
Abstract
Microbial lipids are considered promising and environmentally friendly substitutes for fossil fuels and plant-derived oils. They alleviate the depletion of limited petroleum storage and the decrement of arable lands resulting from the greenhouse effect. Microbial lipids derived from oleaginous yeasts provide fatty acid profiles similar to plant-derived oils, which are considered as sustainable and alternative feedstocks for use in the biofuel, cosmetics, and food industries. Rhodotorula toruloides is an intriguing oleaginous yeast strain that can accumulate more than 70% of its dry biomass as lipid content. It can utilize a wide range of substrates, including low-cost sugars and industrial waste. It is also robust against various industrial inhibitors. However, precise control of the fatty acid profile of the lipids produced by R. toruloides is essential for broadening its biotechnological applications. This mini-review describes recent progress in identifying fatty synthesis pathways and consolidated strategies used for specific fatty acid-rich lipid production via metabolic engineering, strain domestication. In addition, this mini-review summarized the effects of culture conditions on fatty acid profiles in R. toruloides. The perspectives and constraints of harnessing R. toruloides for tailored lipid production are also discussed in this mini-review.
Graphical abstract

Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Owing to the depletion of petroleum storage and the growing world population, lipids derived from plants and animals have been exploited as an alternative to petroleum for producing biofuel and oleochemicals (Chemat et al. 2021; Ndiaye et al. 2019; Riazi et al. 2020; Shaah et al. 2021). However, production limitations (long harvesting time and unstable production affected by geographical and climate factors) and environmental issues (environmental pollution and competence of arable land for food crops) of plant- and animal-based lipids have forced scientists to seek new alternatives (Singh et al. 2020a; Meijaard et al. 2020; Toldrá-Reig et al. 2020). To date, much attention has been paid to the development of microbial lipids because of their short harvesting time and fewer environmental concerns (Bao et al. 2021; Liu et al. 2021c; Aamer Mehmood et al. 2021; Uthandi et al. 2021).
Rhodotorula toruloides is a non-conventional red yeast, also known as Rhodosporidium toruloides, R. rubescens, R. glutinis, or R. gracilis, that belongs to the subphylum Pucciniomycotina within the division Basidiomycota (Abeln and Chuck 2021; Koh et al. 2014). This strain is an excellent microbial lipid producer and has received great attention owing to its outstanding lipogenic ability. R. toruloides can accumulate up to 70% of its dry cellular weight as lipid content under nutrient-limiting conditions (nitrogen, phosphate, and sulfate) (Wang et al. 2018; Wu et al. 2010, 2011). It is also well known for its robustness, as it can produce lipids by utilizing a wide range of substrates from low-cost industrial by-products (Zhao et al. 2022; Wen et al. 2020; Saini et al. 2020; Park et al. 2018b). Numerous studies have described ways to improve lipid production in R. toruloides, including robust mutant selection, bioprocess optimization, and metabolic engineering (Zhao et al. 2022; Wen et al. 2020; Saini et al. 2020; Park et al. 2018b). In addition, available genome sequence (Dinh et al. 2019; Martín-Hernández et al. 2021; Zhu et al. 2012), characterized lipid metabolism (Jagtap et al. 2021), and accessible genetic tools (Bonturi et al. 2022; Jiao et al. 2019; Otoupal et al. 2019) for R. toruloides have also been established and developed. Two efficient and time-saving transformation methods by using Lithium-acetate (Tsai et al. 2017) and electroporation (Liu et al. 2017) were established to shorten the experimental time of the traditional method via Agrobacterium tumefaciens-mediated transformation (ATMT) (Lin et al. 2014). In addition, RNAi machinery was also proven to be functional for gene downregulation in R. toruloides (Liu et al. 2019). Genome editing toolboxes such as CRISPR-Cas9 system were also reported to achieve single or multi-genes editing in R. toruloides (Jiao et al. 2019; Otoupal et al. 2019; Schultz et al. 2019). All these advanced genetic tools and knowledge accelerated the improvement of the control of lipid production in R. toruloides and broaden its applications.
Microbial lipids rich in triacylglycerols are promising and attractive alternatives to existing petroleum-, plant-, and animal-based lipids. A triacylglycerol molecule consists of three fatty acid (acyl) chains attached to an alcohol glycerol backbone (Yoshinaga 2021). Carbon length, desaturation level, and position determine the physicochemical and biological properties of fatty acid (FA) chains attached to triacylglycerols (Cook and McMaster 2002; Temkov and Muresan 2021; Lee et al. 2022; Saini et al. 2021; Falomir-Lockhart et al. 2019). The lipids from R. toruloides have FA compositions that are similar to those of plant-derived oils. Despite the variation of fatty acid profiles existing in different R. toruloides strains, lipids produced in R. toruloides mainly consist of C16-C18 long-chain FAs (palmitic acid [PA], C16:0, 15–40% of total fatty acid (TFA); palmitoleic acid [POA], C16:1, 1–2% of TFA; stearic acid [SA], C18:0, 10–20% of TFA; oleic acid [OA], C18:1, 40–60% of TFA; linoleic acid [LA], C18:2, 1–10% of TFA; α-linolenic acid [ALA], C18:3, 1–2% of TFA). In addition, a small proportion of FAs, such as myristic acid (MA, C14:0, 2% of TFA), arachidic acid (AA, C20:0, less than 0.5% of TFA), docosanoic acid (DA, C22:0, less than 0.5% of TFA), and tetracosanoic acid (TA, C24:0, less than 1% of TFA), have also been reported in R. toruloides (Krikigianni et al. 2022; Zhang et al. 2022a; Liu et al. 2021a). Therefore, R. toruloides-derived lipids could potentially be exploited as feedstock for plant oil substitutes in different applications, such as biodiesel, biolubricants, cosmetics, nutritional supplements, plastics, and coating materials (Carmona-Cabello et al. 2021; Papadaki et al. 2018; Adrio 2017; Lopez-Huertas 2010; Yu et al. 2014). For example, lipids rich in OA (over 70% of TFA) are preferred for biodiesel production due to their appropriate fluidity and stability during storage (Graef et al. 2009). Lipids with high saturated FA profiles are desired as equivalents to coconut (PA, 23–30%; SA, 32–37%; OA, 30–37%; LA, 2–4% of TFA) and cocoa (PA, 26%; SA, 35%; OA, 33%; LA, 3% of TFA) butter in food industries (Lipp et al. 2001; Papanikolaou and Aggelis 2010). Moreover, lipids rich in polyunsaturated fatty acids (PUFA) such as docosahexaenoic acid (C22:6, DHA) and eicosapentaenoic acid (C20:5, EPA) (16% and 17% of TFA) are considered as substitutes to fish oils (Lee et al. 2019). Lipids containing conjugated linoleic acids (CLAs, a group of isomers of LA), γ-linolenic acid (GLA, C18:3), and nervonic acid (NA, C24:1), which are relatively low in their sources, are considered as valuable lipids because of their clinical benefits (Szczepańska et al. 2022). However, commercially desired lipids are often produced at low concentrations or with inappropriate FA compositions in microbial lipids, which may lead to complex downstream processes and result in high production costs (Ochsenreither et al. 2016; Barta et al. 2021). Therefore, harnessing R. toruloudes for specific FA-rich lipid production is a vital issue for industrialization. Most studies have focused on maximizing the lipid-producing ability of R. toruloides (Zhao et al. 2022; Wen et al. 2020). Hence, few review papers have summarized and discussed strategies for controlling FA composition in R. toruloides. Further, a thorough discussion of the culture conditions that affect FA profiles in R. toruloides has not yet been done. In this mini-review, we summarize the advances in the control and production of tailored lipids and the effects of culture conditions on FA profiles in R. toruloides (Table 1).
Harnessing de novo FA synthesis pathways in R. toruloides
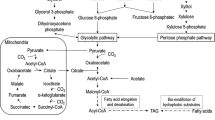
The FA composition of yeasts is precisely synthesized and regulated by FA synthases (Fass), elongases (Elos), and desaturases (Fads) (Heil et al. 2019; Singh et al. 2020b; Takaku et al. 2020; Matsuzawa et al. 2020). FA synthase, an essential enzyme in FA synthesis, catalyzes the reaction between acetyl-CoA and malonyl-CoA to produce C16-C18 FAs (Heil et al. 2019). R. toruloides Fas (RtFas) consists of two subunits (α- and β-subunits), which are encoded by FAS1 (RtFAS1) and FAS2 (RtFAS2), to form a multifunctional enzyme (Fischer et al. 2015; Zhu et al. 2012) (Fig. 1). RtFas1 contains acyltransferase and enoyl reductase domains. RtFas2 contains phosphopantetheinyl transferase, ketoacyl synthase, ketoacyl reductase, two acyl carrier proteins, malonyl/palmitoyl transferase, and dehydratase domains (Fig. 1). Liu et al. (2019) demonstrated that knocking down RtFAS1 and RtFAS2 using RNA interference decreased lipid content, but did not change the FA profile in R. toruloides NP11. Heterologous expression of RtFas in Saccharomyces cerevisiae also showed no effect on the FA composition (Zhou et al. 2016). These studies imply that FAS may not be a good candidate for manipulating FA composition in R. toruloides. However, enzyme engineering of the KS domain of Fas in the oleaginous yeast Yarrowia lipolytica enhanced the content of medium chain FAs (Rigouin et al. 2017), which suggests that engineering Fas may be applied in R. toruloides to modify the FA composition.

Schematic diagram of fatty acid synthase and fatty acid synthesis in R. toruloides cells. Abbreviations indicate components of the fatty acid synthase system. ACP acyl carrier protein, AT acyltransferase, DH dehydratase, ER enoyl reductase, KR ketoacyl reductase, KS ketoacyl synthase, MPT malonyl/palmitoyl transferase, PT phosphopantetheine transferase
Once malonyl-CoA is elongated to the carbon length of the acyl chain to C16, C16 acyl-CoA is released from the barrel-shaped Fas in R. toruloides. The released C16 acyl-CoA is further modified by FA Elos and Fads in R. toruloides (Zhu et al. 2012) (Fig. 2). Fads catalyze the desaturation of FAs by introducing double bonds into C–C bonds, which generate monounsaturated FAs (MUFAs) and polyunsaturated FAs (PUFAs) (Takaku et al. 2020; Wang et al. 2022a). Elos mediate the elongation of FAs by adding extra carbon atoms to the acyl chains. All desaturases and elongases exhibit different substrate specificities and spatial and temporal characteristics (Cerone and Smith 2022; Szczepańska et al. 2022). Therefore, elucidation of the functions of Fads and Elos could assist in the control of FA composition and production of customized lipids in R. toruloides.

Fatty acid elongation and desaturation pathways in R. toruloides. Genes encoding FA synthase, elongases, and desaturases are in red
The functions of Elos have been well studied in the model yeast S. cerevisiae and other yeasts used in the industry (Kihara 2012; Uemura 2012; Rigouin et al. 2018; Matsuzawa et al. 2020); however, few studies have elucidated the functions of Elos in R. toruloides. A patent (Fillet et al. 2016) reported that overexpression of RtELO1 significantly enhanced the OA content to 70% of the TFA in R. toruloides. In another patent (Liu et al. 2018), overexpression of RtELO1 led to an increase in OA. In addition, disruption of RtELO2 did not affect FA profiles (Liu et al. 2018). Conversely, studies on RtFads have also been conducted. R. toruloides Δ9-desaturase (RtOle1) catalyzes the conversion of SA to OA by introducing a cis-double bond at the Δ9 position of acyl-CoA. OA, one of the most essential and abundant FA in R. toruloides, provides appropriate fluidity and excellent thermal and oxidative stability for oleochemical applications such as biodiesel, biolubricants, and hydraulic fluids (Wang et al. 2022b). OA production is determined by RtOle1, which is a rate-limiting enzyme in LA and ALA biosynthesis (Nagao et al. 2019). Hence, RtOle1 plays a vital role in controlling the FA profiles. Overexpression of RtOle1 in R. toruloides have been shown to effectively increase OA content up to 70% and only slightly increase POA content (Tsai et al. 2019). Liu et al. (2021b) suggested that RtOle1 has a strong substrate preference for stearoyl-CoA over palmitoyl-CoA. In addition, blocking the downstream desaturation of OA into LA by disrupting RtFAD2 could also increase the OA content from 30 to 60% TFA. LA and ALA are essential FAs that humans cannot synthesize. Hence, lipids with high LA and ALA contents have high value as nutraceuticals in the human diet (Chen and Liu 2020). Liu et al. (2021b) also demonstrated that R. toruloides FAD2 and FAD4 encode Δ12/Δ15 desaturase and ∆9/Δ12/Δ15 desaturase, respectively, which produce LA and ALA from OA by introducing C–C double bonds at the Δ12 and Δ15 positions of FAs. Additionally, the co-overexpression of RtOLE1 and RtFAD2 has been applied to produce LA-rich lipids (Wu et al. 2021). Nevertheless, downregulation of RtOLE1 expression by elevated LA and ALA was observed in R. toruloides (Wu et al. 2021). Furthermore, two regulatory elements, ORE1 and ORE2, upstream of RtOLE1 have been reported (Liu et al. 2021b). The authors suggested that ORE1 positively regulates R. toruloides FAD gene transcription. Therefore, elucidating the transcription factors associated with the regulatory elements of RtOLE1 is necessary to manipulate the FA desaturation pathway. For example, S. cerevisiae transcriptional factors Spt2 and Mga2 upregulate essential genes involved in unsaturated FA biosynthesis (Zhang et al. 2022b; Sinha et al. 2022). The oleaginous Y. lipolytica Mga2 protein also regulates desaturase gene expression (Liu et al. 2015). Taken together, these studies show that manipulating the endogenous FA synthesis pathway in R. toruloides is an effective strategy for tailoring lipid production. Furthermore, the regulation of the native FA synthesis pathway should also be considered for a rational design to enhance specific FA production.
Metabolic engineering by introduction of exogenous genes for specific high-value lipid production
In addition to manipulating the native FA synthesis pathway, overexpression of genes from plants or other fungi was attempted in R. toruloides to produce specific high-value lipids. Disrupting native aldehyde dehydrogenase (ALD1) and overexpressing FAD2 from Mortierella alpina and FAD3 from Aleurites fordiiek led to a total ALA content of up to 49% of TFA (Liu et al. 2018). Tsai et al. (2019) enhanced the OA content from 50 to 70% of TFA by introducing S. cerevisiae OLE, which encodes a Δ9-desaturase. Wang et al. (2016) successfully enhanced LA content five-fold and achieved final LA titers of up to 1.3 g/L under flask culture conditions by introducing FAD2 from M. alpina and Fusarium verticillioides (Wang et al. 2016). GLA is a valuable omega-6 FA predominantly present in plants in relatively low amounts. GLA has anti-inflammatory properties and has applications in the treatment of several diseases (Liu et al. 2021b). Furthermore, Liu et al. (2021b) succeeded in producing GLA by disrupting native RtFAD2 and co-overexpressing M. alpine FAD2 and FAD6. The OA and GLA contents were further increased to 60.1% and 27.3% of TFA, respectively, in 2-L bioreactors.
FAs with carbon lengths of 20 or more are known as very-long-chain FAs (VLCFAs). VLCFAs, such as erucic acid (EA, C22:1) and NA, are potentially renewable feedstocks for the production of plastics, cosmetics, resins, nylon, and lubricants (Fillet et al. 2017). Additionally, NA has the potential to be used in the treatment of several neurological diseases (Liu et al. 2021a). Hence, EA and NA are high-value FAs. However, R. toruloides cannot naturally produce either EA or NA. To produce VLCFA-rich lipids, Fillet et al. (2017) introduced various plant-derived 3-ketoacyl-CoA synthases (KCSs) into R. toruloides and successfully produced EA and NA. By increasing the KCS copy number and co-overexpressing plant-derived ELOs, the engineered strain produced more EA and NA, which accounted for 27% of TFA in 7-L bioreactors.
Domestication of R. toruloides strains for tailored lipid production
Although metabolic engineering is a powerful strategy for harnessing R. toruloides for tailored lipid production, lack of information of the genetic background is the main limitation for strain engineering. Therefore, other strategies, such as adaptive laboratory evolution (ALE) and mutagenesis by physical/chemical methods, are also popular for isolating mutants with the desired traits for tailored lipid production in R. toruloides (Wen et al. 2020). The mutation rate is one of the main differences between ALE and mutagenesis using physicochemical methods. ALE domesticates parental strains with simultaneous mutation rates by growing several generations under selective pressure conditions (Phaneuf et al. 2020). Mutagenesis by physical and chemical methods commonly introduces a high mutation rate by random DNA double-strand breakage with UV irradiation and chemical agents. Although mutagenesis by physical and chemical methods is an effective way to introduce mutations into the genome, isolated mutants usually possess undesired defects (Arora et al. 2020). Owing to the advantages and disadvantages of the two methods, researchers have employed ALE and mutagenesis by physical and chemical methods in R. toruloides.
Wu et al. (2018) attempted to overcome the high temperatures that occur during fermentation by isolating thermotolerant R. toruloides DMKU3-TK16. The thermotolerant strain L1-1 was isolated under heat stress at 37°C using the ALE method. The isolated strain exhibited improved growth and lipid productivity at 37°C and a high OA content (90%) when cultured at 37°C. Liu et al. (2021d) attempted to improve the tolerance of R. toruloides to toxic lignocellulosic hydrolysates for lipid production. The isolated strains showed improved growth in media containing lignocellulosic biomass hydrolysate. According to their data, the evolved strains CH4 and CH5 showed improved OA and LA content (calculated). However, most evolved strains did not show significant FA profile alterations compared to the parental strain (R. toruloides NRRL Y-1091). Mutagenesis by UV has also been used for strain improvement in R. toruloides. Yamada et al. (2017) attempted to improve the lipid-producing ability of R. toruloides NBRC 8766 by UV irradiation and mutant selection under ethanol and H2O2 or cerulenin stress. Two strains with improved lipid production (8766 2-31 M and 8766 3-11C) were isolated. The FA profiles of these two strains showed considerably increased PA content (12.7% and 13.5%) with decreased MA and LA contents. Another study adopted a similar strategy of UV mutagenesis followed by mutant selection under ethanol and H2O2 or LiCl stress (Guo et al. 2019). The research team obtained two strains (R-ZL2 and R-ZL13) with improved lipid production from R. toruloides strain 2.1389. The R-ZL2 and R-ZL13 strains exhibited higher saturated FA profiles with increased PA (23 and 24%) and SA (12 and 10%) content, respectively. These two studies demonstrated that UV mutagenesis combined with stress selection could effectively domesticate R. toruloides to produce FA-rich lipids. Zhang et al. (2016a) obtained mutants of R. toruloides NP11 using atmospheric and room temperature plasma (ARTP) and nitrosoguanidine (NTG) methods. However, they did not apply stress selection after mutagenesis. According to their results, the isolated mutant XR-2 did not show a significant change in the FA profile compared with the parental strain R. toruloides NP11. In contrast, Qi et al. (2020) applied stress selection using inhibitory lignocellulosic hydrolysates after ARTP. Mutant RM18 exhibited improved ALA content (5.5%, 1.6 times more than the parental strain). These studies (Yamada et al. 2017; Guo et al. 2019; Zhang et al. 2016a; Qi et al. 2020) imply that stress selection after physical or chemical mutagenesis is necessary for screening mutants for desired FA production.
Culture conditions affect FA profiles in R. toruloides
Numerous strategies, such as optimizing culture conditions and waste utilization, have been applied to enhance lipid production in R. toruloides to produce economically competitive lipids (Zhao et al. 2022). However, the effects of each strategy on the FA profiles of lipids varied. Therefore, we have consolidated the progress obtained and provided a wide scope in the following sections. The information can help design rational bioprocesses to enhance specific FA-rich lipid production and assist in tailored lipid production.
Nitrogen, phosphate, and sulfate starvation
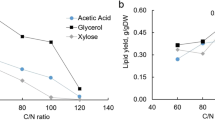
Nitrogen (N), phosphate (P), and sulfate (S) starvation can promote lipid accumulation in R. toruloides (Wang et al. 2018; Wu et al. 2010, 2011), and nitrogen limitation is the most adapted method. Different strategies affect lipid synthesis via different pathways, resulting in different metabolic fluxes. Therefore, the N, P, and S concentrations also affected the FA profiles. Ye et al. (2021) found that, when using sucrose as the sole carbon source and adjusting the C/N ratio to over 80 with ammonium sulfate or ammonium nitrate in the culture media, the FA contents of PA and ALA significantly increased in cells. Increasing the C/P ratio with glucose as a carbon source led to an increase in the OA content from 50 to 67% (Wu et al. 2010). In a study of S-limitation (Wu et al. 2011), lipids produced under increased C/S conditions favored saturated FAs (MA, PA, and SA) up to 60%. According to these studies, the strategies used to trigger lipid production should focus on enhancing the specific FA content.
Carbohydrates
Carbohydrates significantly affect FA profiles by activating key metabolic genes involved in gluconeogenesis, the glyoxylate cycle, and the tricarboxylic acid cycle (Sun et al. 2021; Patel et al. 2015). Most microbes possess a unique carbon catabolite mechanism. Microbes metabolize sugars sequentially because glucose represses the utilization of other sugars (Sun et al. 2021; Patel et al. 2015). Therefore, strategies using a primary carbon substrate or a defined ratio of carbon substrates have been reported for R. toruloides. Wiebe et al. (2012) demonstrated that R. toruloides could utilize C5 (arabinose and xylose) and C6 (glucose) sugars for lipid production. The lipids produced using arabinose and xylose as the sole carbon sources showed increased PA and LA contents compared to glucose, indicating that C5 (xylose and arabinose) sugars significantly changed the FA profiles. However, the FA profiles of the lipids produced using mixed sugars of glucose, arabinose, and xylose did not change significantly compared to those produced using glucose alone. Lignocellulosic biomass hydrolysate is a promising sustainable feedstock for microbial lipid production. After pretreatment and saccharification of lignocellulosic biomass, the complex links among lignin, cellulose, and hemicellulose are broken down, thus releasing fermentable sugars, such as glucose, xylose, or arabinose (Saini et al. 2022b). Osorio-González et al. (2019) demonstrated that R. toruloides strains had higher PA and SA contents when using C5 hydrolysate than C6 hydrolysate. Therefore, selecting carbohydrates and monitoring the carbohydrate content in the media during fermentation is vital for designing a rational fermentation process for desired FA production.
Crude substrates
In addition to using a single carbon source or defined carbon source, non-monomeric substrates containing abundant carbon sources from industrial, agricultural, municipal solid, and biomass wastes have been extensively studied and employed for lipid production in R. toruloides owing to their low cost (Abeln and Chuck 2021; Zhao et al. 2022).
Crude glycerol (CG) is a widely available product in the biodiesel industry. Approximately 10% (w/w) of glycerol is generated in every batch of biodiesel and is considered a waste (Lee et al. 2014; Uprety et al. 2016). Uprety et al. (2017, 2018) indicated that R. toruloides cells grown in media containing CG contained nearly 40% saturated FA (PA and SA), and a reduction in OA (from 60 to 47% of TFA) was observed compared to cells grown with pure glycerol. CG usually consists of glycerol and other impurities, such as glycerol, soap, methanol, ash, FA methyl esters (FAME), and salt. Uprety et al. (2018) reported that the decrease in OA in cells resulted from various impurities, particularly soap. Therefore, impurities in CG should be noted for tailored lipid production.
R. toruloides can utilize volatile FAs (VFAs), such as acetic acid (C2), propionic acid (C3), and butyric acid (C4) (Gao et al. 2017) for lipid production. VFAs can be obtained through the degradation of organic waste biomass by anaerobic digestion (Park et al. 2018a; Llamas et al. 2020), and thus VFAs can be exploited as cheap carbon sources for lipid production in yeasts (Park et al. 2018a, b; Llamas et al. 2020). However, different VFAs caused different FA changes in R. toruloides. R. toruloides grown in media with acetic acid (C2) as the sole carbon source generates high OA-lipids (nearly 50% of TFA) (Huang et al. 2016; Krikigianni et al. 2022). When propionic acid (C3) was used as the sole carbon source, the lipids had a high content of saturated FAs (65.9% of TFA) and a low MUFA content of 16% (Krikigianni et al. 2022). Notably, when propionic acid C3 was used, odd-chain FAs (margaric acid, C17:0, and heptadecenoic acid, C17:1) were generated, accounting for 30.8% of TFA. When fermented brewer’s spent grain contained mixed VFAs, the production of odd-chain FAs (ODFAs) was not observed, indicating that VFA composition influenced the FA profiles in R. toruloides. ODFAs are unusual and rare FAs that are found in natural sources. They are valuable FAs because they are associated with several health benefits, such as regulating allergies, psoriasis, and autoimmune disorders, and reducing the risk of metabolic disorders (Dąbrowski and Konopka 2022). Therefore, using VFAs as a carbon source for lipid production in R. toruloides could alter the native FA profile and produce unusual FA.
Plant-derived essential oils (EOs) can also alter the FA profiles of oleaginous microbes (Uprety and Rakshit 2018; Uprety et al. 2022). EOs are volatile and hydrophobic compounds mainly composed of terpenic hydrocarbons and oxygenated derivatives. EOs can impact the carbon flux toward a specific type of FAs inside microbes. Hence, EOs have been applied to alter the FA profile of R. toruloides (Uprety and Rakshit 2018). The EOs from clove-, cinnamon-, pine-, and orange-supplemented growth media for R. toruloides significantly increased the saturated FA content to 29%, 15%, 14%, and 36% of TFA, respectively. Furthermore, the EOs from orange also elevated PA content to 41% of TFA (Uprety and Rakshit 2018). Therefore, EOs are good inducers of FA production in R. toruloides.
Trace metal salts
Trace metal salts, such as zinc sulfate, copper sulfate, ferric chloride, manganese sulfate, and magnesium sulfate have also been commonly used as micronutrients in growth media to promote microbial growth (Saini et al. 2022a, b). These micronutrients serve as co-factors for several enzymes involved in lipid synthesis and desaturation pathways, and affect their catalytic activities (Singh et al. 2016; Ma et al. 2009; Romero et al. 2018). Furthermore, these metals may alter the catalytic activities of desaturase enzymes, resulting in alterations in the FA composition of lipids (Cai et al. 2020). Metal supplementation with wood hydrolysate as a carbon source causes the production of unusual PUFAs in R. toruloides (Saini et al. 2022a, b). When cells were grown in media supplemented with Mg2+, R. toruloides cells produced eicosatrienoic acid (ETA, C20:3) (0.03% of TFA). Similarly, an unusual DHA (0.05% of TFA) was produced by R. toruloides in media supplemented with Cu2+. In addition, adding Zn2+, Fe2+, and Cu2+ increased the content of GLA, which is also an unusual FA in R. toruloides, from 0.07% to 0.22%, 0.2%, and 0.12% of TFA, respectively. EA, DHA, and GLA are essential FA in humans; hence, they are high-value compounds. These results indicated that the addition of specific trace metals induced the production of high-value PUFAs in R. toruloides.
Oxygen
Oxygen availability is a critical parameter, especially during high cell density and large-scale fermentation, because of the unequal oxygen distribution in the microenvironment of the vessels, which affects cell growth. Oxygen is also a key molecule involved in FA desaturation. Fads introduce C–C double bonds into acyl chains by reducing oxygen (Sperling et al. 2003). Because these are oxygen-dependent reactions, low oxygen availability results in a higher degree of lipid saturation (Abeln and Chuck 2021). In addition, oxygen levels affect the activation of crucial genes, such as S. cerevisiae OLE1 in the FA saturation pathway through transcriptional factors (Romero et al. 2018; Burr et al. 2016). Therefore, oxygen is essential for the desaturation of FA by Fas. Choi et al. (1982) observed that the degree of unsaturation was significantly affected by the specific oxygen uptake rate of R. toruloides. Parreira et al. (2015) also confirmed that the PUFAs fraction increased as oxygen availability increased. These results indicated that oxygen availability is associated with PUFA synthesis in R. toruloides. Furthermore, the oxygen-activation pathways of Fas should be confirmed in R. toruloides.
Temperature
The growth temperature greatly influences various life processes in organisms, including gene transcription, metabolic activity, cell growth, nutrient absorption, cell survival, energy production, and membrane fluidity (Price and Sowers 2004; Fonseca et al. 2019). Microbial cells can modulate membrane fluidity to adapt to environmental temperature. Temperature influences membrane fluidity; decreasing temperature results in reduced viscosity and a more rigid membrane, whereas increasing temperature leads to increased viscosity and a looser membrane (Fonseca et al. 2019; Renne and Kroon 2018). Hence, microbial cells change their FA composition in the membrane lipids to maintain membrane fluidity. An increase in unsaturated FAs content reduces the melting point and increases membrane fluidity (Chen et al. 2022). On the other hand, an increase in saturated FAs content elevates their melting point and decreases membrane fluidity (Mejía-Barajas et al. 2018).
A growth temperature of 30°C is typically adopted for lipid production in R. toruloides. However, higher and lower temperatures significantly influence the FA profile of R. toruloides. The FA profile of the lipids in R. toruloides cells grown at 37°C increased the content of saturated FAs (PA and SA) (30% from 17%), which led to a higher saturated level than that at 30°C (Wu et al. 2020). Notably, the increase in saturated FA at 37°C may result from reduced RtFad2 desaturase activity (Wu et al. 2021). These reports suggest that temperature affects Fad activity and alters membrane FA composition, which may help cells maintain membrane fluidity. In contrast, when R. toruloides cells were grown at 15°C, the content of PUFAs, including LA and ALA, increased significantly compared to that at 25°C. The LA and ALA contents increased from 22 to 35% and from 8 to 21%, respectively (He et al. 2015). He et al. (2015) also demonstrated that the mRNA transcription level of a putative FA desaturase was five-fold higher in cells grown at 15°C than in cells grown at 25°C. In Y. lipolytica, the transcriptional factor Mga2, encoded by YALI0B12342g, may be associated with the upregulation of FAD2 at low temperatures (Tezaki et al. 2017). Therefore, elucidating the mechanism of low-temperature-induced FAD2 expression is necessary for R. toruloides to produce PUFAs.
pH
The commonly set pH for lipid production in R. toruloides is 5–6 (Li et al. 2006; Yang et al. 2014; Zhang et al. 2016b; Zeng et al. 2017). However, different medium pH values influence the FA profiles at different levels (Hall and Ratledge 1977). Similar scenarios were first observed in R. toruloides IIP-30 (formerly known as R. glutinis IIP-30) in the pH range of 3–6 (Johnson et al. 1992). Dias et al. (2016) also reported that medium pH values slightly altered FA profiles in R. toruloides. A slight MUFA percentage (OA) increased in the pH range 4.0–5.5. The PUFA percentages (LA and ALA) decreased at pH 4.5–5.5. Although the alteration in FA profiles did not show strong pH dependency, a specific pH range could assist in specific FA-rich lipid production based on their studies.
Light
Rhodotorula toruloides is a promising strain for carotenoid production. The carotenoid and lipid biosynthesis pathways are related because acetyl-CoA molecules are shared as precursors (Bruder et al. 2019). As a critical photo-inducer, light can promote carotenoid production by increasing microbial growth and activity of enzymes essential for carotenoid biosynthesis (Frengova and Beshkova 2009). In addition, photo-induced reactive oxygen species (ROS) may be associated with the activation of carotenoid synthesis pathways through the transcription factor bZIP in R. toruloides (Lin et al. 2017). Pham et al. (2020) demonstrated that R. toruloides cells grown under light conditions showed higher POA (two-fold) and ALA (four-fold) contents of TFA compared to those grown under dark conditions. Their results indicate that light could be a helpful factor for controlling FA profiles in R. toruloides.
Electro-fermentation
Electro-fermentation (EF) is a promising technique for improving the performance of bioprocesses that has been applied to increase lipid production in R. toruloides (Arbter et al. 2019). EF with the redox mediator Neutral Red (NR) caused a significant shift in the FA profile from OA to PA, resulting in a highly saturated FA composition (from 37 to 50%) in R. toruloides (Arbter et al. 2019). EF with NR is an excellent technique for producing highly saturated lipids as an alternative source of plant lipids such as coconut oil. In addition, EF with different media and strains may be applicable to R. toruloides.
Conclusion
Numerous review articles have summarized the different strategies for maximizing lipid production in R. toruloides. However, few have summarized and compared the impact of each method on FA composition. Thus, this mini-review describes strategies and advanced progress for controlling the FA composition in R. toruloudes through metabolic engineering. In addition, this mini-review also summarized the parameters of culture conditions such as substrate utilization and bioprocess control that have impacts on FA profile in R. toruloides. R. toruloides exhibits flexibility in producing lipids that are rich in saturated FA, MUFA, PUFA, and even unusual FAs. Therefore, R. toruloides is a promising platform for producing specific FA lipids. Further studies are necessary to reveal the mechanisms underlying the described strategies and findings to precisely control FA synthesis and bioprocess in R. toruloides for specific FA lipid production.
Data availability
The data supporting the findings of this mini-review are available in the references cited within this article.
Abbreviations
- AA:
-
Arachidic acid
- ACP:
-
Acyl carrier protein
- ALA:
-
α-Linolenic acid
- ALD:
-
Aldehyde dehydrogenase
- ALE:
-
Adaptive laboratory evolution
- ARTP:
-
Atmospheric and room temperature plasma
- AT:
-
Acyltransferase
- ATMT:
-
Agrobacterium tumefaciens-mediated transformation
- C:
-
Carbon
- CG:
-
Crude glycerol
- DA:
-
Docosanoic acid
- DH:
-
Dehydratase
- DHA:
-
Docosahexaenoic acid
- EA:
-
Erucic acid
- EF:
-
Electro-fermentation
- ELO:
-
Elongase
- EO:
-
Essential oil
- ER:
-
Enoyl reductase
- ETA:
-
Eicosatrienoic acid
- FA:
-
Fatty acid
- FAD:
-
Fatty acid desaturase
- FAME:
-
Fatty acid methyl ester
- FAS:
-
Fatty acid synthase
- GLA:
-
γ-Linolenic acid
- KCS:
-
Ketoacyl-CoA synthase
- KR:
-
Ketoacyl reductase
- KS:
-
Ketoacyl synthase
- LA:
-
Linoleic acid
- MA:
-
Myristic acid
- MPT:
-
Malonyl/palmitoyl transferase
- MUFA:
-
Monounsaturated fatty acid
- N:
-
Nitrogen
- NA:
-
Nervonic acid
- NR:
-
Neutral red
- NTG:
-
Nitrosoguanidine
- OA:
-
Oleic acid
- ODFA:
-
Odd-chain fatty acid
- P:
-
Phosphate
- PA:
-
Palmitic acid
- POA:
-
Palmitoleic acid
- PT:
-
Phosphopantetheine transferase
- PUFA:
-
Polyunsaturated fatty acid
- ROS:
-
Reactive oxygen species
- Rt:
-
Rhodotorula toruloides
- S:
-
Sulfate
- SA:
-
Stearic acid
- TA:
-
Tetracosanoic acid
- TFA:
-
Total fatty acid
- UV:
-
Ultraviolet
- VFA:
-
Volatile fatty acid
- VLCFA:
-
Very-long-chain fatty acid
References
Aamer Mehmood M, Shahid A, Malik S, Wang N, Rizwan Javed M, Nabeel Haider M, Verma P, Umer Farooq Ashraf M, Habib N, Syafiuddin A, Boopathy R (2021) Advances in developing metabolically engineered microbial platforms to produce fourth-generation biofuels and high-value biochemicals. Bioresour Technol 337:125510. https://doi.org/10.1016/j.biortech.2021.125510
Abeln F, Chuck CJ (2021) The history, state of the art and future prospects for oleaginous yeast research. Microb Cell Fact 20:221. https://doi.org/10.1186/s12934-021-01712-1
Adrio JL (2017) Oleaginous yeasts: Promising platforms for the production of oleochemicals and biofuels. Biotechnol Bioeng 114:1915–1920. https://doi.org/10.1002/bit.26337
Arbter P, Sinha A, Troesch J, Utesch T, Zeng AP (2019) Redox governed electro-fermentation improves lipid production by the oleaginous yeast Rhodosporidium toruloides. Bioresour Technol 294:122122. https://doi.org/10.1016/j.biortech.2019.122122
Arora N, Yen H-W, Philippidis GP (2020) Harnessing the power of mutagenesis and adaptive laboratory evolution for high lipid production by oleaginous microalgae and yeasts. Sustainability 12:5125. https://doi.org/10.3390/su12125125
Bao W, Li Z, Wang X, Gao R, Zhou X, Cheng S, Men Y, Zheng L (2021) Approaches to improve the lipid synthesis of oleaginous yeast Yarrowia lipolytica: a review. Renew Sustain Energy Rev 149:111386. https://doi.org/10.1016/j.rser.2021.111386
Barta DG, Coman V, Vodnar DC (2021) Microalgae as sources of omega-3 polyunsaturated fatty acids: Biotechnological aspects. Algal Res 58:102410. https://doi.org/10.1016/j.algal.2021.102410
Bonturi N, Pinheiro MJ, de Oliveira PM, Rusadze E, Eichinger T, Liudžiūtė G, De Biaggi JS, Brauer A, Remm M, Miranda EA, Ledesma-Amaro R, Lahtvee PJ (2022) Development of a dedicated Golden Gate Assembly Platform (RtGGA) for Rhodotorula toruloides. Metab Eng Commun 15:e00200. https://doi.org/10.1016/j.mec.2022.e00200
Burr R, Stewart EV, Shao W, Zhao S, Hannibal-Bach HK, Ejsing CS, Espenshade PJ (2016) Mga2 transcription factor regulates an oxygen-responsive lipid homeostasis pathway in fission yeast. J Biol Chem 291:12171–12183. https://doi.org/10.1074/jbc.M116.723650
Cai Y, Yu XH, Chai J, Liu CJ, Shanklin J (2020) A conserved evolutionary mechanism permits Δ9 desaturation of very-long-chain fatty acyl lipids. J Biol Chem 295:11337–11345. https://doi.org/10.1074/jbc.RA120.014258
Carmona-Cabello M, García IL, Papadaki A, Tsouko E, Koutinas A, Dorado MP (2021) Biodiesel production using microbial lipids derived from food waste discarded by catering services. Bioresour Technol 323:124597. https://doi.org/10.1016/j.biortech.2020.124597
Cerone M, Smith TK (2022) Desaturases: Structural and mechanistic insights into the biosynthesis of unsaturated fatty acids. IUBMB Life 74:1036–1051. https://doi.org/10.1002/iub.2671
Chemat F, Vian MA, Ravi HK (2021) Toward petroleum-free with plant-based chemistry. Curr Opin Green Sustain Chem 28:100450. https://doi.org/10.1016/j.cogsc.2021.100450
Chen J, Liu H (2020) Nutritional indices for assessing fatty acids: a mini-review. Int J Mol Sci 21:5695. https://doi.org/10.3390/ijms21165695
Chen W, Zhang X, Li S, Cui J, Yang X, Zhang Q (2022) The MAP-kinase HOG1 controls cold adaptation in Rhodosporidium kratochvilovae by promoting biosynthesis of polyunsaturated fatty acids and glycerol. Curr Microbiol 79:253. https://doi.org/10.1007/s00284-022-02957-8
Choi SY, Ryu DD, Rhee JS (1982) Production of microbial lipid: Effects of growth rate and oxygen on lipid synthesis and fatty acid composition of Rhodotorula gracilis. Biotechnol Bioeng 24:1165–1172. https://doi.org/10.1002/bit.260240513
Cook HW, McMaster CR (2002) Chapter 7 fatty acid desaturation and chain elongation in eukaryotes. New Compr Biochem 36:181–204. https://doi.org/10.1016/S0167-7306(02)36009-5
Dąbrowski G, Konopka I (2022) Update on food sources and biological activity of odd-chain, branched and cyclic fatty acids—a review. Trends Food Sci Technol 119:514–529. https://doi.org/10.1016/j.tifs.2021.12.019
Dias C, Silva C, Freitas C, Reis A, da Silva TL (2016) Effect of medium pH on Rhodosporidium toruloides NCYC 921 carotenoid and lipid production evaluated by flow cytometry. Appl Biochem Biotechnol 179:776–787. https://doi.org/10.1007/s12010-016-2030-y
Dinh HV, Suthers PF, Chan SHJ, Shen Y, Xiao T, Deewan A, Jagtap SS, Zhao H, Rao CV, Rabinowitz JF, Maranas CD (2019) A comprehensive genome-scale model for Rhodosporidium toruloides IFO0880 accounting for functional genomics and phenotypic data. Metab Eng Commun 9:e00101. https://doi.org/10.1016/j.mec.2019.e00101
Falomir-Lockhart LJ, Cavazzutti GF, Giménez E, Toscani AM (2019) Fatty acid signaling mechanisms in neural cells: fatty acid receptors. Front Cell Neurosci 13:162. https://doi.org/10.3389/fncel.2019.00162
Fillet S, Ronchel C, Callejo C, Fajardo MJ, Moralejo H, Adrio JL (2017) Engineering Rhodosporidium toruloides for the production of very long-chain monounsaturated fatty acid-rich oils. Appl Microbiol Biotechnol 101:7271–7280. https://doi.org/10.1007/s00253-017-8461-8
Fillet SC, Suárez González B, Ronchel Barreno MC, Velasco Álvarez J, Fondevila JLA (2016) Production of microbial oils with an elevated oleic acid content, WO 2016185073. Google Patent. https://patents.google.com/patent/WO2016185073A1/en. Accessed 27 October 2022.
Fischer M, Rhinow D, Zhu Z, Mills DJ, Zhao ZK, Vonck J, Grininger M (2015) Cryo-EM structure of fatty acid synthase (FAS) from Rhodosporidium toruloides provides insights into the evolutionary development of fungal FAS. Protein Sci 24:987–95. https://doi.org/10.1002/pro.2678
Fonseca F, Pénicaud C, Tymczyszyn EE, Gómez-Zavaglia A, Passot S (2019) Factors influencing the membrane fluidity and the impact on production of lactic acid bacteria starters. Appl Microbiol Biotechnol 103:6867–6883. https://doi.org/10.1007/s00253-019-10002-1
Frengova GI, Beshkova DM (2009) Carotenoids from Rhodotorula and Phaffia: Yeasts of biotechnological importance. J Ind Microbiol Biotechnol 36:163. https://doi.org/10.1007/s10295-008-0492-9
Gao R, Li Z, Zhou X, Cheng S, Zheng L (2017) Oleaginous yeast Yarrowia lipolytica culture with synthetic and food waste-derived volatile fatty acids for lipid production. Biotechnol Biofuels 10:247. https://doi.org/10.1186/s13068-017-0942-6
Graef G, LaVallee BJ, Tenopir P, Tat M, Schweiger B, Kinney AJ, Van Gerpen JH, Clemente TE (2009) A high-oleic-acid and low-palmitic-acid soybean: agronomic performance and evaluation as a feedstock for biodiesel. Plant Biotechnol J 7:411–421. https://doi.org/10.1111/j.1467-7652.2009.00408.x
Guo M, Cheng S, Chen G, Chen J (2019) Improvement of lipid production in oleaginous yeast Rhodosporidium toruloides by ultraviolet mutagenesis. Eng Life Sci 19:548–556. https://doi.org/10.1002/elsc.201800203
Hall MJ, Ratledge C (1977) Lipid accumulation in an oleaginous yeast (Candida 107) growing on glucose under various conditions in a one- and two-stage continuous culture. Appl Environ Microbiol 33:577–584. https://doi.org/10.1128/aem.33.3.577-584.1977
He J, Yang Z, Hu B, Ji X, Wei Y, Lin L, Zhang Q (2015) Correlation of polyunsaturated fatty acids with the cold adaptation of Rhodotorula glutinis. Yeast 32:683–690. https://doi.org/10.1002/yea.3095
Heil CS, Wehrheim SS, Paithankar KS, Grininger M (2019) Fatty acid biosynthesis: chain length regulation and control. Chembiochem 20:2298–2321. https://doi.org/10.1002/cbic.201800809
Huang XF, Liu JN, Lu LJ, Peng KM, Yang GX, Liu J (2016) Culture strategies for lipid production using acetic acid as sole carbon source by Rhodosporidium toruloides. Bioresour Technol 206:141–149. https://doi.org/10.1016/j.biortech.2016.01.073
Jagtap SS, Deewan A, Liu JJ, Walukiewicz HE, Yun EJ, Jin YS, Rao CV (2021) Integrating transcriptomic and metabolomic analysis of the oleaginous yeast Rhodosporidium toruloides IFO0880 during growth under different carbon sources. Appl Microbiol Biotechnol 105:7411–7425. https://doi.org/10.1007/s00253-021-11549-8
Jiao X, Zhang Y, Liu X, Zhang Q, Zhang S, Zhao ZK (2019) Developing a CRISPR/Cas9 system for genome editing in the Basidiomycetous yeast Rhodosporidium toruloides. Biotechnol J 14:e1900036. https://doi.org/10.1002/biot.201900036
Johnson V, Singh M, Saini VS, Sista VR, Yadav NK (1992) Effect of pH on lipid accumulation by an oleaginous yeast: Rhodotorula glutinis IIP-30. World J Microbiol Biotechnol 8:382–384. https://doi.org/10.1007/BF01198749
Kihara A (2012) Very long-chain fatty acids: elongation, physiology and related disorders. J Biochem 152:387–395. https://doi.org/10.1093/jb/mvs105
Koh CMJ, Liu Y, Moehninsi LL (2014) Molecular characterization of KU70 and KU80 homologues and exploitation of a KU70-deficient mutant for improving gene deletion frequency in Rhodosporidium toruloides. BMC Microbiol 14:50. https://doi.org/10.1186/1471-2180-14-50
Krikigianni E, Matsakas L, Rova U, Christakopoulos P, Patel A (2022) Investigating the bioconversion potential of volatile fatty acids: Use of oleaginous yeasts Rhodosporidium toruloides and Cryptococcus curvatus towards the sustainable production of biodiesel and odd-chain fatty acids. Appl Sci 12:6541. https://doi.org/10.3390/app12136541
Lee JJ, Chen L, Shi J, Trzcinski A, Chen WN (2014) Metabolomic profiling of Rhodosporidium toruloides grown on glycerol for carotenoid production during different growth phases. J Agric Food Chem 62:10203–10209. https://doi.org/10.1021/jf502987q
Lee AV, You L, Oh SY, Li Z, Code A, Zhu C, Fisher-Heffernan RE, Regnault TRH, De Lange CFM, Huber LA, Karrow NA (2019) Health benefits of supplementing nursery pig diets with microalgae or fish oil. Animals 9:80. https://doi.org/10.3390/ani9030080
Lee YY, Tang TK, Chan ES, Phuah ET, Lai OM, Tan CP, Wang Y, Ab Karim NA, Mat Dian NH, Tan JS (2022) Medium chain triglyceride and medium-and long chain triglyceride: Metabolism, production, health impacts and its applications—a review. Crit Rev Food Sci Nutr 62:4169–4185. https://doi.org/10.1080/10408398.2021.1873729
Li YH, Liu B, Zhao ZB, Bai FW (2006) Optimization of culture conditions for lipid production by Rhodosporidium toruloides. Chin J Biotechnol 22:650–656. https://doi.org/10.1016/S1872-2075(06)60050-2
Lin X, Wang Y, Zhang S, Zhu Z, Zhou YJ, Yang F, Sun W, Wang X, Zhao ZK (2014) Functional integration of multiple genes into the genome of the oleaginous yeast Rhodosporidium toruloides. FEMS Yeast Res 14:547–555. https://doi.org/10.1111/1567-1364.12140
Lipp M, Simoneau C, Ulberth F, Anklam E, Crews C, Brereton P, de Greyt W, Schwack W, Wiedmaier C (2001) Composition of genuine cocoa butter and cocoa butter equivalents. J Food Compost Anal 14:399–408. https://doi.org/10.1006/jfca.2000.0984
Liu L, Markham K, Blazeck J, Zhou N, Leon D, Otoupal P, Alper HS (2015) Surveying the lipogenesis landscape in Yarrowia lipolytica through understanding the function of a Mga2p regulatory protein mutant. Metab Eng 31:102–111. https://doi.org/10.1016/j.ymben.2015.07.004
Liu H, Jiao X, Wang Y, Yang X, Sun W, Wang J, Zhang S, Zhao ZK (2017) Fast and efficient genetic transformation of oleaginous yeast Rhodosporidium toruloides by using electroporation. FEMS Yeast Res. https://doi.org/10.1093/femsyr/fox017
Liu X, Zhang Y, Liu H, Jiao X, Zhang Q, Zhang S, Zhao ZK (2019) RNA interference in the oleaginous yeast Rhodosporidium toruloides. FEMS Yeast Res 19:031. https://doi.org/10.1093/femsyr/foz031
Liu F, Wang P, Xiong X, Zeng X, Zhang X, Wu G (2021) A review of nervonic acid production in plants: Prospects for the genetic engineering of high nervonic acid cultivars plants. Front Plant Sci 12:626625. https://doi.org/10.3389/fpls.2021.626625
Liu Y, Koh CMJ, Yap SA, Cai L, Ji L (2021) Understanding and exploiting the fatty acid desaturation system in Rhodotorula toruloides. Biotechnol Biofuels 14:73. https://doi.org/10.1186/s13068-021-01924-y
Liu Z, Moradi H, Shi S, Darvishi F (2021) Yeasts as microbial cell factories for sustainable production of biofuels. Renewab Sustain Energy Rev 143:110907. https://doi.org/10.1016/j.rser.2021.110907
Liu Z, Radi M, Mohamed ETT, Feist AM, Dragone G, Mussatto SI (2021) Adaptive laboratory evolution of Rhodosporidium toruloides to inhibitors derived from lignocellulosic biomass and genetic variations behind evolution. Bioresour Technol 333:125171. https://doi.org/10.1016/j.biortech.2021.125171
Liu Y, Koh CM, Ji L (2018) Methods for efficient production of polyunsaturated fatty acids (PUFA) in Rhodosporidium and Rhodotorula species. Google patent. https://patents.google.com/patent/US10081821B2/en. Accessed 27 October 2022.
Llamas M, Dourou M, González-Fernández C, Aggelis G, Tomás-Pejó E (2020) Screening of oleaginous yeasts for lipid production using volatile fatty acids as substrate. Biomass Bioenergy 138:105553. https://doi.org/10.1016/j.biombioe.2020.105553
Lopez-Huertas E (2010) Health effects of oleic acid and long chain omega-3 fatty acids (EPA and DHA) enriched milks. A review of intervention studies. Pharmacol Res 61:200–207. https://doi.org/10.1016/j.phrs.2009.10.007
Ma Z, Chu CH, Cheng D (2009) A novel direct homogeneous assay for ATP citrate lyase. J Lipid Res 50:2131–2135. https://doi.org/10.1194/jlr.D900008-JLR200
Martín-Hernández GC, Müller B, Chmielarz M, Brandt C, Hölzer M, Viehweger A, Passoth V (2021) Chromosome-level genome assembly and transcriptome-based annotation of the oleaginous yeast Rhodotorula toruloides CBS 14. Genomics 113:4022–4027. https://doi.org/10.1016/j.ygeno.2021.10.006
Matsuzawa T, Kamisaka Y, Maehara T, Takaku H, Yaoi K (2020) Identification and characterization of two fatty acid elongases in Lipomyces starkeyi. Appl Microbiol Biotechnol 104:2537–2544. https://doi.org/10.1007/s00253-020-10401-9
Meijaard E, Brooks TM, Carlson KM, Slade EM, Garcia-Ulloa J, Gaveau DLA, Lee JSH, Santika T, Juffe-Bignoli D, Struebig MJ, Wich SA, Ancrenaz M, Koh LP, Zamira N, Abrams JF, Prins HHT, Sendashonga CN, Murdiyarso D, Furumo PR, Macfarlane N, Hoffmann R, Persio M, Descals A, Szantoi Z, Sheil D (2020) The environmental impacts of palm oil in context. Nat Plants 6:1418–1426. https://doi.org/10.1038/s41477-020-00813-w
Mejía-Barajas J, Montoya-Pérez R, Manzo-Avalos S, Cortés-Rojo C, Riveros-Rosas H, Cervantes C, Saavedra-Molina A (2018) Fatty acid addition and thermotolerance of Kluyveromyces marxianus. FEMS Microbiol Lett 365:fny043. https://doi:doi:https://doi.org/10.1093/femsle/fny043
Nagao K, Murakami A, Umeda M (2019) Recent progress in biophysical research of biological membrane systems. Chem Pharm Bull 67:327–332. https://doi.org/10.1248/cpb.c18-01001
Ndiaye M, Arhaliass A, Legrand J, Roelens G, Kerihuel A (2019) Reuse of waste animal fat in biodiesel: Biorefining heavily-degraded contaminant-rich waste animal fat and formulation as diesel fuel additive. Renew Energy 145:1073–1079. https://doi.org/10.1016/j.renene.2019.06.030
Ochsenreither K, Glück C, Stressler T, Fischer L, Syldatk C (2016) Production strategies and applications of microbial single cell oils. Front Microbiol 7:1539. https://doi.org/10.3389/fmicb.2016.01539
Otoupal PB, Ito M, Arkin AP, Magnuson JK, Gladden JM, Skerker JM (2019) Multiplexed CRISPR-Cas9-based genome editing of Rhodosporidium toruloides. mSphere. https://doi.org/10.1128/mSphere.00099-19
Papadaki A, Fernandes KV, Chatzifragkou A, Aguieiras ECG, da Silva JAC, Fernandez-Lafuente R, Papanikolaou S, Koutinas A, Freire DMG (2018) Bioprocess development for biolubricant production using microbial oil derived via fermentation from confectionery industry wastes. Bioresour Technol 267:311–318. https://doi.org/10.1016/j.biortech.2018.07.016
Papanikolaou S, Aggelis George (2010) Yarrowia lipolytica: a model microorganism used for the production of tailor-made lipids. Eur J Lipid Sci Technol 112:639–654. https://doi.org/10.1002/EJLT.200900197
Park YK, Dulermo T, Ledesma-Amaro R, Nicaud JM (2018) Optimization of odd chain fatty acid production by Yarrowia lipolytica. Biotechnol Biofuels 11:158. https://doi.org/10.1186/s13068-018-1154-4
Park YK, Nicaud JM, Ledesma-Amaro R (2018) The engineering potential of Rhodosporidium toruloides as a workhorse for biotechnological applications. Trends Biotechnol 36:304–317. https://doi.org/10.1016/j.tibtech.2017.10.013
Parreira TM, Freitas C, Reis A, Roseiro J, da Silva TL (2015) Carbon concentration and oxygen availability affect lipid and carotenoid production by carob pulp syrup-grown Rhodosporidium toruloides NCYC 921. Eng Life Sci 15:815–823. https://doi.org/10.1002/elsc.201500002
Patel A, Pruthi V, Singh RP, Pruthi PA (2015) Synergistic effect of fermentable and non-fermentable carbon sources enhances TAG accumulation in oleaginous yeast Rhodosporidium kratochvilovae HIMPA1. Bioresour Technol 188:136–144. https://doi.org/10.1016/j.biortech.2015.02.062
Pham KD, Shida Y, Miyata A, Takamizawa T, Suzuki Y, Ara S, Yamazaki H, Masaki K, Mori K, Aburatani S, Hirakawa H, Tashiro K, Kuhara S, Takaku H, Ogasawara W (2020) Effect of light on carotenoid and lipid production in the oleaginous yeast Rhodosporidium toruloides. Biosci Biotechnol Biochem 84:1501–1512. https://doi.org/10.1080/09168451.2020.1740581
Phaneuf PV, Yurkovich JT, Heckmann D, Wu M, Sandberg TE, King ZA, Tan J, Palsson BO, Feist AM (2020) Causal mutations from adaptive laboratory evolution are outlined by multiple scales of genome annotations and condition-specificity. BMC Genom 21:514. https://doi.org/10.1186/s12864-020-06920-4
Price PB, Sowers T (2004) Temperature dependence of metabolic rates for microbial growth, maintenance, and survival. Proc Natl Acad Sci USA 101:4631–4636. https://doi.org/10.1073/pnas.0400522101
Qi F, Shen P, Hu R, Xue T, Jiang X, Qin L, Chen Y, Huang J (2020) Carotenoids and lipid production from Rhodosporidium toruloides cultured in tea waste hydrolysate. Biotechnol Biofuels 13:74. https://doi.org/10.1186/s13068-020-01712-0
Renne MF, de Kroon AIPM (2018) The role of phospholipid molecular species in determining the physical properties of yeast membranes. FEBS Lett 592:1330–1345. https://doi.org/10.1002/1873-3468.12944
Riazi B, Mosby JM, Millet B, Spatari S (2020) Renewable diesel from oils and animal fat waste: implications of feedstock, technology, co-products and ILUC on life cycle GWP. Resour Conserv Recycl 161:104944. https://doi.org/10.1016/j.resconrec.2020.104944
Rigouin C, Gueroult M, Croux C, Dubois G, Borsenberger V, Barbe S, Marty A, Daboussi F, André I, Bordes F (2017) Production of medium chain fatty acids by Yarrowia lipolytica: Combining molecular design and TALEN to engineer the fatty acid synthase. ACS Synth Biol. 6:1870–1879. https://doi.org/10.1021/acssynbio.7b00034
Rigouin C, Croux C, Borsenberger V, Ben Khaled M, Chardot T, Marty A, Bordes F (2018) Increasing medium chain fatty acids production in Yarrowia lipolytica by metabolic engineering. Microb Cell Fact 17:142. https://doi.org/10.1186/s12934-018-0989-5
Romero AM, Jordá T, Rozès N, Martínez-Pastor MT, Puig S (2018) Regulation of yeast fatty acid desaturase in response to iron deficiency. Biochim Biophys Acta Mol Cell Biol Lipids 1863:657–668. https://doi.org/10.1016/j.bbalip.2018.03.008
Saini R, Hegde J, Brar SK, Vezina P (2020) Advanced biofuel production and road to commercialization: An insight into bioconversion potential of Rhodosporidium sp. Biomass Bioenergy 132:105439. https://doi.org/10.1016/j.biombioe.2019.105439
Saini RK, Prasad P, Sreedhar RV, Akhilender Naidu K, Shang X, Keum YS (2021) Omega-3 polyunsaturated fatty acids (PUFAs): Emerging plant and microbial sources, oxidative stability, bioavailability, and health benefits—a review. Antioxidants (Basel) 10:1627. https://doi.org/10.3390/antiox10101627
Saini R, Gonzalez CSO, Hegde K, Brar SK, Vezina P (2022) Evaluating the Effect of Trace metal salts on lipid accumulation ability of Rhodosporidium toruloides-1588 using wood hydrolysate as a carbon source. Bioenerg Res. https://doi.org/10.1007/s12155-022-10521-2
Saini R, Osorio-Gonzalez CS, Hegde K, Brar SK, Vezina P (2022b) Investigating the ability of Rhodosporidium toruloides-1588 to use furfural as a carbon source and its degradation: an enzyme identification study. Sustain Energy Fuels 6:4331–4337. https://doi.org/10.1039/D2SE00772J
Schultz JC, Cao M, Zhao H (2019) Development of a CRISPR/Cas9 system for high efficiency multiplexed gene deletion in Rhodosporidium toruloides. Biotechnol Bioeng 116:2103–2109. https://doi.org/10.1002/bit.27001
Shaah MAH, Hossain MS, Allafi FAS, Alsaedi A, Ismail N, Ab Kadir MO, Ahmad MI (2021) A review on non-edible oil as a potential feedstock for biodiesel: Physicochemical properties and production technologies. RSC Adv 11:25018–25037. https://doi.org/10.1039/D1RA04311K
Singh D, Barrow CJ, Puri M, Tuli DK, Mathur AS (2016) Combination of calcium and magnesium ions prevents substrate inhibition and promotes biomass and lipid production in thraustochytrids under higher glycerol concentration. Algal Res 15:202–209. https://doi.org/10.1016/j.algal.2016.02.024
Singh D, Sharma D, Soni SL, Sharma S, Sharma PK, Jhalani A (2020a) A review on feedstocks, production processes, and yield for different generations of biodiesel. Fuel 262:116553. https://doi.org/10.1016/j.fuel.2019.116553
Singh K, Graf B, Linden A, Sautner V, Urlaub H, Tittmann K, Stark H, Chari A (2020b) Discovery of a regulatory subunit of the yeast fatty acid synthase. Cell 180:1130–1143. https://doi.org/10.1016/j.cell.2020.02.034
Sperling P, Ternes P, Zank TK, Heinz E (2003) The evolution of desaturases. Prostaglandins Leukot Essent Fatty Acids 68:73–95. https://doi.org/10.1016/s0952-3278(02)00258-2
Sun C, Shah AM, Yang J, Wang Z, Zhu L, Song Y (2021) Transcriptome analysis of oleaginous fungus Mucor circinelloides WJ11 in response to exogenous soybean oil as carbon source. Nat Prod Commun. https://doi.org/10.1177/1934578X211023366
Szczepańska P, Hapeta P, Lazar Z (2022) Advances in production of high-value lipids by oleaginous yeasts. Crit Rev Biotechnol 42:1–22. https://doi.org/10.1080/07388551.2021.1922353
Takaku H, Matsuzawa T, Yaoi K, Yamazaki H (2020) Lipid metabolism of the oleaginous yeast Lipomyces starkeyi. Appl Microbiol Biotechnol 104:6141–6148. https://doi.org/10.1007/s00253-020-10695-9
Temkov M, Mureșan V (2021) Tailoring the structure of lipids, oleogels and fat replacers by different approaches for solving the trans-fat issue—a review. Foods 10:1376. https://doi.org/10.3390/foods10061376
Tezaki S, Iwama R, Kobayashi S, Shiwa Y, Yoshikawa H, Ohta A, Horiuchi H, Fukuda R (2017) Δ12-fatty acid desaturase is involved in growth at low temperature in yeast Yarrowia lipolytica. Biochem Biophys Res Commun 488:165–170. https://doi.org/10.1016/j.bbrc.2017.05.028
Toldrá-Reig F, Mora L, Toldrá F (2020) Trends in biodiesel production from animal fat waste. Appl Sci 10:3644. https://doi.org/10.3390/app10103644
Tsai YY, Ohashi T, Kanazawa T, Polburee P, Misaki R, Limtong S, Fujiyama K (2017) Development of a sufficient and effective procedure for transformation of an oleaginous yeast, Rhodosporidium toruloides DMKU3-TK16. Curr Genet 63:359–371. https://doi.org/10.1007/s00294-016-0629-8
Tsai YY, Ohashi T, Wu CC, Bataa D, Misaki R, Limtong S, Fujiyama K (2019) Delta-9 fatty acid desaturase overexpression enhanced lipid production and oleic acid content in Rhodosporidium toruloides for preferable yeast lipid production. J Biosci Bioeng 127:430–440. https://doi.org/10.1016/j.jbiosc.2018.09.005
Uemura H (2012) Synthesis and production of unsaturated and polyunsaturated fatty acids in yeast: current state and perspectives. Appl Microbiol Biotechnol 95:1–12. https://doi.org/10.1007/s00253-012-4105-1
Uprety BK, Rakshit SK (2018) Use of essential oils from various plants to change the fatty acids profiles of lipids obtained from oleaginous yeasts. J Am Oil Chem Soc 95:135–148. https://doi.org/10.1002/aocs.12006
Uprety BK, Chaiwong W, Ewelike C, Rakshit SK (2016) Biodiesel production using heterogeneous catalysts including wood ash and the importance of enhancing byproduct glycerol purity. Energy Convers Manag 115:191–199. https://doi.org/10.1016/j.enconman.2016.02.032
Uprety BK, Dalli SS, Rakshit SK (2017) Bioconversion of crude glycerol to microbial lipid using a robust oleaginous yeast Rhodosporidium toruloides ATCC 10788 capable of growing in the presence of impurities. Energy Convers Manag 135:117–128. https://doi.org/10.1016/j.enconman.2016.12.071
Uprety BK, Samavi M, Rakshit SK (2018) Contribution of specific impurities in crude glycerol towards improved lipid production by Rhodosporidium toruloides ATCC 10788. Bioresour Technol Rep 3:27–34. https://doi.org/10.1016/j.biteb.2018.05.011
Uprety BK, Morrison EN, Neil Emery RJ, Farrow SC (2022) Customizing lipids from oleaginous microbes: Leveraging exogenous and endogenous approaches. Trends Biotechnol 40:482–508. https://doi.org/10.1016/j.tibtech.2021.09.004
Uthandi S, Kaliyaperumal A, Srinivasan N, Thangavelu K, Muniraj IK, Zhan X, Gathergood N, Gupta VK (2021) Microbial biodiesel production from lignocellulosic biomass: new insights and future challenges. Crit Rev Environ Sci Technol 52:2197–2225. https://doi.org/10.1080/10643389.2021.1877045
Wang Y, Zhang S, Pötter M, Sun W, Li L, Yang X, Jiao X, Zhao ZK (2016) Overexpression of Δ12-fatty acid desaturase in the oleaginous yeast Rhodosporidium toruloides for production of linoleic acid-rich lipids. Appl Biochem Biotechnol 180:1497–1507. https://doi.org/10.1007/s12010-016-2182-9
Wang Y, Zhang S, Zhu Z, Shen H, Lin X, Jin X, Jiao X, Zhao ZK (2018) Systems analysis of phosphate-limitation-induced lipid accumulation by the oleaginous yeast Rhodosporidium toruloides. Biotechnol Biofuels 11:148. https://doi.org/10.1186/s13068-018-1134-8
Wang K, Shi TQ, Lin L, Wei P, Ledesma-Amaro R, Ji XJ (2022) Engineering Yarrowia lipolytica to produce tailored chain-length fatty acids and their derivatives. ACS Synth Biol 11:2564–2577. https://doi.org/10.1021/acssynbio.2c00305
Wang K, Shi TQ, Wang J, Wei P, Ledesma-Amaro R, Ji XJ (2022) Engineering the lipid and fatty acid metabolism in Yarrowia lipolytica for sustainable production of high oleic oils. ACS Synth Biol 11:1542–1554. https://doi.org/10.1021/acssynbio.1c00613
Wen Z, Zhang S, Odoh CK, Jin M, Zhao ZK (2020) Rhodosporidium toruloides—a potential red yeast chassis for lipids and beyond. FEMS Yeast Res. https://doi.org/10.1093/femsyr/foaa038
Wiebe MG, Koivuranta K, Penttilä M, Ruohonen L (2012) Lipid production in batch and fed-batch cultures of Rhodosporidium toruloides from 5 and 6 carbon carbohydrates. BMC Biotechnol 12:26. https://doi.org/10.1186/1472-6750-12-26
Wu S, Hu C, Jin G, Zhao X, Zhao ZK (2010) Phosphate-limitation mediated lipid production by Rhodosporidium toruloides. Bioresour Technol 101:6124–6129. https://doi.org/10.1016/j.biortech.2010.02.111
Wu S, Zhao X, Shen H, Wang Q, Zhao ZK (2011) Microbial lipid production by Rhodosporidium toruloides under sulfate-limited conditions. Bioresour Technol 102:1803–1807. https://doi.org/10.1016/j.biortech.2010.09.033
Wu CC, Tsai YY, Ohashi T, Misaki R, Limtong S, Fujiyama K (2018) Isolation of a thermotolerant Rhodosporidium toruloides DMKU3-TK16 mutant and its fatty acid profile at high temperature. FEMS Microbiol Lett 365. https://doi.org/10.1093/femsle/fny203
Wu CC, Ohashi T, Misaki R, Limtong S, Fujiyama K (2020) Ethanol and H2O2 stresses enhance lipid production in an oleaginous Rhodotorula toruloides thermotolerant mutant L1-1. FEMS Yeast Res. https://doi.org/10.1093/femsyr/foaa030
Wu CC, Ohashi T, Kajiura H, Sato Y, Misaki R, Honda K, Limtong S, Fujiyama K (2021) Functional characterization and overexpression of Δ12-desaturase in the oleaginous yeast Rhodotorula toruloides for production of linoleic acid-rich lipids. J Biosci Bioeng 131:631–639. https://doi.org/10.1016/j.jbiosc.2021.02.002
Yamada R, Kashihara T, Ogino H (2017) Improvement of lipid production by the oleaginous yeast Rhodosporidium toruloides through UV mutagenesis. World J Microbiol Biotechnol 33:99. https://doi.org/10.1007/s11274-017-2269-7
Yang X, Jin G, Gong Z, Shen H, Bai F, Zhao ZK (2014) Recycling biodiesel-derived glycerol by the oleaginous yeast Rhodosporidium toruloides Y4 through the two-stage lipid production process. Biochem Eng J 91:86–91. https://doi.org/10.1016/j.bej.2014.07.015
Ye Z, Sun T, Hao H, He Y, Liu X, Guo M, Chen G (2021) Optimising nutrients in the culture medium of Rhodosporidium toruloides enhances lipids production. AMB Express 11:149. https://doi.org/10.1186/s13568-021-01313-6
Yoshinaga K (2021) Development of analytical methods and nutritional studies using synthetic fatty acids and triacylglycerols. J Oleo Sci 70:1–9. https://doi.org/10.5650/jos.ess20196
Yu AQ, Pratomo Juwono NK, Leong SSJ, Chang MW (2014) Production of fatty acid-derived valuable chemicals in synthetic microbes. Front Bioeng Biotechnol 2:78. https://doi.org/10.3389/fbioe.2014.00078
Zeng Y, Bian D, Xie Y, Jiang X, Li X, Li P, Xie T (2017) Utilization of food waste hydrolysate for microbial lipid and protein production by Rhodosporidium toruloides Y2. J Chem Technol Biotechnol 92:666–673. https://doi.org/10.1002/jctb.5049
Zhang C, Shen H, Zhang X, Yu X, Wang H, Xiao S, Wang J, Zhao ZK (2016) Combined mutagenesis of Rhodosporidium toruloides for improved production of carotenoids and lipids. Biotechnol Lett 38:1733–1738. https://doi.org/10.1007/s10529-016-2148-6
Zhang S, Skerker JM, Rutter CD, Maurer MJ, Arkin AP, Rao CV (2016) Engineering Rhodosporidium toruloides for increased lipid production. Biotechnol Bioeng 113:1056–1066. https://doi.org/10.1002/bit.25864
Zhang Y, Kamal R, Li Q, Yu X, Wang Q, Zhao ZK (2022a) Comparative fatty acid compositional profiles of Rhodotorula toruloides haploid and diploid strains under various storage conditions. Fermentation 8:467. https://doi.org/10.3390/fermentation8090467
Zhang Y, Pang J, Liu S, Nie K, Deng L, Wang F, Liu J (2022) Harnessing transcription factor Mga2 and fatty acid elongases to overproduce palmitoleic acid in Saccharomyces cerevisiae. Biochem Eng J 181:108402. https://doi.org/10.1016/j.bej.2022.108402
Zhao Y, Song B, Li J, Zhang J (2022) Rhodotorula toruloides: an ideal microbial cell factory to produce oleochemicals, carotenoids, and other products. World J Microbiol Biotechnol 38:13. https://doi.org/10.1007/s11274-021-03201-4
Zhou YJ, Buijs NA, Zhu Z, Qin J, Siewers V, Nielsen J (2016) Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat Commun 7:11709. https://doi.org/10.1038/ncomms11709
Zhu Z, Zhang S, Liu H, Shen H, Lin X, Yang F, Zhou YJ, Jin G, Ye M, Zou H, Zhao ZK (2012) A multi-omic map of the lipid-producing yeast Rhodosporidium toruloides. Nat Commun 3:1112. https://doi.org/10.1038/ncomms2112
Funding
Open access funding provided by Osaka University. This study was supported in part by the NEDO Project (Project code: P20011) of the Ministry of Economy, Trade, and Industry (METI), Japan, and by the COI-NEXT Program (Grant number: JPMJPF2106) of Japan Science and Technology (JST).
Author information
Authors and Affiliations
Contributions
WCC, HK and KF discussed the content and direction. WCC wrote the main manuscript text and prepared figures and table.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, CC., Honda, K. & Kazuhito, F. Current advances in alteration of fatty acid profile in Rhodotorula toruloides: a mini-review. World J Microbiol Biotechnol 39, 234 (2023). https://doi.org/10.1007/s11274-023-03595-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-023-03595-3