
Abstract
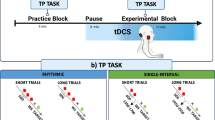
In the present study, we examined the role of the cerebellum in temporal adaptive learning during a coincident timing task, i.e., a baseball-like hitting task involving a moving ball presented on a computer monitor. The subjects were required to change the timing of their responses based on imposed temporal perturbations. Using paired-pulse transcranial magnetic stimulation, we measured cerebellar brain inhibition (CBI) before, during, and after the temporal adaptive learning. Reductions in CBI only occurred during and after the temporal adaptive learning, regardless of the direction of the temporal perturbations. In addition, the changes in CBI were correlated with the magnitude of the adaptation. Here, we showed that the cerebellum is essential for learning about and controlling the timing of movements during temporal adaptation. Furthermore, changes in cerebellar-primary motor cortex connectivity occurred during temporal adaptation, as has been previously reported for spatial adaptation.






Similar content being viewed by others

References
Aso K, Hanakawa T, Aso T, Fukuyama H (2010) Cerebro-cerebellar interactions underlying temporal information processing. J Cogn Neurosci 22:2913–2925. https://doi.org/10.1162/jocn.2010.21429
Baarbe J, Yielder P, Daligadu J, Behbahani H, Haavik H, Murphy B (2014) A novel protocol to investigate motor training-induced plasticity and sensorimotor integration in the cerebellum and motor cortex. J Neurophysiol 111:715–721. https://doi.org/10.1152/jn.00661.2013
Bares M, Lungu OV, Liu T, Waechter T, Gomez CM, Ashe J (2007) Impaired predictive motor timing in patients with cerebellar disorders. Exp Brain Res 180:355–365. https://doi.org/10.1007/s00221-007-0857-8
Bares M, Lungu OV, Husarova I, Gescheidt T (2010) Predictive motor timing performance dissociates between early diseases of the cerebellum and Parkinson’s disease. Cerebellum 9:124–135. https://doi.org/10.1007/s12311-009-0133-5
Bares M, Lungu OV, Liu T, Waechter T, Gomez CM, Ashe J (2011) The neural substrate of predictive motor timing in spinocerebellar ataxia. Cerebellum 10:233–244. https://doi.org/10.1007/s12311-010-0237-y
Bares M, Apps R, Avanzino L, Breska A, D’Angelo E, Filip P, Gerwig M, Ivry RB, Lawrenson CL, Louis ED, Lusk NA, Manto M, Meck WH, Mitoma H, Petter EA (2019) Consensus paper: Decoding the contributions of the cerebellum as a time machine. From Neurons to Clinical Applications. Cerebellum 18:266–286. https://doi.org/10.1007/s12311-018-0979-5
Buhusi CV, Meck WH (2005) What makes us tick? Functional and neural mechanisms of interval timing. Nat Rev Neurosci 6:755–765. https://doi.org/10.1038/nrn1764
Cantarero G, Spampinato D, Reis J, Ajagbe L, Thompson T, Kulkarni K, Celnik PA (2015) Cerebellar direct current stimulation enhances on-line motor skill acquisition through an effect on accuracy. J Neurosci 35:3285–3290. https://doi.org/10.1523/JNEUROSCI.2885-14.2015
Celnik PA (2015) Understanding and modulating motor learning with cerebellar stimulation. Cerebellum 14:171–174. https://doi.org/10.1007/s12311-014-0607-y
Coull JT, Nobre AC (2008) Dissociating explicit timing from temporal expectation with fMRI. Curr Opin Neurobiol 18:137–144. https://doi.org/10.1016/j.conb.2008.07.011
Donchin O, Rabe K, Diedrichsen J, Lally N, Schoch B, Gizewski ER, Timmann D (2012) Cerebellar regions involved in adaptation to force field and visuomotor perturbation. J Neurophyiol 107:134–147. https://doi.org/10.1152/jn.00007.2011
Fernandez L, Major BP, Teo WP, Byrne LK, Enticott PG (2018) Assessing cerebellar brain inhibition (CBI) via transcranial magnetic stimulation (TMS): a systematic review. Neurosci Biobehav Rev 86:176–206. https://doi.org/10.1016/j.neubiorev.2017.11.018
Galea JM, Vazquez A, Pasricha N, de Xivry JJ, Celnik PA (2011) Dissociating the roles of the cerebellum and motor cortex during adaptive learning: the motor cortex retains what the cerebellum learns. Cereb Cortex 21:1761–1770. https://doi.org/10.1093/cercor/bhq246
Gerwig M, Hajjar K, Dimitrova A, Maschke M, Kolb FP, Frings M, Thilmann AF, Forsting M, Diener HC, Timmann D (2005) Timing of conditioned eyeblink responses is impaired in cerebellar patients. J Neurosci 25:3919–3931. https://doi.org/10.1523/JNEUROSCI.0266-05.2005
Hardwick RM, Celnik PA (2014) Cerebellar direct current stimulation enhances motor learning in older adults. Neurobiol Aging 35:2217–2221. https://doi.org/10.1016/j.neurobiolaging.2014.03.030
Hardwick RM, Lesage E, Miall RC (2014) Cerebellar transcranial magnetic stimulation: the role of coil geometry and tissue depth. Brain Stimul 7:643–649. https://doi.org/10.1016/j.brs.2014.04.009
Hirano M, Kubota S, Tanabe S, Koizume Y, Funase K (2015) Interactions among learning stage, retention, and primary motor cortex excitability in motor skill learning. Brain Stimul 8:1195–1204. https://doi.org/10.1016/j.brs.2015.07.025
Hore J, Timmann D, Watts S (2002) Disorders in timing and force of finger opening in overarm throws made by cerebellar subjects. Ann N Y Acad Sci 978:1–15. https://doi.org/10.1111/j.1749-6632.2002.tb07551.x
Ivry RB (1996) The representation of temporal information in perception and motor control. Curr Opin Neurobilo 6:851–857. https://doi.org/10.1016/S0959-4388(96)80037-7
Ivry RB, Keele SW (1989) Timing functions of the cerebellum. J Cogn Neurosci 1:136–152. https://doi.org/10.1162/jocn.1989.1.2.136
Ivry RB, Keele SW, Diener HC (1988) Dissociation of the lateral and medial cerebellum in movement timing and movement execution. Exp Brain Res 73:167–180. https://doi.org/10.1007/BF00279670
Ivry RB, Spencer RM, Zelaznik HN, Diedrichsen J (2002) The cerebellum and event timing. Ann N Y Acad Sci 978:302–317. https://doi.org/10.1111/j.1749-6632.2002.tb07576.x
Izawa J, Criscimagna-Hemminger SE, Shadmehr R (2012) Cerebellar contributions to reach adaptation and learning sensory consequences of action. J Neurosci 32:4230–4239. https://doi.org/10.1523/JNEUROSCI.6353-11.2012
Jayaram G, Galea JM, Bastian AJ, Celnik PA (2011) Human locomotor adaptive learning is proportional to depression of cerebellar excitability. Cereb Cortex 21:1901–1909. https://doi.org/10.1093/cercor/bhq263
Kassavetis P, Hoffland BS, Saifee TA, Bhatia KP, Van De Warrenburg BP, Rothwell JC, Edwards MJ (2011) Cerebellar brain inhibition is decreased in active and surround muscles at the onset of voluntary movement. Exp Brain Res 209:437–442. https://doi.org/10.1007/s00221-011-2575-5
Markova L, Bares M, Lungu OV, Filip P (2020) Quantitative but not qualitative performance changes in predictive motor timing as a result of overtraining. Cerebellum 19:201–207. https://doi.org/10.1007/s12311-019-01100-x
Morton SM, Bastian AJ (2006) Cerebellar contributions to locomotor adaptations during splitbelt treadmill walking. J Neurosci 26:9107–9116. https://doi.org/10.1523/JNEUROSCI.2622-06.2006
Oldfield RC (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113. https://doi.org/10.1016/0028-3932(71)90067-4
O’Reilly JX, Mesulam MM, Nobre AC (2008) The Cerebellum predicts the timing of perceptual events. J Neurosci 28:2252–2260. https://doi.org/10.1523/JNEUROSCI.2742-07.2008
Penhune VB, Steele CJ (2012) Parallel contributions of cerebellar, striatal and M1 mechanisms to motor sequence learning. Behav Brain Res 226:579–591. https://doi.org/10.1016/j.bbr.2011.09.044
Pinto D, Chen R (2001) Suppression of the motor cortex by magnetic stimulation of the cerebellum. Exp Brain Res 4:505–510. https://doi.org/10.1007/s002210100862
Prsa M, Their P (2011) The role of the cerebellum in saccadic adaptation as a window into neural mechanisms of motor learning. Eur J Neurosci 33:2114–2128. https://doi.org/10.1111/j.1460-9568.2011.07693.x
Rabe K, Livne O, Gizewski ER, Aurich V, Beck A, Timmann D, Donchin O (2009) Adaptation to visuomotor rotation and force field perturbation is correlated to different brain areas in patients with cerebellar degeneration. J Neurophysiol 101:1961–1971. https://doi.org/10.1152/jn.91069.2008
Schlerf JE, Spencer RMC, Zelaznik HN, Ivry RB (2007) Timing of rhythmic movements in patients with cerebellar degeneration. Cerebellum 6:221–231. https://doi.org/10.1080/14734220701370643
Schlerf JE, Galea JM, Bastian AJ, Celnik PA (2012) Dynamic modulation of cerebellar excitability for abrupt, but not gradual, visuomotor adaptation. J Neurosci 32:11610–11617. https://doi.org/10.1523/JNEUROSCI.1609-12.2012
Schlerf JE, Xu J, Klemfuss NM, Griffiths TL, Ivry RB (2013) Individuals with cerebellar degeneration show similar adaptation deficits with large and small visuomotor errors. J Neurophyiol 109:1164–1173. https://doi.org/10.1152/jn.00654.2011
Spampinato DA, Celnik PA (2018) Deconstructing skill learning and its physiological mechanisms. Cortex 104:90–102. https://doi.org/10.1016/j.cortex.2018.03.017
Spampinato DA, Celnik PA (2020) Multiple motor learning processes in processes in humans: defining their neurophysiological bases. The Neuroscientist. https://doi.org/10.1177/1073858420939552
Spampinato DA, Block HJ, Celnik PA (2017) Cerebellar-M1 connectivity changes associated with motor learning are somatotopic specific. J Neurosci 37:2377–2386. https://doi.org/10.1523/JNEUROSCI.2511-16.2017
Spampinato DA, Celnik PA, Rothwell JC (2020) Cerebellar-motor cortex connectivity: one or two different networks? J Neurosci 40:4230–4239. https://doi.org/10.1523/JNEUROSCI.2397-19.2020
Torriero S, Oliveri M, Koch G, Lo Gerfo E, Ferlazzo SS, F, Caltagirone C, Petrosini L, (2011) Changes in cerebello-motor connectivity during procedural learning by actual execution and observation. J Cogn Neurosci 23:338–348. https://doi.org/10.1162/jocn.2010.21471
Uehara S, Mawase F, Celnik PA (2018) Learning similar actions by reinforcement sensory-prediction errors rely on distinct physiological mechanisms. Cereb Cortex 28:3478–3490. https://doi.org/10.1093/cercor/bhx214
Ugawa Y, Uesaka Y, Terao Y, Hanajima R, Kanazawa I (1995) Magnetic stimulation over the cerebellum in humans. Ann Neurol 37:703–713. https://doi.org/10.1002/ana.410370603
Acknowledgements
We thank Dr. M. Shin-ya for his helpful comments regarding the custom-made program used for the CoIT task. Dr. M. Hirano was supported as a Research Fellow of the Japan Society for the Promotion of Science (15J03540). The authors thank Medical English Service Co., Ltd., Kyoto, Japan, for their help with the English language editing. This study was supported by Technology of Japan and a Grant-in-aid for scientific research (19K11578) from the Japanese Society for the Promotion of Science.
Author information
Authors and Affiliations
Contributions
ST, MH, and KF conceived and designed the research; ST performed the experiments; ST analyzed the data; ST, MH, and KF interpreted the results of the experiments; ST prepared the figures; ST and KF drafted the manuscript; and ST, HM, and KF approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Additional information
Communicated by Winston D Byblow.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
221_2020_5963_MOESM1_ESM.tif
Supplementary file1 Supplementary Fig. 1. The relationships between the CT and CE in each of the temporal perturbation conditions (A: green: cond F, blue: cond S, gray: ctrl). Linear regression lines for the relationships between the CE and CT in each of the conditions are shown. The linear regression lines were obtained from another 900 trials (300 trials conducted in each of the three conditions) performed by one of the authors, and we used them to recalculate the CE based on the CT if the subjects swung and missed the ball. The slope of the regression line became steeper as the rate of change in the swing speed increased (TIF 83 KB)
Rights and permissions
About this article

Cite this article
Tanaka, Sy., Hirano, M. & Funase, K. Modulation of cerebellar brain inhibition during temporal adaptive learning in a coincident timing task. Exp Brain Res 239, 127–139 (2021). https://doi.org/10.1007/s00221-020-05963-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-020-05963-z