
Abstract
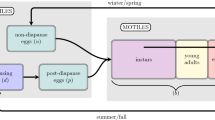
In this paper, we develop a phenologically explicit reaction–diffusion model to analyze the spatial spread of a univoltine insect species. Our model assumes four explicit life stages: adult, two larval, and pupa, with a fourth, implicit, egg stage modeled as a time delay between oviposition and emergence as a larva. As such, our model is broadly applicable to holometabolous insects. To account for phenology (seasonal biological timing), we introduce four time-dependent phenological functions describing adult emergence, oviposition and larval conversion, respectively. Emergence is defined as the per-capita probability of an adult emerging from the pupal stage at a particular time. Oviposition is defined as the per-capita rate of adult egg deposition at a particular time. Two functions deal with the larva stage 1 to larva stage 2, and larva stage 2 to pupa conversion as per-capita rate of conversion at a particular time. This very general formulation allows us to accommodate a wide variety of alternative insect phenologies and lifestyles. We provide the moment-generating function for the general linearized system in terms of phenological functions and model parameters. We prove that the spreading speed of the linearized system is the same as that for nonlinear system. We then find explicit solutions for the spreading speed of the insect population for the limiting cases where (1) emergence and oviposition are impulsive (i.e., take place over an extremely narrow time window), larval conversion occurs at a constant rate, and larvae are immobile, (2) emergence and oviposition are impulsive (i.e., take place over an extremely narrow time window), larval conversion occurs at a constant rate starting at a delayed time from egg hatch, and larvae are immobile, and (3) emergence, oviposition, and larval conversion are impulsive. To consider other biological scenarios, including cases with emergence and oviposition windows of finite width as well as mobile larvae, we use numerical simulations. Our results provide a framework for understanding how phenology can interact with spatial spread to facilitate (or hinder) species expansion. This is an important area of research within the context of global change, which brings both new invasive species and range shifts for native species, all the while causing perturbations to species phenology that may impact the abilities of native and invasive populations to spread.













Similar content being viewed by others

References
Berestycki H, Diekmann O, Nagelkerke CJ, Zegeling PA (2009) Can a species keep pace with a shifting climate? Bull Math Biol 71(2):399–429
Bewick S et al (2016) How resource phenology affects consumer population dynamics. Am Nat 187(2):151–166
Calabrese JM, Fagan WF (2004) Lost in time, lonely, and single: reproductive asynchrony and the allee effect. Am Nat 164:25–37
Etilé E, Despland E (2008) Development variation in the forest tent caterpillar: life history consequences of a threshold size for pupation. Oikos 117(1):135–143
Fagan WF, Bewick S, Cantrell C, Cosner C et al (2014) Phenologically explicit models for species interactions under climate change: a plant-pollinator example. Theor Ecol 7:289–297
Gascoigne J, Berec L, Gregory S, Courchamp F (2009) Dangerously few liaisons: a review of mate-finding Allee effects. Popul Ecol 51:355–372
Gray DR (2004) The gypsy moth life stage model: landscape-wide estimates of gypsy moth establishment using a multi-generational phenology model. Ecol Model 176:155–171
Jepsen JU, Kapari L, Hagen SB, Schott T et al (2011) Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Glob Change Biol 17(6):2071–2083
Johnson DM, Liebhold AM, Tobin PC, Bjrnstad ON (2006) Allee effects and pulsed invasion by the gypsy moth. Nature 444(7117):361–363
Kot M (1992) Discrete-time traveling waves—ecological examples. J Math Biol 30:413–436
Kot M, Lewis MA, VandenDriessche P (1996) Dispersal data and the spread of invading organisms. Ecology 77(7):2027–2042
Lewis MA, Kareiva P (1993) Allee dynamics and the spread of invading organisms. Theor Popul Biol 43(2):141–158
Lewis MA, Li B (2012) Spreading speed, traveling waves, and minimal domain size in impulsive reaction–diffusion models. Bull Math Biol 74:2383–2402
Lewis MA, Schmitz G (1996) Biological invasion of an organism with separate mobile and stationary states: modeling and analysis. Forma 11:1–25
Li B, Weinberger HF, Lewis MA (2005) Spreading speeds as slowest wave speeds for cooperative systems. Math Biosci 196:82–98
Li B et al (2014) Persistence and spread of a species with shifting habitat edge. SIAM J Appl Math 74(5):1397–1417
Li J, Brauer F (2008) Continuous-time age-structured models in population dynamics and epidemiology. In: Brauer F, van den Driessche P, Wu J (eds) Mathematical epidemiology. Springer, New York, pp 205–227
Logan JD (2008) Phenologically-structured predator-prey dynamics with temperature dependence. Bull Math Biol 70(1):1–20
Logan JA, Powell JA (2001) Ghost forests, global warming, and the mountain pine beetle (Coleoptera: Scolytidae). Am Entomol 47(3):160
Lutscher F, Seo G (2011) The effect of temporal variability on persistence conditions in rivers. J Theor Biol 283:53–59
Lutscher F, Van Minh N (2013) Traveling waves in discrete models of biological populations with sessile stages. Nonlinear Anal Real World Appl 14:495–506
Lynch HJ, Rhainds M, Calabrese JM, Cantrell S et al (2014) How climate extremes not means define a species geographic range boundary via a demographic tipping point. Ecol Monogr 84:134–149
Mailleret L (1908) Lemesle V (2009) A note on semi-discrete modelling in the life sciences. Philos Trans R Soc A Math Eng 367:4779–4799
Memmott J, Craze PG, Waser NM, Price MV (2007) Global warming and the disruption of plant–pollinator interactions. Ecol Lett 10(8):710–717
Meyer K, Li B (2013) A spatial model of plants with an age-structured seed bank and juvenile stage. SIAM J Appl Math 73:1676–1702
Miller-Rushing AJ, Hye TT, Inouye DW, Post E (2010) The effects of phenological mismatches on demography. Philos Trans R Soc B Biol Sci 365(1555):3177–3186
Okubo A et al (1989) On the spatial spread of the grey squirrel in britain. Proc R Soc Ser B Biol Sci 238(1291):113–125
Owen MR, Lewis MA (2001) How predation can slow, stop or reverse a prey invasion. Bull Math Biol 63(4):655–684
Pao CV (1992) Nonlinear parabolic and elliptic equations. Plenum Press, New York
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421(6918):37–42
Polyanin A (2002) Handbook of linear partial differential equations for engineers and scientists. Chapman & Hall/CRC, Boca Raton
Post E, Levin SA, Iwasa Y, Stenseth NC (2001) Reproductive asynchrony increases with environmental disturbance. Evolution 55(4):830–834
Rhainds M, Fagan WF (2010) Broad-scale latitudinal variation in female reproductive success contributes to the maintenance of a geographic range boundary in bagworms (Lepidoptera: Psychidae). PLoS ONE 5:e14166
Robinet C, Lance DR, Thorpe KW, Onufrieva KS et al (2008) Dispersion in time and space affect mating success and Allee effects in invading gypsy moth populations. J Anim Ecol 77:966–973
Robinet C, Liebhold A, Gray D (2007) Variation in developmental time affects mating success and Allee effects. Oikos 116:1227–1237
Seo G, Lutscher F (2011) Spread rates under temporal variability: calculation and applications to biological invasions. Math Models Methods Appl Sci 21(12):2469–2489
Tobin PC, Whitmire SL, Johnson DM, Bjrnstad ON et al (2007) Invasion speed is affected by geographical variation in the strength of Allee effects. Ecol Lett 10(1):36–43
Visser ME, Both C (2005) Shifts in phenology due to global climate change: the need for a yardstick. Proc R Soc B Biol Sci 272(1581):2561–2569
Visser ME, Holleman LJ, Gienapp P (2006) Shifts in caterpillar biomass phenology due to climate change and its impact on the breeding biology of an insectivorous bird. Oecologia 147(1):164–172
Walter JA, Meixler MS, Mueller T, Fagan WF et al (2014) How topography induces reproductive asynchrony and alters gypsy moth invasion dynamics. J Anim Ecol 84(1):188–198
Ward NL, Masters GJ (2007) Linking climate change and species invasion: an illustration using insect herbivores. Glob Change Biol 13(8):1605–1615
Weinberger HF, Lewis MA, Li B (2002) Analysis of linear determinacy for spread in cooperative models. J Math Biol 45:183–218
Zhou Y, Kot M (2011) Discrete-time growth-dispersal models with shifting species ranges. Theor Ecol 4:13–25
Author information
Authors and Affiliations
Corresponding author
Additional information
Garrett Otto: This research was partially supported by the National Science Foundation under Grants DMS-1225693 and DMS-1515875. Sharon Bewick and William F. Fagan: This research was partially supported by the National Science Foundation under Grant DMS-1225917. Bingtuan Li: This research was partially supported by the National Science Foundation under Grants DMS-1225693 and DMS-1515875.
Appendices
Appendix A: Proof of Theorem 3.1
We demonstrate that model (1) satisfies Hypotheses 2.1 and the conditions given in Theorem 3.1 in Weinberger et al. (2002), so that the spreading speed of the nonlinear model (1) is the same as that of its linearization.
We show that P(T, x) in (1) is mathematically well defined. There are two cases: case (i) where \(g(t),\,r(t),\,m_i(t)\) are all bounded and piecewise continuous functions, and case (ii) where one of \(g(t),\,r(t),\,m_i(t)\) is a Dirac delta distribution. We first consider case (i) and assume that all phenological functions are bounded and continuous functions. If \(P_n(x)\) is continuous and bounded, Theorem 2.2 in Pao (1992) implies that the first equation in (1) has a unique solution A(t, x) which is continuous and bounded in t and x. The same theorem then implies that the second, third, and fourth equation of (1) have unique solutions \(L_1(t,x)\), \(L_2(t,x)\) and P(t, x) that are continuous and bounded in t and x, respectively, and thus \(P_{n+1}( x)=P(T, x)\) is bounded and continuous in x. Since \(P_0(x)\) is continuous and bounded, induction shows the continuity and boundedness of \(P_n(x)\) for all \(n\ge 1\). If g(t) is piecewise continuous and discontinuous at \(0<s_1<s_2<\cdot \cdot \cdot<s_{\ell }<T\), the first equation of (1) shows the existence, uniqueness, and boundedness of A(t, x) for \(0<t\le s_1\). One can then consider the first equation of (1) for \(s_1<t\le s_2\) with the initial value \(A(s_1, x)\) at \(s_1\) to establish the existence, uniqueness, and boundedness of A(t, x) for \(s_1<t\le s_2\). Induction shows the existence, uniqueness, and boundedness of A(t, x) for \(0<t\le T\). The same method can be used to determine the existence, uniqueness, and boundedness of \(L_i(t, x)\) and P(t, x) for \(\tau <t\le T\) if r(t) or \(m_i(t)\) is piecewise continuous.
We now consider case (ii). If one of \(g(t),\,r(t),\,m_i(t)\) is a Dirac delta function, the corresponding equation is converted into an autonomous equation with an appropriate initial condition.
If \(g(t)=\delta (t-t_{emg})\), then the equation for A(t, x) in (1) is equivalent to the following classical system:
Theorem 2.2 in Pao (1992) again works to show the existence and uniqueness of a continuous and bounded solution for (1).
Similarly, if \(r(t)=\alpha _2\delta (t-t_o)\) (\(t_o>t_{emg}\) if \(g(t)=\delta (t-t_{emg})\)), then the equation for \(L_1(t,x)\) in (1) is equivalent to the following classical system:
For \(m_1(t)=\gamma _1\,\delta (t-t_{l_1})\), pupation occurs impulsively at time \(t_{l_1}\) and therefore \(L_1(t,x)\) is not continuous with respect to time at \(t=t_{l_1}\). In this case, the \(L_1\) and \(L_2\) equations in (1) may be viewed as
To see \(L_1(t_{l_1}^+,x)=e^{-\gamma _1}L_1(t_{l_1}^-,x)\), in (1), we replace \(m_1(t)\) by its approximation \(m_\lambda (t)=\frac{m_0}{\lambda }\phi \left( \frac{t-t_3}{\lambda }\right) \) where \(\phi \) is a nonnegative bounded continuous function with support \([-1,1]\) and \(\int ^{1}_{-1}\phi (s)\mathrm {d}s=1\), integrate the \(L_1\) equation from \(t^-\) to \(t^+\) with \(t^-<t_{l_1}<t^+\), and use the boundedness and second order differentiability of the solution and take the limit as \(\lambda {\rightarrow }0^+\), \(t^-{\rightarrow }t^-_3\) and \(t^+{\rightarrow }t^+_3\).
On the other hand, since all stage 1 larva that defect at \(t_{l_1}\) are assumed to convert to stage 2 larva, \(L_2(t_{l_1}^+,x)=(1-e^{-\gamma _1})L_1(t_{l_1}^-,x)\).
For the case that \(m_2(t)=\gamma _2\,\delta (t-t_{l_2})\), the result is analogous to that for \(m_1(t)=\gamma _1\,\delta (t-t_{l_1})\), with \(P(t_{l_2}^+,x)=\big (1-e^{-\gamma _2}\big )\,L_2(t_{l_2}^-,x)\).
We have justified that the solution to model (1) maps the pupa density distribution at the end of the production season from year n, \(P_{n}(x)\), to the pupa density distribution at the end of the production season from year \(n+1\), \(P_{n+1}(x)\). Mathematically model (1) can be rewritten as an abstract discrete equation in the form of \(P_{n+1}(x)=Q[P_{n}](x)\) with \(Q[P_{n}](x)\) determined by P(T, x).
Clearly, \(P_0(x)\equiv 0\) implies \(P_n(x)\equiv 0\) for all n. That is \(Q[0]=0\). The comparison principals for scalar parabolic equations and ordinary differential equations show that \(Q[p](x)\ge Q[q](x)\) if \(p(x)\ge q(x)\ge 0\). That is, Q is a monotone operator. One can easily see that for the ODE model corresponding to (1), a sufficiently large number M is an upper solution. This M is also an upper solution for the PDE model (1), i.e., \(Q[M]\le M\). It follows from elementary properties of parabolic systems that Q is translation and reflection invariant, and Q is continuous in the topology of uniform convergence on bounded sets. Hypotheses 2.1 (i)–(iv) given in Weinberger et al. (2002) are satisfied by Q if Q has a positive equilibrium.
The linearized system of model (1) is given by
To find a solution to model (8) subject to same initial conditions as model (1), we will solve for A(t, x) and then sequentially work our way to P(t, x). An explanation of how to solve this equation using integral methods can be found in Polyanin (2002). From the first equation of model (8) with initial pupal population \(P_n(x)\), we find
Substituting this result into the second equation of (8) gives the first stage larval population for the system:
where \(M_1(t)\) is an anti-derivative of \(m_1(t)\). The second stage larva density is given by
where \(M_2(t)\) is an anti-derivative of \(m_2(t)\). Finally, the pupal density at time T is given by
Equation (12) describes the number of larvae that successfully mature into pupae by the end of the year. Because it is also proportional to the emerging adult population in the following year, Eqs. (9)–(12) completely describe the species life cycle. Consequently, these equations can be combined to give the following year-to-year linear mapping
The right-hand side of this recursion defines an linear operator R. That is, (13) can be written as \(P_{n+1}(x)=R[P_n](x)\). As indicated in Weinberger et al. (2002), the moment-generating function for R is then given by:
The zero solution for R is unstable if \(\Lambda (0)>1\).
Assume that \(\Lambda (0)>1\). For any positive integer \(\kappa >1\), we define \(R^{(\kappa )}[P_n](x)\) to be the solution operator of the linear system (8) with the death rate \(\nu _i\) replaced by \(\nu ^{(\kappa )}_i=\nu _i+\kappa ^{-1}\). Let \(\Lambda ^{(\kappa )}(\mu )=R^{(\kappa )}[e^{-\mu x}](0)\). It is easily seen that \(\Lambda ^{(\kappa )}(\mu )\) is given by \(\Lambda (\mu )\) with \(\nu _i\) replaced by \(\nu ^{(\kappa )}_i\) for \(i\in \{a,\,l_1,\,l_2,\,p\}\). Clearly, the limit of \(\Lambda ^{(\kappa )}(\mu )\) as \(\kappa \) approaches \(\infty \) is \(\Lambda (\mu )\). For any \(\kappa >1\), there is \(\delta _{\kappa }>0\) such that for any \(0<\alpha \le \delta _{\kappa }\) such that
We now observe that \( R^{(\kappa )}[\delta _{\kappa }]=\Lambda ^{(\kappa )}(0)\delta _{\kappa }\). Since \(\Lambda (0)>1\), \(\Lambda ^{(\kappa )}(0)>1\) for large \(\kappa \). Choose \(\omega =\delta /\Lambda ^{(\kappa )}(0)\). Then, a standard comparison theorem shows that for \(0\le v(x) \le \omega \), \(Q[v](x)\ge R^{(\kappa )}[v](x)\). We have verified Hypothesis 2.1.vi in Weinberger et al. (2002). Hypothesis 2.1.v in the paper is automatically satisfied. On the other hand, \(Q[v](x)\ge R^{(\kappa )}[v](x)\) for \(0\le v(x) \le \omega \) implies that for small positive \(\alpha \), \(Q[\alpha ]>\alpha \). This together with \(Q[M]\le M\) shows that Q has a positive equilibrium \(P^*\) satisfying \(Q[P^*]=P^*\). We have shown that Hypotheses 2.1 in Weinberger et al. (2002) hold if \(\Lambda (0)>1\).
In model (1), the nonlinear quadratic terms are all non-positive. This implies that Q is dominated by R, and thus for the scalar operator Q the conditions in Theorem 3.1 in Weinberger et al. (2002) are automatically satisfied. It follows from this theorem that the spreading speed of Q is the same as that of R, which is given by
if \(\Lambda (0)>1\). On the other hand, since at least one diffusion coefficient is positive, the basic properties of reaction–diffusion equations show that the operator P is compact in the sense that every sequence \(v_n(x)\) with \(0\le v_n(x)\le P^*\) has a subsequence \(v_{n_{\ell }}(x)\) such that \(P[v_{n_{\ell }} ](x)\) converges uniformly on every bounded set. It follows from Theorem 3.1 in Li et al. (2005) that \(c^*\) is the slowest speed of a class of traveling wave solutions connecting 0 with \(P^*\).
Appendix B: Spreading Speed for Impulsive Emergence, Oviposition with no Larval Dispersal, and Constant Pupation Rate
We define \(g,\,r,\,m_1,\,m_2\) as
and set \(d_{l_1}=d_{l_2}=0\). Applying this to Eq. (4), we arrive at the following integral for the moment-generating function,
The \(\mathrm {d}s_1\) and \(\mathrm {d}{s_2}\) integrals are straightforward with the non-delta portion of the integrand being evaluated at \(s_1=0\) and \(s_2=t_o+\tau \), respectively. The lower bounds of the \(\mathrm {d}s_3\) and \(\mathrm {d}s_4\) integral must be replaced with \(t_o+\tau \), and then these integrals can be carried out in a straightforward manner. After some simplification, it is found that
Invoking the non-extinction condition, \(\Lambda (0)>1\), we find the parameters must satisfy
To find \(c^*\), we must find \(\inf _{\mu >0}\,\frac{\ln (\Lambda (\mu ))}{\mu }\). Taking the natural log of \(\Lambda (\mu )\), we find
where
From elementary calculus, we find \(c^*=\inf _{\mu >0}\,\frac{A+B\mu ^2}{\mu }=2\sqrt{AB}\), and thus
Appendix C: Spreading Speed for Impulsive Emergence, Oviposition, and Larval Conversion
To evaluate the moment generator, we will need to employ the following lemma:
Lemma 4.1
Suppose \(f:{\mathbb {R}}\mapsto {\mathbb {R}}\) is continuous at \(x=0\), and \(g(x):{\mathbb {R}}\mapsto {\mathbb {R}}\) is continuous and nonnegative on [0, 1] then:
Proof
Let \(\phi (x)\) be any function with the following properties: \(\phi (x)\in \mathbf{C }({\mathbb {R}})\), \(\phi (x)\) is nonnegative, \(\phi (x)=0\) if \(|x|\ge 1\), \(\int _{-\infty }^{\infty }\!\phi (x)\,\mathrm {d}x=1.\)
We define \(\phi _{\lambda }(x)=\frac{1}{\lambda }\,\phi \left( \frac{x}{\lambda }\right) \) for \(\lambda >0\). We note the support of \(\phi _{\lambda }(x)\) is a subset of \([-\lambda ,\lambda ]\), and \(\int _{-\infty }^{\infty }\!\phi _{\lambda }(x)\,\mathrm {d}x=1\). Using substitution, we find \(\forall \,\lambda >0\):
Let \(\epsilon >0\), then by continuity of f(x) at \(x=0\) there \(\exists \;\bar{\lambda }\) such that if \(x\in [-\bar{\lambda },\bar{\lambda }]\), then
We note:
We note if \(0<\lambda <\bar{\lambda }\), then:
Since \(\epsilon \) can be chosen to be arbitrarily small, we finally see:
thus concluding the proof of the Lemma.
We define \(g(t)=\delta (t)\), \(r(t)=\alpha _2e^{-\nu _e\tau }\,\delta (t-t_2)\), \(m_1(t)=\gamma _1\,\delta (t-t_{l_1})\), \(m_2(t)=\gamma _2\,\delta (t-t_{l_2})\). We further assume \(t_o+\tau<t_{l_1}<t_{l_2}<T\), if this condition is not held it easy to see that the population will face extinction as the larvae will never have an opportunity to pupate. It follows from Eq. (4) that:
The \(\mathrm {d}s_1,\,\mathrm {d}s_2\) integrals are straightforward, with \(s_1,\,s_2\) being replaced by \(0,\,t_o+\tau \), respectively, and replacing the lower bounds of the \(\mathrm {d}s_4,\,\mathrm {d}s_3\) integrals with \(t_o+\tau \). The \(\mathrm {d}s_4\) and \(\mathrm {d}s_3\) integral are more complicated and require subsequent applications of Lemma 4.1, with the \(g(\cdot )\) of Lemma 4.1 being identified as \(\exp (-\gamma _1\,\varvec{\cdot }),\;\exp (-\gamma _2\,\varvec{\cdot })\) respectively.
The moment generator thus becomes
The condition of positive spread speed, \(\Lambda (0)>1\), is equivalent to
Taking the natural log of \(\Lambda (\mu )\), we find
We can thus rewrite \(\frac{\ln (\Lambda (\mu ))}{\mu }\) as
where \(A,\,B\) are defined by
If \(\Lambda (0)>1\), then \(A>0\), while B is always positive. It then follows from elementary calculus that
Therefore,
\(\square \)
Appendix D: Numerical Validation of Results in Sect. 3.2
As a further validation of the analytic result for \(c^*\) in Eq. (5), we numerically integrate equations in model (1), starting with a smooth compact piecewise polynomial distribution. For subsequent years, we generate the end of the year pupa population densities \(P_n(x)\) function. We then define \(\epsilon \) to be some small fraction of the equilibrium population, and for year n we define \(x_n>0\) such that \(P_n(x_n)=\epsilon \). Taking a linear least square fit to the tuples \((n,x_n)\) and rejecting \((0,x_0)\) through \((5,x_5)\) so as to not include transients, we are able to numerically approximate the spreading speed as the slope of the fitted line. We summarize the results in Fig. 14. The value of the spreading speed obtained from the slope of fitted line is 1.06, a \(6\%\) relative error of the analytically determined speed 1 given by Eq. (5).

Wave propagation and spreading speed. Parameter values are chosen as \(d_a = 1,\ T= 1.23,\ t_o = 0.23,\ m_1=m_2=3,\ \nu _a=\nu _e=\nu _{l_1} =\nu _{l_2}=1,\ \nu _p=0.5,\ \alpha _1 =0.8,\ \alpha _2 =12.5,\ \beta _a =0.4,\ \beta _{l_1}=\beta _{l_2}=0.06,\ \tau =.01\) and \(\epsilon =5\). The slope of the fitted line is 1.06, the calculated speed is 1.00. We see on panel b that the position of the threshold population is nearly linear with time. a Wave fronts for even years, b \((n, x_n)\) and fitted line (Color figure online)
Rights and permissions
About this article

Cite this article
Otto, G., Bewick, S., Li, B. et al. How Phenological Variation Affects Species Spreading Speeds. Bull Math Biol 80, 1476–1513 (2018). https://doi.org/10.1007/s11538-018-0409-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11538-018-0409-3