to [116], graphene nanosheets can be suitable as a vector for gene delivery, since they protect DNA from degradation, ensure high transfection efficiency, and are easily absorbed by cells.
The largest number of scientific studies in the field of targeted drug delivery is devoted to NDs. Various aspects of the use of NDs in neurosciences, including the therapeutic potential of these NPs, are analyzed in review [119]. When bound with detonation nanodiamonds (DNDs), even quite complex substances such as proteins and enzymes retain their natural activity almost completely. In the case of using pharmaceuticals that are poorly soluble in water, DND clusters can facilitate their transfer to a soluble phase while maintaining a therapeutic effect [120]. When analyzing complexes of DNDs (mechanically deagglomerated) with several water-insoluble therapeutic agents, it turned out that, these pharmaceuticals in complex with NDs were easily soluble in water, indicating the applicability of NDs as agents for the delivery of such drugs [121]. Composite ND particles coated with PEG–PEI copolymer were used for the intracellular delivery of poorly soluble molecules [122]. The effect of the noncovalent or covalent attachment of hydrophilic polymers or surfactants to NDs on the efficacy of oral delivery of the water-insoluble agent curcumin was compared in [123]. The data obtained showed that covalent attachment was more efficient and biocompatible than pure NPs and NPs with a noncovalent coating. A system consisting of NDs, d-α-tocopherol, PEG, and succinate (TPGS) was developed in the work [124] also devoted to optimizing the oral delivery of curcumin. Curcumin was loaded into ND clusters, then TPGS was applied to this complex. The nanocomplex showed a high loading efficiency and delayed drug release. At the same time, there are few data suggesting that NPs, when administered orally, can penetrate the BBB [102]. It should be noted that curcumin is a strong natural antioxidant and has recently been studied as a neuroprotective agent [125]. The study [126] is concerned with elucidating the question of how the chemistry of the ND surface affects the adsorption and release of thiopronin, an antioxidant with low bioavailability used to treat cystinuria and rheumatoid arthritis. The study showed a high loading capacity in aminated NDs (NDs–NH2), while hydroxylated NDs (NDs–OH) were more efficient for gradual drug release. That is, by changing the chemical composition of the surface, it is possible to use NDs for specific purposes. In [127], the problem of loading NDs with a slow calcium channel blocker, amlodipine, was successfully solved. Amlodipine is able to pass through the BBB, but the authors believe that their work will open up new possibilities for other pharmaceuticals.
In some cases, NDDs are determined by hereditary factors, and then gene therapy may be effective. Positively charged DNDs can form complexes with nucleic acids and serve for gene delivery. A positive charge on the surface of DNDs can be generated by a polymer coating. The possibility of using NDs to deliver the CRISPR-Cas9 system for genome editing was analyzed [128]. Carboxylated NDs 3 nm in diameter were covalently conjugated to the mCherry fluorescent protein, to which two DNA constructs were attached: one encoded with the Cas9 endonuclease and the GFP reporter; the other encoded with sgRNA and contained an HDR template insert designed to introduce the c.625C>T mutation into the RS1 gene associated with the development of congenital X-linked retinoschisis (XLRS). As a result, it was possible to introduce a mutation into the mouse retina and observe several effects specific to XLRS. Such NPs have also been successfully delivered to human induced pluripotent stem cells (iPSCs). In [129], microRNA (miR-181a) was transfected into iPSCs using NDs and it has been shown that this promotes the differentiation of iPSCs into functional cardiomyocytes. That is, such a method of delivery may have clinical potential.
3 DIAGNOSTICS AND IMAGING. BIOSENSORS
CNPs represent a powerful platform for the design of biosensors and various diagnostic tools. Probes and sensors based on them can detect and measure the enzymatic activity of proteases, ions, and metabolites in neural cells in normal and pathological conditions, and study biochemical and physiological processes in neurons and glia [130]. Using CNPs, it is possible to make existing diagnostic methods more efficient. For example, fullerenes can be used to transfer unstable atoms into cells. Metal atoms placed inside fullerenes form endometallofullerenes. Unlike complexes containing bifunctional chelators that prevent the direct binding of a toxic or radioactive metal ion to the body environment and provide thermodynamic stability, the isotope in endometallofullerenes is packed in a closed carbon shell, so they are relatively nontoxic and more resistant to metabolism [131]. Endometallofullerenes can be used as contrast agents for magnetic resonance imaging and visualization using single-photon emission computed tomography.
The optical properties of graphene make it promising for bioimaging. For example, GQDs exhibit fluorescence, which can be used to improve imaging techniques, and detect and diagnose diseases at an early stage [132]. The intrinsic photoluminescence of OG sheets was used to visualize living cells in the visible and IR ranges [117]. Review [7] discusses various strategies for designing biosensors based on graphene and directions for future research. At present, many biosensors, including those containing aptamers, are used in healthcare. The authors summarized data on the interaction of GO with DNA and information on the use of both its intrinsic fluorescence and properties used to develop fluorescent sensors, such as the quenching of luminescence and, conversely, signal amplification. As a result, they came to the conclusion that graphene is ideal for the multiplex detection of various targets, including DNA, proteins, metal ions, etc. In addition, GO can simultaneously be a means for the delivery of DNA- and RNA-based sensors into cells where they cannot penetrate on their own, and even applied to the semiquantitative determination of target molecules. For example, using the strategy of exonuclease-III-aided amplification, a turn-on aptasensor on the GO platform for detecting lysozyme, which is known to be an amyloid protein, was constructed. The fluorescence determined by this method linearly depended on the protein concentration. The sensitivity of the sensor was very high. It is assumed that other proteins can also be detected using this method [133].
NDs have strong stable fluorescence and therefore also have great prospects for the visualization of biological objects. For example, fluorescent NDs coated with PEG based on human serum albumin have been found to pass through the BBB and enter the brains of mice. They can be traced at the level of individual cells, neurons and astrocytes, i.e., they can be used to visualize the latter [134]. Fluorescent NDs exhibiting brightness and photostability were also used to measure axon transport, the disruption of which is characteristic of hereditary forms of NDDs, which made it possible to detect NDs inside neurons [135]. Fluorescent ND with biocompatible properties was used to study the differentiation of neural cells [136]. The latter were obtained from stem cells, and the presence of NDs in neurons was determined using confocal microscopy. We note that NPs did not affect the morphology, cell viability, and expression of the neuron-specific marker β-III-tubulin.
4 PLATFORMS AND SUBSTRATES FOR THE GROWTH AND DIFFERENTIATION OF NEURAL CELLS
One of the areas of neuroscience is the construction of implants that can eliminate defects in the nervous system. In addition, cell-replacement therapy involves the introduction of in vitro grown neurons into brain structures. Currently, there are a number of works proving the prospects of this developing area. Several reviews systematize studies of CNTs as substrates for the growth of neural cells [4, 113, 137]. It is believed that their shape and properties, in particular electrical conductivity, contribute to this. These reviews provide examples of studies in which neural cells were successfully cultured on SWCNT substrates, forming tight contacts with the latter. At the same time, the neurons responded to external electrical stimulation through CNTs; i.e., complete biocompatibility of neurons and SWCNTs was observed. For example, SH-SY5Y human neuroblastoma cells were differentiated into neurons on the surface of glass coated with a SWCNT–PEG complex. The cell morphology and neurite length were assessed after seven days. They were longer and more complex than in the control [12]. CNTs can be successfully used not only to study the growth of neurons and the organization of neural networks, but also to increase the efficiency of signal transmission in the nervous system and to construct nanoelectrodes that stimulate neurons as efficiently as metal ones, but cause a lesser inflammatory response [138].
Graphene-coated substrates also have the potential for neural-cell growth and stimulation. Their electrical properties and conductivity are much better than those of graphite, and the resistivity is lower. Due to the same properties, micromolar concentrations of neurotransmitters, such as dopamine, can be detected on graphene-coated surfaces [139]. To improve neuron attachment, 2D and 3D polymer scaffolds were constructed based on graphene. Experiments on cell cultures have shown that constructs promote both their adhesion and growth [140]. The 3D scaffolds mimic the natural extracellular matrix microenvironment and do not elicit a strong immune response in vivo; so, their construction is of great importance in biomedicine, for example, for repairing damaged nerves. In [141], the case in point is a nanocomposite scaffold consisting of a conducting polymer and GO nanosheets. Molecules of interferon-γ or platelet growth factor, which promoted the differentiation and growth of neurons or oligodendrocytes, respectively, were covalently attached to the surface of the nanocomposite through the functional groups of carboxylic acid. Thus, the scaffold could be changed depending on the goals and objectives of the study. It is assumed that the addition of GO to polymer substrates for growing cells, including neurons, can improve their mechanical properties due to oxygen-containing functional groups on its surface interacting with hydroxyl or amino groups of polymers, providing interfacial bonding between them and GO [139]. The topic of using graphene for the growth and differentiation of neurons, their regeneration, stimulation, and detection of their activity is well covered in reviews [116, 142].
Due to their chemical inertness and biocompatibility, NDs are also being actively studied as a potential substrate for the cultivation of neural cells. The characteristics of the crystal structure of NDs, which are optimal for the differentiation and growth of neurons, were studied [5]. Smaller crystals have been shown to contribute to a higher neurite density. In this case, boron as a dopant included in the structure of NDs had no effect on cells. Neuroblastoma–glioma hybrid cells, as well as neurons and Schwann cells isolated from rats, were cultured on a substrate of amino-functionalized NDs [143]. Human neural stem cells grew well and spontaneously differentiated into neurons on glass coated with NDs (10 nm) with oxygen and hydrogen terminal groups. The use of NDs with an oxygen group increased the total length of neurites, their branching, and density. In this case, NDs were attached to glass using simple sonication [144]. When studying various ND coatings, the ability to increase the adhesion of neural cells was shown to be affected by the size and curvature of NPs [145]. It was also found that neurons can be cultured on various substrates coated with NDs just as successfully as on standard materials coated with protein [146]. Thus, the interaction of insect circadian-pacemaker neurons with the surfaces of ultra-crystalline NDs was studied [147]. It was shown that the cells were quickly and firmly attached to the substrate without using adhesion proteins, which made it possible to significantly improve the protocol for the preparation of primary cultures of neural cells, as well as the methods of their immunocytochemical staining. A new material made from ultra-nanocrystalline diamond using nitrogen was reported. Treatment of the resulting films with oxygen plasma affected their electrochemical capacity and biocompatibility. This material was used for the growth of rat cerebral-cortex neurons, and the surface roughness turned out to play a significant role [148]. Thus, surfaces coated with NDs in simple and inexpensive ways promote neuronal growth. This may be of importance for neurobiological research, as well as for the design of medical implants and devices with diamond-coated parts that come into contact with the nervous system.
NPs for the thermal stimulation of neurons in deep areas of the brain or as an alternative to optogenetic methods can be used as magnetic ones, in addition to the CNPs described above [130]. Electrodes made from CNP-based composites have a unique molecular structure and good electrical conductivity [4]. GO can be considered as a promising candidate for the photothermal treatment of AD, which has better selectivity and fewer side effects compared to traditional chemotherapy and radiotherapy, since only the affected area of the body is exposed to light [66].
CONCLUSION
Thus, CNPs find application in various areas of neuroscience due to their unique properties. But if the use of CNPs as substrates for the growth of neural cells does not raise questions, then the developments associated with their entry into the human body cause distrust in both the scientific and medical communities. The main questions are related to the toxicity of CNPs and their accumulation inside the body, and this despite numerous studies that contradict these claims. However, recently it has become obvious that these disagreements can be resolved, since a sufficient number of works have been accumulated, indicating that the toxicity of CNPs and their behavior in the body depend on controlled factors, such as drug purity, functionalization features, and physico-chemical characteristics. By taking these factors into account, it is possible to obtain NPs that can do much more good than harm.
REFERENCES
Y. Yamakoshi, N. Umezawa, A. Ryu, et al., J. Am. Chem. Soc. 125, 12803 (2003). https://doi.org/10.1021/ja0355574
S. Trajkovic, S. Dobric, V. Jacevic, et al., Colloids Surf., B 58, 39 (2007). https://doi.org/10.1016/j.colsurfb.2007.01.005
A. A. John, M. V. Vellayappan, A. Balaji, et al., Int. J. Nanomed. 10, 4267 (2015). https://doi.org/10.2147/IJN.S83777
C. Xiang, Y. Zhang, W. Guo, and X. J. Liang, Acta Pharm. Sin. B 10, 239 (2020). https://doi.org/10.1016/j.apsb.2019.11.003
P. A. Nistor, P. W. May, F. Tamagnini, et al., Biomaterials 61, 139 (2015). https://doi.org/10.1016/j.biomaterials.2015.04.050
P. Carneiro, S. Morais, and M. C. Pereira, Nanomaterials (Basel) 9, 1663 (2019). https://doi.org/10.3390/nano9121663
Z. Liu, B. Liu, J. Ding, and J. Liu, Anal. Bioanal. Chem. 406, 6885 (2014). https://doi.org/10.1007/s00216-014-7888-3
M. Nikpour, A. Sharafi, M. Hamidi, and S. Andalib, ACS Chem. Neurosci. 11, 2549 (2020). https://doi.org/10.1021/acschemneuro.0c00213
V. Vorobyov, V. Kaptsov, R. Gordon, et al., J. Alzheimers Dis. 45, 217 (2015). https://doi.org/10.3233/JAD-142469
S. H. Alawdi, E. S. El-Denshary, M. M. Safar, et al., Mol. Neurobiol. 54, 1906 (2017). https://doi.org/10.1007/s12035-016-9762-0
S. Ye, T. Zhou, D. Pan, et al., J. Biomed. Nanotechnol. 12, 1820 (2016). https://doi.org/10.1166/jbn.2016.2281
O.-H. Kim, J. H. Park, J. I. Son, et al., Int. J. Nanomed. 15, 7615 (2020). https://doi.org/10.2147/IJN.S276380
H. J. Lee, J. Park, O. J. Yoon, et al., Nat. Nanotechnol. 6, 121 (2011). https://doi.org/10.1038/nnano.2010.281
S. U. Moon, J. Kim, K. K. Bokara, et al., Int. J. Nanomed. 7, 2751 (2012). https://doi.org/10.2147/IJN.S30273
M. Lv, Y. Zhang, L. Liang, et al., Nanoscale 4, 3861 (2012). https://doi.org/10.1039/c2nr30407d
P. K. Singh and S. K. Maji, Cell. Biochem. Biophys. 64, 29 (2012). https://doi.org/10.1007/s12013-012-9364-z
M. Stefani, Biochim. Biophys. Acta 1739, 5 (2004). https://doi.org/10.1016/j.bbadis.2004.08.004
A. Bellova, E. Bystrenova, M. Koneracka, et al., Nanotechnology 21, 065103 (2010). https://doi.org/10.1088/0957-4484/21/6/065103
L. C. Serpell, C. C. Blake, and P. E. Fraser, Biochemistry 39, 13269 (2000). https://doi.org/10.1021/bi000637v
M. Sunde and C. Blake, Adv. Protein Chem. 50, 123 (1997). https://doi.org/10.1016/s0065-3233(08)60320-4
F. Chiti, M. Stefani, N. Taddei, et al., Nature (London, U.K.) 424 (6950), 805 (2003). https://doi.org/10.1038/nature01891
A. G. Bobylev, L. G. Bobyleva, I. M. Vikhlyantsev, A. D. Ulanova, N. N. Salmov, and Z. A. Podlubnaya, Biophysics 58, 754 (2013).
F. Liu, W. Wang, J. Sang, et al., ACS Chem. Neurosci. 10, 588 (2019). https://doi.org/10.1021/acschemneuro.8b00441
P. D. Q. Huy, and M. S. Li, Phys. Chem. Chem. Phys. 16, 20030 (2014). https://doi.org/10.1039/c4cp02348j
L. Xie, Y. Luo, D. Lin, et al., Nanoscale 6, 9752 (2014). https://doi.org/10.1039/c4nr01005a
Z. Bednarikova, P. D. Q. Huy, M.-M. Mocanu, et al., Phys. Chem. Chem. Phys. 18, 18855 (2016). https://doi.org/10.1039/c6cp00901h
A. G. Bobylev, O. A. Kraevaya, L. G. Bobyleva, et al., Colloids Surf., B 183, 110426 (2019). https://doi.org/10.1016/j.colsurfb.2019.110426
A. G. Bobylev, L. G. Marsagishvili, and Z. A. Podlubnaya, Biophysics 55, 699 (2010).
A. G. Bobylev, L. G. Marsagishvili, M. D. Shpagina, V. S. Romanova, R. A. Kotelnikova, and Z. A. Podlubnaya, Biophysics 55, 353 (2010).
A. D. Slobodina, O. I. Bolshakova, A. E. Komissarov, N. V. Surina, S. B. Landa, P. A. Melent’ev, and S. V. Sarantseva, Nanotechnol. Russ. 15, 212 (2020).
Z. Liu, Y. Zou, Q. Zhang, et al., Int. J. Mol. Sci. 20, 2048 (2019). https://doi.org/10.3390/ijms20082048
E. G. Makarova, R. Y. Gordon, and I. Y. Podolski, J. Nanosci. Nanotechnol. 12, 119 (2012). https://doi.org/10.1166/jnn.2012.5709
H. M. Huang, H. C. Ou, S. J. Hsieh, and L. Y. Chiang, Life Sci. 66, 1525 (2000). https://doi.org/10.1016/s0024-3205(00)00470-7
J. E. Kim and M. Lee, Biochem. Biophys. Res. Commun. 303, 576 (2003). https://doi.org/10.1016/S0006-291X(03)00393-0
C.-M. Lee, S. T. Huang, S. H. Huang, et al., Nanomed. Nanotechnol. Biol. Med. 7, 107 (2011). https://doi.org/10.1016/j.nano.2010.06.009
I. Ya. Podolski, Z. A. Podlubnaya, and O. V. Godukhin, Biophysics 55, 71 (2010).
I. Y. Podolski, Z. A. Podlubnaya, E. A. Kosenko, et al., J. Nanosci. Nanotechnol. 7, 1479 (2007). https://doi.org/10.1166/jnn.2007.330
C. Bai, D. Lin, Y. Mo, et al., Phys. Chem. Chem. Phys. 21, 4022 (2019). https://doi.org/10.1039/C8CP07501H
I. Golomidov, O. Bolshakova, A. Komissarov, et al., Biochem. Biophys. Res. Commun. 523, 446 (2020). https://doi.org/10.1016/j.bbrc.2019.12.075
L. G. Marsagishvili, A. G. Bobylev, M. D. Shpagina, P. A. Troshin, and Z. A. Podlubnaya, Biophysics 54, 135 (2009).
A. Peigney, Ch. Laurent, E. Flahaut, et al., Carbon 39, 507 (2001). https://doi.org/10.1016/S0008-6223(00)00155-X
C. Ge, J. Du, L. Zhao, et al., Proc. Natl. Acad. Sci. U. S. A. 108, 16968 (2011). https://doi.org/10.1073/pnas.1105270108
C. Li and R. Mezzenga, Nanoscale 5, 6207 (2013). https://doi.org/10.1039/c3nr01644g
S. Linse, C. Cabaleiro-Lago, W.-F. Xue, et al., Proc. Natl. Acad. Sci. U. S. A. 104, 8691 (2007). https://doi.org/10.1073/pnas.0701250104
V. L. Colvin and K. M. Kulinowski, Proc. Natl. Acad. Sci. U. S. A. 104, 8679 (2007). https://doi.org/10.1073/pnas.0703194104
A. V. Ghule, K. M. Kathir, T. K. S. Kumar, et al., Carbon 45, 1586 (2007). https://doi.org/10.1016/j.carbon.2007.03.043
A. E. Porter, M. Gass, K. Muller, et al., Nat. Nanotechnol. 2, 713 (2007). https://doi.org/10.1038/nnano.2007.347
H.-X. Ren, X. Chen, J.-H. Liu, et al., Mater. Today 13, 6 (2010). https://doi.org/10.1016/S1369-7021(10)70002-X
J.-P. Kaiser, M. Roesslein, T. Buerki-Thurnherr, and P. Wick, Curr. Med. Chem. 18, 2115 (2011). https://doi.org/10.2174/092986711795656171
H. Li, Y. Luo, P. Derreumaux, and G. Wei, Biophys. J. 101, 2267 (2011). https://doi.org/10.1016/j.bpj.2011.09.046
Z. Fu, Y. Luo, P. Derreumaux, and G. Wei, Biophys. J. 97, 1795 (2009). https://doi.org/10.1016/j.bpj.2009.07.014
J. Shin, S. Lee, and M. Cha, Med. Chem. Commun. 8, 625 (2017). https://doi.org/10.1039/c6md00716c
F. Liu, W. Wang, J. Sang, et al., ACS Chem. Neurosci. 10, 588 (2019). https://doi.org/10.1021/acschemneuro.8b00441
Y. Mo, S. Brahmachari, J. Lei, et al., ACS Chem. Neurosci. 9, 2741 (2018). https://doi.org/10.1021/acschemneuro.8b00166
D. M. Bozinovski, P. V. Petrovic, M. R. Belic, and S. D. Zaric, ChemPhysChem 19, 1226 (2018). https://doi.org/10.1002/cphc.201700847
W. E. Gregory, B. Sharma, L. Hu, et al., Biointerphases 15, 031010 (2020). https://doi.org/10.1116/6.0000019
Z. Yang, C. Ge, J. Liu, et al., Nanoscale 7, 18725 (2015). https://doi.org/10.1039/c5nr01172h
Q. Li, L. Liu, S. Zhang, et al., Chem. Eur. J. 20, 7236 (2014). https://doi.org/10.1002/chem.201402022
S. Bag, A. Sett, S. DasGupta, and S. Dasgupta, RSC Adv. 6 (105), 103242 (2016). doi.org/https://doi.org/10.1039/C6RA23570K
Y. Liu, L. P. Xu, W. Dai, et al., Nanoscale 7, 19060 (2015). doi.org/https://doi.org/10.1039/C5NR06282A
Z. Yang, C. Ge, J. Liu, et al., Nanoscale 7, 18725 (2015). doi.org/https://doi.org/10.1039/C5NR01172H
M. Mahmoudi, O. Akhavan, M. Ghavami, et al., Nanoscale 4, 7322 (2012). https://doi.org/10.1039/c2nr31657a
M. Ghaeidamini, D. Bernson, N. Sasanian, et al., Nanoscale 12, 19450 (2020). https://doi.org/10.1039/d0nr05003b
P. Nedumpully-Govindan, E. N. Gurzov, P. Chen, et al., Phys. Chem. Chem. Phys. 18, 94 (2016). https://doi.org/10.1039/c5cp05924k
M. Wang, Y. Sun, X. Cao, et al., Nanoscale 10, 19995 (2018). https://doi.org/10.1039/c8nr07180b
M. Li, X. Yang, J. Ren, et al., Adv. Mater. 24, 1722 (2012). https://doi.org/10.1002/adma.201104864
X. Yu, Q. Wang, Y. Lin, et al., Langmuir 28, 6595 (2012). https://doi.org/10.1021/la3002306
X. Mao, Y. Wang, L. Liu, et al., Langmuir 25, 8849 (2009). https://doi.org/10.1021/la901342r
A. Singh, R. Kukreti, L. Saso, and S. Kukreti, Molecules 24, 1583 (2019). https://doi.org/10.3390/molecules24081583
J. Grebowski, P. Kazmierska, and A. Krokosz, BioMed Res. Int. 2013, 751913 (2013). https://doi.org/10.1155/2013/751913
J. Emerit, M. Edeas, and F. Bricaire, Biomed. Pharmacother. 58, 39 (2004). https://doi.org/10.1016/j.biopha.2003.11.004
V. A. Chistyakov, Y. O. Smirnova, E. V. Prazdnova, and A. V. Soldatov, BioMed Res. Int. 2013, 821498 (2013). https://doi.org/10.1155/2013/821498
S. Bosi, T. da Ros, G. Spalluto, and M. Prato, Eur. J. Med. Chem. 38, 913 (2003). https://doi.org/10.1016/j.ejmech.2003.09.005
H. Jin, W. Q. Chen, X. W. Tang, et al., J. Neurosci. Res. 62, 600 (2000). https://doi.org/10.1002/1097-4547(20001115)62:4<600::AID-JNR15>3.0.CO;2-F
L. L. Dugan, J. K. Gabrielsen, S. P. Yu, et al., Neurobiol. Dis. 3, 129 (1996). https://doi.org/10.1006/nbdi.1996.0013
L. L. Dugan, D. M. Turetsky, C. Du, et al., Proc. Natl. Acad. Sci. U. S. A. 94, 9434 (1997). https://doi.org/10.1073/pnas.94.17.9434
M. Bisaglia, B. Natalini, R. Pellicciari, et al., J. Neurochem. 74, 1197 (2000). https://doi.org/10.1046/j.1471-4159.2000.741197.x
Q. Liu, X. Zhang, X. Zhang, et al., ACS Appl. Mater. Interfaces 5, 11101 (2013). https://doi.org/10.1021/am4033372
F. Chirico, C. Fumelli, A. Marconi, et al., Exp. Dermatol. 16, 429 (2007). https://doi.org/10.1111/j.1600-0625.2007.00545.x
S. S. Ali, J. I. Hardt, and L. L. Dugan, Nanomed. Nanotechnol. Biol. Med. 4, 283 (2008). https://doi.org/10.1016/j.nano.2008.05.003
J. Lotharius, L. L. Dugan, and K. L. O’Malley, J. Neurosci. 19, 1284 (1999).
X. Cai, H. Jia, Z. Liu, et al., J. Neurosci. Res. 86, 3622 (2008). https://doi.org/10.1002/jnr.21805
L. L. Dugan, L. Tian, K. L. Quick, et al., Ann. Neurol. 76, 393 (2014). https://doi.org/10.1002/ana.24220
J. I. Hardt, J. S. Perlmutter, C. J. Smith, et al., Eur. J. Drug Metab. Pharmacokinet. 43, 543 (2018). https://doi.org/10.1007/s13318-018-0464-z
A. M. Y. Lin, S. F. Fang, S. Z. Lin, et al., Neurosci. Res. 43, 317 (2002). https://doi.org/10.1016/S0168-0102(02)00056-1
A. A. Corona-Morales, A. Castell, A. Escobar, et al., J. Neurosci. Res. 71, 121 (2003). https://doi.org/10.1002/jnr.10456
A. S. Basso, D. Frenkel, F. J. Quintana, et al., J. Clin. Invest. 118, 1532 (2008). https://doi.org/10.1172/JCI33464
E. O. Etem, R. Bal, A. E. Akagac, et al., J. Recept. Signal Transduct. Res. 34, 317 (2014). https://doi.org/10.3109/10799893.2014.896381
S. S. Huang, S. K. Tsai, C. L. Chih, et al., Free Radic. Biol. Med. 30, 643 (2001). https://doi.org/10.1016/S0891-5849(00)00505-0
S. M. Mirkov, A. N. Djordjevic, N. L. Andric, et al., Nitric Oxide 11, 201 (2004). https://doi.org/10.1016/j.niox.2004.08.003
C. Cusan, T. da Ros, G. Spalluto, and S. Foley, Eur. J. Org. Chem. 17, 2928 (2002). https://doi.org/10.1002/1099-0690(200209)2002:17<2928::AID-EJOC2928>3.0.CO;2-I
R. M. Lucente-Schultz, V. C. Moore, A. D. Leonard, et al., J. Am. Chem. Soc. 131, 3934 (2009). https://doi.org/10.1021/ja805721p
Y. Chang, S.-T. Yang, J.-H. Liu, et al., Toxicol. Lett. 200, 201 (2011). https://doi.org/10.1016/j.toxlet.2010.11.016
T. M. Chen, X. M. Tian, L. Huang, et al., Nanoscale 9, 15673 (2017). https://doi.org/10.1039/C7NR05629J
K. Santacruz-Gomez, A. Sarabia-Sainz, M. Acosta-Elias, et al., Nanotechnology 29, 125707 (2018). https://doi.org/10.1088/1361-6528/aaa80e
D. Lim, K. Kim, S. Lim, et al., Int. J. Nanomed. 11, 2381 (2016). https://doi.org/10.2147/IJN.S104859
S. E. Kim, S. Choi, J.-Y. Hong, et al., Nanomaterials 10, 50 (2019). https://doi.org/10.3390/nano10010050
R. Singh and J. W. Lillard, Exp. Mol. Pathol. 86, 215 (2009). https://doi.org/10.1016/j.yexmp.2008.12.004
S. Cunha, M. Amaral, J. Lobo, and A. Silva, Curr. Med. Chem. 23, 3618 (2016). https://doi.org/10.2174/0929867323666160824162401
S. V. Sarantseva, O. I. Bol’shakova, S. I. Timoshenko, A. A. Kolobov, et al., Byull. Eksp. Biol. Med. 150 (10), 402 (2011).
S. Ramanathan, G. Archunan, M. Sivakumar, et al., Int. J. Nanomed. 13, 5561 (2018). https://doi.org/10.2147/IJN.S149022
A. F. Hubbs, L. M. Sargent, D. W. Porter, et al., Toxicol. Pathol. 41, 395 (2013). https://doi.org/10.1177/0192623312467403
K. T. Nguyen, M. N. Pham, T. V. Vo, et al., Curr. Drug Metab. 18, 786 (2017). https://doi.org/10.2174/1389200218666170125114751
Y. Zorkina, O. Abramova, V. Ushakova, et al., Molecules (Basel) 25, 5294 (2020). https://doi.org/10.3390/molecules25225294
L. Fenart, A. Casanova, B. Dehouck, et al., J. Pharmacol. Exp. Ther. 291, 1017 (1999).
T. I. Rokitskaya and Y. N. Antonenko, Biochim. Biophys. Acta: Biomembr. 1858, 1165 (2016). https://doi.org/10.1016/j.bbamem.2016.02.009
D. Bedrov, G. D. Smith, H. Davande, and L. Li, J. Phys. Chem. B 112, 2078 (2008). https://doi.org/10.1021/jp075149c
A. Borowik, Y. Prylutskyy, Ł. Kawelski, et al., Colloids Surf., B 164, 134 (2018). https://doi.org/10.1016/j.colsurfb.2018.01.026
H. Isobe, W. Nakanishi, N. Tomita, et al., Mol. Pharm. 3, 124 (2006). https://doi.org/10.1021/mp050068r
E. Alipour, F. Alimohammady, A. Yumashev, and A. Maseleno, J. Mol. Model. 26, 7 (2020). https://doi.org/10.1007/s00894-019-4267-1
P. N. Samanta and K. K. Das, J. Mol. Graph. Model. 72, 187 (2017). https://doi.org/10.1016/j.jmgm.2017.01.009
L. Lacerda, A. Bianco, M. Prato, and K. Kostarelos, Adv. Drug Deliv. Rev. 58, 1460 (2006). https://doi.org/10.1016/j.addr.2006.09.015
I. V. Mitrofanova, I. V. Mil’to, I. V. Sukhodolo, and G. Yu. Vasyukov, Byull. Sib. Med. 13 (1), 135 (2014). https://doi.org/10.20538/1682-0363-2014-1-135-144
Y. Cao and Y. Luo, Toxicol. Appl. Pharmacol. 385, 114801 (2019). https://doi.org/10.1016/j.taap.2019.114801
Q. Guo, H. You, X. Yang, et al., Nanoscale 9, 10832 (2017). https://doi.org/10.1039/C7NR02682J
S. R. Shin, Y.-C. Li, H. L. Jang, et al., Adv. Drug Deliv. Rev. 105, 255 (2016). https://doi.org/10.1016/j.addr.2016.03.007
X. Sun, Z. Liu, K. Welsher, et al., Nano Res. 1, 203 (2008). https://doi.org/10.1007/s12274-008-8021-8
A. Paul, A. Hasan, H. A. Kindi, et al., ACS Nano 8, 8050 (2014). https://doi.org/10.1021/nn5020787
J. Saraf, K. Kalia, P. Bhattacharya, and R. K. Tekade, Drug Discov. Today. 24, 584 (2019). https://doi.org/10.1016/j.drudis.2018.10.012
L. B. Piotrovskii, D. N. Nikolaev, and O. A. Shenderova, in Detonation Nanodiamonds. Technology, Structure, Properties and Applications, Ed. by A. Ya. Vul’ and O. A. Shenderova (FTI im. A. F. Ioffe, St. Petersburg, 2016), p. 317 [in Russian].
M. Chen, E. D. Pierstorff, R. Lam, et al., ACS Nano 3, 2016 (2009). https://doi.org/10.1021/nn900480m
N. Prabhakar, T. Nareoja, E. von Haartman, et al., Nanoscale 5, 3713 (2013). https://doi.org/10.1039/c3nr33926b
D. Liu, S. Qiao, B. Cheng, et al., AAPS PharmSciTech. 21, 187 (2020). https://doi.org/10.1208/s12249-020-01721-0
B. Cheng, H. Pan, D. Liu, et al., Int. J. Pharm. 540, 162 (2018). https://doi.org/10.1016/j.ijpharm.2018.02.014
A. Askarizadeh, G. E. Barreto, N. C. Henney, et al., Int. J. Pharm. 585, 119476 (2020). https://doi.org/10.1016/j.ijpharm.2020.119476
J. Beltz, A. Pfaff, I. M. Abdullahi, et al., Diamond Relat. Mater. 100, 107590 (2019). https://doi.org/10.1016/j.diamond.2019.107590
S. H. Alawdi, H. Eidi, M. M. Safar, and M. A. Abdel-Wahhab, Nanotechnol. Sci. Appl. 12, 47 (2020). https://doi.org/10.2147/NSA.S232517
T.-C. Yang, C.-Y. Chang, A. A. Yarmishyn, et al., Acta Biomater. 101, 484 (2020). https://doi.org/10.1016/j.actbio.2019.10.037
C. Y. Liu, M. C. Lee, H. F. Lin, et al., J. Chin. Med. Assoc. 84, 177 (2021). https://doi.org/10.1097/JCMA.0000000000000441
D. Maysinger, J. Ji, E. Hutter, and E. Cooper, Front Neurosci. 9, 480 (2015). https://doi.org/10.3389/fnins.2015.00480
R. Bakry, R. M. Vallant, M. Najam-ul-Haq, et al., Int. J. Nanomed. 2, 639 (2007).
T. K. Henna, V. R. Raphey, R. Sankar, et al., Int. J. Pharm. 587, 119701 (2020). https://doi.org/10.1016/j.ijpharm.2020.119701
C. Chen, J. Zhao, J. Jiang, and R. Yu, Talanta 101, 357 (2012). https://doi.org/10.1016/j.talanta.2012.09.041
P. Moscariello, M. Raabe, W. Liu, et al., Small 15, 1902992 (2019). https://doi.org/10.1002/smll.201902992
S. Haziza, N. Mohan, Y. Loe-Mie, et al., Nat. Nanotechnol. 12, 322 (2017). https://doi.org/10.1038/nnano.2016.260
T. C. Hsu, K. K. Liu, H. C. Chang, et al., Sci. Rep. 4, 5004 (2015). https://doi.org/10.1038/srep05004
E. B. Malarkey and V. Parpura, Brain Edema XIV 106, 337 (2010). https://doi.org/10.1007/978-3-211-98811-4_62
F. Vitale, S. R. Summerson, B. Aazhang, et al., ACS Nano 9, 4465 (2015). https://doi.org/10.1021/acsnano.5b01060
Y. Shin, S.-J. Song, S. Hong, et al., Nanomaterials 7 (11), 369 (2017). https://doi.org/10.3390/nano7110369
K. Zhou, G. A. Thouas, C. C. Bernard, et al., ACS Appl. Mater. Interfaces 4, 4524 (2012). https://doi.org/10.1021/am3007565
C. L. Weaver and X. T. Cui, Adv. Healthc. Mater. 4, 1408 (2015). https://doi.org/10.1002/adhm.201500056
S. Goenka, V. Sant, and S. Sant, J. Control. Release 173, 75 (2014). https://doi.org/10.1016/j.jconrel.2013.10.017
A. P. Hopper, J. M. Dugan, A. A. Gill, et al., Biomed. Mater. 9, 045009 (2014). https://doi.org/10.1088/1748-6041/9/4/045009
A. C. Taylor, C. H. González, P. Ferretti, and R. B. Jackman, Adv. Biosyst. 3, 1800299 (2019). .https://doi.org/10.1002/adbi.201800299
R. J. Edgington, A. Thalhammer, J. O. Welch, et al., J. Neural. Eng. 10, 056022 (2013). https://doi.org/10.1088/1741-2560/10/5/056022
A. Thalhammer, R. J. Edgington, L. A. Cingolani, et al., Biomaterials 31, 2097 (2010). https://doi.org/10.1016/j.biomaterials.2009.11.109
A. Voss, H. Wei, Y. Zhang, et al., Mater. Sci. Eng. C 64, 278 (2016). https://doi.org/10.1016/j.msec.2016.03.092
W. Tong, K. Fox, A. Zamani, et al., Biomaterials 104, 32 (2016). https://doi.org/10.1016/j.biomaterials.2016.07.006
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
We declare that we have no conflicts of interest.
Additional information
Translated by G. Levit
The original online version of this article was revised: Due to a retrospective Open Access order.
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bolshakova, O.I., Slobodina, A.D. & Sarantseva, S.V. Carbon Nanoparticles as Promising Neuroprotectors: Pro et Contra. II. Application of Carbon Nanoparticles in Neurobiology and Neurology. Nanotechnol Russia 17, 141–154 (2022). https://doi.org/10.1134/S2635167622020069
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S2635167622020069