
Abstract
Mesenchymal stem cells (mesenchymal stromal cells, MSC) are multipotent stem cells that can differentiate into cells of at least three mesodermal lineages, namely adipocytes, osteoblasts, and chondrocytes, and have potent immunomodulatory properties. Epigenetic modifications are critical regulators of gene expression and cellular differentiation of mesenchymal stem cells (MSCs). Epigenetic machinery controls MSC differentiation through direct modifications to DNA and histones. Understanding the role of epigenetic machinery in MSC is crucial for the development of effective cell-based therapies for degenerative and inflammatory diseases. In this review, we summarize the current understanding of the role of epigenetic control of MSC differentiation and immunomodulatory properties.
Similar content being viewed by others
Facts
-
Major transcription factors determining MSCs’ are epigenetically controlled.
-
C/EBPα (encoded by CBPA) and PPARγ (encoded by PPARG) are recognized as master regulators of adipocyte differentiation. The expression of CEBPA depends on HDAC1 activity. Opening of PPARG promoter region depends on SWI/SNF complex and simultaneous acetylation of histone H3 residues.
-
RUNX2 and OSX major orchestrators of osteogenic differentiation are controlled by, methylation at histone H3 residues, acetylation at histone H3 and H4 residues, and DNA methylation signatures.
-
Chondrogenic differentiation depends on MSCs condensation to form 3D structures. SOX9 is a major transcription factor in early chondrogenesis, and its expression is controlled by histone H3/H4 acetylation.
-
MSCs licensing with proinflammatory cytokines, hypoxia conditions, and bioactive molecules induces their immune-modulatory activities.
Open questions
-
Do differentially sourced MSCs differ in the epigenetic landscape, showing higher environmentally defined potential for differentiation or immune modulation (trained effect)?
-
What is the role of ncRNAs in the regulation of epigenetic machinery in MSCs?
-
What are the epigenetic mechanisms controlling MSCs immunomodulatory properties after licensing with proinflammatory cytokines?
-
How stress microenvironment is changing the epigenetic landscape of MSCs?
-
How to modulate MSCs epigenetic landscape to improve their therapeutic properties?
Introduction
Mesenchymal stem cells (MSCs, also known as mesenchymal stromal cells and multipotent stromal cells) were first identified in the bone marrow by Alexander Friedenstein et al. in the late '60s [1, 2]. They were described by Owen et al. as spindle-shaped, colony-forming unit fibroblasts (CFU-Fs) [3]. Nowadays, MSCs are successfully isolated and characterized from almost every tissue, including adipose tissue, lungs, liver, kidney, peripheral blood, umbilical cord, Wharton’s jelly, and dental pulp [4, 5]. To date, several phenotypical and functional variances among differentially sourced MSCs have been reported, which started a scientific debate over the homogeneity and their therapeutic properties. In 2005, the International Society for Cellular Therapy proposed three minimal criteria to define human MSCs, namely: i) plastic or glass surface adherence; ii) the ability to differentiate into at least adipogenic, chondrogenic, and osteogenic lineages in vitro; and iii) the presence of specific phenotype associated with the expression of CD29, CD90, CD73 with simultaneous lack of CD14, CD34, CD45, and HLA-DR expression [6,7,8].
MSCs gained considerable interest due to their promising therapeutic potential in degenerative and inflammatory diseases. The lack of immunogenicity offers the possibilities of their applications both in autologous and allogeneic systems [9]. Notably, MSCs activities such as self-renewal, differentiation, and immunoregulation can be modulated by microenvironmental factors. Consequently, MSCs’ stability remains the major safety issue, limiting their broad use in clinical settings. Therefore, a better understanding of MSCs’ biology and the effects of microenvironmental stimulation on their function is crucial to approximate their therapeutical usage. Notably, our understanding of the role of epigenetic machinery in cellular biology allowed us to define complex molecular mechanisms controlling cell phenotype and functional properties. It seems that, similarly to other stem and adult cells, epigenetic modifications play an important role in MSCs’ fate decisions and functions. Here, we aimed to summarize current progress in understanding the role of epigenetic mechanisms controlling MSCs’ differentiation and immunomodulatory function.
The mechanism of epigenetic modifications
All cellular processes and functions depend on the expression of genes encoding proteins or regulatory molecules. In fact, gene expression is a fundamental process that enables the decoding of the DNA sequences into the final functional gene product (cellular proteins). Notably, DNA is tightly packed within the nucleus of eukaryotic cells in highly organized and compact structures, making the gene hardly accessible to transcription machinery. Nucleosomes represent the first level of DNA packing, formed by DNA wrapped around histone proteins. Histones are nucleosome core proteins providing structural support to form chromatin. Five types of histones were identified. The nucleosome is formed by core histones, namely H2A, H2B, H3, and H4, while H1 and H5 are involved in higher-order chromatin structures [10]. Gene expression depends on chromatin accessibility, defined as the extent of physical interaction between nuclear macromolecules and “chromatinized” DNA. The structure of chromatin may be lightly packed (referred to as euchromatin) or tightly packed - more condensed (referred to as heterochromatin). Chromatin accessibility is controlled by the occupancy and topological arrangement of nucleosomes, along with the presence of various chromatin-binding factors, which bind either directly or indirectly to DNA, that modulate DNA accessibility to transcription machinery (extensively reviewed in ref. [11]). Progress in understanding molecular mechanisms regulating gene expression allowed us to define epigenetics.
Epigenetics refers to the heritable changes in gene expression and long-term alteration in the transcriptional potential at the chromatin level [12] (Fig. 1). The first evidence was reported by Waddington C.H. et al. in 1942 and defined as a phenomenon above genetics [13]. Currently, we understand that epigenetic regulation of gene expression may be mediated by DNA methylation, histone modifications, and chromatin remodeling, which occur without changes in the DNA sequences. In addition to the above-mentioned mechanisms associated with direct chromatin remodeling, control of gene expression by regulatory non-coding RNAs (ncRNAs) is usually considered a part of the epigenetic machinery. In this chapter, we will briefly introduce the essential epigenetic mechanisms associated with direct chromatin modifications that will be further discussed in the context of MSCs’ fate and immunomodulatory function.

Chromatin is composed of proteins (histones) and DNA that form chromosomes, and the spatial organization of chromatin is critical for gene expression regulation. Epigenetic machinery, including methylation of DNA and histone modifications, directly orchestrates chromatin activity, mediating the balance between an inactive (heterochromatin) or an active form (euchromatin). DNA methylation occurs by adding a methyl group (-CH3) to cytosine. This process is catalyzed by enzymes such as the activity of DNA methyltransferases (DNMTs). Histone modification includes methylation (Me), acetylation (Ac), phosphorylation (P), ubiquitination (U), and citrullination (C) on N-terminal tails. ISWI, CHD, SWI/SNF, and INO80 are the major subfamilies of ATP-dependent chromatin remodelers. In contrast to above mentioned epigenetic modifications, miRNAs are not directly involved in chromatin activity; however, as potent posttranslational regulators of gene expression, miRNAs may regulate the expression of enzymes involved in direct modifications of chromatin, including DNMTs, ten-eleven translocation (TETs) methyl dioxygenases (involved in DNA demethylation), histone deacetylases (HDACs), and histone-lysine N-methyltransferase enzymes belonging to EZH family (enhancer of zest homolog). Created with BioRender.com.
DNA methylation
DNA methylation is a biological process involving direct chemical modification of the DNA without changing its sequence [14]. This process is controlled by catalytic enzymes belonging to the family of methyltransferases (DNMTs), transferring a methyl group (-CH3) from S-adenosylmethionine (AdoMet/SAM) to cytosine in a dinucleotide sequence CpG (extensively reviewed in ref. [15]). Approximately 70% of gene promoter sites are associated with CpG islands (CGIs) which are responsible for transcriptional initiation [16]. The canonical DNMT enzymes, namely DNMT3a and DNMT3b, can establish de novo methylation to unmodified sequences [17]. At the same time, DNMT1 maintains the methyl group transfer from parental to newly synthesized DNA strand during DNA replication [18, 19]. DNA methylation was shown to play a crucial role in genomic imprinting, regulation of tissue-specific gene expression, and X chromosome inactivation; therefore, it is recognized as a conservative and long-lasting mechanism [20, 21].
Histone modifications
Chromatin structure is dense and hardly accessible for enzymes involved in transcription processes. Therefore, histones undergo post-transcriptional modifications to control DNA accessibility. To date, four mechanisms of histone modifications are best described and functionally characterized, namely acetylation, methylation, ubiquitination, and phosphorylation [22].
Histone acetylation is controlled by acetyltransferases (HATs) that transfer acetyl groups from acetyl co-enzyme A to lysine residues of the N-terminal H3 and H4 tails [23, 24]. Consequently, the positive charge of histone is neutralized, which reduces chromatin condensation, allowing the initialization of gene transcription [25]. This process may be reversed by activating histone deacetylases (HDACs), which remove acetyl groups from lysine residues making chromatin denser and more unavailable for transcriptional machinery and thus repressing gene expression [26, 27].
Similarly, to DNA methylation, methyl groups can be transferred to amino acids of histone proteins by methyltransferases; however, this process is independent of DNA methylation. Histone methylations can both repress or activate transcriptional processes depending on the methylation region, and a number of modifications occurred [28]. Histone methylation may occur on all histones, targeting especially lysine and arginine residues [29].
Ubiquitination refers to modifying the epsilon amino group of lysine by attaching one (monoubiquitinylation) or more (polyubiquitination) ubiquitin monomers. Ubiquitin is a small protein found in the vast majority of human tissues. Similarly, to the processes mentioned above, ubiquitination occurs in lysine residues of the N-terminal histone tails. Histones H2A and H2B seem to be the most ubiquitinated proteins in the nucleus [30, 31]. However, some reports showed that H1, H3, and H4 could also be modified by ubiquitination [32]. The process is controlled by several different enzymes such as Ubiquitin C-termin Hydrolase 1 (UCHL1), and Ubiquitin Specific Peptidase 1 (USP1) (for an extensive review, please see ref. [33]) and may control transcription processes, DNA damage responses, stem cell maintenance, and differentiation, among others [34]. However, it seems that H2A monoubiquitinylation is frequently associated with the repression of gene transcription [35, 36], while H2B monoubiquitinylation is often associated with increased transcription [37,38,39].
Contrary to the abovementioned processes, histone phosphorylation occurs on serine, thyronine, and threonine. Their role in the regulation of transcription remains not fully elucidated, although a number of proteins with phosphor-binding domains recognizing phosphorylated histones have been characterized [40, 41]. It seems that histone phosphorylation is primarily associated with the response to DNA damage (for review, please see ref. [42]).
Adenosine triphosphate-dependent chromatin remodeling
Adenosine triphosphate (ATP)-dependent chromatin alternations are mediated by multi-subunit chromatin-remodeling complexes utilizing the energy from ATP hydrolysis. These ATP-dependent chromatin remodeling enzymes were classified within the RNA/DNA helicase 2 superfamily and, based on their ATPase domain sequence similarities, may be divided into four groups/subfamilies, namely chromodomain helicase DNA-binding (CHD), SWItch/Sucrose non-fermentable (SWI/SNF), imitation SWI/SNF (ISWI), and INO80 (reviewed extensively in ref. [43]). Their direct role in developmental and physiological processes is well established. Interestingly, they may also be recruited by tissue-specific transcription factors to gene promoters and interact with other epigenetic modifiers such as HATS, HDACs, and histone methyltransferases (HMTs) [44].
Non-coding RNAs
ncRNAs are widely present in the eukaryotic genome and in contrast to messenger RNA do not translate into proteins. They are classified into housekeeping or regulatory ncRNAs. Housekeeping ncRNAs include ribosomal RNA (rRNA), transfer RNA (tRNA), small nuclear RNA (snRNA), small nucleolar RNA (snoRNA), and telomerase RNA (TERC). They are involved in fundamental cellular processes, including mRNA translation (rRNA, tRNA), pre-mRNA splicing (snRNA), RNA modifications (snoRNA), and telomeric DNA synthesis (TERC). In contrast, regulatory ncRNAs are considered functional components of gene expression, which are usually classified into two subclasses, namely small ncRNAs including microRNA (miRNA; 18-22 nt), small interfering RNA (siRNA; 20-25 nt), piwi RNA (piRNA; 26-31 nt), small cajal body-specific RNA (scaRNA; 200-300 nt), and long ncRNAs including long intergenic ncRNA (lincRNA; 1 kb nt), circular RNA (circRNA; 100-999 nt), and natural antisense transcript (NAT; >200 nt). The importance and function of ncRNAs in cellular biology were extensively reviewed in ref. [45]. In fact, regulatory ncRNAs play a crucial role in almost all cellular processes, regulating gene expression at the posttranscriptional level [46, 47]. However, ncRNAs do not directly modify DNA or chromatin structure. Although they can indirectly influence epigenetic modifications, such as recruiting histone modifiers or regulating the expression of proteins and enzymes involved in this process, they do not directly modify the epigenetic landscape of the genome [48,49,50]. Therefore, in this review, we will summarize the function of ncRNAs directly regulating the expression and activation of molecules involved in the epigenetic modifications (Table 1).
Epigenetic control of mesenchymal stem cell differentiation
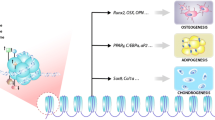
As mentioned, MSCs present the plasticity to differentiate into mesodermal lineage cells, namely adipocytes, osteocytes, and chondrocytes. Nowadays, it has become clear that epigenetic reprogramming in response to the mediators regulating MSCs’ fate (growth factors, cytokines, and metabolites) plays a vital role during differentiation. Here we will summarize our understanding of epigenetic control of MSCs differentiation.
Adipogenic differentiation
The adipogenic differentiation of MSCs required changes in the epigenetic landscape, allowing activation of the specific transcription machinery inducing cellular plasticity and further adipocyte maturation. Differentiation toward adipocytes is usually considered a two-step process, namely determination (described as a commitment to the adipocyte lineage) followed by differentiation (adipogenesis) (Fig. 2).

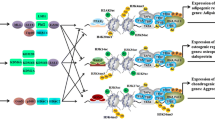
Adipogenesis occurs in a two-step process, namely the determination phase, followed by the differentiation phase. Adipogenic differentiation induction attracts CCAAT/enhancer-binding proteins beta and delta (C/EBPβ and C/EBPδ, respectively) activation and binding to peroxisome proliferator-activated receptor gamma (PPARG) promoter encoding PPARγ Simultaneously HDAC1 activity is decreased, which in turn activates the transcription of C/EBPα. C/EBPα and PPARγ are the major transcription factors whose expression is controlled by the acetylation (ac) of histone H3 lysine 9 and 27 (H3K9ac mark and H3K27ac mark, respectively). Further, di-methylation of histone H3 lysine 4 (H3K4me2 mark) upregulates adiponectin (apM1), glucose transporter type 4 (glut4), g-patch domain protein 1 (gdp1), and leptin (lep) gene expression. Created with BioRender.com.
The CCAAT/enhancer-binding protein alpha (C/EBPα encoded by CEBPA) and the peroxisome proliferator-activated receptor gamma (PPARγ, encoded by PPARG) are recognized as master regulators of adipocyte differentiation that control both stages of the process. The PPARG expression is activated after a few hours of differentiation induction by CCAAT/enhancer-binding proteins beta and delta (C/EBPβ and C/EBPδ, respectively) binding to the PPARG2 promoter [51,52,53]. In addition, PPARγ phosphorylation is reduced by miR-143 mediated downregulation of dual specificity mitogen-activated protein kinase kinase 5 (MAP2K5) expression and subsequent blocking of MAP2K5–ERK5 (extracellular signal-regulated kinase 5) signaling pathway [54].
From the perspective of the epigenetic machinery, the opening of the PPARG promoter region seems to depend on the activity of the SWI/SNF complex and is accompanied by acetylation of histone H3, namely acetylation of the 9th lysine (K9ac) residue of histone H3 (described as H3K9ac mark) and H3K27ac mark [55]. SWI/SNF complex activity has been shown to be regulated by lncRNAs such as SWINGN [56]. However, to date, it remains elusive whether this mechanism may be involved in the regulation of adipogenic differentiation of MSCs. The expression of CEBPA gene is initially blocked by HDAC1 activity, which is further downregulated upon accumulation of PPARγ through 26 S proteasome degradation, allowing C/EBPα expression [57, 58]. Moreover, C/EBPα activity promotes the expression of lncRNA TINCR, which acts as a sponge to reduce miR-31-5p and, in turn, unblock C/EBPα overexpression [59]. C/EBPα controls the expression of downstream genes by recruiting SWI/SNF complex to the promoter region of its target genes [60].
Committed cells remain undifferentiated and possess the phenotypic characteristic of MSCs. During the determination step, DNA demethylation in the promoter regions of genes critical for cell fate decision occurs. This process is accompanied by di-methylation of the 4th lysine (K4me2) residue of histone H3 (described as H3K4me2 mark) in the promoter regions of late adipogenic genes such as adiponectin (apM1), glucose transporter type 4 (glut4), g-patch domain protein 1 (gdp1), and leptin (lep) [61]. Interestingly, however, this activation mark is not associated with the active transcription of those genes in preadipocytes. Further, during the differentiation step, their active transcription is associated with DNA demethylation of their promoters, with simultaneous histone demethylation and histone H3 acetylation [62, 63]. Histone H3 acetylation seems to be mediated by CREB-binding protein (CBP), p300HAT, and simultaneous reduction of HDAC1 and HDAC3 activity [64, 65]. The process may be controlled by Sirt 1 (NAD-dependent) and HDAC3 (Zn-dependent) deacetylases to inhibit adipogenesis by decreasing PPARG expression [66]. In addition, upregulation of HDAC4-6 activity was observed during adipogenic differentiation of bone marrow-delivered MSCs [67]. Despite an increasing body of functional evidence indicating the significant importance of class II HDACs in the adipogenic differentiation of MSCs, the effect of their activity on changes in the epigenetic landscape remains elusive [68,69,70]. However, it becomes clear that HDAC4-6 expression is controlled by lncRNA H19 activity and lncRNA H19-derived miR-675 activity, which are downregulated during adipogenesis [71].
The differentiation process leads to the maturation of adipocytes and results in acquiring morphological and phenotypical characteristics as well as gene expression profiles of mature adipocytes. Importantly, once the adipogenic program is activated, MSCs lose their ability to differentiate towards other lineage cells and, therefore, are referred to as preadipocytes. Likewise, osteogenic differentiation inducers associated with the Wnt signaling pathway repress PPARG expression and, in consequence, block adipogenic differentiation [72]. The canonical Wnt/β-catenin pathway directly inhibits the expression of the PPARG gene. In contrast, the noncanonical Wnt pathway members such as Wnt5a promote adipogenesis at the initial stage of differentiation [73].
Osteogenic differentiation
In contrast to adipogenesis, osteogenic differentiation is characterized as a three-step process starting with the proliferative phase, then matrix maturation, and mineralization (Fig. 3). Runt-related transcription factor 2 (RUNX2, also known as a core-binding factor-α-CBFA1) and Osterix (OSX; also known as Sp7) represent the main transcription factors for osteogenic differentiation [74,75,76]. Their expression is accompanied by increased miR-27b, miR-130a, activity that directly targets PPARγ expression [77]. Both transcription factors control the expression of numerous downstream genes encoding proteins crucial for cellular plasticity induction and osteoblast phenotype establishment. Activation of RUNX2 induces transformation into osteoblast-lineage cells by increasing the expression of hedgehog (Indian Hedgehog Signaling Molecule (Ihh), Gli Family Zinc Finger 1 (Gli1), and Patched 1 (Ptch1)), fibroblast growth factor (Fgf; Fgfr1, Fgfr2, and Fgfr3), Wnt family member (Transcription Factor 7 (Tcf7), Wnt1 and Wnt10b), and parathyroid hormone-like hormone (Pthlh; Parathyroid Hormone 1 Receptor (Pthr1)) signaling genes [78]. In addition, RUNX2 modulates the expression of bone-related genes, including osteocalcin (OCN), collagen I (COL1a1), osteopontin (OPN), bone sialoprotein (BSP), alkaline phosphatase (ALP), and the parathyroid receptor (PTHR) [79, 80]. However, both RUNX2 and OSX with simultaneous canonical Wnt and BMP signaling pathway activation are required for mature osteoblast phenotype establishment and their proliferation [81, 82].

Osteogenesis is a three-step process consisting of the proliferative phase, matrix maturation, and mineralization. The process is controlled by RUNX2 and OSX transcription factors. In MSCs, their expression is inhibited by repressive marks, namely histone H3 mono-methylation at the 4th lysine residue (H3K4me1), histone H3 tri-methylation at the 9th lysine residue (H3K9me3), histone H3 tri-methylation of 27th lysine residue (H3K27me3), and DNA methylation at the 5th carbon of cytosine (5mCpG). After induction of differentiation, expression of methyltransferases (KMT2C and KMT2D) is reduced, and repressive marks are removed by SWItch/Sucrose non-fermentable complex (SWI/SNF) and histone demethylases (KDM4B and KDM6B) activity. They are replaced by activation marks, namely histone H3 tri-methylation at the 4th lysine residue (H3K4me3) and acetylation (ac) of histones H3 and H4, which opened the chromatin for RUNX2 and OSX expression. Histone acetylation is mediated by p300HAT (p300 histone acetyltransferase) and accompanied by decreased expression of HDAC1-3 (Histone deacetylases 1-3), while H3K4me3 is mediated by SET1/COMPASS and KMT2B activity and accompanied by reduced expression of KDM5B. Created with BioRender.com.
Notably, transcription of both mentioned orchestrators of osteogenic differentiation undergoes epigenetic control. To date, described mechanisms are associated with changes in i) methylation at histone H3 residues; ii) acetylation at histone H3 and H4 residues; and iii) DNA methylation signatures [83, 84]. All the mechanisms attract both promoter regions of the described transcription factors. Repressive marks, which are associated mainly with histone H3 modifications (such as mono-methylation at the 4th lysine residue (H3K4me1 mark), tri-methylation of 9th lysine residue (H3K9me3 mark), and tri-methylation of 27th lysine residue (H3K27me3 mark)) and direct DNA methylation at the 5th carbon of cytosine (5mCpG), are reduced and replaced with active marks such as trimethylation of 4th lysine on histone H3 (H3K4me3 mark) and acetylation of histone H3 (H3ac) and H4 (H4ac) [85]. Osteogenic differentiation decreases the activity of histone-H3K4-specific demethylase KDM5B and consequently increases the H3K4me3 mark activating Runx2 transcription [86, 87]. This process seems to be associated with increased activity of lncRNA TCONS_00041960, TCONS_00023297, and CALB2 acting as a sponge for Runx2 targeting miR-204-5p, miR-608-5p, and miR-30b-3p respectively [88,89,90]. On the other hand, H3K4me3 mark enrichment in both promoters of described transcription factors may be mediated by histone-H3K4-specific methylases belonging to the Complex Proteins Associated with Set1 (COMPASS) family complexes, namely SET1/COMPASS and MLL2/COMPASS-like (also known as KMT2B) [91,92,93]. In contrast, MLL3/COMPASS-like (also known as KMT2C) and MLL4/COMPASS-like (also referred to as KMT2D) seem to be responsible for the maintenance of repressive marks and are downregulated after induction of osteogenic differentiation by monomethylation of the 4th lysine of histone H3 (H3K4me1 mark) [94, 95]. In addition, H3K9me3 and H3K27me3 repressive marks are reduced by the activity of SWI/SNF (belonging as mentioned above to the ATP-dependent chromatin remodeling complexes family), histone demethylases (KDM4B and KDM6B, respectively) resulting in increased osteogenic potential [96,97,98,99]. Furthermore, histone H3 and H4 acetylation are associated with a reduced expression of HDAC1-3 activity and a simultaneous increase of histone acetyltransferase p300 (p300 HAT) activity, allowing effective acetylation in both promoter regions of RUNX2 and OSX [100, 101]. This process is also controlled by the lncRNA H19/miR-675 regulation of HDAC4-6 expression [67]. However, similarly to adipogenic differentiation, the role of HDAC4-6 in orchestrating the epigenetic landscape remains not fully elucidated. It has been proposed that inhibition of HDAC4-6 reduces acetylation of histone H4 at the endogenous OCN promoter induced by TGF−β1 [102].
The process of bone formation decreases with aging and during inflammatory bone diseases. High levels of IL-1β or TNF pro-inflammatory cytokines suppress osteogenesis by BMP/Smad signaling inhibition [103, 104]. However, Sirt1 activation increases MSC potential to osteogenic differentiation by modulation of NF-κβ transcription factor and RUNX2 upregulation [105]. Moreover, HOXB7 increased expression influence ON, OCN, BSP, and COL1A2 gene expression and further RUNX2 [106]. Importantly, RUNX2 high expression during proliferation phase promotes, in turn, distal-less homeobox 5 (DLX5) and bone sialoprotein (BSP) transcription factors [107, 108]. Furthermore, Satb2 contributes to MSC osteogenic differentiation [109]. However, the maturation and mineralization phase seems to be promoted by OSX, the second major transcription factor during osteogenesis, which adjusts the expression of Ocn, Opn, and Osteonectin (On) mature osteoblast genes [110]. Moreover, ALP expression increases through Runx2 influence [111].
Chondrogenic differentiation
MSCs differentiation toward chondrocytes strictly depends on cellular density (condensation of cells into 3D structures that occurs with the central role of N-cadherin) and growth factors stimulation (tumor growth factor beta (TGFβ) family members are well established in this process, namely TGFβ1, TGFβ2, and TGFβ3) (Fig. 4) [112, 113].

Chondrogenesis begins with mesenchymal condensation, followed by chondrocyte differentiation and maturation. SRY-Box Transcription Factor (SOX) 9, together with cofactors SOX5 and SOX6) represents a key transcription factor responsible for Collagen (COL2a1, COL9a1, and COL11a1) and Aggrecan (Acan) gene expression. The maturation phase is controlled by Histone H3 and H4 acetylation (ac) by p300HAT (p300 histone acetyltransferase) and Tip60HAT (Tip60 histone acetyltransferase). Chondrocyte maturation and hypertrophy are controlled by Runx2 expression. Its expression is associated with the demethylation of the 9th lysine (K9) at histone H3 (H3K9me mark) by Phf2 (PHD Finger Protein 2) demethylase and histone deacetylase 4 (HDAC4) downregulation. Created with BioRender.com.
Chondrogenesis commences with mesenchymal condensations (morphogenetic transformation mediated by cell adhesion), chondrocyte differentiation, and maturation. SRY-box containing gene 9 (SOX9) is a key transcription factor in early chondrogenesis [114]. Together with its cofactors, namely SRY-box containing gene 5 (SOX5) and SRY-box containing gene 6 (SOX6), control expression of cartilage-specific extracellular matrix genes (such as COL2A1, COL9A1, COL11A2, and Aggrecan (ACAN)) [115]. SOX9 expression is directly regulated by miR-145 and miR-495, and indirectly by miR-574-3p inhibiting retinoid x receptor alpha (RXRα) an inhibitor of chondrogenesis [116,117,118,119]. Moreover, Sox9 interacts with a number of differentiation stage-specific transcriptional regulators, such as WW domain-containing E3 Ubiquitin Protein Ligase 2 (Wwp2) transport to the cell nucleus. Wwp2 as a co-factor leads to increased expression of Sox9 [120]. Additionally, the transcription factor Zinc finger protein (Znf)219, associated by the C-terminal region with Sox9 factor activity on the Col2a1 promoter, upregulate Sox9 expression during chondrocyte differentiation and lead to increased expression of Col2a1, Col11a2, and ACAN [121]. AT-rich interaction domain (Arid)5a, as another transcriptional regulator, stimulates chondrogenesis by binding directly to the Col2a1 gene promoter [122]. Importantly, these activities are accompanied by changes in the epigenetic landscape involving both DNA methylation and histone modifications. In fact, chondrocyte-specific gene promoter regions (such as COL10A1) are hypomethylated in the course of MSC differentiation [123, 124]. In addition, histone acetylation has been shown to support chondrocyte differentiation [125, 126]. p300 HAT supports Sox9-mediated expression of cartilage-specific genes such as Col2a1, a major component of cartilage surrounding extracellular matrix, by acetylation of histones H3 and H4 in the enhancer regions [127]. Similarly, Tip60 HAT interacts with Sox9 and enhances its transcriptional activity in chondrocyte differentiation by the acetylation mark of lysine 61, 253, and 398 residues [128].
On the other hand, HDAC1- and HDAC2-mediated histone deacetylation significantly lowers the expression of extracellular matrix genes, including Col2a1, Col9a1, and Acan, as well as cartilage oligomeric matrix protein (a non-collagenous cartilage matrix protein) [129,130,131]. HDAC4 has been shown to inhibit Runx2 expression, thus significantly reducing chondrocyte differentiation and hypertrophy [132]. In addition, nicotinamide adenine dinucleotide (NAD)-dependent deacetylase Sirtuin1 has been reported to reduce the expression of the COL2A1 gene [133].
Unfortunately, our understanding of histone methylation in this process remains elusive. Similarly, our understanding of the role of ncRNAs in the regulation of epigenetic machinery during MSCs chondrogenic differentiation needs significantly more attention in the future. Some reports indicate an important role of H3K4me3 and H3K36me3 active marks [134, 135]. Moreover, H3K9 methylation has been shown to inhibit chondrocyte maturation and hypertrophy through the downregulation of Runx2 expression. This repression mark may be erased by the involvement of AT-rich interactive domain 5b (Arid5b) recruited histone demethylase Phf2 to promote chondrogenesis [136].
Epigenetic control of mesenchymal stem cells immunomodulatory properties
MSCs acquire immunosuppressive functions in a proinflammatory microenvironment (in response to cytokine stimulation such as IL-1α/β, IFNγ, TNF, and IL-17) [137, 138], hypoxia conditions [139], or in response to pharmacological drugs (bortezomib, dexamethasone) [140, 141] and chemical/biological agents (LL-37, LPS, curcumin, α-synuclein) [142,143,144,145], including epigenetic modifiers (HDAC/DNMT inhibitors) [146, 147]. To date, several MSCs mediated immunomodulatory mechanisms have been described, including (1) reduction of T cell-mediated responses by induction of T cell apoptosis, inhibition of T cell proliferation, and supporting of regulatory T cell differentiation [148]; (2) limitation of B cell responses by regulation B cell proliferation and differentiation towards plasma cells [149]; (3) inhibition of the cytotoxic function of natural killer (NK) cells [150]; (4) induction of dendritic cells (DCs) [151] and monocyte/macrophage (Mo/Ma) tolerogenic properties [152]; and (5) limitation of inflammatory mediator secretion (Fig. 5) [153].

TNF – tumor necrosis factor; IFN – interferon; IL – interleukin; IDO - indoleamine 2,3-dioxygenase; PGE2 – prostaglandin E2; COX2 – cyclooxygenase 2; HGF - hepatocyte growth factor; CCL5 – CC chemokine superfamily member; CXCL – CXC chemokine superfamily member; TSG-6 - Tumor necrosis factor-inducible gene 6 protein; 5-aza – 5-Aza-2′-deoxycytidine; PTGES - prostaglandin E synthase; TSA - trichostatin A, VPA - valproic acid. Created with BioRender.com.
The MSCs stimulation with pro-inflammatory cytokines leads to an increase in indoleamine 2,3-dioxygenase (IDO) levels, high IL-10 and prostaglandin E2 (PGE2) secretion, and induction of immunoinhibitory checkpoint (coinhibitory) molecule expression; all of these activities are recognized as hallmarks of their immunomodulatory functions [154, 155]. Interestingly, increased immunosuppressive activities have been observed after MSC priming with a combination of proinflammatory IFNγ and polyinosinic-polycytidylic acid (poly(I:C), a synthetic analog of double-stranded RNA, a ligand for toll-like receptor 3 (TLR3) stimulation [156,157,158]. However, it remains elusive how epigenetic machinery is involved in the proinflammatory cytokine-mediated MSCs licensing. In fact, our understanding of epigenetic mechanisms that control immunosuppressive functions of MSCs comes mainly from the reports describing the in vitro effect of chemical stimulations, including hypomethylating agents (5-aza-2’-deoxycytidine) (5-aza), HDAC inhibitors (trichostatin A and valproic acid), and drugs known to act on epigenetic marks (chlorzoxazone, sodium butyrate) [159,160,161,162].
5-aza is a chemical analog of cytidine, which possess hypomethylating activity and direct cytotoxicity in higher doses. In clinical practice, it is used to treat some types of malignancies, such as myelodysplastic syndrome (MDS) and acute myeloid leukemia (AML) [163]. In MSCs, 5-aza-2’-deoxycytidine has been shown to alter their differentiation potential and gene expression profiles, including enhancement of immunomodulatory gene expression. It leads to the demethylation of CXCR2, CXCR4, and CXCR6 promoters, which enhances MSCs migration potential in response to IL-8, C-X-C Motif Chemokine Ligand (CXCL1), and SDF-1 gradient [164]. The combination treatment of 5-aza with histone deacetylase inhibitors (trichostatin A and valproic acid-selectively inhibiting class I and II histone deacetylases) resulted in elevated expression of IDO and IL10. These licensed MSCs exerted a regulatory effect on Th1 and Th17 cell differentiation and significantly decreased IFNγ, IL-2, and IL-17 levels [165]. Microarray and methylation analyses revealed the demethylation of 7734 gene promoters and methylation of 5615 genes induced by 5-aza treatment of MSCs in inflammatory conditions. The pretreatment with 5-aza decreases methylation of immunomodulatory promoters COX2 and PTGES, and migration factors CXCR2 and CXCR4, subsequently contributing to the increased PGE2 production by MSC and suppress lymphocyte proliferation [147].
Deng L. et al. recently proposed chlorzoxazone for improving MSCs immunosuppressive properties in rat acute kidney injury model. Chlorzoxazone is a benzoxazole derivative used in clinical practice as a muscle-relaxing drug. Chlorzoxazone, in contrast to IFNγ stimulation, does not induce changes in the biological characteristic of MSCs and their MHC I and MHC II expression while similarly inhibiting the T cell activation and proliferation via the increase in IDO expression [160]. Moreover, chlorzoxazone elevates the expression of Cyclooxygenase (COX2), IL-4, Hepatocyte Growth Factor (HGF), TNF-stimulated gene 6 (TSG-6), C-C Motif Chemokine Ligand 5 (CCL5; RANTES), C-X-C Motif Chemokine Ligand (CXCL9, and CXCL10 (IP-10)) [160]. At the epigenetic level, chlorzoxazone has been shown to regulate the phosphorylation of transcription factor Forkhead Box O3 (FOXO3) independently to AKT and ERK signaling pathways [160]. Administration of chlorzoxazone-educated MSCs resulted in the reduction of renal tissue infiltration with immune cells and impairment of fibrinoid necrosis within the glomeruli, compared to naïve MSCs [160]. In addition, Yu T. et al. showed that inhibition of Tet-1 and Tet-2 demethylases in periodontium-derived MSCs (periodontal ligament stem cells) enhances their immunosuppressive functions. Tet-1 and Tet-2 knockdown results in DKK1 promoter hypermethylation and a decrease of DKK1 expression, leading to the activation of the Wnt/b-catenin signaling pathway and modulation of Fas ligand (FasL) expression. Consequently, MSCs acquire the ability to induce T cell apoptosis and increase the frequency of Tregs [166].
Similarly, to the above-mentioned drugs, stimulation of MSCs with IFNγ causes an increase in IDO enzyme level, resulting in the accumulation of toxic tryptophan metabolites that in turn, hamper T cell proliferation [167]. The expression of Ido1 gene seems to be associated with a combination of hypomethylating agents and histone deacetylation inhibitors (HDACIs) [168]. The increase in H3K9ac is accompanied simultaneously by a reduction in H3K9me3 at the promoter region [169].
Taking together, differentially sourced MSCs may vary in their immunosuppressive properties, and distinct strategies for empowering therapy should be assessed with a focus on the changes in biological characteristics, immune privilege, and functionality of MSCs [170,171,172]. Therefore, there is a substantial need to strengthen our understanding of the mechanisms and immunomodulatory potential of MSCs isolated from different tissues [165, 166, 173, 174]. This certainly would bring us closer to their use in clinical practice.
Perspective for improving clinical strategies involving MSCs
The approaches utilizing infusion or transfer of MSCs into patients have begun to develop in the 90’s of the last century. Over the past decades, the advances in MSCs administration procedures and safety strategies allowed for registering more than 950 clinical trials involving ~10,000 treated patients [175]. The considerable interest of the scientific community in optimizing MSC-based therapies in inflammatory and degenerative diseases resulted in the registration of more than 1000 clinical trials with a targeted enrollment of almost 50,000 patients in 2011–2018 [175]. Unfortunately, this great focus on clinical settings did not provide potent progress in the introduction of the first MSC-based therapy for routine medical practice.
Therefore, further efforts should be focused on enhancing the stability and effectiveness of MSCs transfer regardless of various conditions affecting the replicability of individual transplants. Given the complexity and diversity of the isolation procedures and expansion methods of MSCs, along with donor diversity and the heterogeneity in the inflammatory profiles of the recipient microenvironment, the achievement of the desired therapeutic outcome is very difficult. The advancement of two strategies might be useful in order to minimize currently defined issues, namely: (1) optimization of the MSC-related effects in controlled in vitro production conditions containing the standardized procedures using effective stabilizing/enhancing agents (such as inflammatory mediators, epigenetic or genetic modifiers); or (2) prediction suited properties and modification the MSC for personalized clinical indication (extensively reviewed in ref. [175]).
Recently developed methods, including single-cell sequencing (with multi-omics approaches) and further exploring of next-generation technologies, may progress our understanding of MSCs heterogeneity being the result of donors’ diversity, culturing conditions (hypoxia, length of culture), and isolation tissue sources [176]. Those advances may help to predict particular epigenetic manipulations or gene silencing, which might improve the desired therapeutic effect in individual clinical indications. On the other hand, the acquirement of a sufficient number of MSCs (hundreds of millions) for transplantation requires extensive expansion of cells in a standardized laboratory environment. The senescence of cells during culturing remains one of the leading problems in the advancement of cellular therapies. Aging MSCs may manifest loss of stemness, differentiation abilities, and immunomodulatory potential (for review please see ref. [177]). Therefore, epigenetic manipulation may maintain their beneficial properties or reverse the effects of senescence.
The improvement and substantial progress in the development of stem cell-based therapies require extensive research in the field of epigenetic machinery driving immunomodulatory activities and the differentiation potential of MSC. It is tempting to speculate that a better understanding of the role of epigenetic machinery and dynamic changes of the epigenetic landscape during fate decision, differentiation, and induction of immunomodulatory activities will allow us to better control the stability and enhance the therapeutic potential of MSCs.
References
Friedenstein AJ, Piatetzky-Shapiro II, Petrakova KV. Osteogenesis in transplants of bone marrow cells. J Embryol Exp Morphol. 1966;16:381–90.
Friedenstein AJ, Petrakova KV, Kurolesova AI, Frolova GP. Heterotopic of bone marrow. Analysis of precursor cells for osteogenic and hematopoietic tissues. Transplantation. 1968;6:230–47.
Owen M. Marrow stromal stem cells. J Cell Sci Suppl. 1988;10:63–76.
Lopez-Santalla M, Conde C, Rodriguez-Trillo A, Garin MI. Assessment of mesenchymal stem/stromal cell-based therapy in K/BxN serum transfer-induced arthritis. Front Immunol. 2022;13:943293.
Steens J, Klar L, Hansel C, Slama A, Hager T, Jendrossek V, et al. The vascular nature of lung-resident mesenchymal stem cells. Stem Cells Transl Med. 2021;10:128–43.
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini F, Krause D, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8:315–7.
Horwitz EM, Le Blanc K, Dominici M, Mueller I, Slaper-Cortenbach I, Marini FC, et al. Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy. 2005;7:393–5.
Tynecka M, Moniuszko M, Eljaszewicz A. Old friends with unexploited perspectives: current advances in mesenchymal stem cell-based therapies in asthma. Stem Cell Rev Rep. 2021;17:1323–42.
van Megen KM, van ‘t Wout ET, Lages Motta J, Dekker B, Nikolic T, Roep BO. Activated mesenchymal stromal cells process and present antigens regulating adaptive immunity. Front Immunol. 2019;10:694.
Wang S, Vogirala VK, Soman A, Berezhnoy NV, Liu ZB, Wong ASW, et al. Linker histone defines structure and self-association behaviour of the 177 bp human chromatosome. Sci Rep. 2021;11:380.
Klemm SL, Shipony Z, Greenleaf WJ. Chromatin accessibility and the regulatory epigenome. Nat Rev Genet. 2019;20:207–20.
Zhang L, Lu Q, Chang C. Epigenetics in health and disease. Adv Exp Med Biol. 2020;1253:3–55.
Waddington CH. The epigenotype. 1942. Int J Epidemiol. 2012;41:10-3.
Moore LD, Le T, Fan G. DNA methylation and its basic function. Neuropsychopharmacology. 2013;38:23–38.
Jaenisch R, Bird A. Epigenetic regulation of gene expression: how the genome integrates intrinsic and environmental signals. Nat Genet. 2003;33:245–54.
Deaton AM, Bird A. CpG islands and the regulation of transcription. Genes Dev. 2011;25:1010–22.
Gao L, Anteneh H, Song J. Dissect the DNMT3A- and DNMT3B-mediated DNA co-methylation through a covalent complex approach. J Mol Biol. 2020;432:569–75.
Lee PP, Fitzpatrick DR, Beard C, Jessup HK, Lehar S, Makar KW, et al. A critical role for Dnmt1 and DNA methylation in T cell development, function, and survival. Immunity. 2001;15:763–74.
Tao C, Liu J, Li Z, Lai P, Zhang S, Qu J, et al. DNMT1 is a negative regulator of osteogenesis. Biol Open. 2022;11:bio058534.
Andergassen D, Smith ZD, Kretzmer H, Rinn JL, Meissner A. Diverse epigenetic mechanisms maintain parental imprints within the embryonic and extraembryonic lineages. Dev Cell. 2021;56:2995–3005.e4.
Wang C, Liu X, Gao Y, Yang L, Li C, Liu W, et al. Reprogramming of H3K9me3-dependent heterochromatin during mammalian embryo development. Nat Cell Biol. 2018;20:620–31.
Kimura H. Histone modifications for human epigenome analysis. J Hum Genet. 2013;58:439–45.
Lei I, Tian S, Gao W, Liu L, Guo Y, Tang P, et al. Acetyl-CoA production by specific metabolites promotes cardiac repair after myocardial infarction via histone acetylation. Elife. 2021;10:e60311.
Ray A, Khan P, Nag Chaudhuri R. Deacetylation of H4 lysine16 affects acetylation of lysine residues in histone H3 and H4 and promotes transcription of constitutive genes. Epigenetics. 2021;16:597–617.
Erler J, Zhang R, Petridis L, Cheng X, Smith JC, Langowski J. The role of histone tails in the nucleosome: a computational study. Biophys J. 2014;107:2911–22.
Slaughter MJ, Shanle EK, Khan A, Chua KF, Hong T, Boxer LD, et al. HDAC inhibition results in widespread alteration of the histone acetylation landscape and BRD4 targeting to gene bodies. Cell Rep. 2021;34:108638.
Martin BJE, Brind’Amour J, Kuzmin A, Jensen KN, Liu ZC, Lorincz M, et al. Transcription shapes genome-wide histone acetylation patterns. Nat Commun. 2021;12:210.
Francis M, Gopinathan G, Foyle D, Fallah P, Gonzalez M, Luan X, et al. Histone methylation: achilles heel and powerful mediator of periodontal homeostasis. J Dent Res. 2020;99:1332–40.
Wilson JR. Determination of histone methyltransferase structure by crystallography. Methods Mol Biol. 2022;2529:137–47.
Hao S, Wang Y, Zhao Y, Gao W, Cui W, Li Y, et al. Dynamic switching of crotonylation to ubiquitination of H2A at lysine 119 attenuates transcription-replication conflicts caused by replication stress. Nucleic Acids Res. 2022;50:9873–92.
Shema-Yaacoby E, Nikolov M, Haj-Yahya M, Siman P, Allemand E, Yamaguchi Y, et al. Systematic identification of proteins binding to chromatin-embedded ubiquitylated H2B reveals recruitment of SWI/SNF to regulate transcription. Cell Rep. 2013;4:601–8.
Wang H, Zhai L, Xu J, Joo HY, Jackson S, Erdjument-Bromage H, et al. Histone H3 and H4 ubiquitylation by the CUL4-DDB-ROC1 ubiquitin ligase facilitates cellular response to DNA damage. Mol Cell. 2006;22:383–94.
Cao J, Yan Q. Histone ubiquitination and deubiquitination in transcription, DNA damage response, and cancer. Front Oncol. 2012;2:26.
Gu Y, Ding X, Huang J, Xue M, Zhang J, Wang Q, et al. The deubiquitinating enzyme UCHL1 negatively regulates the immunosuppressive capacity and survival of multipotent mesenchymal stromal cells. Cell Death Dis. 2018;9:459.
Wang YZ, Liu C, Zhao J, Yu J, Luo A, Xiao X, et al. H2A mono-ubiquitination differentiates FACT’s functions in nucleosome assembly and disassembly. Nucleic Acids Res. 2022;50:833–46.
Sijm A, Atlasi Y, van der Knaap JA, Wolf van der Meer J, Chalkley GE, Bezstarosti K, et al. USP7 regulates the ncPRC1 Polycomb axis to stimulate genomic H2AK119ub1 deposition uncoupled from H3K27me3. Sci Adv. 2022;8:eabq7598.
Sengupta B, Huynh M, Smith CB, McGinty RK, Krajewski W, Lee TH. The Effects of Histone H2B Ubiquitylations on the Nucleosome Structure and Internucleosomal Interactions. Biochemistry. 2022;61:2198–205.
Ai H, Sun M, Liu A, Sun Z, Liu T, Cao L, et al. H2B Lys34 ubiquitination induces nucleosome distortion to stimulate Dot1L activity. Nat Chem Biol. 2022;18:972–80.
Zhao Y, Pan J, Cao C, Liang X, Yang S, Liu L, et al. RNF20 affects porcine adipocyte differentiation via regulation of mitotic clonal expansion. Cell Prolif. 2021;54:e13131.
Liu T, Wang Y, Cheung SK, Or PM, Wong CW, Guan J, et al. The mitotic regulator RCC2 promotes glucose metabolism through BACH1-dependent transcriptional upregulation of hexokinase II in glioma. Cancer Lett. 2022;549:215914.
Wang Z, Li M, Jiang H, Luo S, Shao F, Xia Y, et al. Fructose-1,6-bisphosphatase 1 functions as a protein phosphatase to dephosphorylate histone H3 and suppresses PPARα-regulated gene transcription and tumour growth. Nat Cell Biol. 2022;24:1655–65.
Strahl BD, Allis CD. The language of covalent histone modifications. Nature. 2000;403:41–5.
Clapier CR, Iwasa J, Cairns BR, Peterson CL. Mechanisms of action and regulation of ATP-dependent chromatin-remodelling complexes. Nat Rev Mol Cell Biol. 2017;18:407–22.
Adachi A, Senmatsu S, Asada R, Abe T, Hoffman CS, Ohta K, et al. Interplay between chromatin modulators and histone acetylation regulates the formation of accessible chromatin in the upstream regulatory region of fission yeast fbp1. Genes Genet Syst. 2018;92:267–76.
Zhang P, Wu W, Chen Q, Chen M. Non-coding RNAs and their integrated networks. J Integr Bioinform. 2019;16:20190027.
Rusek AM, Abba M, Eljaszewicz A, Moniuszko M, Niklinski J, Allgayer H. MicroRNA modulators of epigenetic regulation, the tumor microenvironment and the immune system in lung cancer. Mol Cancer. 2015;14:34.
Hausser J, Zavolan M. Identification and consequences of miRNA-target interactions–beyond repression of gene expression. Nat Rev Genet. 2014;15:599–612.
Lan Y, Xie H, Jin Q, Zhao X, Shi Y, Zhou Y, et al. Extracellular vesicles derived from neural EGFL-Like 1-modified mesenchymal stem cells improve acellular bone regeneration via the miR-25-5p-SMAD2 signaling axis. Bioact Mater. 2022;17:457–70.
Bordin A, Chirivì M, Pagano F, Milan M, Iuliano M, Scaccia E, et al. Human platelet lysate-derived extracellular vesicles enhance angiogenesis through miR-126. Cell Prolif. 2022;55:e13312.
Peschansky VJ, Wahlestedt C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics. 2014;9:3–12.
Takenaka Y, Inoue I, Nakano T, Shinoda Y, Ikeda M, Awata T, et al. A novel splicing variant of peroxisome proliferator-activated receptor-γ (Pparγ1sv) cooperatively regulates adipocyte differentiation with Pparγ2. PLoS ONE. 2013;8:e65583.
Zwamborn RA, Slieker RC, Mulder PC, Zoetemelk I, Verschuren L, Suchiman HE, et al. Prolonged high-fat diet induces gradual and fat depot-specific DNA methylation changes in adult mice. Sci Rep. 2017;7:43261.
Medina-Gomez G, Gray SL, Yetukuri L, Shimomura K, Virtue S, Campbell M, et al. PPAR gamma 2 prevents lipotoxicity by controlling adipose tissue expandability and peripheral lipid metabolism. PLoS Genet. 2007;3:e64.
Chen L, Hou J, Ye L, Chen Y, Cui J, Tian W, et al. MicroRNA-143 regulates adipogenesis by modulating the MAP2K5-ERK5 signaling. Sci Rep. 2014;4:3819.
Wanior M, Preuss F, Ni X, Krämer A, Mathea S, Göbel T, et al. Pan-SMARCA/PB1 bromodomain inhibitors and their role in regulating adipogenesis. J Med Chem. 2020;63:14680–99.
Grossi E, Raimondi I, Goñi E, González J, Marchese FP, Chapaprieta V, et al. A lncRNA-SWI/SNF complex crosstalk controls transcriptional activation at specific promoter regions. Nat Commun. 2020;11:936.
Kuzmochka C, Abdou HS, Haché RJ, Atlas E. Inactivation of histone deacetylase 1 (HDAC1) but not HDAC2 is required for the glucocorticoid-dependent CCAAT/enhancer-binding protein α (C/EBPα) expression and preadipocyte differentiation. Endocrinology. 2014;155:4762–73.
Duan P, Wang H, Yi X, Zhang H, Chen H, Pan Z. C/EBPα regulates the fate of bone marrow mesenchymal stem cells and steroid-induced avascular necrosis of the femoral head by targeting the PPARγ signalling pathway. Stem Cell Res Ther. 2022;13:342.
Liu Y, Wang Y, He X, Zhang S, Wang K, Wu H, et al. LncRNA TINCR/miR-31-5p/C/EBP-α feedback loop modulates the adipogenic differentiation process in human adipose tissue-derived mesenchymal stem cells. Stem Cell Res. 2018;32:35–42.
Pedersen TA, Kowenz-Leutz E, Leutz A, Nerlov C. Cooperation between C/EBPalpha TBP/TFIIB and SWI/SNF recruiting domains is required for adipocyte differentiation. Genes Dev. 2001;15:3208–16.
Musri MM, Corominola H, Casamitjana R, Gomis R, Párrizas M. Histone H3 lysine 4 dimethylation signals the transcriptional competence of the adiponectin promoter in preadipocytes. J Biol Chem. 2006;281:17180–8.
Jang MK, Kim JH, Jung MH. Histone H3K9 demethylase JMJD2B activates adipogenesis by regulating H3K9 methylation on PPARγ and C/EBPα during adipogenesis. PLoS ONE. 2017;12:e0168185.
Tateishi K, Okada Y, Kallin EM, Zhang Y. Role of Jhdm2a in regulating metabolic gene expression and obesity resistance. Nature. 2009;458:757–61.
Wang M, Chen Z, Zhang Y. CBP/p300 and HDAC activities regulate H3K27 acetylation dynamics and zygotic genome activation in mouse preimplantation embryos. EMBO J. 2022;41:e112012.
Weinert BT, Narita T, Satpathy S, Srinivasan B, Hansen BK, Schölz C, et al. Time-resolved analysis reveals rapid dynamics and broad scope of the CBP/p300 acetylome. Cell. 2018;174:231–44.e12.
Fajas L, Egler V, Reiter R, Hansen J, Kristiansen K, Debril MB, et al. The retinoblastoma-histone deacetylase 3 complex inhibits PPARgamma and adipocyte differentiation. Dev Cell. 2002;3:903–10.
Huang Y, Zheng Y, Jin C, Li X, Jia L, Li W. Long non-coding RNA H19 inhibits adipocyte differentiation of bone marrow mesenchymal stem cells through epigenetic modulation of histone deacetylases. Sci Rep. 2016;6:28897.
Huang S, Wang S, Bian C, Yang Z, Zhou H, Zeng Y, et al. Upregulation of miR-22 promotes osteogenic differentiation and inhibits adipogenic differentiation of human adipose tissue-derived mesenchymal stem cells by repressing HDAC6 protein expression. Stem Cells Dev. 2012;21:2531–40.
Nebbioso A, Dell’Aversana C, Bugge A, Sarno R, Valente S, Rotili D, et al. HDACs class II-selective inhibition alters nuclear receptor-dependent differentiation. J Mol Endocrinol. 2010;45:219–28.
Lee S, Park JR, Seo MS, Roh KH, Park SB, Hwang JW, et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009;42:711–20.
Dey BK, Pfeifer K, Dutta A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014;28:491–501.
Ji H, Liu Y, Zhao X, Zhang M. N-acetyl-L-cysteine enhances the osteogenic differentiation and inhibits the adipogenic differentiation through up regulation of Wnt 5a and down regulation of PPARG in bone marrow stromal cells. Biomed Pharmacother. 2011;65:369–74.
Nishizuka M, Koyanagi A, Osada S, Imagawa M. Wnt4 and Wnt5a promote adipocyte differentiation. FEBS Lett. 2008;582:3201–5.
Lai CF, Shen J, Balic A, Pagella P, Schwab ME, Mitsiadis TA. Nogo-A regulates the fate of human dental pulp stem cells toward osteogenic, adipogenic, and neurogenic differentiation. Cells. 2022;11:3415.
Zhou L, Li H, Sun S, Zhang T, Yu Y, Xu L, et al. Thrap3 promotes osteogenesis by inhibiting the degradation of Runx2. FASEB J. 2022;36:e22231.
Xu Y, Chen S, Huang L, Han W, Shao Y, Chen M, et al. Epimedin C alleviates glucocorticoid-induced suppression of osteogenic differentiation by modulating PI3K/AKT/RUNX2 signaling pathway. Front Pharmacol. 2022;13:894832.
Seenprachawong K, Tawornsawutruk T, Nantasenamat C, Nuchnoi P, Hongeng S, Supokawej A. miR-130a and miR-27b enhance osteogenesis in human bone marrow mesenchymal stem cells via specific down-regulation of peroxisome proliferator-activated receptor γ. Front Genet. 2018;9:543.
Qin X, Jiang Q, Miyazaki T, Komori T. Runx2 regulates cranial suture closure by inducing hedgehog, Fgf, Wnt and Pthlh signaling pathway gene expressions in suture mesenchymal cells. Hum Mol Genet. 2019;28:896–911.
Ma L, Rao N, Jiang H, Dai Y, Yang S, Yang H, et al. Small extracellular vesicles from dental follicle stem cells provide biochemical cues for periodontal tissue regeneration. Stem Cell Res Ther. 2022;13:92.
Zheng H, Guo Z, Ma Q, Jia H, Dang G. Cbfa1/osf2 transduced bone marrow stromal cells facilitate bone formation in vitro and in vivo. Calcif Tissue Int. 2004;74:194–203.
Wang M, Li J, Ye Y, He S, Song J. SHED-derived conditioned exosomes enhance the osteogenic differentiation of PDLSCs via Wnt and BMP signaling in vitro. Differentiation. 2020;111:1–11.
Zhang B, Wang M, Gong A, Zhang X, Wu X, Zhu Y, et al. HucMSC-exosome mediated-Wnt4 signaling is required for cutaneous wound healing. Stem Cells. 2015;33:2158–68.
Montecino M, Carrasco ME, Nardocci G. Epigenetic control of osteogenic lineage commitment. Front Cell Dev Biol. 2020;8:611197.
Li K, Han J, Wang Z. Histone modifications centric-regulation in osteogenic differentiation. Cell Death Discov. 2021;7:91.
Zhang YX, Sun HL, Liang H, Li K, Fan QM, Zhao QH. Dynamic and distinct histone modifications of osteogenic genes during osteogenic differentiation. J Biochem. 2015;158:445–57.
Rojas A, Aguilar R, Henriquez B, Lian JB, Stein JL, Stein GS, et al. Epigenetic control of the bone-master Runx2 gene during osteoblast-lineage commitment by the histone demethylase JARID1B/KDM5B. J Biol Chem. 2015;290:28329–42.
Kidder BL, Hu G, Zhao K. KDM5B focuses H3K4 methylation near promoters and enhancers during embryonic stem cell self-renewal and differentiation. Genome Biol. 2014;15:R32.
Shang G, Wang Y, Xu Y, Zhang S, Sun X, Guan H, et al. Long non-coding RNA TCONS_00041960 enhances osteogenesis and inhibits adipogenesis of rat bone marrow mesenchymal stem cell by targeting miR-204-5p and miR-125a-3p. J Cell Physiol. 2018;233:6041–51.
Wang H, Wei P, Zhang Y, Li Y, Yin L. LncRNA TCONS_00023297 regulates the balance of osteogenic and adipogenic differentiation in bone marrow mesenchymal stem cells and the coupling process of osteogenesis and angiogenesis. Front Cell Dev Biol. 2021;9:697858.
Tu S, Wu J, Chen L, Tian Y, Qin W, Huang S, et al. LncRNA CALB2 sponges miR-30b-3p to promote odontoblast differentiation of human dental pulp stem cells via up-regulating RUNX2. Cell Signal. 2020;73:109695.
Sze CC, Cao K, Collings CK, Marshall SA, Rendleman EJ, Ozark PA, et al. Histone H3K4 methylation-dependent and -independent functions of Set1A/COMPASS in embryonic stem cell self-renewal and differentiation. Genes Dev. 2017;31:1732–7.
Denissov S, Hofemeister H, Marks H, Kranz A, Ciotta G, Singh S, et al. Mll2 is required for H3K4 trimethylation on bivalent promoters in embryonic stem cells, whereas Mll1 is redundant. Development. 2014;141:526–37.
Heintzman ND, Hon GC, Hawkins RD, Kheradpour P, Stark A, Harp LF, et al. Histone modifications at human enhancers reflect global cell-type-specific gene expression. Nature. 2009;459:108–12.
Hu D, Gao X, Morgan MA, Herz HM, Smith ER, Shilatifard A. The MLL3/MLL4 branches of the COMPASS family function as major histone H3K4 monomethylases at enhancers. Mol Cell Biol. 2013;33:4745–54.
Fagan RJ, Dingwall AK. COMPASS Ascending: Emerging clues regarding the roles of MLL3/KMT2C and MLL2/KMT2D proteins in cancer. Cancer Lett. 2019;458:56–65.
Busby T, Chen Y, Godfrey TC, Rehan M, Wildman BJ, Smith CM, et al. Baf45a mediated chromatin remodeling promotes transcriptional activation for osteogenesis and odontogenesis. Front Endocrinol (Lausanne). 2021;12:763392.
Hemming S, Cakouros D, Isenmann S, Cooper L, Menicanin D, Zannettino A, et al. EZH2 and KDM6A act as an epigenetic switch to regulate mesenchymal stem cell lineage specification. Stem Cells. 2014;32:802–15.
Deng P, Chang I, Wang J, Badreldin AA, Li X, Yu B, et al. Loss of KDM4B impairs osteogenic differentiation of OMSCs and promotes oral bone aging. Int J Oral Sci. 2022;14:24.
Ye L, Fan Z, Yu B, Chang J, Al Hezaimi K, Zhou X, et al. Histone demethylases KDM4B and KDM6B promotes osteogenic differentiation of human MSCs. Cell Stem Cell. 2012;11:50–61.
Iwata T, Kaneda-Ikeda E, Takahashi K, Takeda K, Nagahara T, Kajiya M, et al. Regulation of osteogenesis in bone marrow-derived mesenchymal stem cells via histone deacetylase 1 and 2 co-cultured with human gingival fibroblasts and periodontal ligament cells. J Periodontal Res. 2023;58:83–96.
Lagger G, O’Carroll D, Rembold M, Khier H, Tischler J, Weitzer G, et al. Essential function of histone deacetylase 1 in proliferation control and CDK inhibitor repression. EMBO J. 2002;21:2672–81.
Huang Y, Zheng Y, Jia L, Li W. Long noncoding RNA H19 promotes osteoblast differentiation via TGF-β1/Smad3/HDAC signaling pathway by deriving miR-675. Stem Cells. 2015;33:3481–92.
Mao CY, Wang YG, Zhang X, Zheng XY, Tang TT, Lu EY. Double-edged-sword effect of IL-1β on the osteogenesis of periodontal ligament stem cells via crosstalk between the NF-κB, MAPK and BMP/Smad signaling pathways. Cell Death Dis. 2016;7:e2296.
Liu C, Zhang H, Tang X, Feng R, Yao G, Chen W, et al. Mesenchymal stem cells promote the osteogenesis in collagen-induced arthritic mice through the inhibition of TNF-α. Stem Cells Int. 2018;2018:4069032.
Constanze B, Popper B, Aggarwal BB, Shakibaei M. Evidence that TNF-β suppresses osteoblast differentiation of mesenchymal stem cells and resveratrol reverses it through modulation of NF-κB, Sirt1 and Runx2. Cell Tissue Res. 2020;381:83–98.
Gao RT, Zhan LP, Meng C, Zhang N, Chang SM, Yao R, et al. Homeobox B7 promotes the osteogenic differentiation potential of mesenchymal stem cells by activating RUNX2 and transcript of BSP. Int J Clin Exp Med. 2015;8:10459–70.
Artigas N, Gámez B, Cubillos-Rojas M, Sánchez-de Diego C, Valer JA, Pons G, et al. p53 inhibits SP7/Osterix activity in the transcriptional program of osteoblast differentiation. Cell Death Differ. 2017;24:2022–31.
Egusa H, Kayashima H, Miura J, Uraguchi S, Wang F, Okawa H, et al. Comparative analysis of mouse-induced pluripotent stem cells and mesenchymal stem cells during osteogenic differentiation in vitro. Stem Cells Dev. 2014;23:2156–69.
Deng Y, Wu S, Zhou H, Bi X, Wang Y, Hu Y, et al. Effects of a miR-31, Runx2, and Satb2 regulatory loop on the osteogenic differentiation of bone mesenchymal stem cells. Stem Cells Dev. 2013;22:2278–86.
Fu H, Doll B, McNelis T, Hollinger JO. Osteoblast differentiation in vitro and in vivo promoted by Osterix. J Biomed Mater Res A. 2007;83:770–8.
Weng JJ, Su Y. Nuclear matrix-targeting of the osteogenic factor Runx2 is essential for its recognition and activation of the alkaline phosphatase gene. Biochim Biophys Acta. 2013;1830:2839–52.
Roger Y, Sydow S, Burmeister L, Menzel H, Hoffmann A. Sustained release of TGF-β. Colloids Surf B Biointerfaces. 2020;189:110843.
Lamparelli EP, Ciardulli MC, Giudice V, Scala P, Vitolo R, Dale TP, et al. 3D. Front Bioeng Biotechnol. 2022;10:986310.
Zhang Q, Ji Q, Wang X, Kang L, Fu Y, Yin Y, et al. SOX9 is a regulator of ADAMTSs-induced cartilage degeneration at the early stage of human osteoarthritis. Osteoarthritis Cartilage. 2015;23:2259–68.
Ylöstalo J, Smith JR, Pochampally RR, Matz R, Sekiya I, Larson BL, et al. Use of differentiating adult stem cells (marrow stromal cells) to identify new downstream target genes for transcription factors. Stem Cells. 2006;24:642–52.
Yang B, Guo H, Zhang Y, Chen L, Ying D, Dong S. MicroRNA-145 regulates chondrogenic differentiation of mesenchymal stem cells by targeting Sox9. PLoS ONE. 2011;6:e21679.
Lee S, Yoon DS, Paik S, Lee KM, Jang Y, Lee JW. microRNA-495 inhibits chondrogenic differentiation in human mesenchymal stem cells by targeting Sox9. Stem Cells Dev. 2014;23:1798–808.
Guérit D, Philipot D, Chuchana P, Toupet K, Brondello JM, Mathieu M, et al. Sox9-regulated miRNA-574-3p inhibits chondrogenic differentiation of mesenchymal stem cells. PLoS ONE. 2013;8:e62582.
Weston AD, Chandraratna RA, Torchia J, Underhill TM. Requirement for RAR-mediated gene repression in skeletal progenitor differentiation. J Cell Biol. 2002;158:39–51.
Nakamura Y, Yamamoto K, He X, Otsuki B, Kim Y, Murao H, et al. Wwp2 is essential for palatogenesis mediated by the interaction between Sox9 and mediator subunit 25. Nat Commun. 2011;2:251.
Takigawa Y, Hata K, Muramatsu S, Amano K, Ono K, Wakabayashi M, et al. The transcription factor Znf219 regulates chondrocyte differentiation by assembling a transcription factory with Sox9. J Cell Sci. 2010;123:3780–8.
Amano K, Hata K, Muramatsu S, Wakabayashi M, Takigawa Y, Ono K, et al. Arid5a cooperates with Sox9 to stimulate chondrocyte-specific transcription. Mol Biol Cell. 2011;22:1300–11.
Zimmermann P, Boeuf S, Dickhut A, Boehmer S, Olek S, Richter W. Correlation of COL10A1 induction during chondrogenesis of mesenchymal stem cells with demethylation of two CpG sites in the COL10A1 promoter. Arthritis Rheum. 2008;58:2743–53.
Barter MJ, Bui C, Cheung K, Falk J, Gómez R, Skelton AJ, et al. DNA hypomethylation during MSC chondrogenesis occurs predominantly at enhancer regions. Sci Rep. 2020;10:1169.
Chen W, Chen L, Zhang Z, Meng F, Huang G, Sheng P, et al. MicroRNA-455-3p modulates cartilage development and degeneration through modification of histone H3 acetylation. Biochim Biophys Acta. 2016;1863:2881–91.
Mao G, Zhang Z, Huang Z, Chen W, Huang G, Meng F, et al. MicroRNA-92a-3p regulates the expression of cartilage-specific genes by directly targeting histone deacetylase 2 in chondrogenesis and degradation. Osteoarthritis Cartilage. 2017;25:521–32.
Furumatsu T, Tsuda M, Yoshida K, Taniguchi N, Ito T, Hashimoto M, et al. Sox9 and p300 cooperatively regulate chromatin-mediated transcription. J Biol Chem. 2005;280:35203–8.
Hattori T, Coustry F, Stephens S, Eberspaecher H, Takigawa M, Yasuda H, et al. Transcriptional regulation of chondrogenesis by coactivator Tip60 via chromatin association with Sox9 and Sox5. Nucleic Acids Res. 2008;36:3011–24.
Huh YH, Ryu JH, Chun JS. Regulation of type II collagen expression by histone deacetylase in articular chondrocytes. J Biol Chem. 2007;282:17123–31.
Liu CJ, Zhang Y, Xu K, Parsons D, Alfonso D, Di Cesare PE. Transcriptional activation of cartilage oligomeric matrix protein by Sox9, Sox5, and Sox6 transcription factors and CBP/p300 coactivators. Front Biosci. 2007;12:3899–910.
Hong S, Derfoul A, Pereira-Mouries L, Hall DJ. A novel domain in histone deacetylase 1 and 2 mediates repression of cartilage-specific genes in human chondrocytes. FASEB J. 2009;23:3539–52.
Vega RB, Matsuda K, Oh J, Barbosa AC, Yang X, Meadows E, et al. Histone deacetylase 4 controls chondrocyte hypertrophy during skeletogenesis. Cell. 2004;119:555–66.
Wu WT, Chen YR, Lu DH, Senatov FS, Yang KC, Wang CC. Silymarin modulates catabolic cytokine expression through Sirt1 and SOX9 in human articular chondrocytes. J Orthop Surg Res. 2021;16:147.
Herlofsen SR, Bryne JC, Høiby T, Wang L, Issner R, Zhang X, et al. Genome-wide map of quantified epigenetic changes during in vitro chondrogenic differentiation of primary human mesenchymal stem cells. BMC Genomics. 2013;14:105.
Cheung K, Barter MJ, Falk J, Proctor CJ, Reynard LN, Young DA. Histone ChIP-Seq identifies differential enhancer usage during chondrogenesis as critical for defining cell-type specificity. FASEB J. 2020;34:5317–31.
Hata K, Takashima R, Amano K, Ono K, Nakanishi M, Yoshida M, et al. Arid5b facilitates chondrogenesis by recruiting the histone demethylase Phf2 to Sox9-regulated genes. Nat Commun. 2013;4:2850.
Hackel A, Aksamit A, Bruderek K, Lang S, Brandau S. TNF-α and IL-1β sensitize human MSC for IFN-γ signaling and enhance neutrophil recruitment. Eur J Immunol. 2021;51:319–30.
Watanabe Y, Fukuda T, Hayashi C, Nakao Y, Toyoda M, Kawakami K, et al. Extracellular vesicles derived from GMSCs stimulated with TNF-α and IFN-α promote M2 macrophage polarization via enhanced CD73 and CD5L expression. Sci Rep. 2022;12:13344.
Zhu Z, Zhang Y, Huang Z, Hao H, Yan M. Hypoxic culture of umbilical cord mesenchymal stem cell-derived sEVs prompts peripheral nerve injury repair. Front Cell Neurosci. 2022;16:897224.
Park HY, Kim CE, Lee SM, Ahn JM, Yoon EH, Yoo M, et al. Priming mesenchymal stem/stromal cells with a combination of a low dose of IFN-γ and bortezomib results in potent suppression of pathogenic Th17 immunity through the IDO1-AHR axis. Stem Cells. 2023;41:64–76.
Rawat S, Dadhwal V, Mohanty S. Dexamethasone priming enhances stemness and immunomodulatory property of tissue-specific human mesenchymal stem cells. BMC Dev Biol. 2021;21:16.
Oliveira-Bravo M, Sangiorgi BB, Schiavinato JL, Carvalho JL, Covas DT, Panepucci RA, et al. LL-37 boosts immunosuppressive function of placenta-derived mesenchymal stromal cells. Stem Cell Res Ther. 2016;7:189.
Liu GY, Liu Y, Lu Y, Qin YR, Di GH, Lei YH, et al. Short-term memory of danger signals or environmental stimuli in mesenchymal stem cells: implications for therapeutic potential. Cell Mol Immunol. 2016;13:369–78.
Yang Q, Leong SA, Chan KP, Yuan XL, Ng TK. Complex effect of continuous curcumin exposure on human bone marrow-derived mesenchymal stem cell regenerative properties through matrix metalloproteinase regulation. Basic Clin Pharmacol Toxicol. 2021;128:141–53.
Shin JY, Kim DY, Lee J, Shin YJ, Kim YS, Lee PH. Priming mesenchymal stem cells with α-synuclein enhances neuroprotective properties through induction of autophagy in Parkinsonian models. Stem Cell Res Ther. 2022;13:483.
Marquez-Curtis LA, Qiu Y, Xu A, Janowska-Wieczorek A. Migration, proliferation, and differentiation of cord blood mesenchymal stromal cells treated with histone deacetylase inhibitor valproic Acid. Stem Cells Int. 2014;2014:610495.
Lee S, Kim HS, Roh KH, Lee BC, Shin TH, Yoo JM, et al. DNA methyltransferase inhibition accelerates the immunomodulation and migration of human mesenchymal stem cells. Sci Rep. 2015;5:8020.
Li X, Xu Z, Bai J, Yang S, Zhao S, Zhang Y, et al. Umbilical cord tissue-derived mesenchymal stem cells induce T lymphocyte apoptosis and cell cycle arrest by expression of indoleamine 2, 3-dioxygenase. Stem Cells Int. 2016;2016:7495135.
Luz-Crawford P, Djouad F, Toupet K, Bony C, Franquesa M, Hoogduijn MJ, et al. Mesenchymal stem cell-derived interleukin 1 receptor antagonist promotes macrophage polarization and inhibits B cell differentiation. Stem Cells. 2016;34:483–92.
Yigitbilek F, Ozdogan E, Abrol N, Park WD, Hansen MJ, Dasari S, et al. Liver mesenchymal stem cells are superior inhibitors of NK cell functions through differences in their secretome compared to other mesenchymal stem cells. Front Immunol. 2022;13:952262.
Peng YQ, Wu ZC, Xu ZB, Fang SB, Chen DH, Zhang HY, et al. Mesenchymal stromal cells-derived small extracellular vesicles modulate DC function to suppress Th2 responses via IL-10 in patients with allergic rhinitis. Eur J Immunol. 2022;52:1129–40.
Li J, Pan Y, Yang J, Wang J, Jiang Q, Dou H, et al. Tumor necrosis factor-α-primed mesenchymal stem cell-derived exosomes promote M2 macrophage polarization. Front Immunol. 2022;13:945234.
Martire A, Bedada FB, Uchida S, Pöling J, Krüger M, Warnecke H, et al. Mesenchymal stem cells attenuate inflammatory processes in the heart and lung via inhibition of TNF signaling. Basic Res Cardiol. 2016;111:54.
Barrachina L, Remacha AR, Romero A, Vázquez FJ, Albareda J, Prades M, et al. Effect of inflammatory environment on equine bone marrow derived mesenchymal stem cells immunogenicity and immunomodulatory properties. Vet Immunol Immunopathol. 2016;171:57–65.
Lin T, Pajarinen J, Nabeshima A, Lu L, Nathan K, Jämsen E, et al. Preconditioning of murine mesenchymal stem cells synergistically enhanced immunomodulation and osteogenesis. Stem Cell Res Ther. 2017;8:277.
Sheng H, Wang Y, Jin Y, Zhang Q, Zhang Y, Wang L, et al. A critical role of IFNgamma in priming MSC-mediated suppression of T cell proliferation through up-regulation of B7-H1. Cell Res. 2008;18:846–57.
Barrachina L, Remacha AR, Romero A, Vázquez FJ, Albareda J, Prades M, et al. Priming equine bone marrow-derived mesenchymal stem cells with proinflammatory cytokines: implications in immunomodulation-immunogenicity balance, cell viability, and differentiation potential. Stem Cells Dev. 2017;26:15–24.
Lim JY, Kim BS, Ryu DB, Kim TW, Park G, Min CK. The therapeutic efficacy of mesenchymal stromal cells on experimental colitis was improved by the IFN-γ and poly(I:C) priming through promoting the expression of indoleamine 2,3-dioxygenase. Stem Cell Res Ther. 2021;12:37.
Killer MC, Nold P, Henkenius K, Fritz L, Riedlinger T, Barckhausen C, et al. Immunosuppressive capacity of mesenchymal stem cells correlates with metabolic activity and can be enhanced by valproic acid. Stem Cell Res Ther. 2017;8:100.
Deng L, Li H, Su X, Zhang Y, Xu H, Fan L, et al. Chlorzoxazone, a small molecule drug, augments immunosuppressive capacity of mesenchymal stem cells via modulation of FOXO3 phosphorylation. Cell Death Dis. 2020;11:158.
Kornicka K, Marycz K, Marędziak M, Tomaszewski KA, Nicpoń J. The effects of the DNA methyltranfserases inhibitor 5-Azacitidine on ageing, oxidative stress and DNA methylation of adipose derived stem cells. J Cell Mol Med. 2017;21:387–401.
Hsu WT, Huang WJ, Chiang BL, Tseng PH. Butyrate modulates adipose-derived stem cells isolated from polygenic obese and diabetic mice to drive enhanced immunosuppression. Cytotherapy. 2021;23:567–81.
Wenk C, Garz AK, Grath S, Huberle C, Witham D, Weickert M, et al. Direct modulation of the bone marrow mesenchymal stromal cell compartment by azacitidine enhances healthy hematopoiesis. Blood Adv. 2018;2:3447–61.
Pelekanos RA, Ting MJ, Sardesai VS, Ryan JM, Lim YC, Chan JK, et al. Intracellular trafficking and endocytosis of CXCR4 in fetal mesenchymal stem/stromal cells. BMC Cell Biol. 2014;15:15.
Kim KW, Kim HJ, Kim BM, Kwon YR, Kim HR, Kim YJ. Epigenetic modification of mesenchymal stromal cells enhances their suppressive effects on the Th17 responses of cells from rheumatoid arthritis patients. Stem Cell Res Ther. 2018;9:208.
Yu T, Liu D, Zhang T, Zhou Y, Shi S, Yang R. Inhibition of Tet1- and Tet2-mediated DNA demethylation promotes immunomodulation of periodontal ligament stem cells. Cell Death Dis. 2019;10:780.
Croitoru-Lamoury J, Lamoury FM, Caristo M, Suzuki K, Walker D, Takikawa O, et al. Interferon-γ regulates the proliferation and differentiation of mesenchymal stem cells via activation of indoleamine 2,3 dioxygenase (IDO). PLoS ONE. 2011;6:e14698.
Marchi E, Zullo KM, Amengual JE, Kalac M, Bongero D, McIntosh CM, et al. The combination of hypomethylating agents and histone deacetylase inhibitors produce marked synergy in preclinical models of T-cell lymphoma. Br J Haematol. 2015;171:215–26.
Perillo B, Di Santi A, Cernera G, Galasso G, Pocsfalvi G, Castoria G, et al. Acetylation/methylation at lysine 9 in histone H3 as a mark of nucleosome asymmetry in human somatic breast cells. Cell Death Discov. 2020;6:39.
Roubelakis MG, Pappa KI, Bitsika V, Zagoura D, Vlahou A, Papadaki HA, et al. Molecular and proteomic characterization of human mesenchymal stem cells derived from amniotic fluid: comparison to bone marrow mesenchymal stem cells. Stem Cells Dev. 2007;16:931–52.
Bieback K, Kern S, Klüter H, Eichler H. Critical parameters for the isolation of mesenchymal stem cells from umbilical cord blood. Stem Cells. 2004;22:625–34.
Mennan C, Garcia J, Roberts S, Hulme C, Wright K. A comprehensive characterisation of large-scale expanded human bone marrow and umbilical cord mesenchymal stem cells. Stem Cell Res Ther. 2019;10:99.
Qi J, Tang X, Li W, Chen W, Yao G, Sun L. Mesenchymal stem cells inhibited the differentiation of MDSCs via COX2/PGE2 in experimental sialadenitis. Stem Cell Res Ther. 2020;11:325.
Azevedo RI, Minskaia E, Fernandes-Platzgummer A, Vieira AIS, da Silva CL, Cabral JMS, et al. Mesenchymal stromal cells induce regulatory T cells via epigenetic conversion of human conventional CD4 T cells in vitro. Stem Cells. 2020;38:1007–19.
Pittenger MF, Discher DE, Péault BM, Phinney DG, Hare JM, Caplan AI. Mesenchymal stem cell perspective: cell biology to clinical progress. NPJ Regen Med. 2019;4:22.
Wang Z, Chai C, Wang R, Feng Y, Huang L, Zhang Y, et al. Single-cell transcriptome atlas of human mesenchymal stem cells exploring cellular heterogeneity. Clin Transl Med. 2021;11:e650.
Al-Azab M, Safi M, Idiiatullina E, Al-Shaebi F, Zaky MY. Aging of mesenchymal stem cell: machinery, markers, and strategies of fighting. Cell Mol Biol Lett. 2022;27:69.
Acknowledgements
This work was financially supported by the Medical University of Bialystok No. SUB/1/DN/21/003/1199. MT was supported by funds from Foundation for Polish Science.
Author information
Authors and Affiliations
Contributions
AW, AJ, and MT - draft preparation; AW – figure preparation; AW and AJ – table preparation; and MM and AE – final manuscript preparation. all authors read and accepted the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
AW and AJ declare no competing interests. MT reports National Science Centre Grant No. 2020/37/N/NZ5/04144; MM reports personal payments from Astra Zeneca, GSK, Sanofi, Berlin-Chemie/Menarini, Chiesi, Lek-AM, Takeda, Teva, Novartis, CSL Behring, Celon and support for attending meetings from Chiesi, Astra Zeneca, GSK, Berlin-Chemie/Menarini; AE: reports National Science Centre Grant No. 2020/37/N/NZ5/04144, National Centre for Research and Development No. STRATEGMED2/269807/14/NCBR/2015, National Centre for Research and Development (POLTUR3/MT-REMOD/2/2019).
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by Professor Yufang Shi
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Walewska, A., Janucik, A., Tynecka, M. et al. Mesenchymal stem cells under epigenetic control – the role of epigenetic machinery in fate decision and functional properties. Cell Death Dis 14, 720 (2023). https://doi.org/10.1038/s41419-023-06239-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-023-06239-4
- Springer Nature Limited