
Abstract
Many of the recently emerging highly virulent zoonotic diseases have a likely bat origin, for example Hendra, Nipah, Ebola and diseases caused by coronaviruses. Presumably because of their long history of coevolution, most of these viruses remain subclinical in bats, but have the potential to cause severe illnesses in domestic and wildlife animals and also humans. Spillovers from bats to humans either happen directly (via contact with infected bats) or indirectly (via intermediate hosts such as domestic or wildlife animals, by consuming food items contaminated by saliva, faeces or urine of bats, or via other environmental sources). Increasing numbers of breakouts of zoonotic viral diseases among humans and livestock have mainly been accounted to human encroachment into natural habitat, as well as agricultural intensification, deforestation and bushmeat consumption. Persecution of bats, including the destruction of their roosts and culling of whole colonies, has led not only to declines of protected bat species, but also to an increase in virus prevalence in some of these populations. Educational efforts are needed in order to prevent future spillovers of bat-borne viruses to humans and livestock, and to further protect bats from unnecessary and counterproductive culling.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Over the past decades, the emergence of zoonotic viruses (those that are naturally transmitted between vertebrate animals and humans ) from bats has been the subject of increasing attention from both scientists and the general public (e.g. Quammen 2013). During outbreaks of diseases in humans and livestock, bats are now often the primary focus of searches for a reservoir host (Chua et al. 2002a; Leroy et al. 2005; Li et al. 2005; Halpin et al. 2007; Towner et al. 2007; Lau et al. 2010; Wibbelt et al. 2010; Memish et al. 2013). Identification of bats as natural host s for emerging viruses has important implications for bat conservation. We review the current state of research of four important families of emerging zoonotic viruses for which bats are natural reservoir hosts and discuss direct and indirect conservation implications.
2 Emerging Viral Diseases: Why Bats?
Although bats have been identified as carriers of many highly virulent human pathogens (Chen et al. 2014), evidence of pathogen-related clinical signs or disease in bats is scarce, particularly for intracellular pathogens such as viruses (Brook and Dobson 2015). Post-infection survival is supported by the frequent identification of antibodies to known viruses in apparently healthy bats and long-term survival of these bats (e.g. Hayman et al. 2010). Additionally, viruses isolated or genetically detected from bat populations are highly diverse and often ancestral to related viruses in human and other mammalian species (e.g. Towner et al. 2009; Drexler et al. 2012; Baker et al. 2013a; Tong et al. 2013; Vidgen et al. 2015). Together, these findings suggest a long history of coevolution between many bat-virus relationships identified to date. Recent progress in the field of bat immunology and genomics has identified key differences in bat immunity and physiology that evolved concomitantly with the evolution of flight , resulting not only in apparently increased immunotolerance of intracellular pathogens, but also in increased longevity and decreased tumour production (Baker et al. 2013b; Zhang et al. 2013; Brook and Dobson 2015). Immunotolerance and incomplete clearance of viral infections are also likely to favour the establishment of persistent infections (Virgin et al. 2009), as proposed for a number of bat-borne viruses (Plowright et al. 2015).
Various ecological and life-history factors play a key role in the susceptibility of individuals and populations to pathogens (Allen et al. 2009; Turmelle et al. 2010; Schneeberger et al. 2013), and notable differences exist between bats and terrestrial mammals such as rodents (Luis et al. 2013). For example, the often high-population densities and the usually gregarious roosting behaviour of bats increase the likelihood of both intra- and interspecies transmission of viruses (Luis et al. 2013; Streicker et al. 2010). Large-scale movements of bats due to their ability for powered flight are also likely to facilitate viral transmission within and among species, including the exchange of novel viruses and virus variants across biomes or even continents (Calisher et al. 2006; Epstein et al. 2009; Peel et al. 2013). The extreme relative longevity of bats compared to other mammals of similar size (Wilkinson and South 2002) and the potential for persistent and/or subclinical viral infections could further increase transmission potential (Calisher et al. 2006). Reduction of body temperature associated with hibernation of temperate zone bats lowers both viral activity and the metabolism of hosts, leading to increased incubation periods and therefore reduced likelihood of epizootic fadeout (of rabies , for example; George et al. 2011). Bats are ancient mammals in evolutionary terms, and virus utilisation of highly conserved cellular receptors could facilitate transmission to other mammals (Calisher et al. 2006), for example, as has been suggested for henipaviruses (Negrete et al. 2005). Lastly, it was recently speculated that, similar to the febrile response of other mammals, the relatively high body temperature (about 38–41 °C) and metabolism of bats during flight may select for viruses tolerant to such conditions, meaning the normal febrile defence mechanism of other mammals is ineffective (“Flight as fever hypothesis ”, O’Shea et al. 2014), making bat-borne viruses potentially more virulent and lethal for other, non-flying mammals.
3 Zoonotic Viruses of Bats and Their SpillOver
3.1 Rhabdoviruses
Rabies virus (RABV) is the longest and best-known member of the genus Lyssavirus (family Rhabdoviridae ) and still one of the most significant zoonoses known from bats (recent reviews include: Banyard et al. 2011; Banyard et al. 2014 and Kuzmin 2014). The genus is rapidly expanding, with 14 of the currently recognised species (plus another known from genetic material only), and all but two (Mokola and Ikoma virus es) having been isolated from bats (Table 10.1). Lyssaviruses spill over directly from bats to domestic animals , other wildlife and humans , or indirectly to humans via these other species. All lyssaviruses are potentially neurotropic , meaning that the virus infects nerve cells and replicates in the brain, resulting in clinical signs consistent with classical rabies (Schnell et al. 2009). Although isolated from a variety of tissues and body fluids in the late stages of infection , the predominant route of transmission is via saliva (mostly via biting; Kuzmin 2014).
Lyssaviruses can be divided into two distinct “phylogroups” (Badrane et al. 2001, Table 10.1), reflecting biological and genetic differences, and they are distributed globally in bats. Classical rabies virus occurs in bats across North , Central and South America (Messenger et al. 2003; Banyard et al. 2011) and was first associated with vampire bats following an outbreak in cattle in South America in 1911 (Carini 1911). It is reported most frequently in the common vampire bat ( Desmodus rotundus ; Kuzmin et al. 2011a), which has a wide distribution across Mexico, Central America, and South America. Bites from this species appear to be responsible for the majority of human and domestic animal rabies infections of bat origin in South and Central America, with increased prey availability via expansion of livestock into new areas across the region hypothesised to be contributing to increasing incidences (Schneider et al. 2009; Ruiz and Chávez 2010). In Canada and the USA , 51 cases of human rabies transmitted by non-haematophagous bats were recognised or inferred between 1951 and 2006 (mostly silver-haired bats ( Lasionycteris noctivagans ), eastern pipistrelle bat s ( Perimyotis subflavus ) and Brazilian/Mexican free-tailed bat s ( Tadarida brasiliensis )) (Constantine and Blehert 2009; Banyard et al. 2011). However, across the Americas, only 15 % of human rabies cases between 1993 and 2002 were reported as resulting from encounters with bats (Belotto et al. 2005).
Reported antibody prevalences against RABV in D. rotundus include 3–28 % in Peru (Streicker et al. 2012) and 12 % in Brazil (Almeida et al. 2011). Depending on the year, location and species, prevalence in other bats varies from relatively low 2 % in T. brasiliensis in New Mexico (Steece and Altenbach 1989) and 2.5 % in the little brown bats ( Myotis lucifugus ) in New York (Trimarchi and Debbie 1977), to 58 % in Seba’s short-tailed bat ( Carollia perspicillata ) in Peru (Salmón-Mulanovich et al. 2009) and 67 % in T. brasiliensis in Texas (Baer and Smith 1991). As with other lyssaviruses discussed below, the potential for high antibody prevalences in bat populations and infrequent reports of mortality suggest that many individuals exposed to the virus survive, contrary to the overwhelmingly lethal nature of lyssavirus infections in other mammalian species (reviewed in Banyard et al. 2011). The mechanisms for this remain unclear.
Seven bat lyssaviruses have been isolated in Eurasia (Table 10.1). European bat lyssavirus type 1 and type 2 (EBLV-1 and EBLV-2; Bourhy et al. 1992) are the most widely recognised and studied. Five fatal cases of human infections with EBLV have so far been reported, three from EBLV-1 (Roine et al. 1988; Selimov et al. 1989; Botvinkin et al. 2005) and two from EBLV-2 (Lumio et al. 1986; Fooks et al. 2003; Nathwani et al. 2003). Spillover of EBLV-1 into other mammals has also been observed, but rarely, with examples including zoo bats (Rønsholt et al. 1998), sheep (Tjørnehøj et al. 2006), domestic cats (Dacheux et al. 2009) and a stone marten (Müller et al. 2004). While EBLV-1 and EBLV-2 have been detected in a range of bat species (reviewed in Schatz et al. 2013), they are most frequently associated with serotine bats ( Eptesicus serotinus ) and Daubenton’s bat ( Myotis daubentonii ), respectively. The dynamics of EBLV infections in their natural hosts is poorly understood, but banding and recapture data and the frequent capture of apparently healthy bats with antibodies against EBLV suggest that many bats survive infection (Serra-Cobo et al. 2002; Amengual et al. 2007; Schatz et al. 2013). In cases where bats develop clinical symptoms of EBLV infection, the affected individuals are often unable to fly, are generally weak and show abnormal behaviour, including attempts to bite (Banyard et al. 2011). Experimental studies suggest that variable development of clinical signs may be related to inoculation route and dose (reviewed in Banyard et al. 2011).
Comparatively, little is known about the remaining Eurasian bat lyssaviruses, which have each been isolated from bats only once: West Caucasian bat virus (WCBV, Botvinkin et al. 2003), Bokeloh bat lyssavirus (BBLV, Freuling et al. 2011), Aravan virus (ARAV, Kuzmin et al. 1991), Irkut virus (IRKV, Botvinkin et al. 2003) and Khujand virus (KHUV, Kuzmin et al. 2001), or is only known from partial genetic sequence data (Lleida virus , Ceballos et al. 2013, Table 10.1). Of these, only IRKV has been detected in other mammals (a human who developed rabies after a bat bite, Leonova et al. 2009). WCBV appears to have a large geographical range. It was isolated from Miniopterus schreibersii in Russia , but cross-reactive antibodies have also been detected in Miniopterus bats in Kenya (Kuzmin et al. 2008a). The relatively wide distribution and migratory behaviour of Miniopterus spp. may facilitate cross-continental transmission of this virus. Alternatively, given the close relationship between WCBV and Ikoma virus (IKOV), which was recently isolated in neighbouring Tanzania, the serological findings from Kenya could in fact indicate exposure to IKOV or another related lyssavirus rather than WCBV (Marston et al. 2012; Horton et al. 2014). Similarly, serological surveys have detected antibodies against ARAV virus and KHUV virus in Indian flying foxes ( Pteropus giganteus ) from Bangladesh (Kuzmin et al. 2006), and ARAV, KHUV, IRKV or Australian bat lyssavirus in Lyle’s flying fox es (P. lylei ) and dawn bats ( Eonycteris spelaea ) from Thailand (Lumlertdacha et al. 2005). Yet, given the limited lyssavirus surveillance in bats performed to date in this region and that individuals in these studies tested positive to multiple viruses, these results likely represent cross-reactivity of serological assays to unknown lyssaviruses.
Africa also hosts significant lyssavirus diversity, with five species identified, though only three of these isolated from bats to date (Table 10.1). Duvenhage virus (DUVV, Meredith et al. 1971) is the only phylogroup I lyssavirus in Africa and is more closely related to RABV, ABLV and the majority of the European species than other known African lyssaviruses. Since it was first isolated from a human in 1970, two more fatal human infections of DUVV have been reported, one in South Africa in 2006 (Paweska et al. 2006) and one from the Netherlands in 2007 after obtaining the infection in Kenya (van Thiel et al. 2008). DUVV has been isolated from bats twice, once from a presumed M. schreibersii bat in South Africa and once from an Egyptian slit-faced bat ( Nycteris thebaica ) in Zimbabwe (Schneider et al. 1985; Foggin 1988; Paweska et al. 2006). No further information is so far available on this apparently rare African lyssavirus.
In contrast, Lagos bat virus (LBV ) is the most widely detected lyssavirus in Africa (Banyard et al. 2011). In 1956, this virus was first isolated from a straw-coloured fruit bat ( Eidolon helvum ; Boulger and Porterfield 1958). Since then, the virus has been isolated and neutralising antibodies detected in a variety of fruit bat species, one insectivorous bat species, domestic cats , domestic dogs and a water mongoose , but not in humans (reviewed in Banyard et al. 2011). E. helvum and Rousettus aegyptiacus are likely primary reservoir hosts for LBV, with seroprevalences ranging from 6 to 80 % and 29 to 46 %, respectively, depending on the region (Hayman et al. 2008, 2012; Kuzmin et al. 2008b; Dzikwi et al. 2010; Peel et al. 2013). LBV has been isolated from healthy, rabid and dead bats (reviewed in Banyard et al. 2011), but longitudinal studies in Ghana (Hayman et al. 2012) and surveys across continental Africa (Peel et al. 2010, 2013) suggest widespread exposure, no difference in survival between seropositive and seronegative E. helvum, and viral persistence in very small, isolated island populations. Early infection experiments with LBV suggested that LBV and other phylogroup II viruses were less pathogenic than other lyssaviruses (Boulger and Porterfield 1958; Badrane et al. 2001). However, recent experimental infections indicated the potential for comparable mortality between LBV and RABV and indicated that significant differences might instead exist between different LBV isolates (Kuzmin et al. 2010; Markotter et al. 2009).
Of the other Africa n lyssaviruses, only Shimoni bat virus (SHIBV) has been detected in bats (Commerson’s leaf-nosed bat ( Hipposideros commersoni ) in Kenya ; Kuzmin et al. 2010) and only Mokola virus (MOKV) has been detected in humans (on two occasions in Nigeria , Familusi and Moore 1972; Familusi et al. 1972). MOKV has also been isolated from cats and small wild mammals, however, the natural reservoir host is unknown (Nel 2001). Ikoma virus was isolated from a rabid African civet ( Civettictis civetta ), but it is believed that the civet was a spillover host and the true reservoir host is yet to be identified (Horton et al. 2014).
The only lyssavirus detected in Australia to date— Australian bat lyssavirus (ABLV)—has two known lineages, one circulating in flying foxes and one in an insectivorous bat (Fraser et al. 1996; Gould et al. 2002; Warrilow 2005). In 1996, shortly after ABLV was first isolated from a black flying fox (P. alecto ) that was unable to fly (Fraser et al. 1996), a 39-year-old woman died of clinical rabies after being bitten by a yellow-bellied sheath-tail bat ( Saccolaimus flaviventris ; Gould et al. 2002). Two subsequent human cases have been identified, a woman who died in 1998, 27 months after being bitten by a flying fox (Hanna et al. 2000), and a child who died in 2014 after being scratched by a flying fox (Francis et al. 2014). Experimental infection of grey-headed flying foxes (P. poliocephalus ) with ABLV resulted in clinical signs of weakness, trembling and limb paralysis in three out of ten individuals (McColl et al. 2002). As with other bat lyssaviruses, a small proportion of ABLV-positive bats succumb to encephalitis -like symptoms (Hooper et al. 1997), yet serological tests show a high prevalence of antibodies in populations of surviving bats (McColl et al. 2000).
3.2 Paramyxoviruses
The most notable viruses from the Paramyxoviridae family in bats are those of the genus Henipavirus , which are the subject of many reviews (e.g. Halpin and Rota 2015; Smith and Wang 2013, Luby and Gurley 2012; Clayton et al. 2013; Middleton and Weingartl 2012; Field and Kung 2011). The first recognised henipavirus, Hendra virus (HeV ), was first detected during an outbreak of infectious respiratory disease in horses and then humans in Hendra, Australia , in 1994 (Murray et al. 1995). Ultimately, 13 of 20 infected horses died or were euthanised, and of two humans working closely with horses who became infected, one died from acute pneumonia (Murray et al. 1995; Plowright et al. 2015). This spillover was preceded a month earlier by another involving two horses and one human over 800 km away in Mackay, but which went unrecognised until 1995 (Rogers et al. 1996; O’Sullivan et al. 1997). An initial serological survey of 46 wildlife species (excluding bats) failed to identify a reservoir host ; however, serological evidence of HeV infection was later identified in all four species of flying foxes native to Australia (Young et al. 1996). Virus isolation (Halpin et al. 2000) and experimental studies (Halpin et al. 2011) have confirmed pteropodid bats as reservoir hosts for henipaviruses (with a lack of clinical signs ), with evidence that black (P. alecto ) and spectacled flying fox es (P. conspicillatus ) are the main reservoir species for HeV (Smith et al. 2014; Goldspink et al. 2015).
Because HeV is frequently detected in the urine of wild flying foxes (Smith et al. 2014), the predominant transmission route to horses is likely via material recently contaminated with bat urine (e.g. pastures) or via direct transmission (Martin et al. 2015). Recognised spillover events from bats to horses occurred sporadically from 1994 to 2004 and annually since 2006, with five spillover events resulting in ongoing transmission to humans in close contact with horses (a total of seven human cases and four deaths; Field et al. 2010). Spillover events are spatiotemporally clustered, occurring year-round in the northern tropics, but seasonally clustered in winter with a peak in July in subtropical regions (Plowright et al. 2015). The relative importance of various hypothesised drivers of HeV dynamics in bats and subsequent spillover to horses is still unclear (Plowright et al. 2015).
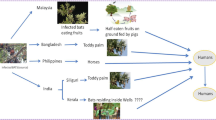
Nipah virus (NiV), the second henipavirus to be recognised, was first isolated in 1999 from pigs and encephalitic pig workers in Malaysia (Center of Disease Control and Prevention 1999). NiV spillover has not been observed since this time in Malaysia; however, annual seasonal outbreak s with high case fatality (average 73 %) have occurred in people in Bangladesh since 2001 (Hsu et al. 2004; Luby et al. 2009; Luby and Gurley 2012), with occasional spillover also occurring in neighbouring India (Chadha et al. 2006; Harit et al. 2006). Due to the close relatedness of HeV and NiV, fruit bats were targeted, and serological evidence quickly identified them to be the natural reservoir of NiV (Enserink 2000; Yob et al. 2001). This was subsequently supported by isolation of NiV from the urine of P. hypomelanus (Chua et al. 2002a), P. vampyrus (Rahman et al. 2010) and P. lylei (Reynes et al. 2005), and seroconversion in the absence of clinical signs following experimental infections in P. vampyrus (Halpin et al. 2011). Antibodies against NiV and NiV-related viruses have now been detected in a variety of bat species (including non-pteropid bats) across a wide geographical area (summarised in Breed et al. 2013). NiV transmission to humans appears to occur via a wider variety of routes compared with HeV. Infection of domestic animal intermediate hosts (via consumption of saliva - or urine - contaminated partially eaten fruits or raw date palm sap ) has been implicated as a source of human infection s in both Malaysia and Bangladesh (Chua et al. 2002b; Chowdhury et al. 2014). In Malaysia, human infection s resulted from direct contacts with infected pigs (Chua et al. 1999; Paton et al. 1999; Parashar et al. 2000), whereas in Bangladesh, transmission to humans regularly occurs via consumption of contaminated date palm sap (Luby et al. 2006; Rahman et al. 2012) or directly from human to human (e.g. via nursing sick individuals or preparation for burial; Hughes et al. 2009). The risk of direct human infection with NiV from bats is considered to be lower than horizontal transmission once the virus enters the human population (Gurley et al. 2007; Luby et al. 2009; Chong et al. 2003).
A third henipavirus, Cedar Virus (Marsh et al. 2012), has been isolated from urine collected under a mixed P. alecto /P. scapulatus roost in Australia . In contrast to HeV and NiV, however, it appears to be of low pathogenicity and failed to induce clinical signs in experimentally infected laboratory animal species (Marsh et al. 2012). Serological evidence from South-East Asia and Australasia (Breed et al. 2013) and the wide diversity of paramyxovirus sequences detected in Australia (Vidgen et al. 2015) suggest more henipaviruses are yet to be found. Additionally, although henipaviruses were long thought to be restricted to Asia and Australia, antibodies cross-reactive to HeV and NiV were detected in Madagascar in 2007, suggesting a potentially wider geographical distribution of henipa-related paramyxoviruses (Iehlé et al. 2007). This was supported by serological findings and molecular detection of henipa- or henipa-like viruses in mainland Africa and its offshore islands (Hayman et al. 2008, 2012; Peel et al. 2010, 2013; Drexler et al. 2012). Indeed, a recent serological study indicates that these viruses are also occasionally transmitted to humans in Africa (Pernet et al. 2014), though no African henipavirus has been isolated to date.
Viruses from the paramyxovirus genus Rubulavirus (a genus which includes the human mumps virus ) have also been frequently detected in bats (Barr et al. 2015). Menangle virus was isolated from pigs following the birth of unusually high numbers of stillborn and deformed piglets in Australia (Philbey et al. 1998). Two piggery personnel had neutralising antibodies against Menangle virus after having recovered from an unexplained febrile illness (Philbey et al. 1998). Flying fox colonies roosting in close proximity to the piggeries were a suspected source of infection for pigs, with subsequent transmission to humans (Philbey et al. 1998). This was supported by serological evidence from P. poliocephalus , P. alecto and P. conspicillatus , and recent virus isolation from P. alecto (Barr et al. 2012). Other isolated bat rubulaviruses with unknown or limited understanding of their zoonotic potential include Tioman virus from Malaysia (Chua et al. 2001), Tuhokovirus 1, 2 and 3 from China (Lau et al. 2010), Achimota virus 1 and 2 from Ghana (Baker et al. 2013c) and Hervey, Grove, Teviot and Yeppoon paramyxoviruses from Australia (Barr et al. 2015). Neutralising antibodies to Tioman virus and Achimota viruses have been detected in humans , suggesting previous exposure and infection with the virus (Yaiw et al. 2007; Baker et al. 2013c). Pigs experimentally infected with Tioman virus produced neutralising antibodies and excreted virus in saliva , but were either asymptomatic or developed only a fever (Yaiw et al. 2008). Undetected infection in pigs could therefore facilitate transmission to humans.
Finally, viral fragments related to rubulaviruses and the proposed genus Jeilongvirus have also been detected outside the range of fruit bats, in European insectivorous bat species (Kurth et al. 2012). However, nothing is yet known about the relevance of these viruses as potentially zoonotic threats to humans .
3.3 Coronaviruses
Bat coronaviruses were first identified from species of the genus Miniopterus (Poon et al. 2005), however, with unknown zoonotic potential. The most prominent coronavirus, the one causing severe acute respiratory syndrome ( SARS ), was followed by a pandemic spread in humans after the first outbreak in China in 2002 (Rota et al. 2003). Soon after the outbreak, the virus was detected in masked palm civet ( Paguma larvata ) and raccoon dog s (Nyctereutes procyonoides) in a market in Guangdong Province, where SARS was first reported (Guan et al. 2003). A survey of common wildlife species in the area identified bats to be the natural reservoir of SARS coronavirus , with viruses from bats showing greater genetic diversity than the ones isolated from other species, including humans (Li et al. 2005). Bats can regularly be found in markets in China, which makes direct transmission of the virus from bats to humans likely (Li et al. 2005). The followed pandemic spread with 8096 confirmed cases of which 774 were fatal can be accounted to rapid interindividual transmission of the virus once it entered the human population (World Health Organization 2003).
Outside Asia , SARS-like coronaviruses have been detected in the lesser horseshoe bat ( Rhinolophus hipposideros) from Europe (Rihtarič et al. 2010), in Chaerephon sp. from Kenya (Tong et al. 2009) and in Hipposideros commersoni from Nigeria (Quan et al. 2010). Antibodies against SARS coronavirus are present in various Africa n bat species (Müller et al. 2007). As with many newly detected viruses, their potential threat as a zoonotic disease is yet unclear.
Since the outbreak of SARS in Asia has been traced to bats as natural hosts of the virus, the same was suspected to be the case for Middle East respiratory syndrome ( MERS ), an infection that has been occasionally spreading among humans of the Arabia n peninsula since 2012 (Zaki et al. 2012). Most human infections have been traced down to close contacts with dromedary camels ( Camelus dromedarius ), which carry a virus with a similar genome organisation as human MERS (Hemida et al. 2014). There is at least one report of direct transmission of the virus from camels to humans via contact with infected animals (Memish et al. 2014). However, a small fragment of a coronavirus PCRed from an Egyptian tomb bat ( Taphozous perforatus ) showed 100 % nucleotide identity to virus from the human index case-patient of MERS , suggesting that this species may be one of the putative natural reservoir s of the virus (Memish et al. 2013). Bat-derived MERS virus has been shown to be able to use human receptors and thus could potentially infect human cells (Yang et al. 2014). However, given the generally low prevalence of MERS virus in bat populations, a direct spillover from bats to humans is unlikely, and transmission probably happens mainly via camels as intermediate hosts (Memish et al. 2013). In fact, no other bat has yet been found to carry MERS virus since the one reported by Memish and colleagues in 2013.
The intensified search for viruses in bats worldwide has led to the detection of coronaviruses other than SARS and MERS, whose potential to be or become zoonotic has yet to be investigated (Woo et al. 2006; Tang et al. 2006; Dominguez et al. 2007; Carrington et al. 2008; Brandão et al. 2008; Misra et al. 2009; Pfefferle et al. 2009; Donaldson et al. 2010; Watanabe et al. 2010; Drexler et al. 2010; Falcón et al. 2011; Annan et al. 2013; Ge et al. 2013; Anthony et al. 2013; Ithete et al. 2013). No clinical symptoms associated with infections with SARS-like and other coronaviruses have yet been described for bats.
3.4 Filoviruses
Ebola virus is the most prominent filovirus, causing severe haemorrhagic fever in humans with high mortality and fast spreading among African populations. The recent outbreak in 2013 in west Africa has resulted in the most severe epidemy of Ebola so far, with more than 11,000 lethal cases (as by September 2015; according to World Health Organization;http://apps.who.int/ebola/ebola-situation-reports).
All Ebola outbreaks recorded until 2004 in Gabon and the Republic of the Congo have been linked to handling of gorilla , chimpanzee or duiker carcasses, species that can carry the Ebola virus (Leroy et al. 2004; Pigott et al. 2014). It has thus became apparent that spillover from animals to humans occurs through hunting, butchering and consumption of bushmeat (Gonzalez et al. 2005; Li and Chen 2014; Chap. 12), followed by fast human-to-human transmission (World Health Organization 2014). An outbreak of Ebola in Congo in 2007 that resulted in 260 infected humans of whom 186 died has been traced to a potential direct transmission from a dead fruit bat that the first human victim bought from hunters to eat (Leroy et al. 2009). Antibodies against Ebola virus have since been detected in a total of 14 bat species, with seroprevalences of up to 44 % depending on species and location (Olival and Hayman 2014). Experimental infection of several bat species with Ebola led to high replication of the virus, but to no apparent signs of illness, suggesting that Ebola infections are subclinical in these species (Swanepoel et al. 1996). One Eidolon helvum has survived for at least 13 months after being tested seropositive for Ebola virus and Lagos bat virus , indicating long-term survival of an individual bat following exposure to these viruses (Hayman et al. 2010). The recent outbreak of Ebola in Guinea and neighbouring countries in 2013—countries that are at significant distance to the previous outbreaks in central Africa —has caused speculations about a possible transmission of the virus by migrating fruit bats (Bausch and Schwarz 2014; Vogel 2014). However, as the strain of the west African Ebola virus is a genetic outlier within the known Ebola viruses, it has been argued that the west African variant may have emerged from local wildlife populations rather than from migrating individuals (Gatherer 2014). Furthermore, although speculated (Saéz et al. 2015), it is yet not clear whether the spillover of Ebola virus in west Africa originated from bats.
Marburg virus is the only filovirus that has so far been directly isolated from bats (Towner et al. 2009; Amman et al. 2012; Pourrut et al. 2005). The first outbreak of the virus was caused by a spillover from laboratory monkeys to humans in Marburg, Germany, in 1967 (Jacob and Solcher 1968). In 2007, mine workers in a cave in Uganda were diagnosed with Marburg haemorrhagic fever that potentially resulted from a spillover of the virus from a colony of Rousettus aegyptiacus , where 5.1 % of tested individuals carried the virus (Towner et al. 2009). The high divergence of the genome sequence of Marburg in this population suggests a long-term association of the virus with the host, leading to the assumption that bats are the natural reservoir (Towner et al. 2009). However, given that no other bat species has yet tested positive for the virus (Towner et al. 2007), and seroprevalence being generally low in R. aegyptiacus (Pourrut et al. 2009), spillovers from bats to humans may be rare events.
The Reston Ebolavirus has first been detected in 1989 in crab-eating macaques ( Macaca fascicularis ) imported from the Philippines to be used for animal testing in laboratories in Reston, USA (Jahrling et al. 1990). During a second outbreak in 1990, animal handlers developed antibodies but did not get sick (Center for Disease Control and Prevention 1990). In 2008, Reston Ebolavirus was isolated from pigs in the Philippines (Marsh et al. 2011), and soon after, some sampled R. amplexicaudatus had antibodies against the virus, while 16 other bat species tested negative against Reston Ebolavirus (Taniguchi et al. 2011). Screening for antibodies of the Ebola virus and Reston Ebolavirus in bats in Bangladesh has found seropositive R. leschenaultii , suggesting that these filoviruses or related strains are distributed at a much larger geographic range than previously assumed (Olival et al. 2013).
4 Main Conservation Issues Related to Bat Viruses
4.1 Direct Effect: Viruses Killing Bats
From all the viruses described above, only a few seem to affect bats. Although experimental infection with RABV leads to mortalities between 40 and 90 % depending on the bat species (Sétien et al. 1998; Jackson et al. 2008; Turmelle et al. 2010), there are no observed mass mortalities in natural populations (Pawan 1959). The only virus that may be largely lethal for bats is the Lloviu virus, which is closely related to Ebola and Marburg virus , but not yet of zoonotic relevance. It was detected during investigations of a massive die-off of Miniopterus schreibersii in a cave in Spain (Negredo et al. 2011). However, a causal connection between the detected virus and death of the bats has not yet been confirmed, and other bat species roosting in the same caves appeared to remain unaffected (Roué and Nemoz 2004).
The lack of reports of viruses that are detrimental for bat health should not imply that viruses in general are not of importance for the conservation of bat populations. Similar to white-nose syndrome causing mass mortalities in North American bats (Frick et al. 2010), newly emerging viruses may put local populations at threat. This may be especially the case if pathogens cross geographical borders and infect naïve bat populations. Pseudogymnoascus destructans —the causative fungus responsible for white-nose syndrome—likely originated from Europe , where it seemingly causes no bat fatalities, in contrast to North America (Puechmaille et al. 2010; Frick et al. 2010; Frick et al. 2016, Chap. 9).
4.2 Indirect Effects: Biased Public Perception
Generally, the public perception of bats as aesthetically less appealing mammals as well as folklore s that often associate bats with negative stigma makes bat-related conservation efforts time-consuming and demanding (Fenton 1997; Allen 2004; Knight 2008). The recent outbreak s of viral zoonotic diseases with the identification of bats as putative natural host s have further complicated bat conservation efforts (Li et al. 2005; Knight 2008). Following numerous and often lurid reports of fatal zoonotic diseases by the media , public perception of bats is mostly skewed by fear and lack of information (Kingston 2016, Chap. 18). Therefore, it is important to highlight the context of bat-associated infections in order to provide more evidence-based information about the emergence and transmission of bat-related zoonotic diseases , which may lead to a more balanced reputation of bats. Depending on educational, cultural, legal and medial background of the targeted audience, specific aspects need to be taken into account.
In Europe and North America , rabies is, so far, the only viral disease that is associated with bats. The fact that lyssaviruses are occasionally found in temperate zone bats sometimes finds its way to the media, not always in favour of bats. Biased newspaper articles or press campaigns may result in the public misconception that bats are aggressive animals or that their mere presence can lead to human infections with these viruses. Although there are anecdotal reports of unprovoked attacks of bats on humans and dogs (Baer and Smith 1991), bats, as is the case of most mammals, usually only bite when handled or provoked. Furthermore, once bitten or scratched by a bat, immediate post-exposure vaccination can prevent a person from contracting rabies (see Sect. 10.5.2). In the case of the 37-year-old woman who died from a bat lyssavirus infection in Kenya , staff members of the health facility which the woman visited after being scratched by a bat were unaware of the possibility of rabies transmission (van Thiel et al. 2009). Likewise, two persons in Europe who worked regularly with bats and died from rabies after being bitten and scratched by bats received neither pre- nor post-exposure treatment (Roine et al. 1988; Nathwani et al. 2003). These two cases triggered a Europe-wide serological screening effort involving more than 11,000 bats, with seroprevalences varying depending on the species and location (Racey et al. 2012). EBLV-1 was most commonly detected in the serotine bat ( Eptesicus serotinus ), while EBLV-2 was very uncommon in all bat species. As a result, the public has been persuaded not to handle bats or to do so only with gloves and, in the case of bat workers, to receive pre- and/or post-exposure immunisation. Two fatal cases in which persons contracted rabies in Australia (Samaratunga et al. 1998; Hanna et al. 2000) triggered a similar campaign on this continent (Speare et al. 1997, but see Francis et al. 2014). Efficient education of medical professionals worldwide seems to be pivotal for implementing the correct treatment after scratches or bites from bats. In addition, vaccination should be mandatory for those who are frequently exposed to bats (Rupprecht and Gibbons 2004). Studies on animal models have shown that rabies vaccine also provides protection against other, although not all, lyssaviruses’ variants (Brookes et al. 2005; Hanlon et al. 2005). However, there is no known case of a person developing bat-associated rabies despite having been vaccinated, neither pre- nor post-exposure. Thus, getting infected by some sort of bat-related virus is unlikely in Europe and North America and decreases virtually to zero if people who experienced bat bites and scratches are treated appropriately.
There is no case known for paramyxoviruses having spilled over to humans by direct contact with bats. An extensive serological survey among people frequently handling bats in Australia revealed no antibodies against Hendra virus (Arklay et al. 1996). The virus apparently needs horses as amplifier host s, from where the virus can further be transmitted to persons in close contact with infected individuals. Nevertheless, the outbreak of Hendra increased the unpopularity of flying foxes in Australia, making conservation of the four native species challenging (Thiriet 2011). Unlike Hendra, Nipah virus has likely been acquired by humans via consumption of contaminated date palm sap (Luby et al. 2006; Rahman et al. 2012), followed by person-to-person transmission (Gurley et al. 2007). Although diseases associated with Hendra virus and Nipah virus have high mortality rates, the risk of infection for humans seems to be low (Chong et al. 2003), and countermeasures may be taken in order to prevent future spillover events (see Sect. 10.5.2). MERS, just as Hendra virus, apparently needs livestock as an amplifier host . In contrast to dromedaries (Hemida et al. 2014), seroprevalence of MERS seems to be low in bats (Memish et al. 2013), making direct transmission from bats to humans unlikely. As long as details on MERS infections in dromedaries and how to mitigate them are missing, it is hard to give recommendations to people who might be at risk.
In contrast to MERS, the spillover of SARS into the human populations most likely happened via the wildlife market , either directly from a bat, or from other wildlife species. Likewise, the hunting, butchering and consumption of chimpanzees , gorillas and bats seem to have been sources of Ebola spillover s from wildlife to humans. The education of local communities needs to carefully balance information about the potential risk of acquiring infectious diseases by consuming bushmeat , without implying that bats need to be eradicated in order to prevent spillovers. The recent outbreak of Ebola resulting in several thousand human victims, and with bats frequently being reported as the likely source of origin, has undoubtly led to severe loss of reputation of bats on this continent, which makes the conservation of threatened populations and species even more challenging, not only in Africa , but also worldwide.
4.3 Indirect Effect—Culling
The direct persecution of bats often seems to be the most effective way to deal with bat-borne diseases to members of the public . Killing of bats has long been acceptable, even if they are protected (Chap. 14). Even though culling may be officially banned and thus not supported by authorities or governmental programs, large-scale killing of bats or the destruction of roost trees may still be commonly practiced in areas where zoonotic diseases are spreading.
In Australia , for example, flying foxes are frequently harassed and killed, both legally (under permits issued by state wildlife management agencies) and illegally. This happened most prominently during periods when Hendra virus emerged in Australian flying fox populations (Roberts et al. 2012). Half of the flying fox species native to Australia have declined about 30 % in population size during the last decade, and killing of bats usually does not lead to legal measures (Booth 2005). Furthermore, large-scale culling leads to a change of movement behaviour of bats, with new, susceptible individuals being recruited from nearby colonies (Field 2009). Instead of reducing the viral prevalence , this may therefore lead to the exact opposite (see below).
In the attempt to reduce rabies incidences, vampire bats are regularly culled in many parts of Latin America (Streicker et al. 2012). In Brazil , for example, governmental programs are in action that involve targeted campaigns against vampire bats. During these measures, vampire bats are captured and poisoned or coated with anticoagulant and released, so that allogrooming kills their conspecifics (Medellin 2003). Furthermore, bat roosts are destroyed using fire and explosives (Mayen 2003), which also leads to dramatic declines of non-target bats (Furey and Racey 2016, Chap. 15). Besides the questionable methods involved, instead of reducing viral abundance in the population, culling of wildlife can lead to an increase in viral spreading. New hosts are recruited and the dispersal probability of infected individuals increases, which results in transmission of the disease to naïve hosts (Donnelly et al. 2005; Choisy and Rohani 2006; Streicker et al. 2012). This was the case for vampire bats in Peru , where culling failed to reduce seroprevalence of rabies in bat populations, but rather had the opposite effect (Streicker et al. 2012). Therefore, persecution of bats as potential carriers of zoonotic diseases has been denounced as useless and even counterproductive by both conservationists and experts on disease transmission (Hutson and Mickleburgh 2001; Knight 2008).
4.4 Indirect Effect—Killing of Bats for Virus Surveys
In the scope of recently emerging zoonotic diseases, the search for new bat-borne viruses has become a well-funded field in the scientific community. While research is important to advance our understanding about the emergence of diseases and to possibly prevent further spillover events, the methods involved in these surveys are sometimes questionable from the perspective of bat conservation (Racey 2015). Some of the investigated bat species are listed as near threatened or vulnerable by the International Union for the Conservation of Nature (IUCN), with decreasing population sizes even in many species of least concern. While most surveillance studies that involve species of conservation concern use non-lethal methods such as antibody screening in blood (Hayman et al. 2008; Young et al. 1996; Lumlertdacha et al. 2005; Wacharapluesadee et al. 2005; Reynes et al. 2005), others have involved the killing of a considerable number of bats of various conservation status (e.g. in Yob et al. 2001; Kuzmin et al. 2008b, 2010, 2011b; Dzikwi et al. 2010 and Sasaki et al. 2012). In order to limit such detrimental surveys, the Food and Agriculture Organization of the United Nations (2011) has published a guideline for investigating the role of bats in emerging zoonotic diseases, including non-invasive protocols, which not only reduce the impact on bat populations, but also minimise the transmission risk of viral diseases. Such protocols have now been widely adopted, as for example by Ecohealth Alliance and other international research groups and networks.
5 Counter Measures in Favour of Bat Conservation
5.1 Preventing the Emergence of New Viral Diseases
In general, preventing the emergence of infectious diseases in wildlife populations is extremely challenging and usually underfunded, with only few practical suggestions being discussed (Daszak et al. 2000). For example, it is important that translocation s of animals across geographical borders need to follow strict guidelines in order to prevent the introduction of exotic pathogens in novel areas (e.g. Woodroofe 1999). Furthermore, an integration of knowledge about disease dynamics, as well as ecological and immunological aspects of the host, may contribute to a better understanding of emerging infectious diseases in wildlife species such as bats (Daszak et al. 2000).
5.2 Educational Efforts
As many bat-borne viral diseases have high lethality rates for humans, preventing spillover events are of central importance. In particular, spillover by direct contact to bats, such as via bites or bat consumption, may bear severe risks to humans that could be minimised by educational programs (Kingston 2016, Chap. 18). Reducing the risk of outbreaks of zoonotic viruses may also lead to more positive attitudes towards bats, which may further be increased by highlighting their ecological importance as pollinators, seed dispersers and pest control for agriculture (Ghanem and Voigt 2012). Moreover, conservation measures that promote the preservation of bat habitats serve a dual role as they can decrease the contact zone between bats and humans, thus reducing the risk of spillover .
As aforementioned vaccination against rabies and other lyssaviruses should be mandatory for persons working with bats and recommended for other people at risk. A significant problem is that both pre- and post-exposure treatments are expensive and thus may not be readily available in developing countries, such as in Central and South America. Here, building houses in a bat-proof manner in order to avoid vampire bites during sleep and decreasing the risk of direct contact with other bats has so far been the best solution (Greenhall 1964; Voigt et al. 2016, Chap. 14).
A different issue is the transmission of Nipah viruses via consuming raw date palm sap contaminated by urine , faeces or saliva of bats (Luby et al. 2006; Rahman et al. 2012). Here, cooking the sap at temperatures above the level that viruses tolerate is an effective measure to prevent spillover (Hughes et al. 2009). Additionally, preventing bats from accessing date palms and thus contaminating the sap has been proved to be both efficient and relatively cheap (Nahar et al. 2010, 2013). The traditional “bamboo skirt” method for example uses inexpensive, recyclable bamboo to cover the part of the date palm where the sap is collected, preventing bats and other vertebrates from getting access. Furthermore, in contrast to bird nets, this measure is non-lethal to the bats and therefore of high conservation value to local populations. However, such protective measures are reported to be rarely used in Bangladesh (Nahar et al. 2010, 2013). This could potentially be changed by encouraging local farmers to use this method, emphasising its inexpensiveness and efficiency while highlighting the reduced risk of acquiring Nipah virus disease.
One of the key issues both for conservation and public health is the direct transmission of SARS and Ebola via wildlife markets. In South-East Asia , flying foxes are hunted regularly for the purpose of food (Mickleburgh et al. 2002; Mildenstein et al. 2016, Chap. 12), sometimes even authorised by the local Wildlife Department such as in Malaysia (Breed et al. 2006). Likewise, fruit bats are consumed regularly throughout Africa (Mickleburgh et al. 2009; Mildenstein et al. 2016, Chap. 12). Since bats are suggested as potential reservoir for the recent outbreak of Ebola, Guinea banned bats for sale from markets (Gatherer 2014). Educational efforts to reduce the threat both to public health by zoonotic diseases and to the conservation of local bat populations are challenging, as they are usually impeded by the lack of understanding of entrenched cultural behaviours and social components (Pooley et al. 2015; Kingston 2016, Chap. 18). In Ghana, for example, where the consumption of bats is part of the local culture and traditions, a survey revealed that knowledge about the ecological and economical value of bats would not make people refrain from killing and eating bats (Kamins et al. 2014). Usually, the direct economic benefit from selling hunted bats is more valuable to an individual person than the indirect, not always obvious economic value of bats, for example, for agriculture. However, about half of the hunters stated they would stop hunting bats if they could make them sick (Kamins et al. 2014). This highlights the potential effectiveness of public education, but careful consideration is needed to avoid demonising bats in the process (Pooley et al. 2015). The recent Ebola epidemic in western Africa for example has led to an increase in the persecution of bats, with roosts being destroyed and colonies being killed by communities (Fig. 10.1). Although preventing bats from being consumed may have higher priorities due to public health reasons, the culling of whole colonies as a likely result may be much more of a threat for the conservation of bats than the bushmeat trade (Pooley et al. 2015).

Intact trees with colonies of Eidolon helvum (left) in Yaoundé, Cameroon, as compared to former roosting trees that have been cut (right) after bats were suspected to be the source of the recent Ebola outbreak in western Africa (photograph credits: Simon Ghanem)
5.3 Environmental Conservation
Combining knowledge about the ecology of the host species as well as the disease dynamics of the virus may be crucial for establishing efficient disease prevention programs (e.g. Plowright et al. 2015). Here, it needs to be noted that the emergence of zoonotic diseases from bats also seems to be a consequence of anthropogenic alteration of natural environments (e.g. Daszak et al. 2001). For example, in Central and South America, the conversion of forested habitats into pastures shifted the dominant food source for vampire bats from native vertebrates to livestock . This has increased rabies transmission from vampire bats to livestock and domestic animals in many parts of Latin America (Schneider et al. 2009). Where bat habitats have been converted largely into agricultural farmland, the remaining bat populations are forced to concentrate in patches that provide them with resources they need. Flying foxes , for example, are highly sensitive to landscape modifications, as they require large forested areas for foraging. Where natural habitats are scarce, flying foxes may use fruiting or flowering trees in agricultural, suburban and urban areas, which increases the contact zone and spillover risk between bats and livestock or humans (Daszak et al. 2006; Plowright et al. 2015). Indeed, contact between bats and naïve hosts as a consequence of human landscape modification and encroachment likely sparked the transmission of Hendra viruses to horses (Epstein et al. 2006) and Nipah virus to pigs (Chua et al. 1999; Field et al. 2001).
5.4 Conservation of Bat Populations and Population Dynamics
Removing individuals or colonies from regional populations, either by unsustainable hunting or culling, can cause an increase in relative local resource availability, creating regional gradients along which bats from other populations may move, which may lead to an increase of virus movement (Field 2009). In Australia , for example, roosts that became empty after culling , disturbing or relocating colonies of flying foxes are usually reoccupied by immigrating individuals (Roberts et al. 2012).
Anthropogenic transformation of bat habitats in Australia has also been shown to lead to decreased migration in Pteropus bats, which can itself lead to a decline in population immunity (Plowright et al. 2011). This could give rise to more viral shedding after local viral reintroduction, a mechanism that may be facilitated by urban habituation of fruit bat and the resulting increased contact with human and domestic animal populations (Epstein et al. 2006; Plowright et al. 2011). In Australia, all recently emerged bat-associated viruses—Hendra , Menangle and Australian bat lyssavirus —are hypothesised to be associated with habitat loss due to deforestation and agricultural intensification (Jones et al. 2013). Therefore, protection of remaining natural habitats of bats along with farm management aiming at decreasing the contact zone between bats and livestock as well as education plans increasing awareness of environmental issues and safety may play a crucial role in the avoidance of future spillover s of bat-borne diseases to livestock and human populations, and promote further protection of local bat populations.
6 Conclusion
Bats harbour viruses that may become zoonotic. Circumstances facilitating spillover include direct contact with bats (bites, scratches, consumption of bats), contact with material contaminated by bat saliva , faeces or urine and amplification via intermediate hosts such as domestic animals or other wildlife species . Conservational actions are not only important to prevent spillovers, but also because emerging zoonotic viruses often lead to persecution of bats. In order to reduce the transmission risk of viruses from bats to human and livestock and to protect bat species at threat, educational efforts are needed. However, entrenched cultural and social components often act as barriers to efficient changes on how people think about and respond to bats. Whenever possible, educational efforts should be done in an informative, non-lurid way, presenting the facts rather than provoking additional fears to the already bad reputation of bats. Wherever possible, solutions should be found to enable the existence of bats in anthropogenic landscape, including the development of more affordable and readily available vaccinations (e.g. against rabies ), and the reduction of potential contact between bats and humans and livestock . This however also includes that the natural habitats of bats need to be better protected to provide bat populations with sufficient space and to prevent range expansion into urban and suburban areas, where contact with humans and livestock may increase the risk of spillover events. Bat-borne viruses should be considered during bat conservation efforts, and it should be equally noticed that appropriate conservation measures may even reduce the risk of viral spillover from bat populations into human populations.
References
Allen GM (2004) Bats: biology, behavior, and folklore. Dover Publications
Allen LC, Turmelle AS, Mendonça MT, Navara KJ, Kunz TH, McCracken GF (2009) Roosting ecology and variation in adaptive and innate immune system function in the Brazilian free-tailed bat (Tadarida brasiliensis). J Comp Physiol B 179(3):315–323
Almeida MFD, Martorelli LFA, Sodré MM et al (2011) Rabies diagnosis and serology in bats from the State of São Paulo, Brazil. Rev Soc Bras Med Trop 44(2):140–145
Amengual B, Bourhy H, López-Roig M, Serra-Cobo J (2007) Temporal dynamics of European bat Lyssavirus type 1 and survival of Myotis myotis bats in natural colonies. PLoS ONE 2(6):e566
Amman BR, Carroll SA, Reed ZD et al (2012) Seasonal pulses of Marburg virus circulation in juvenile Rousettus aegyptiacus bats coincide with periods of increased risk of human infection. PLoS Pathog 8:e1002877
Annan A, Baldwin HJ, Corman VM et al (2013) Human betacoronavirus 2c EMC/2012–related viruses in bats, Ghana and Europe. Emerg Infect Dis 19(3):456
Anthony S, Ojeda-Flores R, Rico-Chávez O et al (2013) Coronaviruses in bats from Mexico. J Gen Virol 94(Pt 5):1028–1038
Arklay A, Selvey L, Taylor R, Gerrard J (1996) Screening of bat carers for antibodies to equine morbillivirus. Commun Dis Intell 20:477
Badrane H, Bahloul C, Perrin P, Tordo N (2001) Evidence of two Lyssavirus phylogroups with distinct pathogenicity and immunogenicity. J Virol 75(7):3268–3276
Baer GM, Smith J (1991) Rabies in nonhematophagous bats. Nat Hist Rabies 2:105–120
Baker KS, Leggett RM, Bexfield NH et al (2013a) Metagenomic study of the viruses of African straw-coloured fruit bats: detection of a chiropteran poxvirus and isolation of a novel adenovirus. Virology 441(2):95–106
Baker ML, Schountz T, Wang LF (2013b) Antiviral immune responses of bats: a review. Zoonoses Public Health 60(1):104–116
Baker KS, Todd S, Marsh GA et al (2013c) Novel, potentially zoonotic paramyxoviruses from the African straw-colored fruit bat Eidolon helvum. J Virol 87(3):1348–1358
Banyard AC, Hayman D, Johnson N, McElhinney L, Fooks AR (2011) Bats and lyssaviruses. Adv Virus Res 79:239–289
Banyard AC, Evans JS, Luo TR, Fooks AR (2014) Lyssaviruses and bats: emergence and zoonotic threat. Viruses 6(8):2974–2990
Barr JA, Smith C, Marsh GA, Field H, Wang LF (2012) Evidence of bat origin for Menangle virus, a zoonotic paramyxovirus first isolated from diseased. J Gen Virol 93:2590–2594
Barr J, Smith C, Smith I et al (2015) Isolation of multiple novel paramyxoviruses from pteropid bat urine. J Gen Virol 96:24–29
Barrett JL (2004) Australian bat lyssavirus. Ph.D. thesis, University of Queensland, Brisbane
Bausch DG, Schwarz L (2014) Outbreak of Ebola virus disease in Guinea: where ecology meets economy. PLoS Neglected Trop Dis 8(7):e3056
Belotto A, Leanes L, Schneider M, Tamayo H, Correa E (2005) Overview of rabies in the Americas. Virus Res 111(1):5–12
Booth C (2005) Time to stop the killing. The Australasian Bat Society Newsletter 24
Botvinkin AD, Poleschuk EM, Kuzmin IV et al (2003) Novel lyssaviruses isolated from bats in Russia. Emerg Infect Dis 9(12):1623
Botvinkin A, Selnikova O, Antonova L, Moiseeva A, Nesterenko EY, Gromashevsky L (2005) Human rabies case caused from a bat bite in Ukraine. Rabies Bull Eur 29(3):5–7
Boulger L, Porterfield J (1958) Isolation of a virus from Nigerian fruit bats. Trans R Soc Trop Med Hyg 52(5):421–424
Bourhy H, Kissi B, Lafon M, Sacramento D, Tordo N (1992) Antigenic and molecular characterization of bat rabies virus in Europe. J Clin Microbiol 30(9):2419–2426
Brandão PE, Scheffer K, Villarreal LY et al (2008) A coronavirus detected in the vampire bat Desmodus rotundus. Braz J Infect Dis 12(6):466–468
Breed AC, Field HE, Epstein JH, Daszak P (2006) Emerging henipaviruses and flying foxes–conservation and management perspectives. Biol Conserv 131(2):211–220
Breed AC, Meers J, Sendow I et al (2013) The distribution of Henipaviruses in Southeast Asia and Australasia: Is Wallace’s line a barrier to Nipah Virus? PLoS ONE 8(4):e61316
Brook CE, Dobson AP (2015) Bats as ‘special’ reservoirs for emerging zoonotic pathogens. Trends Microbiol 23:172–180
Brookes S, Healy D, Fooks A (2005) Ability of rabies vaccine strains to elicit cross-neutralising antibodies. Dev Biol (Basel) 125:185–193
Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T (2006) Bats: important reservoir hosts of emerging viruses. Clin Microbiol Rev 19(3):531–545
Carini A (1911) About one large epizootie of rabies. Ann Inst Pasteur 25:843–846
Carrington CV, Foster JE, Zhu HC et al (2008) Detection and phylogenetic analysis of group 1 coronaviruses in South American bats. Emerg Infect Dis 14 (12):1890
Ceballos NA, Morón SV, Berciano JM et al (2013) Novel lyssavirus in bat, Spain. Emerg Infect Dis 19(5):793
Center for Disease Control and Prevention (1990) Epidemiologic notes and reports updates: filovirus infection in animal handlers. Morb Mortal Wkly Rep 39:221
Center for Disease Control and Prevention (1999) Outbreak of Hendra-like virus–Malaysia and Singapore, 1998–1999. Morb Mortal Wkly Rep 48:265
Chadha MS, Comer JA, Lowe L et al (2006) Nipah virus-associated encephalitis outbreak, Siliguri, India. Emerg Infect Dis 12(2):235–240
Chen L, Liu B, Yang J, Jin Q (2014) DBatVir: the database of bat-associated viruses. Database:bau021. doi:10.1093/database/bau021
Choisy M, Rohani P (2006) Harvesting can increase severity of wildlife disease epidemics. Proc R Soc B: Biol Sci 273(1597):2025–2034
Chong HT, Tan C, Goh K, Lam S, Chua K (2003) The risk of human Nipah virus infection directly from bats (Pteropus hypomelanus) is low. Neurol J Southeast Asia 8:31–34
Chowdhury S, Khan SU, Crameri G et al (2014) Serological evidence of Henipavirus exposure in cattle, goats and pigs in Bangladesh. PLoS Neglected Trop Dis 8:e3302
Chua KB, Goh KJ, Wong KT et al (1999) Fatal encephalitis due to Nipah virus among pig-farmers in Malaysia. Lancet 354(9186):1257–1259
Chua K, Wang L, Lam SK et al (2001) Tioman virus, a novel paramyxovirus isolated from fruit bats in Malaysia. Virology 283 (2):215–229
Chua KB, Lek Koh C, Hooi PS et al (2002a) Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect 4(2):145–151
Chua KB, Chua BH, Wang CW (2002b) Anthropogenic deforestation, El Nino and the emergence of Nipah virus in Malaysia. Malays J Pathol 24(1):15–21
Clayton BA, Wang LF, Marsh GA (2013) Henipaviruses: an updated review focusing on the pteropid reservoir and features of transmission. Zoonoses Public Health 60(1):69–83
Constantine DG, Blehert DS (2009) Bat rabies and other lyssavirus infections. US Geological Survey
Dacheux L, Larrous F, Mailles A et al (2009) European bat lyssavirus transmission among cats, Europe. Emerg Infect Dis 15(2):280
Daszak P, Cunningham AA, Hyatt AD (2000) Emerging infectious diseases of wildlife–threats to biodiversity and human health. Science 287(5452):443–449
Daszak P, Cunningham AA, Hyatt AD (2001) Anthropogenic environmental change and the emergence of infectious diseases in wildlife. Acta Trop 78:103–116
Daszak P, Plowright R, Epstein J et al (2006) The emergence of Nipah and Hendra virus: pathogen dynamics across a wildlife-livestock-human continuum. Dis Ecol: Community Struct Pathog Dyn: 186–201
Dominguez SR, O’Shea TJ, Oko LM, Holmes KV (2007) Detection of group 1 coronaviruses in bats in North America. Emerg Infect Dis 13(9):1295
Donaldson EF, Haskew AN, Gates JE, Huynh J, Moore CJ, Frieman MB (2010) Metagenomic analysis of the viromes of three North American bat species: viral diversity among different bat species that share a common habitat. J Virol 84(24):13004–13018
Donnelly CA, Woodroffe R, Cox D et al (2005) Positive and negative effects of widespread badger culling on tuberculosis in cattle. Nature 439(7078):843–846
Drexler JF, Gloza-Rausch F, Glende J et al (2010) Genomic characterization of severe acute respiratory syndrome-related coronavirus in European bats and classification of coronaviruses based on partial RNA-dependent RNA polymerase gene sequences. J Virol 84(21):11336–11349
Drexler JF, Corman VM, Müller MA et al (2012) Bats host major mammalian paramyxoviruses. Nat Commun 3:796
Dzikwi AA, Kuzmin II, Umoh JU, Kwaga JK, Ahmad AA, Rupprecht CE (2010) Evidence of Lagos bat virus circulation among Nigerian fruit bats. J Wildl Dis 46(1):267–271
Enserink M (2000) Malaysian researchers trace Nipah virus outbreak to bats. Science 289(5479):518–519
Epstein JH, Field HE, Luby S, Pulliam JR, Daszak P (2006) Nipah virus: impact, origins, and causes of emergence. Curr Infect Dis Rep 8(1):59–65
Epstein JH, Olival KJ, Pulliam JRC et al (2009) Pteropus vampyrus, a hunted migratory species with a multinational home-range and a need for ro regional management. J Appl Ecol 46:991–1002
Falcón A, Vázquez-Morón S, Casas I et al (2011) Detection of alpha and betacoronaviruses in multiple Iberian bat species. Arch Virol 156(10):1883–1890
Familusi JB, Moore DL (1972) Isolation of a rabies related virus from the cerebrospinal fluid of a child with ‘aseptic meningitis’. Afr J Med Sci 3(1):93–96
Familusi JB, Osunkoya BO, Moore DL, Kemp GE, Fabiyi A (1972) A fatal human infection with Mokola virus. Am J Trop Med Hyg 21(6):959–963
Fenton MB (1997) Science and the conservation of bats. J Mammal: 1–14
Field H, Young P, Yob JM, Mills J, Hall L, Mackenzie J (2001) The natural history of Hendra and Nipah viruses. Microbes Infect 3(4):307–314
Field HE (2009) Bats and emerging zoonoses: Henipaviruses and SARS. Zoonoses Public Health 56:278–284
Field H, Schaaf K, Kung N et al (2010) Hendra virus outbreak with novel clinical features, Australia. Emerg Infect Dis 16(2):338
Field H, Kung N (2011) Henipaviruses—unanswered questions of lethal zoonoses. Curr Opin Virol 1(6):658–661
Foggin C (1988) Rabies and rabies-related viruses in Zimbabwe: historical, virological and ecological aspects. University of Zimbabwe
Food and Agricultural Organisation of the United Nations (2011) Investigating the role of bats in emerging zoonoses: balancing ecology, conservation and public health interests. In: Newman SH, Field HE, de Jong CE, Epstein JH (eds) FAO animal production and health manual no 12, Rome
Fooks AR, McElhinney LM, Pounder DJ et al (2003) Case report: isolation of a European bat lyssavirus type 2a from a fatal human case of rabies encephalitis. J Med Virol 71(2):281–289
Francis JR, Nourse C, Vaska VL et al (2014) Australian bat lyssavirus in a child: the first reported case. Pediatrics 133(4):e1063–e1067
Fraser GC, Hooper PT, Lunt RA et al (1996) Encephalitis caused by a Lyssavirus in fruit bats in Australia. Emerg Infect Dis 2(4):327
Freuling CM, Beer M, Conraths FJ et al (2011) Novel lyssavirus in Natterer’s bat, Germany. Emerg Infect Dis 17(8):1519–1522
Frick WF, Pollock JF, Hicks AC et al (2010) An emerging disease causes regional population collapse of a common North American bat species. Science 329:679–682
Frick WF, Puechmaille SJ, Willis CKR (2016) White-nose syndrome in bats. In: Voigt CC, Kingston T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 245–259
Furey N, Racey P (2016) Conservation ecology of cave bats. In: Voigt, CC, Kingston, T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 463–492
Gatherer D (2014) The 2014 Ebola virus disease outbreak in West Africa. J Gen Virol: vir 0.067199-067190
Ge X-Y, Li J-L, Yang X-L et al (2013) Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 503:535–538
George DB, Webb CT, Farnsworth ML et al (2011) Host and viral ecology determine bat rabies seasonality and maintenance. Proc Natl Acad Sci 108(25):10208–10213
Ghanem SJ, Voigt CC (2012) Increasing awareness of ecosystem services provided by bats. Adv Study Behav 44:279–302
Gonzalez J-P, Herbreteau V, Morvan J, Leroy EM (2005) Ebola virus circulation in Africa: a balance between clinical expression and epidemiological silence. Bull de la Société de Pathol Exotique 98(3):210–217
Gould AR, Kattenbelt JA, Gumley SG, Lunt RA (2002) Characterisation of an Australian bat lyssavirus variant isolated from an insectivorous bat. Virus Res 89(1):1–28
Goldspink LK, Edson D, Vidgen M, Bingham J, Field H, Smith C (2015) Natural Hendra virus infection in flying-foxes, tissue tropism and risk factors. PLoS ONE 10(6):e0128835
Greenhall AM (1964) Bats: their public health importance and control with special reference to Trinidad. In: Proceedings of the 2nd vertebrate pest control conference, p. 18
Guan Y, Zheng B, He Y et al (2003) Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 302(5643):276–278
Gurley ES, Montgomery JM, Hossain MJ et al (2007) Person-to-person transmission of Nipah virus in a Bangladeshi community. Emerg Infect Dis 13(7):1031
Halpin K, Young P, Field H, Mackenzie J (2000) Isolation of Hendra virus from pteropid bats: a natural reservoir of Hendra virus. J Gen Virol 81(8):1927–1932
Halpin K, Hyatt AD, Plowright RK et al (2007) Emerging viruses: coming in ona wrinkled wing and a prayer. Clin Infect Dis 44:711–717
Halpin K, Hyatt AD, Fogarty R et al (2011) Pteropid bats are confirmed as the reservoir hosts of henipaviruses: a comprehensive experimental study of virus transmission. Am J Trop Med Hyg 85(5):946–951
Halpin K, Rota P (2015) A review of Hendra virus and Nipah virus infections in man and other animals. In: Zoonoses-infections affecting humans and animals. Springer Netherlands, pp. 997–1012
Hanlon CA, Kuzmin IV, Blanton JD, Weldon WC, Manangan JS, Rupprecht CE (2005) Efficacy of rabies biologics against new lyssaviruses from Eurasia. Virus Res 111(1):44–54
Hanna JN, Carney IK, Smith GA et al (2000) Australian bat lyssavirus infection: a second human case, with a long incubation period. Med J Aust 172(12):597–599
Harit AK, Ichhpujani RL, Gupta S et al (2006) Nipah/Hendra virus outbreak in Siliguri, West Bengal, India in 2001. Indian J Med Res 123(4):553–560
Hayman DT, Fooks AR, Horton D et al (2008) Antibodies against Lagos bat virus in megachiroptera from West Africa. Emerg Infect Dis 14(6):926
Hayman DT, Emmerich P, Yu M et al (2010) Long-term survival of an urban fruit bat seropositive for Ebola and Lagos bat viruses. PLoS ONE 5(8):e11978
Hayman DT, Fooks AR, Rowcliffe JM et al (2012) Endemic Lagos bat virus infection in Eidolon helvum. Epidemiol Infect 140(12):2163–2171
Hemida MG, Chu D, Poon L, Perera R, Alhammadi M, Ng H (2014) MERS coronavirus in dromedary camel herd, Saudi Arabia. Emerg Infect Dis 20(7):1231–1234
Hooper P, Lunt R, Gould A et al (1997) A new lyssavirus—fhe first endemic-related virus recognized in Australia. Bull de l’Institut Pasteur 95(4):209–218
Horton DL, Banyard AC, Marston DA et al (2014) Antigenic and genetic characterization of a divergent African virus, Ikoma lyssavirus. J Gen Virol 95:1025–1032
Hsu VP, Hossain MJ, Parashar UD et al (2004) Nipah virus encephalitis reemergence, Bangladesh. Emerg Infect Dis 10(12):2082–2087
Hughes JM, Wilson ME, Luby SP, Gurley ES, Hossain MJ (2009) Transmission of human infection with Nipah virus. Clin Infect Dis 49(11):1743–1748
Hutson AM, Mickleburgh SP (2001) Microchiropteran bats: global status survey and conservation action plan, vol 56. IUCN
Iehlé C, Razafitrimo G, Razainirina J (2007) Henipavirus and Tioman virus antibodies in pteropodid bats, Madagascar. Emerg Infect Dis 13:159
Ithete NL, Stoffberg S, Corman VM et al (2013) Close relative of human Middle East respiratory syndrome coronavirus in bat, South Africa. Emerg Infect Dis 19(10):1697
Jackson FR, Turmelle AS, Farino DM, Franka R, McCracken GF, Rupprecht CE (2008) Experimental rabies virus infection of big brown bats (Eptesicus fuscus). J Wildl Dis 44(3):612–621
Jacob H, Solcher H (1968) An infectious disease transmitted by Cercopithecus aethiops (“marburg disease”) with glial nodule encephalitis. Acta Neuropathol 11(1):29
Jahrling P, Geisbert T, Johnson E, Peters C, Dalgard D, Hall W (1990) Preliminary report: isolation of Ebola virus from monkeys imported to USA. Lancet 335(8688):502–505
Jones BA, Grace D, Kock R et al (2013) Zoonosis emergence linked to agricultural intensification and environmental change. Proc Natl Acad Sci 110(21):8399–8404
Kamins AO, Rowcliffe JM, Ntiamoa-Baidu Y, Cunningham AA, Wood JL, Restif O (2014) Characteristics and risk perceptions of Ghanaians potentially exposed to bat-borne zoonoses through bushmeat. EcoHealth:1–17
Kingston T (2016) Cute, Creepy, or Crispy–how values, attitudes and norms shape human behavior toward bats. In: Voigt, CC, Kingston, T (eds) Bats in the anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 571–588
Knight AJ (2008) “Bats, snakes and spiders, Oh my!” How aesthetic and negativistic attitudes, and other concepts predict support for species protection. J Environ Psychol 28(1):94–103
Kurth A, Kohl C, Brinkmann A et al (2012) Novel paramyxoviruses in free-ranging European bats. PLoS ONE 7(6):e38688
Kuzmin IV, Botvinkin AD, Rybin SN, Baialiev AB (1991) A lyssavirus with an unusual antigenic structure isolated from a bat in southern Kyrgyzstan. Vopr Virusol 37(5–6):256–259
Kuzmin IV, Botvinkin AD, Khabilov TK (2001) The lyssavirus was isolated from a whiskered bat in northern Tajikistan. Plecotus et al 4:75–81
Kuzmin IV, Niezgoda M, Carroll DS et al (2006) Lyssavirus surveillance in bats, Bangladesh. Emerg Infect Dis 12(3):486
Kuzmin IV, Niezgoda M, Franka R et al (2008a) Possible emergence of West Caucasian bat virus in Africa. Emerg Infect Dis 14(12):1887
Kuzmin IV, Niezgoda M, Franka R et al (2008b) Lagos bat virus in Kenya. J Clin Microbiol 46(4):1451–1461
Kuzmin IV, Mayer AE, Niezgoda M et al (2010) Shimoni bat virus, a new representative of the Lyssavirus genus. Virus Res 149(2):197–210
Kuzmin IV, Bozick B, Guagliardo SA et al (2011a) Bats, emerging infectious diseases, and the rabies paradigm revisited. Emerg Health Threats J 4
Kuzmin IV, Turmelle AS, Agwanda B et al (2011b) Commerson’s leaf-nosed bat (Hipposideros commersoni) is the likely reservoir of Shimoni bat virus. Vector-Borne Zoonotic Dis 11(11):1465–1470
Kuzmin IV (2014) Basic facts about Lyssaviruses. Curr Lab Tech Rabies Diagn Res Prev 1:3
Lau SK, Woo PC, Wong BH et al (2010) Identification and complete genome analysis of three novel paramyxoviruses, Tuhoko virus 1, 2 and 3, in fruit bats from China. Virology 404(1):106–116
Leonova GN, Belikov SI, Kondratov IG et al (2009) A fatal case of bat lyssavirus infection in Primorye territory of the Russian far East. Rabies Bull Europe 33(4):5–8
Leroy EM, Rouquet P, Formenty P et al (2004) Multiple Ebola virus transmission events and rapid decline of central African wildlife. Science 303(5656):387–390
Leroy EM, Kumulungui B, Pourrut X et al (2005) Fruit bats as reservoirs of Ebola virus. Nature 438(7068):575–576
Leroy EM, Epelboin A, Mondonge V et al (2009) Human Ebola outbreak resulting from direct exposure to fruit bats in Luebo, democratic Republic of Congo, 2007. Vector-Borne Zoonotic Dis 9(6):723–728
Li W, Shi Z, Yu M et al (2005) Bats are natural reservoirs of SARS-like coronaviruses. Science 310(5748):676–679
Li Y, Chen S (2014) Evolutionary history of Ebola virus. Epidemiol Infect 142(06):1138–1145
Luby SP, Rahman M, Hossain MJ et al (2006) Foodborne transmission of Nipah virus, Bangladesh. Emerg Infect Dis 12(12):1888
Luby SP, Hossain MJ, Gurley ES et al (2009) Recurrent zoonotic transmission of Nipah virus into humans, Bangladesh, 2001–2007. Emerg Infect Dis 15(8):1229
Luby SP, Gurley ES (2012) Epidemiology of henipavirus disease in humans. In: Henipavirus. Springer Berlin Heidelberg, pp. 25–40
Luis AD, Hayman DT, O’Shea TJ et al (2013) A comparison of bats and rodents as reservoirs of zoonotic viruses: are bats special? Proc R Soc B: Biol Sci 280(1756):20122753
Lumio J, Hillbom M, Roine R et al (1986) Human rabies of bat origin in Europe. Lancet 327(8477):378
Lumlertdacha B, Boongird K, Wanghongsa S et al (2005) Survey for bat lyssaviruses, Thailand. Emerg Infect Dis 11(2):232
Markotter W, Kuzmin IV, Rupprecht CE, Nel LH (2009) Lagos bat virus virulence in mice inoculated by the peripheral route. Epidemiol Infect 137(8):1155
Marsh GA, Haining J, Robinson R et al (2011) Ebola Reston virus infection of: clinical significance and transmission potential. J Infect Dis 204(Suppl 3):S804–S809
Marsh GA, De Jong C, Barr JA et al (2012) Cedar virus: a novel henipavirus isolated from Australian bats. PLoS Pathog 8(8):e1002836
Marston DA, Horton DL, Ngeleja C et al (2012) Ikoma lyssavirus, highly divergent novel lyssavirus in an African civet. Emerg Infect Dis 18(4):664–667
Martin G, Plowright R, Chen C, Kault D, Selleck P, Skerratt L (2015) Hendra virus survival does not explain spillover patterns and implicates relatively direct transmission routes from flying foxes to horses. J Gen Virol 96(6):1229–1237
Mayen F (2003) Haematophagous bats in Brazil, their role in rabies transmission, impact on public health, livestock industry and alternatives to an indiscriminate reduction of bat population. J Vet Med Ser B 50(10):469–472
McColl KA, Tordo N, Aguilar SA (2000) Bat lyssavirus infections. Revue scientifique et technique (Int Office Epizootics) 19(1):177–196
McColl K, Chamberlain T, Lunt R, Newberry K, Middleton D, Westbury H (2002) Pathogenesis studies with Australian bat lyssavirus in grey-headed flying foxes (Pteropus poliocephalus). Aust Vet J 80(10):636–641
Medellin RA (2003) Diversity and conservation of bats in Mexico: research priorities, strategies and Actions. Wildl Soc Bull 31:87–97
Memish ZA, Mishra N, Olival KJ et al (2013) Middle East respiratory syndrome coronavirus in bats, Saudi Arabia. Emerg Infect Dis 19(11):1819
Memish ZA, Cotten M, Meyer B et al (2014) Human infection with MERS coronavirus after exposure to infected camels, Saudi Arabia, 2013. Emerg Infect Dis 20(6):1012
Meredith C, Prossouw A, Koch H (1971) An unusual case of human rabies thought to be of chiropteran origin. South African Med J=Suid-Afrikaanse tydskrif vir geneeskd 45(28):767
Messenger SL, Rupprecht CE, Smith JS (2003) Bats, emerging virus infections, and the rabies paradigm. In: Kunz TH, Fenton MB (eds) Bat ecology. The University of Chicago Press, Chicago, pp 622–679
Mickleburgh SP, Hutson AM, Racey PA (2002) A review of the global conservation status of bats. Oryx 36(01):18–34
Mickleburgh S, Waylen K, Racey P (2009) Bats as bushmeat: a global review. Oryx 43(02):217–234
Middleton DJ, Weingartl HM (2012) Henipaviruses in their natural animal hosts. In: Henipavirus. Springer Berlin Heidelberg, pp 105–121
Mildenstein T, Tanshi I, Racey PA (2016) Exploitation of bats for bushmeat and medicine. In: Voigt, CC, Kingston, T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp 325–376
Misra V, Dumonceaux T, Dubois J et al (2009) Detection of polyoma and corona viruses in bats of Canada. J Gen Virol 90(8):2015–2022
Müller T, Cox J, Peter W et al (2004) Spill-over of European bat Lyssavirus type 1 into a Stone Marten (Martes foina) in Germany. J Vet Med Ser B 51(2):49–54
Müller MA, Paweska JT, Leman PA et al (2007) Coronavirus antibodies in African bat species. Emerg Infect Dis 13(9):1367
Murray K, Selleck P, Hooper P et al (1995) A morbillivirus that caused fatal disease in horses and humans. Science 268(5207):94–97
Nahar N, Sultana R, Gurley ES, Hossain MJ, Luby SP (2010) Date palm sap collection: exploring opportunities to prevent Nipah transmission. EcoHealth 7(2):196–203
Nahar N, Mondal UK, Sultana R et al (2013) Piloting the use of indigenous methods to prevent Nipah virus infection by interrupting bats’ access to date palm sap in Bangladesh. Health Promot Int 28(3):378–386
Nathwani D, McIntyre P, White K et al (2003) Fatal human rabies caused by European bat Lyssavirus type 2a infection in Scotland. Clin Infect Dis 37(4):598–601
Negredo A, Palacios G, Vázquez-Morón S et al (2011) Discovery of an ebolavirus-like filovirus in europe. PLoS Pathog 7:e1002304
Negrete OA, Levroney EL, Aguilar HC et al (2005) EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 436(7049):401–405
Nel LH (2001) Mokola virus: a brief review of the status quo. In Sixth SEARG meeting, Lilongwe/Malawi:80–86
O’Shea TJ, Cryan PM, Cunningham AA et al (2014) Bat flight and zoonotic viruses. Emerg Infect Dis 20(5):741
O’Sullivan J, Allworth A, Paterson D et al (1997) Fatal encephalitis due to novel paramyxovirus transmitted from horses. Lancet 349:93–95
Olival KJ, Islam A, Yu M et al (2013) Ebola virus antibodies in fruit bats, Bangladesh. Emerg Infect Dis 19:270–273
Olival KJ, Hayman DTS (2014) Filoviruses in bats: current knowledge and future directions. Viruses 6:1759–1788
Parashar UD, Sunn LM, Ong F et al (2000) Case-control study of risk factors for human infection with a new zoonotic paramyxovirus, Nipah virus, during a 1998–1999 outbreak of severe encephalitis in Malaysia. J Infect Dis 181:1755–1759
Paton NI, Leo YS, Zaki SR et al (1999) Outbreak of Nipah-virus infection among abattoir workers in Singapore. Lancet 354:1253–1256
Pawan J (1959) Rabies in the vampire bat of Trinidad with special reference to the clinical course and the latency of infection. Carib Med J 21:137–156
Paweska JT, Blumberg LH, Liebenberg C et al (2006) Fatal human infection with rabies-related Duvenhage virus, South Africa. Emerg Infect Dis 12:1965–1967
Peel AJ, Feldman K, Hayman DTS et al (2010) Widespread seroprevalence to Lagos bat virus and henipavirus in straw coloured fruit bats (Eidolon helvum) across East Africa. In: Proceedings of the 2nd International Berlin bat meeting: bat biology and infectious diseases, pp 15–16
Peel AJ, Sargan DR, Baker KS et al (2013) Continent-wide panmixia of an African fruit bat facilitates transmission of potentially zoonotic viruses. Nat Commun 4:2770
Pernet O, Schneider BS, Beaty SM et al (2014) Evidence for henipavirus spillover into human populations in Africa. Nat Commun 5:5342
Pfefferle S, Oppong S, Drexler JF et al (2009) Distant relatives of severe acute respiratory syndrome coronavirus and close relatives of human coronavirus 229E in bats, Ghana. Emerg Infect Dis 15:1377–1384
Philbey AW, Kirkland PD, Ross AD et al (1998) An apparently new virus (family Paramyxoviridae) infectious for pigs, humans, and fruit bats. Emerg Infect Dis 4:269–271
Pigott DM, Golding N, Mylne A et al (2014) Mapping the zoonotic niche of Ebola virus disease in Africa. Elife 3:e04395
Plowright RK, Foley P, Field HE et al (2011) Urban habituation, ecological connectivity and epidemic dampening: the emergence of Hendra virus from flying foxes (Pteropus spp.). Proc R Soc B 278:3703–3712
Plowright RK, Eby P, Hudson PJ et al (2015) Ecological dynamics of emerging bat virus spillover. Proc R Soc B 282(1798):20142124
Pooley S, Fa JE, Nasi R (2015) No conservation silver lining to Ebola. Conserv Biol 29(3):965–967
Poon L, Chu D, Chan K et al (2005) Identification of a novel coronavirus in bats. J Virol 79:2001–2009
Pourrut X, Kumulungui B, Wittmann T et al (2005) The natural history of Ebola virus in Africa. Microbes Infect/Inst Pasteur 7:1005–1014
Pourrut X, Souris M, Towner JS et al (2009) Large serological survey showing cocirculation of Ebola and Marburg viruses in Gabonese bat populations, and a high seroprevalence of both viruses in Rousettus aegyptiacus. BMC Infect Dis 9:159
Puechmaille SJ, Verdeyroux P, Fuller H, Gouilh MA, Bekaert M, Teeling EC (2010) White-nose syndrome fungus (Geomyces destructans) in bat, France. Emerg Infect Dis 16(2):290
Quammen D (2013) Spillover: animal infections and the next human pandemic. WW Norton & Company
Quan P-L, Firth C, Street C et al (2010) Identification of a severe acute respiratory syndrome coronavirus-like virus in a leaf-nosed bat in Nigeria. MBio 1:e00208–e00210
Racey PA, Hutson AM, Lina PHC (2012) Bat rabies, public health and european bat conservation. Zoonoses Public Health (Special Issue—Bats). 60:58–68
Racey PA (2015) The uniqueness of bats. In: Lin-Fa W, Cowled C (eds) Bats and Viruses: A New Frontier of Emerging Infectious Diseases. Wiley Blackwell. p. 1–22
Rahman SA, Hassan SS, Olival KJ et al (2010) Characterization of Nipah virus from naturally infected Pteropus vampyrus bats, Malaysia. Emerg Infect Dis 16(12):1990
Rahman MA, Hossain MJ, Sultana S et al (2012) Date palm sap linked to Nipah virus outbreak in Bangladesh, 2008. Vector-Borne Zoonotic Dis 12:65–72
Reynes J-M, Counor D, Ong S et al (2005) Nipah virus in Lyle’s flying foxes, Cambodia. Emerg Infect Dis 11:1042–1047
Rihtarič D, Hostnik P, Steyer A, Grom J, Toplak I (2010) Identification of SARS-like coronaviruses in horseshoe bats (Rhinolophus hipposideros) in Slovenia. Arch Virol 155:507–514
Roberts BJ, Catterall CP, Eby P, Kanowski J (2012) Long-distance and frequent movements of the flying-fox Pteropus poliocephalus: implications for management. PLoS ONE 7:e42532
Rogers RJ, Douglas IC, Baldock FC et al (1996) Investigation of a second focus of equine morbillivirus infection in coastal Queensland. Aust Vet J 74(3):243–244
Roine R, Hillbom M, Valle M et al (1988) Fatal encephalitis caused by a bat-borne rabies-related virus. Clinical findings. Brain: a journal of neurology 111:1505–1516
Rønsholt L, Sørensen K, Bruschke C et al (1998) Clinically silent rabies infection in (zoo) bats. Vet Rec 142:519–520
Rota PA, Oberste MS, Monroe SS et al (2003) Characterization of a novel coronavirus associated with severe acute respiratory syndrome. Science 300:1394–1399
Roué SY, Nemoz M (2004) Unusual mortality in Schreiber’s long fingered bat, at several nurseries. Wild Dis Assoc Newsl 14:4
Ruiz M, Chávez CB (2010) Rabies in Latin America. Neurol Res 32:272–277
Rupprecht CE, Gibbons RV (2004) Prophylaxis against rabies. New Engl J Med 351:2626–2635
Saéz AM, Weiss S, Nowak K, Lapeyre V, Zimmermann F et al (2015) Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol Med 7(1):17–23
Salmón-Mulanovich G, Vásquez A, Albújar C, Guevara C (2009) Human rabies and rabies in vampire and nonvampire bat species, Southeastern Peru, 2007. Emerg Inf Dis 15:1308–1310
Samaratunga H, Searle J, Hudson N (1998) Non-rabies Lyssavirus human encephalitis from fruit bats: Australian bat Lyssavirus (pteropid Lyssavirus) infection. Neuropathol Appl Neurobiol 24:331–335
Sasaki M, Setiyono A, Handharyani E et al (2012) Molecular detection of a novel paramyxovirus in fruit bats from Indonesia. Virol J 9:240
Schatz J, Fooks AR, McElhinney L et al (2013) Bat rabies surveillance in Europe. Zoonoses Public Health 60(1):22–34
Schneeberger K, Czirják GÁ, Voigt CC (2013) Measures of the constitutive immune system are linked to diet and roosting habits of neotropical bats. PLoS ONE 8:e54023
Schneider L, Barnard B, Schneider H et al (1985) Application of monoclonal antibodies for epidemiological investigations and oral vaccination studies. In: Rabies in the tropics. Springer, Verlag
Schneider MC, Romijn PC, Uieda W et al (2009) Rabies transmitted by vampire bats to humans: an emerging zoonotic disease in Latin America? Rev panam de salud pública 25:260–269
Schnell MJ, McGettigan JP, Wirblich C, Papaneri A (2009) The cell biology of rabies virus: using stealth to reach the brain. Nat Rev Microbiol 8:51–61
Selimov MA, Tatarov AG, Botvinkin AD et al (1989) Rabies-related Yuli virus; identification with a panel of monoclonal antibodies. Acta Virol 33(6):542–546
Serra-Cobo J, Amengual B, Abellán C, Bourhy H (2002) European bat lyssavirus infection in Spanish bat populations. Emerg Inf Dis 8:413–420
Sétien A, Brochier B, Tordo N et al (1998) Experimental rabies infection and oral vaccination in vampire bats (Desmodus rotundus). Vaccine 16:1122–1126
Smith I, Wang LF (2013) Bats and their virome: an important source of emerging viruses capable of infecting humans. Curr Opin Virol 3(1):84–91
Smith C, Skelly C, Kung N, Roberts B, Field H (2014) Flying-fox species density-a spatial risk factor for Hendra virus infection in horses in Eastern Australia. PLoS ONE 9(6):e99965
Speare R, Skerratt L, Foster R et al (1997) Australian bat lyssavirus infection in three fruit bats from north Queensland. Commun Dis Intell 21:117–119
Steece R, Altenbach JS (1989) Prevalence of rabies specific antibodies in the Mexican free-tailed bat (Tadarida brasiliensis mexicana) at Lava Cave, New Mexico. J Wildl Dis 25:490–496
Streicker DG, Turmelle AS, Vonhof MJ et al (2010) Host phylogeny constrains cross-species emergence and establishment of rabies virus in bats. Science 329(5992):676–679
Streicker DG, Recuenco S, Valderrama W et al (2012) Ecological and anthropogenic drivers of rabies exposure in vampire bats: implications for transmission and control. Proc R Soc B 279:3384–3392
Swanepoel R, Leman PA, Burt FJ et al (1996) Experimental inoculation of plants and animals with Ebola virus. Emerg Inf Dis 2:321–325
Tang X, Zhang J, Zhang S et al (2006) Prevalence and genetic diversity of coronaviruses in bats from China. J Virol 80:7481–7490
Taniguchi S, Watanabe S, Masangkay JS et al (2011) Reston Ebolavirus antibodies in bats, the Philippines. Emerg Infect Dis 17:1559–1560
Thiriet D (2011) Conservation shouldn’t be a popularity contest. Conversation:1–4
Tjørnehøj K, Fooks AR, Agerholm JS, Rønsholt L (2006) Natural and experimental infection of sheep with European bat lyssavirus type-1 of Danish bat origin. J Comp Pathol 134(2):190–201
Tong S, Conrardy C, Ruone S et al (2009) Detection of novel SARS-like and other coronaviruses in bats from Kenya. Emerg Inf Dis 15:482–485
Tong S, Zhu X, Li Y et al (2013) New world bats harbor diverse influenza A viruses. PLoS Pathog 9(10):e1003657
Towner JS, Pourrut X, Albariño CG et al (2007) Marburg virus infection detected in a common African bat. PLoS ONE 2:e764
Towner JS, Amman BR, Sealy TK et al (2009) Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog 5:e1000536
Trimarchi C, Debbie J (1977) Naturally occurring rabies virus and neutralizing antibody in two species of insectivorous bats of New York State. J Wildl Dis 13:366–369
Turmelle A, Jackson F, Green D, McCracken G, Rupprecht C (2010) Host immunity to repeated rabies virus infection in big brown bats. J Gen Virol 91:2360–2366
van Thiel P, Van den Hoek J, Eftimov F et al (2008) Fatal case of human rabies (Duvenhage virus) from a bat in Kenya: The Netherlands, December 2007. Euro Surveill 13:118
van Thiel P-PA, de Bie RM, Eftimov F et al (2009) Fatal human rabies due to Duvenhage virus from a bat in Kenya: failure of treatment with coma-induction, ketamine, and antiviral drugs. PLoS Negl Trop Dis 3:e428
Vidgen ME, de Jong C, Rose K et al (2015) Novel paramyxoviruses in Australian flying-fox populations support host-virus coevolution. J Gen Virol 96(7):1619–1625
Virgin HW, Wherry EJ, Ahmed R (2009) Redefining chronic viral infection. Cell 138(1):30–50
Vogel G (2014) Are bats spreading ebola across sub-saharan Africa? Science 344:140
Voigt, CC, Phelps KL, Aguirre L, Schoeman MC, Vanitharani J, Zubaid A (2016) Bats and buildings: the conservation of synanthropic bats. In: Voigt, CC, Kingston, T (eds) Bats in the Anthropocene: conservation of bats in a changing world. Springer International AG, Cham, pp. 427–453
Wacharapluesadee S, Lumlertdacha B, Boongird K et al (2005) Bat Nipah virus, Thailand. Emerg Inf Dis 11:1949–1951
Warrilow D (2005) Australian bat lyssavirus: a recently discovered new rhabdovirus. In: Fu ZF (ed) The world of Rhabdoviruses. Springer, Berlin
Watanabe S, Masangkay JS, Nagata NK et al (2010) Bat coronaviruses and experimental infection of bats, the Philippines. Emerg Inf Dis 16:1217–1223
Wilkinson GS, South JM (2002) Life history, ecology and longevity in bats. Aging Cell 1(2):124–131
Woodroffe R (1999) Managing disease threats to wild mammals. Anim Conserv 2(03):185–193
World Health Organisation (2003) Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003 (http://www.who.int/csr/sars/country/table2004_04_21)
World Health Organization (2014) Ebola virus disease, Fact Sheet. (http://www.who.int/mediacentre/factsheets/fs103/en)
Wibbelt G, Moore MS, Schountz T, Voigt CC (2010) Emerging diseases in Chiroptera: why bats? Biol Lett 6:438–440
Woo PC, Lau SK, Li KS et al (2006) Molecular diversity of coronaviruses in bats. Virology 351:180–187
Yaiw KC, Crameri G, Wang L et al (2007) Serological evidence of possible human infection with Tioman virus, a newly described paramyxovirus of bat origin. J Infect Dis 196:884–886
Yaiw KC, Bingham J, Crameri G et al (2008) Tioman virus, a paramyxovirus of bat origin, causes mild disease in pigs and has a predilection for lymphoid tissues. J Virol 82:565–568
Yang Y, Du L, Liu C et al (2014) Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus. Proc Nat Acad Sci USA 111:12516–12521
Yob JM, Field H, Rashdi AM et al (2001) Nipah virus infection in bats (order Chiroptera) in peninsular Malaysia. Emerg Inf Dis 7:439–441
Young PL, Halpin K, Selleck PW et al (1996) Serologic evidence for the presence in Pteropus bats of a paramyxovirus related to equine morbillivirus. Emerg Inf Dis 2:239–240
Zaki AM, Van Boheemen S, Bestebroer TM et al (2012) Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. New Engl J Med 367:1814–1820
Zhang G, Cowled C, Shi Z et al (2013) Comparative analysis of bat genomes provides insight into the evolution of flight and immunity. Science 339(6118):456–460
Acknowledgments
We thank Alison Peel, Paul Racey and an anonymous reviewer for constructive comments that helped to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2016 The Author(s)
About this chapter
Cite this chapter
Schneeberger, K., Voigt, C.C. (2016). Zoonotic Viruses and Conservation of Bats. In: Voigt, C., Kingston, T. (eds) Bats in the Anthropocene: Conservation of Bats in a Changing World. Springer, Cham. https://doi.org/10.1007/978-3-319-25220-9_10
Download citation
DOI: https://doi.org/10.1007/978-3-319-25220-9_10
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-25218-6
Online ISBN: 978-3-319-25220-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)