
Abstract
Soil organic matter largely controls the ecosystem functions of soil and is a source of energy and a nutrient substrate for the soil microbial community as well. The postagrogenic dynamics of organic matter stock (content of organic carbon, Corg; total nitrogen, Ntot; and their ratio, C/N), basic soil properties, and indicators of soil microbial and enzyme activities are analyzed for the chronosequence of Retic Albic Podzol (southern taiga zone, Kostroma oblast, Russia). Different plots of the chronosequence have been withdrawn from agricultural use 11, 16, and 40 years ago. The stratification ratios (SRs) 0–5 : 5–10, 0–10 : 10–20, and 0–10 : 20–30 cm layers for Corg and Ntot contents are also estimated. A statistically significant increase in the contents of Corg and Ntot during the postagrogenic succession is observed in the topsoil layer of the old arable horizon (0–5 and 5–10 cm) as well as an increase in the stratification of their distribution, especially after a closed tree canopy has developed on the former arable land. Postagrogenic succession promotes an increase in the soil basal respiration rate and the content of microbial biomass, the maximum values of which are most often recordable in the soil under a 40-year-old secondary small-leaved forest. During the natural reforestation of the former arable lands, a statistically significant increase in the phosphatase activity is observed within the entire old arable horizon. The peaks of oxidoreductase activity correspond to the stages of succession with the maximum uptake of easily decomposable plant litter. Over the 40 years of postagrogenic succession, all analyzed parameters (except for pHKCl) have increased in a statistically significant manner at all considered depths of the old arable layer, with the highest annual increase in 0–5-cm layer, most enriched for organic matter.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
INTRODUCTION
The organic matter (OM) of soil is its vital component, which largely controls its ecosystem functions and acts as a source of energy and a nutrient substrate for the soil microbial community [18, 33, 35]. The amount and qualitative composition of soil OM are determined by the corresponding natural climatic zone, where these characteristics depend on the type of cenosis, land use, and agricultural load [64, 67, 79]. A change in the land use type also considerably influences the OM stock in soil [39, 59, 60]. In particular, the involvement of virgin soils in agricultural use causes inevitable losses in soil OM by mineralization and erosion [38, 65, 74, 75]. An opposite process, namely, cessation of agricultural use, first and foremost as tillage, leads to the restoration of natural vegetation and OM accumulation in the former arable layer [44, 47, 50, 51, 55, 58].
In the southern taiga zone, natural reforestation is observed during postagrogenic succession on the former farmlands [43, 45, 46, 53] and the ratio of main soil OM pools changes with the growth of tree stands [48, 62, 71, 72]. A considerable pool of weakly decomposed OM is formed thanks to the development of forest litter at the stage when closed canopy is formed [17, 44, 62]; this pool represents a large part of the total pool of soil carbon [43, 45, 76]. In addition, the litter structure becomes ever more complex with the progress of reforestation succession, thereby increasing the degree of spatial variation in the morphological characteristics of organogenic horizon [72, 73], and the biochemical composition of plant litter is cardinally changed [56]. In general, the balance between the mineralization and humification of soil OM changes during postagrogenic reforestation [9]. The OM redistribution along the profile is especially pronounced in the light soils; this appears as a decrease in the content and stock of organic carbon in the mineral part of the profile at the expense of their increase in the litter [43].
The change in the OM content and stock in the abandoned soils during natural reforestation in the southern taiga subzone is sufficiently well studied [10, 11, 45, 55, 61]. As has been shown, the carbon content in former arable soils can increase [17, 51, 55], decrease [12, 16, 58], or remain at the same level [42, 62] depending on the degree of their state of cultivation. Cessation of the agricultural use of soils in the forest zone also leads to an increase in their respiratory activity and the pool of microbial carbon [40, 54, 55, 57, 69, 78].
Although the microbial activity and enzyme pool of soil play an important role in the cycling of soil OM, the enzyme status of soils during postagrogenic evolution has been insufficiently studied so far. It is known that the changes in microbial and enzyme activities are observable long before the changes in other characteristics of soil quality [81]. Extracellular soil enzymes are responsible for oxidation and hydrolysis of polymeric and oligomeric organic compounds, implementing the balance between the decomposition of plant residues entering soil and formation of stable humic compounds [31, 79]. As is believed, hydrolytic enzymes trigger the initial stages in the destruction of fresh plant residues and are responsible for the cleavage of hydrolyzable compounds, such as carbohydrates (cellulose and chitin) and proteins [20, 31, 37, 77]. Extracellular hydrolytic enzymes catalyze most of the biochemical reactions in soil [70], cleaving high molecular weight compounds into monomers, which can be directly used by microorganisms [34]. The enzymes belonging to oxidoreductases (phenoloxidases and peroxidases) are involved in the mineralization of phenolic compounds by triggering the oxidation of complex aromatic structures of lignin and humic acids [23, 36, 41, 80]. The reactions of synthesis of specific humic acids can proceed concurrently with the oxidation and spontaneous condensation of phenolic substrates with different molecular weights [30, 80]. Thus, the activity of extracellular enzymes determines both the production rate of soluble organic compounds and the intensity of soil OM transformation.
The activity of different enzyme groups is an important characteristic in the function of soil microbial community [6, 14, 28]. A rapid response of the enzyme pool to various environmental changes [32, 78] makes enzyme activities a popular indicator of microbial activity and soil fertility [27, 29]. A few published data demonstrate that the invertase activity restored to almost background level in the soddy podzol of the Kostroma oblast (Russia) over 40 years of postagrogenic development versus, for example, cellulase activity, which did not restore [72]. The catalase activity in the soils of the chronosequence increased during the overall succession, which is most likely associated with a decrease in the soil density and improvement of aeration. A study of the postagrogenic gray forest soils [14] demonstrates a low activity of hydrolytic enzymes in young abandoned lands, while the hydrolase activities in the abandoned lands of 30 years old are comparable to the soil under a secondary forest. The oxidase activities were the highest in the soils of the “outermost” time stages of succession, namely, tillage and secondary forest cenosis. The dynamics as well as many aspects in the interaction of enzyme activity and other soil properties are to be explained. The study into the structural changes in the composition of enzyme pool will clarify the specific features in the transformation of soil OM during the self-restoration of soils after their agricultural use was stopped.
The goals of this work were (1) to study the postagrogenic dynamics of the contents of Corg and Ntot, C/N ratio, pH, and characteristics of microbial and enzyme activities in the Retic Albic Podzol of the Kostroma oblast (Russia); (2) to assess the stratification ratios (SRs) for Corg and Ntot contents within the old arable horizon; and (3) to clarify the relations of the microbial and enzyme pools of soils with other soil characteristics. We assume that the restoration of natural vegetation on the former arable soils, which entails the changes in the volume, availability, and biochemical diversity of plant litter, will lead to an increase in the OM stock in soils as well as the corresponding increase in the microbial and enzyme pools. The postagrogenic changes in the studied soil features within the former arable horizon will go on with different rates.
OBJECTS AND METHODS
Climate, vegetation, and soils in the study area. The study was performed in the Manturovo district (Kostroma oblast, Russia) in the chronosequence of abandoned lands of different ages (Table 1). The climate in the region is temperate continental; the average annual air temperature over the last 20 years is 3.6°C and average annual precipitation, 644 mm [1]. This territory belongs to the northeastern subregion of the Atlantic continental forest region [4]. The soil-forming conditions in the studied region are characteristic of the southern taiga, where the natural communities are represented by grass–dwarf shrub spruce stands [15]. Soddy podzolic soils (Albic Podzol) and podzols (Retic Albic Podzol) are the prevalent soil types [7]. The parental rock there is various glacial and fluvioglacial deposits. Most frequently, this rock is represented by binomial deposits or more lithologically complex inhomogeneous deposits although the upper 40–45 cm (including the former arable layer) are rather uniform and have a sandy loam particle size distribution [72].
The chronosequence of former arable plots with restoring vegetation comprises the following stages (Table 1): (1) tillage (stage 0) under the oat or perennial grasses, which is poorly cultivated and plowed without fertilizers for the last 25–30 years; (2) 11-year-old abandoned soil with meadow vegetation; (3) 16-year-old abandoned soil with tree stands of the goat willow and rich forb; (4) small-leaved aspen–spruce secondary forest with an age of 40 years; (5) and birch–spruce forest with an age of approximately 100 years (control).
The data on the history and duration of land use was kindly provided by the Ecological Station with the Severtsov Institute of Ecology and Evolution, Russian Academy of Sciences (Manturovo district, Kostroma oblast). The age of tree stands was determined using an increment borer. Starting from 2008, the plots have been monitored for the changes in plant composition and soil properties [72, 73, 76]. The vegetation was described and the biomass of grass (grass–dwarf shrub) layer was sampled on an almost annual basis; the weight of tree stand and its fractions was assessed allometrically [5]; the aboveground weight of grass–dwarf shrub horizon was assessed by cutting technique and of roots, by the technique of monoliths (in five replicates).
Soil sampling and assay of soil basic chemical properties. In each of the five plots constituting the chronosequence, mixed soil samples were taken from the depths of 0–5, 5–10, 10–20, 20–30 cm using an envelope technique. The soil bulk density (BD, g/cm3) was measured at the same depths using a Kachinsky borer [22]. Soil samples were air-dried and sieved (mesh, 2 mm). The roots and coarse organic material were manually removed. The contents of organic carbon (Corg) and total nitrogen (Ntot) were determined in a CHNS (Leco, United States) analyzer in triplicate. The Corg stock (ST-Corg, t C/ha) in assayed layer was calculated taking into account the Corg content (g C/kg soil), soil bulk density (g/cm3), and layer thickness (H, cm) as
The Corg stock in the 20-cm layer was calculated as the sum of ST-Corg in each of the constituent layers (0–5, 5–10, and 10–20 cm); pH was measured in 1 M KCl solution (soil to solution ratio, 1 : 2.5) in a Metler-Toledo (Switzerland) pH-meter in triplicate.
Analysis of soil microbial activity. Soil basal respiration (BR) was determined in triplicate according to the intensity of CO2 emission from soil after 7 days of preincubation at a humidity corresponding to 70–75% of their water holding capacity (WHC) and a temperature of 22–24°C [52]:
where C0 and C1 are the initial and final CO2 concentrations in vial, vol %; Vvial is the volume of vial, mL; t, incubation time, days; m, soil weighed sample; 12, molecular weight of carbon, g; and 22.4, molar volume, L.
The content of microbial carbon (Cmic) was assessed using the substrate-induced respiration technique [24] in the soil samples assayed for BR and supplemented with glucose solution (10 mg glucose per 1 g soil). The rate of the substrate-induced respiration (VSIR) was calculated using Eq. (2) and expressed in µL CO2/g soil h). The content of Cmic (mg C/kg soil) was computed as [24]:
The obtained microbial characteristics (BR and Cmic) and Corg content were used to assess the relative indices characterizing the function of soil microbial community and the OM availability to decomposition during postagrogenic evolution [40, 49, 66, 68]:
(1) Microbial metabolic coefficient (qCO2) or specific respiration of microbial biomass, which is the BR/Cmic ratio (mg CO2–C/g Cmic/day) and indicates the ecophysiological state of soil microbial community [26];
(2) The BR/Corg as the indicator of availability of soil Corg to decomposition [52, 53].
Analysis of soil enzyme activities. The oxidoreductase activities (polyphenol oxidase and peroxidase) were determined using the Karyagina–Mikhailova technique [20]. For this purpose, a soil sample (1 g) was supplemented with fresh 1% hydroquinone solution (10 mL) and 0.05% hydrogen peroxide solution (1 mL). The suspension was thoroughly mixed and placed into a thermostat for 30 min at +30°C. The mixture of hydroquinone and hydrogen peroxide solutions without soil was used as the control. After the incubation, the flasks were supplemented with ethanol (10 mL); the suspension was thoroughly mixed and filtered through double blue ribbon filter. The optical density of alcohol extracts (colored yellow) was measured at a wavelength of 450 nm in a UNICO 2080 (United States) spectrophotometer. The peroxidase (PO) activity was expressed in arbitrary units corresponding to the transformation of 1 g of benzoquinone (BQ) per 1 g soil over 30 min of incubation (mg BQ/g soil/30 min) at a temperature of 30°C [20].
Polyphenol oxidase (PPO) activity was determined analogously to PO activity but without adding hydrogen peroxidase since peroxidation of phenols requires the oxygen of air [10]; soil weighed sample, 2 g.
Phosphatase (PP) activity was determined according to Galstyan and Arutyunyan technique [20]. For this purpose, a soil weighed sample (1 g) was moistened to 90% of the water holding capacity, supplemented with fresh 1% sodium phenolphthalein phosphate (1 mL) and 1–2 drops of toluene; the suspension was thoroughly mixed and placed into a thermostat for 1 h at 30°C. After the incubation, the flasks were supplemented with distilled water (45 mL), potassium alum (1 mL), and 10% ammonium solution (2 mL). The suspension was thoroughly mixed and filtered through a blue ribbon filter. The optical density of the extracts, colored violet, was determined at a wavelength of 553 nm in a UNICO 2080 (United States) spectrophotometer (mg P2O5/100 g soil/h [21]).
Data processing. Statistical significance of the postagrogenic changes in all soil properties over 40 years was assessed using a linear regression model (y = ax + b), showing the dependence of soil properties (y, arithmetic means) on the age of abandoned soil (x). The arable land was assumed 0 years old and the forest cenosis was excluded from the analysis. The degree of differentiation of the old arable horizon in the Corg and Ntot contents after the forest stand has grown there was estimated using the stratification ratio (SR), which is the ratio of Corg (Ntot) content in different soil layers: SR(0–5 : 5–10), SR(0–10 : 10–20), and SR(0–10 : 20–30).
The correlation between the studied soil properties (arithmetic means) within the full chronosequence including the old-aged forest was assessed using Pearson’s correlation coefficient (F-test). The data were visualized using MS Excel 2013. Tables and plots show the means and standard errors (SEs). MS Excel 2013 software package and STATISTICA 6 were used for statistical analysis (α = 0.05).
RESULTS AND DISCUSSION
Changes in the ecosystem pools of carbon. During the restorative succession, the composition and structure of plant communities considerably change. In particular, individual trees appear approximately 7–8 years after cultivation is stopped [76] and a poorly closed tree stand of the goat willow appears after 15 years. Spruce undergrowth, potentially able to form spruce stands, appears 35–40 years of restorative succession in the secondary small-leaved forest. Thus, the biomass of tree stands increases during the postagrogenic restorative succession (Table 2) and reaches 138 t/ha in the 40-year-old forest. On the contrary, the biomass of grass and dwarf shrub layer decreases from 3.4 t/ha in the abandoned land of 11-year old to 0.50 t/ha in the abandoned land under 40-year-old forest stand to further increase to 0.92 t/ha in 100-year-old forest at the expense of a considerable share of dwarf shrubs. As for the 100-year-old forest, the spruce is prevalent; correspondingly, the share of needles and cones in the litter increases to 40% [19, 73], which cannot but be reflected in the rate of organic matter decomposition.
During the restorative succession, the annual stock of readily decomposable plant litter entering soil (tree leaves, aboveground parts of grass–dwarf shrub layer, and part of its root biomass) has two maximums. The first maximum is observed at the stage of meadow (at the expense of highly productive grass canopy) and the second is characteristic of the stage of small-leaved forest (thanks to leaf litter). With the development of tree stands, the litter stock and thickness increase so that its weight in an old-aged forest is almost tenfold higher as compared with a young aspen–birch stand. The ash content in the upper litter layer, which is an indirect characteristic of the mineralization rate of the input plant litter, almost halves during the succession. The Corg stock in the 0–20-cm layer is 1.5-fold higher as early as the stage of 16-year-old abandoned land and remains at the same level in the soil under a young forest. This suggests the restoration of the carbon sequestering function of soils at later stages of the postagrogenic development thanks to a gradual restoration of the natural vegetation. As for the soil of old-aged forest, the Corg stock in the 0–20-cm layer exceeds fourfold the corresponding stock in an arable soil. Thus, the capacity of biological turnover, indirectly determined by the volume of plant biomass, increases with the restoration of natural vegetation and the intensity of biological turnover, indirectly assessable from the ash content in the litter, decreases.
Postagrogenic dynamics of basic soil properties. During the postagrogenic succession, soil acidity increases in the upper part of the profile, which complies with the earlier data for forest soils [11, 13]. Within the old arable horizon (0–30 cm), the most pronounced gradient of postagrogenic acidification is observed in the upper (0–5 cm) layer (Fig. 1a), where pHKCl value decreases by almost unity as early as 10–11 years after plowing was stopped. The pHKCl values do not exceed 4.0 in the soil of forest succession stages, when the closed canopy of tree stands is formed; this is associated with a sharp decrease in the input of grass litter and development of forest litter, the decomposition products of which has an acidification effect on the soil. A decrease in pHKCl during 40-year postagrogenic succession displays a statistically significant linear trend (p < 0.05) at all depths (Table 3). The rate of a decrease in the pHKCl values is the highest in the 0–5-cm layer (–0.032 units/year) and is approximately the same (–0.023 to –0.025 units/year) in the remaining layers.

Postagrogenic dynamics of (a) pHKCl, (b) Corg content, (c) Ntot content, and (d) C/N ratio at different depths of the agrosoddy podzol arable horizon: 1, 0–5 cm; 2, 5–10 cm; 3, 10–20 cm; and 4, 20–30 cm.
The Corg content during the postagrogenic succession considerably increases, being especially pronounced in the upper part of the old arable horizon. In particular, the Corg content in the 0–5-cm layer over 11 years after abandonment mode doubles as compared with arable soil, increases fourfold at the stage of the secondary small-leaved forest, and sevenfold at the stage of the old-age birch–spruce forest. The change in the Corg content in the 0–5-cm layer is the most pronounced in the old-aged forest as compared with the small-leaved 40-year-old forest (Fig. 1b). As for the deeper layer of the old arable horizon (10–20 and 20–30 cm), an increase in the Corg content there changed only to the stage of 40-year-old small-leaved forest stand either remaining the same in the soil under the old-aged forest (10–20 cm) or decreasing (20–30 cm). The postagrogenic dynamics of Ntot content displays the trends similar to the above-described ones for the Corg content (Fig. 1c). As for the C/N ratio, it increases during the postagrogenic succession at almost all analyzed depths, suggesting a decrease in the intensity of soil OM decomposition especially at later stages of the succession, which correspond to the forest communities.
The linear trends of the changes in Corg content demonstrate that it increases in a statistically significant manner in the entire old arable horizon (except for the 20–30-cm layer) during the 40 years of postagrogenic succession. The highest rate of the increase (0.64 g C/kg soil per yr) is observable in the upper layer (0–5 cm). A statistically significant increase in the Ntot content is recorded in the 0–5- and 20–30-cm layers with linear regression coefficients of 0.038 and 0.019 g N/kg soil per yr, respectively. As a rule, the increase in Corg and Ntot contents in deeper layers becomes less pronounced during the restorative succession. A negative correlation of the Corg content, Ntot content, and C/N ratio with pHKCl is characteristic of the 0–5- and 5–10-cm layers (r = –0.75 to –0.90, p < 0.05) in the soils of the studied chronosequence (Table 4); this was also earlier reported for the soils of this chronosequence [55].
Changes in the Corg and Ntot stratification ratios during postagrogenic development. The Corg and Ntot are rather uniformly distributed in the studied 0–30-cm layer of arable plot and insignificantly change with depth (Figs. 1b, 1c). With the restoration of vegetation, the OM is considerably redistributed within the old arable layer. In particular, the Corg content at depths of 0–5 and 20–30 cm in the soil under the 11-year-old abandoned land differs twofold and in the soil under old-aged forest, more than fourfold. At the earlier stages of the postagrogenic succession, this is determined by separation of the soddy horizon and at the later stages, by formation of the organoprofile typical of the forest soils, in which a poorly humic upper part of the profile is almost immediately replaced by mineral horizons poor in OM.
Using stratification ratios (SRs), we quantitatively assessed the specific features in redistribution of the Corg and Ntot contents within the old arable horizon (Fig. 2). Their gradual stratification commences in the 0–10-cm layer: the SR (0–5 : 5–10) values in arable soil do not exceed 1.2 and amount to 1.8 in the soil under the secondary small-leaved forest. As for the soil of the old-aged birch–spruce stand, where the organic substances from the litter enter the uppermost part of the mineral profile, the SR (0–5 : 5–10) values reach 2.6. The SR (0–10 : 10–20) and SR (0–10 : 20–30) for Corg and Ntot change in a similar manner to the stage of the 40-year-old small-leaved forest. As for the soils under the old-aged forest, the SRs ratio of Corg and Ntot there for the 0–10-cm and for the deeper layers reach 6.0–8.1 and 6.3–9.5, respectively. This change in the OM distribution along the profile during the succession suggests a gradual degradation of the old arable horizon and development of the litter in the soils under forest stands formed during the restorative succession.

Changes in the stratification ratios (SRs) of the contents of (a) Corg and (b) Ntot in different layers of the old arable horizon during the postagrogenic development of agrosoddy podzols: 1, SR(0–5 : 5–10); 2, SR(0–10 : 10–20); and 3, SR(0–10 : 0–30).
The soil OM under natural vegetation is, as a rule, more stratified in the depth as compared with the arable soils; correspondingly, the SR values for Corg and Ntot increase during the natural reforestation, suggesting an improvement of the natural soil quality in terms of ecology [49, 61]. SRs are also frequently used to assess the carbon sequestration in soils [63, 82]. The earlier case study of two chronosequences of abandoned lands in the Kostroma oblast (Russia) using SR (0–10 : 20–30) has demonstrated a gradual differentiation of old arable soil according to the Corg content and stock, with its maximum in the soils of subclimax mixed forests with an age of 90–100 years [61]. In addition, a noticeable stratification of the upper part of old arable horizon during the natural reforestation is observed for all soil characteristics positively correlated with Corg content [55].
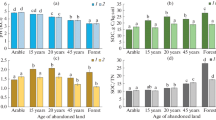
Postagrogenic dynamics of soil microbial activity characteristics. The characteristics of soil biological activity, such as the basal respiration and the content of microbial biomass, most rapidly respond to a change in the local ecological conditions [25], including the pattern of vegetation during the natural postagrogenic reforestation [51, 53, 56]. Next to an increase in the OM stock in soils during the postagrogenic succession, the Cmic content grows in a most pronounced manner in the 0–5-cm layer (Fig. 3a). Arable soil is characterized by the minimum Cmic content in all layers of the old arable horizon (140–180 mg C/kg soil), where the root biomass remaining after crop harvesting is the sole source of OM. Consequently, the fresh readily decomposable plant material is constantly in deficiency, thereby influencing the state of microbial community. The Cmic in the 0–5-cm layer increases more than twofold in 11-year-old abandoned land due to the forming a meadow cenosis with rich forb. Content Cmic reaches a maximum value (936 mg C/kg soil) after 40 years at the stage of the small-leaved forest. Presumably, this is determined by a considerable increase in the stock of input readily decomposable plant litter with a relatively high content of ash elements (Table 2). However, the acidity of the plant litter and the upper soil level is not high enough to inhibit the activity of soil microbiota. It is the acid pH of the upper soil layer and needle–leaf litter in the old-aged mixed forest that caused a drastic decrease in the Cmic content in the 0–5-cm layer (to 245 mg C/kg soil). A similar pattern of the postagrogenic Cmic dynamics is also observed in the other layers of the old arable horizon; however, the pattern is leveled with depth.

Postagrogenic dynamics of (a) microbial carbon content, Cmic; (b) rate of basal respiration, BR; (c) BR/Corg ratio; and (d) metabolic coefficient qCO2 at different depths of the agrosoddy podzol arable horizon: 1, 0–5 cm; 2, 5–10 cm; 3, 10–20 cm; and 4, 20–30 cm.
The postagrogenic dynamics of BR rate in all layers of the old arable horizon in general repeats the dynamics of Cmic content (Fig. 3b): the minimum BR values are observed in the arable plot; a threefold increase, at the meadow stage; and the maximum BR value, at the secondary small-leaved forest. The most pronounced changes in BR rate are recorded in soils of different succession stages in the upper soil layer (0–10 cm).
The linear trends of the changes in microbial characteristics demonstrate a statistically significant increase in the Cmic in the 0–5-cm layer over 40 years of postagrogenic succession and in the BR rate, at all depths of the old arable layer except for the 10–20-cm layer. The most pronounced increase in these characteristics is observable in the 0–5-cm layer; it amounts to 18.51 mg C/(kg soil year) for Cmic and 1.48 mg C/(kg soil day year) for BR rate (Table 5), while the duration of postagrogenic restoration explains 97–99% of the variance of Cmic content and BR rate, respectively. The increase in Cmic content and BR value in the 5–10-cm layer decreases 4.4- and 3-fold, respectively. In the 0–5- and 5–10-cm layers, the BR rate and Cmic content are correlated in a statistically significant manner (r = 0.94, p < 0.05; Table 4); this pattern has been observed during postagrogenic soil evolution in other bioclimatic zones as well [51, 57].
BR/Corg ratio may act as an indicator of soil OM availability for decomposition [51, 52]. Analysis of the BR/Corg postagrogenic dynamics demonstrates that this characteristic increases two- to fourfold in the upper part of the old arable horizon (0–5- and 5–10-cm layers) during the first 40 years of the restorative succession (Fig. 3c). This suggests that the availability of the OM freshly formed during the postagrogenic succession increases at the expense of the input readily decomposable plant litter. Consequently, the OM stability decreases after the cultivation is stopped, which has been observed for the soils of the studied chronosequence as well as earlier [55]. In the soil of mixed old-aged forest, the BR/Corg ratio (0–10-cm layer) is threefold lower as compared with arable soil; presumably, this is explainable with the fact that the OM formed of coniferous–deciduous litter is much less decomposable. These patterns in general correspond to the earlier data on the chronosequence of postagrogenic gray forest soils [51, 53].
The linear trends in the BR/Corg change over the 40 years of postagrogenic succession demonstrate a statistically significant increase only in the 0–5- and 5–10-cm layers, while the duration of postagrogenic restoration explains 93–96% of the variance of this parameter (Table 5). Quite expectedly, the BR/Corg ratio and Cmic content are correlated in a statistically significant manner in the 0–5- and 5–10-cm layers (r = 0.82, p < 0.05; Table 4).
Microbial metabolic coefficient qCO2, which is the ratio of BR rate to Cmic content, indicates the degree of environmental enrichment with the substrate necessary for the life activities of microbiota, i.e., this ratio demonstrates the availability of OM to soil microorganisms [3, 40]. The lowest qCO2 values (0.13–0.16 mg CO2/(mg Cmic day)) are characteristic of arable, where the microbial community suffers a deficiency in the nutrient substrate because of a very small input of plant litter (Table 3). During the 40-year restorative succession of former arable soil to the stage of small-leaved forest, the qCO2 value gradually increases in all soil layers of the old arable horizon (Fig. 3d). Although the qCO2 values within the 0–30-cm layer are as a rule close, the coefficient is higher at a depth of 5–10 cm in the soils under 16-year-old abandoned land transitional to forest and secondary 40-year-old forest stand (0.28–0.30 mg CO2/(mg Cmic day)). This is associated not only with an increase in the input of plant litter stock, available to rapid microbial decomposition, but also with its high biochemical diversity (tree leaves and grasses of various biomorphological groups). The higher qCO2 values in the 5–10-cm layer and deeper at the stage of rich grass cover are also explainable with the presence of readily available root mortmass in these layers. The maximum qCO2 value of 0.46 mg CO2/(mg Cmic day) is recorded in the 0–5-cm layer of the soil under the old-aged forest most likely because of a higher input of the plant litter containing the compounds with different degrees of availability. As for the 0–10-cm layer, the qCO2 values there are positively correlated with the Corg and Ntot contents and C/N ratio as well as negatively correlated with pHKCl (r = –0.74, p < 0.05; Table 4). Thus, a relatively high OM content and sufficient input of the plant material available for decomposition at the middle stages of succession enhance the favorable ecophysiological state of microbial communities and their active function during the postagrogenic succession.
Postagrogenic dynamics of soil enzyme activity. Enzyme activities are also a sensitive indicator of the postagrogenic changes induced by removal of agricultural load [14, 56]. The hydrolytic enzyme activities, including phosphatases, are largely determined by the OM stock [20]; correspondingly, the PP activity is rather low in arable soil and the young abandoned land (Fig. 4a). The most pronounced increase in PP activity during the postagrogenic evolution is observed in the 0–5-cm layer at the stage of the small-leaved forest; this activity reaches its maximum in the old-aged forest. This pattern of the change in PP activity is retained in the 5–10-cm layer but at a lower level. As for the lower part of the old arable layer (10–30 cm) with its rather low Corg content, the PP activity is considerably decreased. A tight positive correlation of PP activity with the Corg and Ntot content (r = 0.99, p < 0.05) as well as with qCO2 (r = 0.83, p < 0.05; Table 4) is observed in the 0–10-cm layer. On the other hand, the PP activity is negatively correlated with pH (r = –0.73, p < 0.05). The linear trends of the changes in PP activity over the 40 years of postagrogenic succession demonstrate a statistically significant increase at all depths of the old arable horizon (Table 5). The annual increase in PP activity in the 0–5-cm layer is almost doubled as compared with the other layers of the old arable horizon and the duration of postagrogenic restoration explains 90–98% of the variance of this parameter.

Postagrogenic dynamics of enzyme activities: (a) phosphatase, PP; (b) polyphenol oxidase, PPO; (c) peroxidase, PO; and (d) PPO/PO ratio.
The postagrogenic dynamics of oxidoreductases differs from that of PP because these enzyme activities are primarily associated with OM transformation, namely, the balance between OM mineralization and humification. With the restoration of vegetation, the PO activity in the 0–5-cm layer gradually increases to reach its maximum in the soils of secondary small-leaved and old-aged mixed forests (Fig. 4c). During the postagrogenic evolution, the PO activity in the deeper layers of old arable horizon also displays the general trend of an increase but sharply decreases in the soil under the old-aged forest. This can be associated with that OM almost does not accumulate in the deeper podzol layers formed under the old-aged forest. A lower PO activity in the arable soil as compared with abandoned lands is explained with that the OM entering the soil with crop residues is evidently insufficient for active mineralization processes. Most likely, the OM stock having accumulated by the stage of 40-year-old small-leaved forest is sufficient for a sharp increase in the PO activity. A positive correlations of PO activity with the BR rate, qCO2 value, and PP activity are observed in the 0–5- and 5–10-cm layers (r = 0.78, p < 0.05; Table 4). These correlations are explainable with the fact that the changes in the above-mentioned parameters are determined by the common factor, most likely, the amount of readily decomposed substrate entering soil.
The postagrogenic dynamics of PPO activity is more complex as compared with PO although they are rather tightly correlated (r = 0.78, p < 0.05; Table 4). During the restorative succession, two PPO activity peaks are observed in the 0–5- and 5–10-cm layers, namely, at the stages of young highly productive meadow (11 years) and small-leaved forest (40 years). A decrease in PPO activity in the soil of 16-year-old abandoned land is most likely determined by a certain time gap between the highly productive grass phytocenosis and forest phytocenosis, which creates a certain deficiency in the input of readily decomposable substances with the plant litter and decreases their transformation activity. In addition, the phenolic compounds contained in willow leaves are able to inhibit plant residues decomposition [55].
The linear trends of the changes in PO and PPO activities over the 40 years of postagrogenic succession demonstrate a statistically significant growth in these parameters in all layers of the old arable horizon except for the 10–20-cm layer for PPO activity and 5–10- and 20–30-cm layers for PO activity (Table 5). The rate of increase in PO activity is the highest in the 0–5-cm layer, while the PPO activity displays a similar rate of increase over the entire old arable horizon.
Some researchers [2, 8] believe that the ratio of PPO to PO activities (PPO/PO) indicated the degree of prevalence of the humification over mineralization processes, that is, it can be used as a humification coefficient. Any distinct trend of this characteristic is unobservable in the studied chronosequence (Fig. 4d); however, the PPO/PO coefficient at a depth of 0–5 cm has the minimum value (0.51) at the stage of 16-year-old abandoned land and the maximum (1.31), at the stage of 40-year-old forest. As for the 5–10-cm layer, the minimum PPO/PO value corresponds to arable soil and the maximum, (2.05), to 11-year-old abandoned land. The PPO/PO ratio in both layers of the old-aged forest is approximately 1.0. This suggests that humification processes are more pronounced as compared with mineralization when the input of plant litter is insufficient, which is characteristic of arable soil when only crop afterharvest residues is the sole substrate for decomposition. The maximum values of PPO/PO ratio are observable in the 5–10- and 10–20-cm layers at the stage of highly productive meadow owing to a more active humification of the underground fraction of grass biomass, which is maximal at this stage (Table 2). The PPO/PO value is one of the lowest in the soils under 16-year-old abandoned land with the tree stand of the goat willow and well-developed grass layer (Fig. 4d). We have earlier observed somewhat similar patterns using HIX1 value estimated with the help of the fluorescence spectra of soil water extracts as the humification index [55]. As has been shown, the humification index of water-soluble OM during natural reforestation decreases to its lowest values at the stage of goat willow stands, the leaf litter of which is rich in phenolic compounds, able to inhibit an active transformation of organic substrates. Our data suggest that the PPO/PO ratio is rather problematic and dubious as an indicator of humification. Other instrumental tools, such as solid-phase 13C-NMR spectroscopy, are required for a more adequate assessment of the degree of transformation of soil OM during postagrogenic development.
We used the principle component method to integrate and visualize our results on the changes in a wide range of properties at four different depths of the old arable horizon in five plots constituting the chronosequence of abandoned lands. The overall set of the studied properties distinctly distinguishes the old-aged mixed forest from the secondary small-leaved forest and the arable soil from the 16-year-old abandoned land (Fig. 5). The youngest abandoned land of the chronosequence (11-year-old) is in an intermediate position between the arable and 16-year-old abandoned land. Factor 1, which correlates best with Corg content (r = –0.91), Ntot content (r = –0.90), PP activity (r = –0.93), and pH (r = 0.76), describes 55.0% of the total information and the ordinate distinctly distinguishes the soils of forest plots from the soils of arable and younger abandoned land with the prevalence of meadow vegetation. Factor 2, which most tightly correlates with Corg content (r = –0.79), is responsible for only 19.6%. Thus, the OM content gradually increases within the entire old arable horizon during the restorative succession of abandoned lands in the southern taiga zone. Consequently, the microbial and enzyme activities increase as well, which is most pronounced in the upper layer (0–10 cm) of the soil under the old-aged forest, representing the subclimax stage of postagrogenic reforestation, suggesting that polysaccharide soils may require several tens of years to reach a dynamic equilibrium.

Distribution of the studied soils according to the principle component technique using the values of different soil characteristics at different depths: 1, 0–5 cm; 2, 5–10 cm; 3, 10–20 cm; and 4, 20–30 cm. Properties: pHKCl, potential soil acidity; Corg, the content of organic carbon (g C/kg soil); Ntot, the content of total nitrogen content; Cmic, carbon content in microbial biomass; and PP, phosphatase activity (mg P2O5/(100 g soil h)).
CONCLUSIONS
Natural reforestation of former arable soils in the southern taiga subzone is accompanied by an increase in the input of plant residues, leading to a considerable increase in the content of organic carbon and total nitrogen in the upper part of the old arable horizon with concurrent decrease in pHKCl. Alongside, the stratification of the Corg and Ntot contents in the 0–30-cm layer considerably increases during the postagrogenic succession, which is especially pronounced at the stage of closed tree canopy; this suggests that the carbon-sequestering function of soil restores at the later stages of postagrogenic development. The highest stratification ratios, SRs (0–5 : 5–10, 0–5 : 10–20, and 0–5 : 20–30), are observed in the soil under old-aged forest, which principally differs in the structure of its organoprofile.
The soil enrichment with OM induces a considerable increase in the microbial activity, appearing as an increase in the basal respiration rate and the content of microbial biomass. The most pronounced changes in these characteristics during the natural restorative succession are observed in the upper soil layer (0–10 cm) and their maximum values, in the soil under the 40‑year-old abandoned land, which corresponds to small-leaved forest stand, where a readily decomposed plant litter provides the presence of a large stock of available substrate for microbial community. The microbial metabolic coefficient also gradually increases during the 40 years of restorative succession of former arable soils, which is associated not only with an increase in the input plant litter available for rapid microbial decomposition, but also with its high biochemical diversity. The ratio of basal respiration to total carbon content, which characterizes the OM availability for microbial decomposition, in the upper layer (0–10 cm) of the former arable horizon during the first 40 years of the restorative succession increases two–fourfold and is tightly correlated with the Cmic content.
Soil enzymes, both of hydrolytic group and oxidoreductases, play an important role in the OM transformation. Phosphatase activity increases manifold during the postagrogenic restoration, especially in the upper layer (0–10 cm) of arable horizon; first and foremost, this is associated with an increase in the OM content and is confirmed by a tight positive correlation between these parameters. The oxidoreductase activities do not display any distinct trends during the postagrogenic succession; however, the peaks of these activities, observable at the stages of highly productive meadow (peroxidase) and small-leaved forest (phenoloxidase), may result from an increased input of readily decomposed plant litter of different biochemical compositions (grass or small leaves).
Over the 40 years of postagrogenic succession, almost all analyzed characteristics display the most pronounced increase in the 0–5-cm layer. The changes in all analyzed properties during the restorative succession are less pronounced in the deeper layers. Thus, our hypotheses formulated at the beginning of the paper are fully confirmed.
REFERENCES
Climate data archive. http://climatbase.ru/. Cited June 4, 2020.
Z. Z. Ayupov, L. V. Sidorova, N. S. Anokhina, I. F. Minnebaeva, and N. I. Gareev, “Organic matter and enzymatic activity of leached chernozem depending on the tillage method and fertilizer,” Vestn. Bashkir. Gos. Agrar. Univ., No. 2, 11–16 (2010).
E. V. Blagodatskaya, N. D. Anan’eva, and T. N. Myakshina, “Characterization of the microbial community using the metabolic coefficient,” Pochvovedenie, No. 2, 205–210 (1995).
S. I. Bolysov and Yu. N. Fuzeina, “Physical and geographical conditions of the Kostroma Volga region: geological and geomorphologic structure,” in Kostroma Volga Region: Nature and Man (Moscow, 2001), pp. 36–60.
D. G. Zamolodchikov, A. I. Utkin, and G. N. Korovin, “Determination of carbon stocks by age-dependent plantation using conversion-volume coefficients,” Lesovedenie, No. 3, 84–93 (1998).
A. E. Ivanova, V. V. Nikolaeva, and O. E. Marfenina, “Changes in the cellulolytic activity of urban soils induced by the removal of plant litter (using Moscow as an example),” Eurasian Soil Sci. 48, 501–508 (2015).
L. L. Shishov, V. D. Tonkonogov, I. I. Lebedeva, and M. I. Gerasimova, Classification and Diagnostic System of Russian Soils (Oikumena, Smolensk, 2004) [in Russian].
M. V. Kovalenko and G. K. Markovskaya, “Influence of soil basic tillage on its enzymatic activity,” Vestn. Kazan. Gos. Agrar. Univ., No. 1 (27), 108–111 (2013).
A. V. Litvinovich, O. Yu. Pavlova, and I. A. Plylova, “Changes of the composition of the vegetation cover, content and reserves of humus in cultivated soddy-podzolic gleyic loamy soil during post-agrogenic evolution,” Izv. S.-Peterb. Gos. Agrar. Univ., No. 22, 74–77 (2011).
A. V. Litvinovich, O. Yu. Pavlova, and D. V. Chernov, “Change of humus content in soddy-podzolic soil after anthropogenic impact,” Dokl. Ross. Akad. S-kh. Nauk, No. 6, 26–28 (2002).
A. V. Litvinovich and I. A. Plylova, “Change of acid-base properties of soddy-podzolic loamy soil during postagrogenic evolution,” in Scientific Basis for Development of Agroindustrial Complex under Reformation (St. Petersburg, 2009), pp. 160–164.
D. I. Lyuri, S. V. Goryachkin, N. A. Karavaeva, E. A. Denisenko, and T. G. Nefedova, Dynamics of Agricultural Lands of Russia in the 20th Century and Postagrogenic Restoration of Vegetation and Soils (GEOS, Moscow, 2010) [in Russian].
N. N. Matinyan, K. A. Bakhmatova, and S. S. Alekseev, “Postagrogenic transformation of soils formed on rocks with contrasting granulometric composition,” in Humus and Pedogenesis (St. Petersburg, 2007), pp. 52–60.
L. A. Ovsepyan, I. N. Kurganova, A. S. Mostovaya, V. O. Lopes de Gerenyu, V. I. Lichko, E. V. Blagodatskaya, and Ya. V. Kuzyakov, “Enzymatic activity of postagrogenic gray forest soils in Les na Vorskle upland oak forest,” Izv. Samar. Nauchn. Tsentra, Ross. Akad. Nauk 19 (2), 151–158 (2017).
G. N. Ogureeva, Botanical-Geographic Zonation of the USSR (Moscow State Univ., Moscow, 1991) [in Russian].
A. A. Romanovskaya, V. N. Korotkov, R. T. Karaban’, and N. S. Smirnov, “Dynamics of carbon balance components in fallow arable lands on the Valdai Upland,” Russ. J. Ecol. 43, 373–377 (2012). https://doi.org/10.1134/S106741361204011X
I. M. Ryzhova, A. A. Erokhova, and M. A. Podvezennaya, “Dynamics of carbon reserves in postagrogenic ecosystems as a result of afforestation in Kostroma oblast,” Lesovedenie, No. 4, 307–317 (2015).
V. M. Semenov and B. M. Kogut, Soil Organic Matter (GEOS, Moscow, 2015) [in Russian].
V. M. Telesnina, L. G. Bogatyrev, A. I. Benediktova, Ph. I. Zemskov, and M. N. Maslov, “The dynamics of plant debris input and of some properties of forest litters during postagrogenic reforestation under the conditions of southern taiga,” Moscow Univ. Soil Sci. Bull. 74, 139–145 (2019).
F. Kh. Khaziev, Methods of Soil Enzymology (Nauka, Moscow, 2005) [in Russian].
T. E. Khomutova, T. S. Demkina, N. N. Kashirskaya, and V. A. Demkin, “Phosphatase activity in the surface and buried chestnut soils of the Volga-Don interfluve,” Eurasian Soil Sci. 45, 423–428 (2012).
Theory and Methods of Soil Physics, Ed. by E. V. Shein and L. O. Karpachevskii (Grif i K, Moscow, 2007) [in Russian].
A. V. Yakushev, I. N. Kuznetsova, E. V. Blagodatskaya, and S. A. Blagodatsky, “Temperature dependence of the activity of polyphenol peroxidases and polyphenol oxidases in modern and buried soils,” Eurasian Soil Sci. 47, 459–465 (2014).
J. P. E. Anderson and K. H. Domsch, “A physiological method for the quantitative measurement of microbial biomass in soils,” Soil Biol. Biochem. 10 (3), 215–221 (1978).
T.-H. Anderson, “Physiological analysis of microbial communities in soil: applications and limitations,” in Beyond the Biomass: Compositional and Functional Analysis of Soil Microbial Communities (Wiley, Chichester, 1994), pp. 67–76.
T.-H. Anderson and K. H. Domsch, “The metabolic quotient for CO2 (qCO2) as a specific activity parameter to assess the effects of environmental conditions, such as pH, on the microbial biomass of forest soils,” Soils Biol. Biochem. 25, 393–395 (1993). https://doi.org/10.1016/0038-0717(93)90140-7
H. Aponte, J. Medina, B. Butler, S. Meier, P. Cornejo, and Y. Kuzyakov, “Soil quality indices for metal(loid) contamination: an enzymatic perspective,” Land Degrad. Dev. 31, 2700–2719 (2020).
M. A. Azarenko, K. Sh. Kazeev, O. Yu. Yermolaeva, and S. I. Kolesnikov, “Changes in the plant cover and biological properties of chernozems in the postagrogenic period,” Eurasian Soil Sci. 53, 1645–1654 (2020). https://doi.org/10.1134/S1064229320110034
N. N. Y. Badiane, J. L. Chotte, E. Pate, D. Masse, and C. Rouland, “Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semiarid tropical regions,” Appl. Soil Ecol. 18, 229–238 (2001).
J. M. Bollag, J. Dec, and P. M. Huang, “Formation mechanisms of complex organic structures in soil habitats,” Adv. Agron. 63, 237–265 (1998).
R. G. Burns, J. L. DeForest, J. Marxsen, R. L. Sinsabaugh, M. E. Stromberger, M. D. Wallenstein, M. N. Weintraub, and A. Zoppini, “Soil enzymes in a changing environment: current knowledge and future directions,” Soil Biol. Biochem. 58, 216–234 (2013).
R. P. Dick, D. D. Myrold, and E. A. Kerle, “Microbial biomass and soil enzyme activities in compacted and rehabilitated skid trail soils,” Soil Sci. Soc. Am. J. 52, 512–516 (1988). https://doi.org/10.2136/sssaj1988.03615995005200020038x
S. Duarte-Guardia, P. Peri, W. Amelung, E. Thomas, N. Borchard, G. Baldi, A. Cowie, and B. Ladd, “Biophysical and socioeconomic factors influencing soil carbon stocks: a global assessment,” Mitigation Adapt. Strategies Global Change 25 (6), (2020). https://doi.org/:10.1007/s11027-020-09926-1
F. Eivazi and M. A. Tabatabai, “Factors affecting glucosidase and galactosidase activities in soils,” Soil Biol. Biochem. 22, 891–897 (1990). https://doi.org/10.1016/0038-0717(90)90126-K
U. Falkengren-Grerup, D.-J. ten Brink, and J. Brunet, “Land use effects on soil N, P, C and pH persist over 40–80 years of forest growth on agricultural soils,” For. Ecol. Manage. 225 (1–3), 74–81 (2006). https://doi.org/10.1016/j.foreco.2005.12.027
C. Floch, E. Alarcon-Gutiérrez, and S. Criquet, “ABTS assay of phenol oxidase activity in soil,” J. Microbiol. Methods 71, 319–324 (2007).
D. P. German, K. R. B. Marcelo, M. M. Stone, and S. D. Allison, “The Michaelis–Menten kinetics of soil extracellular enzymes in response to temperature: a cross-latitudinal study,” Global Change Biol. 18, 1468–1469 (2012).
T. Guillaume, D. Muhammad, and Y. Kuzyakov, “Losses of soil carbon by converting tropical forest to plantations: erosion and decomposition estimated by δ13C,” Global Change Biol. 21, 3548–3560 (2015).
L. B. Guo and R. M. Gifford, “Soil carbon stocks and land use change: a metaanalysis,” Global Change Biol. 8, 345–360 (2002). https://doi.org/10.1046/j.1354-1013.2002.00486x
K. V. Ivashchenko, N. D. Ananyeva, V. I. Vasenev, V. N. Kudeyarov, and R. Valentini, “Biomass and respiration activity of soil microorganisms in anthropogenically transformed ecosystems (Moscow region),” Eurasian J. Soil Sci. 47, 892–903 (2014). https://doi.org/10.1134/S1064229314090051
A. R. Johnsen and O. S. Jacobsen, “A quick and sensitive method for the quantification of peroxidase activity of organic surface soil from forests,” Soil Biol. Biochem. 40, 814–821 (2008).
O. Kalinina, O. Chertov, A. V. Dolgikh, S. V. Goryachkin, D. I. Lyuri, S. Vormstein, and L. Giani, “Self-restoration of post-agrogenic albeluvisols: soil development, carbon stocks and dynamics of carbon pools,” Geoderma 207–208, 221–233 (2013). https://doi.org/10.1016/j.geoderma.2013.05.019
O. Kalinina, S. V. Goryachkin, N. A. Karavaeva, D. I. Lyuri, and L. Giani, “Dynamics of carbon pools in post-agrogenic sandy soils of southern taiga of Russia,” Carbon Balance Manage. 5 (1), 1–9 (2010). https://doi.org/10.1186/1750-0680-5-1
O. Kalinina, S. V. Goryachkin, D. I. Lyuri, and L. Giani, “Post-agrogenic development of vegetation, soils, and carbon stocks under self-restoration in different climatic zones of European Russia,” Catena 129, 18–29 (2015). https://doi.org/10.1016/j.catena.2015.02.016
O. Kalinina, S. V. Goryachkin, N. A. Karavaeva, D. I. Lyuri, L. Najdenko, and L. Giani, “Self-restoration of post-agrogenic sandy soils in the southern taiga of Russia: soil development, nutrient status, and carbon dynamics,” Geoderma 152, 35–42 (2009). https://doi.org/10.1016/j.geoderma.2009.05.014
O. Yu. Kalinina, O. Chertov, P. Frolov, S. Goryachkin, P. Kuner, J. Küper, and L. Giani, “Alteration process during the post-agricultural restoration of luvisols of the temperate broad-leaved forest in Russia,” Catena 171, 602–612 (2018). https://doi.org/10.1016/j.catena.2018.08.004
I. Kämpf, N. Hölzel, M. Störrle, G. Broll, and K. Kiehl, “Potential of temperate agricultural soils for carbon sequestration: a meta-analysis of land-use effects,” Sci. Total Environ. 566–567, 428–435 (2016). https://doi.org/10.1016/j.scitotenv.2016.05.067
I. O. Kechaikina, A. G. Ryumin, and S. N. Chukov, “Postagrogenic transformation of organic matter in soddy-podzolic soils,” Eurasian Soil Sci. 44, 1077–1089 (2011). https://doi.org/10.1134/S1064229311100061
Y. Kooch, S. Ehsanib, and M. Akbariniac, “Stratification of soil organic matter and biota dynamics in natural and anthropogenic ecosystems,” Soil Tillage Res. 200, 104621 (2020). https://doi.org/10.1016/j.still.2020.104621
I. Kurganova, V. Lopes de Gerenyu, Y. Kuzyakov, and J. Six, “Carbon cost of collective farming collapse in Russia,” Global Change Biol. 20 (3), 938–947 (2014). https://doi.org/10.1111/gcb.12379
I. Kurganova, A. Merino, V. Lopes de Gerenyu, N. Barros, O. Kalinina, L. Giani, and Y. Kuzyakov, “Climate dependent mechanisms of carbon sequestration and stabilization by restoration of arable soils after abandonment,” Geoderma 354, 113882 (2019). https://doi.org/10.1016/j.geoderma.2019.113882
I. N. Kurganova, V. O. Lopes de Gerenyu, J. F. Gallardo Lancho, and C. T. Oehm, “Evaluation of the rates of soil organic matter mineralization in forest ecosystems of temperate continental, Mediterranean, and tropical monsoon climates,” Eurasian Soil Sci. 45, 68–79 (2012). https://doi.org/10.1134/S1064229312010085
I. N. Kurganova, V. O. Lopes de Gerenyu, A. S. Mostovaya, L. A. Ovsepyan, V. M. Telesnina, V. I. Lichko, and Yu. I. Baeva, “Effect of reforestation on microbiological activity of postagrogenic soils in European Russia,” Contemp. Probl. Ecol. 11, 704–718 (2018). https://doi.org/10.1134/S1995425518070089
I. N. Kurganova, V. M. Semenov, and V. N. Kudeyarov, “Climate and land use as key factors of the stability of organic matter in soils,” Dokl. Biol. Sci. 489, 189–192 (2019).
I. N. Kurganova, V. M. Telesnina, V. O. Lopes de Gerenyu, V. I. Lichko, and E. I. Karavanova, “The dynamics of carbon pools and biological activity of retic albic podzols in southern taiga during the postagrogenic evolution,” Eurasian Soil Sci. 54, 337–351 (2021). https://doi.org/10.1134/S1064229321030108
L. Ovsepyan, I. Kurganova, V. Lopes de Gerenyu, and Ya. Kuzyakov, “Conversion of cropland to natural vegetation boosts microbial and enzyme activities in soil,” Sci. Total Environ. 743, 140829 (2020). https://doi.org/10.1016/j.scitotenv.2020.140829
L. Ovsepyan, I. Kurganova, V. Lopes de Gerenyu, and Ya. Kuzyakov, “Recovery of organic matter and microbial biomass after abandonment of degraded agricultural soils: the influence of climate,” Land Degrad. Dev. 30, 1861–1874 (2019). https://doi.org/10.1002/ldr.3387
L. A. Ovsepyan, I. N. Kurganova, V. O. Lopes de Gerenyu, A. V. Rusakov, and Ya. V. Kuzyakov, “Changes in the fractional composition of organic matter in the soils of the forest-steppe zone during their postagrogenic evolution,” Eurasian Soil Sci. 53, 50–61 (2020). https://doi.org/10.1134/S1064229320010123
C. Poeplau, A. Don, L. Vesterdal, J. Leifeld, B. van Wesemael, J. Schumacher, and A. Gensior, “Temporal dynamics of soil organic carbon after land-use change in the temperate zone—carbon response functions as a model approach: soil organic carbon and land-use change,” Global Change Biol. 17, 2415–2427 (2011). https://doi.org/10.1111/j.1365-2486.2011.02408.x
W. M. Post and K. C. Kwon, “Soil carbon sequestration and land-use change: processes and potential,” Global Change Biol. 6, 317–328 (2000). https://doi.org/10.1046/j.1365-2486.2000.00308.x
I. M. Ryzhova, V. M. Telesnina, and A. A. Sitnikova, “Dynamic of soil properties and carbon stock structure in postagrogenic ecosystems of Southern taiga during natural reforestation,” Eurasian Soil Sci. 53, 230–243 (2020). https://doi.org/10.1134/S0032180X20020100
I. M. Ryzhova, A. A. Erokhova, and M. A. Podvezennaya, “Dynamics and structure of carbon storage in the postagrogenic ecosystems of the southern taiga,” Eurasian Soil Sci. 47, 1207–1215 (2014). https://doi.org/10.1134/S1064229314090117
J. C. M. Sá, and R. Lal, “Stratification ratio of soil organic matter pools as an indicator of carbon sequestration in a tillage chronosequence on a Brazilian oxisol,” Soil Tillage Res. 103 (1), 46–56 (2009). https://doi.org/10.1016/j.still.2008.09.003
P. Smith, “Land use change and soil organic carbon dynamics,” Nutr. Cycl. Agroecosyst. 81, 169–178 (2008). https://doi.org/10.1007/s10705-007-9138-y
J.-F. Soussana, P. Loiseau, N. Vuichard, E. Ceschia, J. Balesdent, T. Chevallier, and D. Arrouays, “Carbon cycling and sequestration opportunities in temperate grasslands,” Soil Use Manage. 20, 219–230 (2004).
M. Spohn, T.J. Novák, J. Incze, and L. Giani, “Dynamics of soil carbon, nitrogen, and phosphorus in calcareous soils after land-use abandonment—A chronosequence study,” Plant Soil 401 (1–2), 185–196 (2016). https://doi.org/10.1007/s11104-015-2513-6
U. Stockmann, J. Padarian, A. McBratney, B. Minasny, D. de Brogniez, L. Montanarella, S. Y. Hong, B. G. Rawlins, and D. J. Field, “Global soil organic carbon assessment,” Global Food Sec. 6, 9–16 (2015). https://doi.org/10.1016/j.gfs.2015.07.001
E. V. Stolnikova, N. D. Ananyeva, and O. V. Chernova, “The microbial biomass and its activity and structure in the soils of old forests in the European Russia,” Eurasian Soil Sci. 44, 437–452 (2011). https://doi.org/10.1134/S1064229311040107
E. A. Susyan, S. Wirth, N. D. Ananyeva, and E. V. Stolnikova, “Forest succession on abandoned arable soils in European Russia—Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity,” Eur. J. Soil Biol. 47, 169–174 (2011). https://doi.org/10.1016/j.ejsobi.2011.04.002
M. J. Swift, O. W. Heal, and J. M. Anderson, Decomposition in Terrestrial Ecosystems, Stud. Ecol., vol. 5 (Blackwell, Oxford, 1979).
V. M. Telesnina, I. N. Kurganova, V. O. Lopes de Gerenyu, L. A. Ovsepyan, V. I. Lichko, A. M. Yermolaev, and D. M. Mirin, “Dynamics of soil properties and plant composition during postagrogenic evolution in different bioclimatic zones,” Eurasian Soil Sci. 50, 1515–1534 (2017). https://doi.org/10.1134/S1064229317120109
V. M. Telesnina, I. E. Vaganov, A. A. Karlsen, A. E. Ivanova, M. A. Zhukov, and S. M. Lebedev, “Specific features of the morphology and chemical properties of coarse-textured postagrogenic soils of the southern taiga, Kostroma oblast,” Eurasian Soil Sci. 49, 102–115 (2016). https://doi.org/10.1134/S1064229316010117
V. M. Telesnina and M. A. Zhukov, “The influence of agricultural land use on the dynamics of biological cycling and soil properties in the course of postagrogenic succession (Kostroma oblast),” Eurasian Soil Sci. 52, 1114–1129 (2019). https://doi.org/10.1134/S1064229319070135
A. A. Titlyanova, G. I. Bulavko, S. Ya. Kudryashova, A. V. Naumov, V. V. Smirnov, and A. A. Tanasienko, “Stocks and losses of organic carbon in soils of Siberia,” Eurasian Soil Sci. 1, 51–59 (1998).
A. A. Titlyanova and A. V. Naumov, “Carbon losses from soils of Western Siberia under agriculture,” Eurasian Soil Sci. 11, 1357–1362 (1995).
A. S. Vladychenskii, V. M. Telesnina, K. A. Rumyantseva, and T. A. Chalaya, “Organic matter and biological activity of postagrogenic soils in the southern taiga using the example of Kostroma oblast,” Eurasian Soil Sci. 46, 518–529 (2013). https://doi.org/10.1134/S1064229313050141
M. D. Wallenstein, S. K. McMahon, and J. P. Schimel, “Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils,” Global Change Biol. 15, 1631–1639 (2009). https://doi.org/10.1111/j.1365-2486.2008.01819.x
B. Wang, G. B. Liu, S. Xue, and B. Zhu, “Changes in soil physicochemical and microbiological properties during natural succession on abandoned farmland in the loess plateau,” Environ. Earth Sci. 62 (5), 915–925 (2011). https://doi.org/10.1007/s12665-010-0577-4
M. Wiesmeier, L. Urbanski, E. Hobley, B. Lang, M. von Lützow, E. Marin-Spiotta, B. van Wesemael, E. Rabot, M. Ließ, N. Garcia-Franco, U. Wollschläger, H. J. Vogel, and I. Kögel-Knabner, “Soil organic carbon storage as a key function of soils—a review of drivers and indicators at various scales,” Geoderma 333, 149–162 (2019). https://doi.org/10.1016/j.geoderma.2018.07.026
A. G. Zavarzina, “Heterophase synthesis of humic acids in soils by immobilized phenol oxidases,” in Soil Enzymology (Springer-Verlag, Berlin, 2011), pp. 207–228.
L. Zhang, W. Chen, M. Burger, L. Yang, P. Gong, and Z. Wu, “Changes in soil carbon and enzyme activity as a result of different long-term fertilization regimes in a greenhouse field,” PLoS One 10, e0118371 (2015). https://doi.org/10.1371/journal.pone.0118371
F. Zhao, G. Yang, X. Han, Y. Feng, and G. Ren, “Stratification of carbon fractions and carbon management index in deep soil affected by the Grain-to-Green Program in China,” PLoS One 9 (6), e99657 (2014). https://doi.org/10.1371/journal.pone.0099657
Funding
This study was supported by the state budget for the Institute of Physicochemical and Biological Problems of Soil Science, Russian Academy of Sciences (project no. 121040800126-5); Ministry of Education and Science of the Russian Federation (project no. 075-15-2021-610); and Government of Tyumen oblast (project of the Western Siberian Interregional Science and Education Center no. 89-DON).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
The authors declare that they have no conflicts of interest.
Additional information
Translated by G. Chirikova
Rights and permissions
Open Access. This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kurganova, I.N., Telesnina, V.M., Lopes de Gerenyu, V.O. et al. Changes in the Carbon Stocks, Microbial and Enzyme Activities of Retic Albic Podzol in Southern Taiga during Postagrogenic Evolution. Eurasian Soil Sc. 55, 895–910 (2022). https://doi.org/10.1134/S1064229322070079
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S1064229322070079