
Abstract
Background
Fabaceae is considered as the third largest family, which includes more than 727 genera and 20,000 species. The genus Vicia has from 180 to 210 species. Vicia species have a great economical and agricultural importance. Karyotype study of chromosomes and SDS-PAGE for seed storage proteins (soluble and non-soluble proteins) were carried out on five Vicia species (Vicia macrocarpa, Vicia sativa, Vicia narbonensis, Vicia ervilia) collected from IPK, Germany, and Vicia faba from Agriculture Research Centre, Giza, Egypt, to find out the phylogenetic relationships among these species.
Result
From karyotype of studied Vicia species chromosomes, it was found that V. macrocarpa, V. sativa, and V. faba had six pairs of chromosome (2n = 12) while V. narbonensis and V. ervilia had seven pairs of chromosome (2n = 14). The most related species was found between V. ervilia and V. narbonensis (77.8%) depending on seed soluble protein similarity level, but between V. narbonensis and V. macrocarpa was 70.0% depending on seed non-soluble protein similarity level, while between V. ervilia and V. narbonensis, the most related species was 69.0% depending on collective data of both soluble and non-soluble seed storage protein.
Conclusion
The phylogenetic relationships between the studied species depending on collective data of protein markers and karyotype characteristic were as follows: V. ervilia is closely related to V. narbonensis, while V. narbonensis is related to V. macrocarpa and V. ervilia, but the degree of relation between V. narbonensis and V. macrocarpa is less than the relation between V. narbonensis and V. ervilia. Equally, while V. sativa is closely related to V. macrocarpa, but V. faba is distant from all other studied species.
Similar content being viewed by others

Introduction
Fabaceae is considered the second family after cereal crops in agricultural importance based on area harvested and total production; this family contains more than 727 genera and 20,000 species (Gepts et al. 2005). The species in genus Vicia (180 to 210 species) are widely distributed throughout the world. This genus has two subgenera, Vicia and Vicilla, and the subgenus Vicilla is considered more primitive and diverse than the subgenus Vicia (Hanelt and Mettin 1989; Maxted 1993). The subgenus Vicilla is divided into 17 sections including forage species. Kupicha (1976) suggested that the subgenus Vicia is smaller and coherent, containing 38 species divided into 5 sections. This subgenus contains the more agriculturally important species of V. faba (section Faba), V. sativa (section Vicia), and V. narbonensis (section Narbonensis).
Karyological studies had an important role in improvement and solving taxonomic problems between the related species (Lavia et al. 2009; Murti et al. 2012). The cytogenetic comparisons based on chromosome size, centromeric index, and banding patterns between related species occurred by staining chromosomes with different dyes such as feulgen, orcein, or carmine (Cremonini 1992; Galasso et al. 1994; Cremonini et al. 1998; Fuchs et al. 1998).
Until recently, cytotaxonomic relationships between species were performed using conventional staining methods to visualize the chromosomes. The development and application of banding techniques for plants have proved to be a practical tool for identifying chromosomes as well as providing much information regarding species relationships. The most popular staining procedures include Q, G, C, R, and silver stain banding which have been developed for bright-field microscopy (Casperson et al. 1970; Howell et al. 1975; Fominaya et al. 1988; Jellen et al. 1993; Jellen and Ladizinsky 2000).
Polyacrylamide gel electrophoresis (PAGE) plays an important role in the analysis of protein profile. PAGE is considered the most widespread technique. Seed storage proteins were used in investigating genetic diversity and evaluation of taxonomic and genetic associations in Vicia (Emre et al. 2010).
The aim of our research is to find out the phylogenetic relationships between five Vicia species (V. macrocarpa (Moris) Betol., V. sativa (L.) ssp. Sativa convar var. sativa, V. narbonensis (L.) var. narbonensis, V. ervilia (L.) Willd, and V. faba var. sakha 3) by studing the karyotype of chromosomes and seed storage protein (soluble and non-soluble proteins) profile of the studied species.
Materials and methods
All the laboratory experiments were carried out in the laboratories of Genetics and Cytology Department, Genetic Engineering and Biotechnology Research Division, National Research Centre, Giza, Egypt.
Plant materials
Seeds of four Vicia species such as V. macrocarpa ((Moris) Betol.), V. sativa L. (ssp. Sativa convar var. sativa), V. narbonensis L. (var. narbonensis), and V. ervilia L. (Willd) were obtained from the germplasm collection of the Institute of Plant Genetics and Crop Plant Research (IPK), Gatersleben, Germany, except for V. faba (var. sakha 3) from the Agricultural Research Center, Giza, Egypt.
Chromosome preparation
The seeds of Vicia species were germinated on moist filter paper at 20 °C. Root tips of about 1–2 cm length were excised. The roots were treated with ice-cold water for 20–22 h to arrest the chromosome at metaphase then fixed in Carnoyʼs solution I (3:1 v/v) absolute ethanol to glacial acetic acid for 24 h at 4 °C, then stored in a refrigerator in 70% ethanol; after that, the roots were incubated in 1% cellulose and 1% pectinase (v/v) which were dissolved in 0.01 M citrate buffer pH 4.8 at 37 °C for 1 h. Root tips were squashed on slides in a drop of 45% acetic acid, frozen on liquid nitrogen to remove the coverslips. After that, slides were washed with Carnoyʼs fixative solution and air dried. The slides were stained with DAPI; after that, they were examined under florescence microscope.
Karyotype characteristics
After photos were captured with a camera connected to a computer, the chromosomes of each cell were arranged using the Adobe Photoshop 6.0 software. After finishing the arrangement of chromosomes of one species, a computer program (Micromeasure 3.3) was used to measure the total length of each chromosome, length of the short arm, length of the long arm, arm ratio (long/short), centromeric index [short/(long + short)], and the relative length (RL) for each chromosome (percentage of total length of haploid complement). The ideograms for the 5 species were drawn in Corel-Draw program.
The procedure of Bebeli and Kaltsikes (1985) was followed to describe the chromosome types in the five species according to the location of the centromere, i.e., metacentric to cover the M-chromosomes with an arm ratio (S:L) between 1:1.35, submetacentric to cover the Sm chromosomes with an arm ratio between 1.36 and 1.75, and subtelocentric to cover the St-chromosomes with an arm ratio greater than 1.76.
Seed storage protein profiles using SDS-PAGE
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to the method proposed by Laemmli (1970), as modified by Studier (1973). Water-soluble proteins (WSP) and water-non-soluble proteins (WNSP) were extracted from the seed of five Vicia species.
Results
Karyotype characteristics
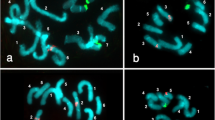
The somatic chromosome number of two species (V. narbonensis and V. ervilia) is 2n = 14, while in the other three species (V. sativa, V. macrocarpa, and V. faba) is 2n = 12. All of the examined species have one secondary constriction except for V. sativa which has two. For karyotypic analysis, chromosomes were captured by a cooled CCD camera and analyzed on a computer with image analysis software (Photoshop 0.6). Chromosomes were randomly numbered, and the total length and lengths of the short arm (S) and long arm (L) were measured for each chromosome. Using chromosome-measuring software, the short and long arms of the homologous chromosome pairs were measured and identification based on chromosome arm ratio (L/S). In the karyogram construction, chromosome pairs were ordered from longest to shortest based on the relative length of each pair of chromosomes. At least 10 well-spread chromosome preparations of each species were analyzed to validate the karyogram construction for each species. The total length and the length of the short arm, long arm, arm ratio, the relative length of each chromosome, and the chromosome types of the five Vicia species are depicted in Table 1. The karyotype of the five Vicia species and their ideograms are shown in Fig. 1.

Karyotype and idiograms of V. macrocarpa, V. sativa, V. narbonensis, V. ervilia, and V. faba (scale = 2 μm)
Vicia macrocarpa
The length of the short arms for the different chromosomes ranged from 7.1 ± 0.83 to 2.47 ± 0.61 μm, whereas the long arms gave average lengths from 9.54 ± 1.21 to 4.44 ± 0.35 μm. Chromosomes no. 1, 5, and 6 are metacentric, having arm ratios of 1.23 ± 0.18, 1.09 ± 0.08, and 1.29 ± 0.21, respectively, while chromosomes no. 2 and 3 are submetacentric, having arm of ratio 1.71 ± 0.39 and 1.65 ± 0.21, respectively. Chromosome no. 4 is subtelocentric, having arm ratio of 3.68 ± 1.08. Chromosome no. 6 has the secondary constriction region on its relatively short arm.
Vicia sativa
The length of the short arms for the different chromosomes ranged from 4.72 ± 0.78 to 1.66 ± 0.21 μm, whereas the long arms gave average lengths from 7.78 ± 1.01 to 3.39 ± 0.29 μm. The chromosome nos. 2, 3, and 5 are metacentric, having arm ratios of 1.18 ± 0.85, 1.29 ± 0.46, and 1.35 ± 0.25, respectively. Chromosome nos. 1, 4, and 6 are subtelocentric, having arm ratios of 2.15 ± 0.38, 2.06 ± 0.60, and 2.07 ± 0.26, respectively.
Vicia narbonensis
The length of the short arms for the different chromosomes ranged from 5.99 ± 0.5 to 4.72 ± 0.57 μm, whereas the long arms gave average lengths from 10.36 ± 0.46 to 6.24 ± 0.52 μm. The chromosome no. 7 is metacentric, having arm ratio of 1.22 ± 0.14%, while chromosome no. 4 is subtelocentric, having arm ratio of 1.76 ± 0.19. Chromosome nos. 1, 2, 3, 5, and 6 are submetacentric, having arm ratios of 1.74 ± 0.16, 1.72 ± 0.26, 1.65 ± 0.26, 1.72 ± 0.26, and 1.55 ± 0.26, respectively.
Vicia ervila
The length of the short arm for the different chromosomes ranged from 7.73 ± 0.46 to 1.98 ± 0.19 μm, whereas the long arms gave average lengths from 8.88 ± 0.33 to 6.13 ± 0.19 μm. Chromosome nos. 1, 2, and 6 are metacentric, having arm ratios of 1.14 ± 0.1, 1.26 ± 0.16, and 1.31 ± 0.1, respectively. Chromosome no. 3 is submetacentric, having an arm ratio of 1.46 ± 0.16. Chromosome nos. 4, 5, and 7 are subtelocentric, having arm ratios of 2.23 ± 0.07, 2.44 ± 0.36, and 3.12 ± 0.40, respectively. Chromosome no. 7 has the secondary constriction region on its long arm.
Vicia faba
The length of the short arms for the different chromosomes ranged from 11.82 ± 1.01 to 1.52 ± 0.25 μm, whereas the long arms gave average lengths from 13.24 ± 1.43 to 8.69 ± 0.74 μm. Chromosome no. 1 is metacentric and has an arm ratio of 1.12 ± 0.12. Chromosome nos. 2, 3, 4, 5, and 6 are subtelocentric, having arm ratios of 7.38 ± 1.18, 6.38 ± 2.04, 2.48 ± 0.42, 6.66 ± 0.72, and 5.4 ± 1.2, respectively.
Seed storage protein profiles using SDS-PAGE
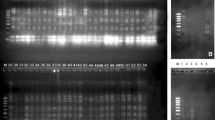
Seed storage proteins (soluble and non-soluble proteins) fractions were used to find out the relationships between the studied five Vicia species (Fig. 2).

Electrophoresis pattern of seed proteins of Vicia species. a Water-soluble proteins (WSP). b Water non-soluble proteins (WNSP). M: protein marker. 1: V. macrocarpa. 2: V. sativa. 3: V. narbonensis. 4: V. ervilia. 5: V. faba
The studied Vicia species were examined for seed storage protein profile, and the data was subjected to unweighted pair-group method with arithmetical average (UPGMA) to find the phylogenetic relationships among the species. Table 2 and Fig. 3 represented the relationship between the studied Vicia species depending on seed soluble protein similarity level, and it was found as follows: V. ervilia and V. narbonensis are the most related species (77.8%), then comes V. macrocarpa which is most related to V. narbonensis (64.5%), V. sativa which is most related to V. macrocarpa (60.0%), and peripheral position comes V. faba which has almost the same relationship with the other studied species, while Table 3 and Fig. 4 showed the relationship between the studied five species depending on seed non-soluble protein similarity level, and it was found as follows: V. narbonensis and V. macrocarpa are the most related species (70.0%), then comes V. sativa, V. ervilia, and V. faba.

UPGMA dendrogram of Vicia species based on frequencies of seed soluble proteins

UPGMA dendrogram of Vicia species based on frequencies of seed non-soluble proteins
The collective data of both soluble and non-soluble seed proteins were analyzed using UPGMA (Table 4 and Fig. 5); the relationship between the studied five species was as follows: V. ervilia and V. narbonensis were the most related species (69.0%), then V. narbonensis and V. macrocarpa (67.2%), V. macrocarpa and V. sativa (60.7%), and V. faba at a peripheral position.

UPGMA dendrogram of Vicia species based on frequencies of collective date of seed soluble proteins and non-soluble proteins
Discussion
The karyotypes of several species have been established based on chromosome size and centromeric index in addition to the traditional process for karyotyping by adding a dye to metaphase chromosomes. Different dyes that affect different areas of the chromosomes are used for a range of identification purposes. One common dye used is Giemsa; this dye is effective because it markedly stains the bands on a chromosome; each chromosome can then be identified by its banding patterns (Cremonini 1992; Galasso et al. 1994; Cremonini et al. 1998; Fuchs et al. 1998); however, this approach is limited by the similar morphology of chromosomes in many species.
Chromosome features and their count have been recorded in cytological characterization of germplasm (Sharma and Sharma 2013). The genus Vicia become an interesting model for studying a plant genome and karyotype evolution due to the variation in basic chromosome number between Vicia species 2n = 10, 12, or 14 (Maxted 1995). El-Bok et al. (2014) mentioned that the chromosome numbers varied between Vicia species and subspecies such as Vicia cordata had 2n = 10, Vicia angustifolia had 2n = 12, Vicia narbonensis, and Vicia monantha ssp. calcarata and ssp. cinerea presented 2n = 14. Both V. sativa ssp. amphicarpa accessions with aerial and underground pods showed 2n = 14 and were first reported. Chromosome numbers of V. sativa ssp. sativa were verified and revised as 2n = 10, 12. Also, Gaffarzadeh-Namazi et al. (2008) found that Vicia species from Iran were different in chromosome number, karyotype formula, and karyotype characteristics such as Vicia villosa (2n = 2x = 14), Vicia hyrcanica (2n = 2x = 12), V. sativa subsp. sativa (2n = 2x = 12), and V. sativa subsp. nigra (2n = 2x = 12).
In our results, V. macrocarpa ((Moris) Betol.) had six pairs of chromosomes (2n = 12): three metacentric, two submetacentric, and one subtelocentric chromosome; this nearly agrees with Raina et al. (2001) who worked on Vicia macrocarpa ((Moris) Arcang).
V. sativa L. (ssp. Sativa convar. var. sativa) had six pairs of chromosomes (2n = 12): three metacentric and three subtelocentric chromosomes; this results are agreement with the results of Davis and Plitmann (1970), Raina and Rees (1983), and Maxted et al. (1991) who found that the chromosome numbers for V. sativa (subsp. sativa) and V. sativa (subsp. macrocarpa) were 2n = 12, while there are different chromosome number reported in V. sativa (subsp. Incise var. incise) (2n = 14) in the study of Çiler and Feruzan (1999). But it was determined to be 2n = 10 for V. sativa subsp. incisa var. cordata as reported by Raina and Rees (1983) and Kamari et al. (1994). V. sativa subsp. nigra was found to have 2n = 12, 14 (Davis and Plitmann 1970; Tutin 1968), while V. sativa subsp. Amphicarpa had 2n = 14 (Tutin 1968).
V. narbonensis L. (var. narbonensis) had seven pairs of chromosomes (2n = 14): one metacentric, one subtelocentric, and five submetacentric chromosomes; this is in agreement with the result of Navrátilová et al. (2003) who worked on Vicia narbonensis (L.) IFYN574, and with the results of Raina et al. (2001) who worked on V. narbonensis (ssp narbonensis).
In our results, V. ervilia L. (Willd) had seven pairs of chromosomes (2n = 14): three metacentric, one submetacentric, and three subtelocentric chromosomes, while V. faba (var. sakha 3) had six pairs of chromosomes (2n = 12): one metacentric and five subtelocentric chromosomes which disagree with Hizume et al. (1980) in chromosome type; they studied the C-banding patterns on V. faba using Giemsa stain and found that the number of chromosomes 2n = 12 (one metacentric, three subtelocentric, and two telocentric); the metacentric chromosome associated with secondary constriction on short arm.
SDS-PAGE considered a genetic markers in analyses of genetic distances between species to determine the taxonomic relationship (Tamkoc and Arslan 2011).
In our study, the five Vicia species were examined for protein profile levels and the data was subjected to unweighted pair-group method with arithmetical average (UPGMA) to find the phylogenetic relationships among the species. First, the relationship between the studied five Vicia species depending on seed soluble protein similarity level was as the follows: V. ervilia and V. narbonensis are the most related species (77.8%), then comes V. macrocarpa which is most related to V. narbonensis (64.5%), and V. sativa which is most related to V. macrocarpa (60.0%), and peripheral position comes V. faba which has almost the same relationship with the other studied species (Table 2 and Fig. 3). Second, the relationship between the studied five species depending on seed non-soluble protein similarity level was as the follows: V. narbonensis and V. macrocarpa are the most related species (70.0%), then comes V. sativa, V. ervilia, and V. faba (Table 3 and Fig. 4), while the relationship between the studied five species depending on the collective data of both soluble and non-soluble proteins using UPGMA was as the follows: V. ervilia and V. narbonensis were the most related species (69.0%), then comes V. narbonensis and V. macrocarpa (67.2%), then comes V. macrocarpa and V. sativa (60.7%), and V. faba at a peripheral position (Table 4 and Fig. 5).
There are a few studies made to determine the genetic diversity between faba bean and its related Vicia species (Haider et al. 2001). In that study, 13 taxa representing 6 Vicia species (V. sativa, V. villosa, V. monantha, V. narbonensis, and V. cinerea, in addition to V. faba) were collected from the north-west coastal region of Egypt. SDS-PAGE indicated clear differences between different Vicia species as well as within the taxa of the same species. The results indicated that V. monantha and V. villosa are most closely related to V. cinerea, V. faba, and V. narbonensis, which are completely distant. The marked differences between V. faba and other wild species indicated that none of the latter can be considered as the wild progenitor of V. faba, while Kahraman et al. (2016) implicated that protein banding patterns for investigating 22 bean genotypes provided a clear classification by view of selection criteria. Similarity dendrogram presented two main groups that showed ranges nearly 20–75% and 50–90%, respectively.
Conclusion
The phylogenetic relationships among the studied species depending on the seed storage proteins profile (soluble and non-soluble proteins) using SDS-PAGE and karyotype characteristic were as follows: V. ervilia was closely related to V. narbonensis, and V. narbonensis was related to V. macrocarpa, but the degree of relation between V. narbonensis and V. macrocarpa was less than the relation between V. narbonensis and V. ervilia which were the most related species, while V. sativa was closely related to V. macrocarpa, but V. faba was distant from all other studied species. This study helps the breeder to perform the breeding program without consumption of a long duration for doing several crossing over for doing a hybridization between different species.
Availability of data and materials
Not applicable.
Abbreviations
- S:
-
Short arm
- L:
-
Long arm
- S:L:
-
Arm ratio
- Sm:
-
Submetacentric
- St:
-
Subtelocentric
- M:
-
Metacentric
- RL:
-
Relative length
- UPGMA:
-
Unweighted pair-group method with arithmetical average
- SDS-PAGE:
-
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis
- WSP:
-
Water-soluble protein
- WNSP:
-
Water non-soluble protein
References
Bebeli PJ, Kaltsikes PJ (1985) Karyotyric analysis of two durum wheat varieties. Can J Genet Cytol 28:42–62
Casperson T, Zech L, Johnsson C (1970) Differential banding of alkylating fluorochromes in human chromosomes. Exp Cell Res 60:315–319
Çiler M, Feruzan D (1999) Karyological studies on Vicia sativa L. subsp. incisa (Bieb.) arc. var. incise. Tr J of Botany 23:63–67
Cremonini R (1992) The chromosomes of Vicia faba: banding patterns and in situ hybridizations. Biol Zent Bl 111:188–203
Cremonini R, Miotto D, Ngu MA, Tota D, Pignone D et al (1998) Cytology of Vicia species. 5. Nuclear chromatin structure, karyomorphological analysis and DNA content in newly discovered relatives of Vicia faba L.: Vicia kalakhensis Khattab, Maxted et Bisby and Vicia eristalioides Maxted. Cytologia 63:371–379 https://doi.org/10.1508/cytologia.63.371
Davis PH, Plitmann U (1970) Vicia L. flora of Turkey and the East Aegean Islands, Davis , P.H., ed. Edinburgh University Press,Edinburgh 3: 274-325.
El-Bok S, Khelil AZ, Brahim TB, Ouji A, Hassen H, Lamine O, Jabri C, Douggari R, El-Gazzah M (2014) Chromosome number and karyotype analysis of some taxa of Vicia genus (Fabaceae): revision and description. Int J Agric Biol 16:1067–1074
Emre I, Turgut-Balık D, Genç H, Şahin A (2010) Total seed storage protein patterns of some Lathyrus species growing in Turkey using SDS-PAGE. Pak J Bot 42(5):3157–3163
Fominaya A, Vega C, Ferrer E (1988) C-banding and nucleolar activity of tetraploid Avena species. Genome 30:336–638 https://doi.org/10.1139/g88-107
Fuchs J, Strehl S, Brandes A, Schweizer D, Schubert I (1998) Molecular cytogenetic characterization of the Vicia faba genome, heterochromatin differentiation, replication patterns and sequence localization. Chromosom Res 6:219–230. https://doi.org/10.1023/A:1009215802737
Gaffarzadeh-Namazi L, Badrzadeh M, Asghari-Zakaria R (2008) Karyotype of several Vicia species from Iran. Asian J Plant Sci 7(4):417–420. https://doi.org/10.3923/ajps.2008.417.420
Galasso I, Piergiovanni AR, Cremonini R, Perrino P, Pignone D (1994) Cytology of Vicia species. 3. Characterization of the chromosomal chromatin of some species of the section Cracca. Cytobios. 77:175–182
Gepts P, Charles EB, Randy C, Stalker H, Norman F et al (2005) Legumes as a model plant family. Genomics for food and feed report of the cross-legume advances through genomics conference. Plant Physiol 137:1228–1239
Haider AS, Bahieldin A, Hassanin R, Mahmoud N, Madkour M (2001) Molecular characterization of some species of genus Vicia. Arab Journal of Biotechnology 4:197–206
Hanelt P, Mettin D (1989) Biosystematics of the genus Vicia L. Leguminosae. Annu Rev Ecol Syst 20:199–223. https://doi.org/10.1146/annurev.es.20.110189.001215
Hizume M, Tanaka A, Yonezawa Y, Tanaka R (1980) A technique for C-banding in Vicia faba chromosomes. Japan J Genetics 55(4):301–305
Howell W, Denton T, Diamond J (1975) Differential staining of the satellite regions of human acrocentric chromosomes. Experientia 31:260–262
Jellen EN, Ladizinsky G (2000) Giemsa C-banding in Avena insularis Ladizinsky. Genet Resour Crop Evol 47:227–230. https://doi.org/10.1023/A:1008769105071
Jellen EN, Phillips R, Rines H (1993) C-Banded karyotype and polymorphisms in hexaploid oat accessions Avena spp. Using Wright stain. Genome 36:1129–1137
Kahraman A, Uysal T, Bozkurt M, Şimşek Sezer EN, Ceyhan E, Ozkan Z (2016) Classification of bean genotypes by protein profiles. Selcuk J Agr Food Sci 30(1):29–33
Kamari G, Felber F, Garbari F (1994) Mediterranean chromosome number reports 4. Flora Mediterranea 4:233–301
Kupicha FK (1976) The infrageneric structure of Vicia L. Notes from the Royal Botanic Garden, Edinburgh 34:287–326
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227:680–685 https://doi.org/10.1038/227680a
Lavia GI, Ortiz AM, Fernandez A (2009) Karyotypic studies in wild germplasm of Arachis (Leguminosae). Genet Resour Crop Evol 56:755–764
Maxted N (1993) A phenetic investigation of Vicia (Leguminosae, Vicieae). Bot J Linn Soc 111:155–182 https://doi.org/10.1017/S0960428600001736
Maxted N (1995) An ecogeographical study of Vicia subgenus Vicia. Systematic and ecogeographic studies on crop genepools. 8. International Plant Genetic Resources Institute, Rome.
Maxted N, Callimassia MA, Bennett MD (1991) Cytotaxonomic studies of eastern mediterranean Vicia species (Leguminosae). Pl Syst Evol 177:221–234
Murti RH, Kim HY, Yeoung YR (2012) Morphological and anatomical characters of ploidy mutants of strawberry. Int J Agric Biol 14:204–210
Navrátilová A, Neumann P, Macas J (2003) Karyotype analysis of four Vicia species using in situ hybridization with repetitive sequences. Ann Bot 91:921–926
Raina SN, Rees H (1983) DNA variation between and within chromosome complements of Vicia species. Heredity 51:335–346
Raina SN, Mukai Y, Kawaguchi K, Goel S, Jain A (2001) Physical mapping of 18S-5.8S-26S and 5S ribosomal RNA gene families in three important vetches (Vicia species) and their allied taxa constituting three species complexes. Theor Appl Genet 103:839–845
Sharma G, Sharma N (2013) Cytology as an important tool for solving evolutionary problems in Angiosperms. Proc Natl Acad Sci, India, Sect B Biol Sci 84:5–12
Studier FW (1973) Analysis of bacteriophage T1 early RNAs and proteins of slab gels. J Mol Biol 79:237–248
Tamkoc A, Arslan E (2011) Inter and intra-specific variation in SDS-page of seed proteins of three Poa L. (poaceae) species. Pak J Bot 43(2):1105–1110
Tutin TG (1968) Flora Europaea 2:129–136
Acknowledgements
I would like to thank the Genetics and Cytology Department, National Research Centre, because this research work was done in the Genetic and Cytology Laboratory.
Funding
This research was not funded by any funding body.
Author information
Authors and Affiliations
Contributions
Authors SAO and HBA designed the study and managed the laboratory experiments and analyzed all data. Authors ZME and SEE managed the literature searches and wrote this manuscript. All authors approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Osman, S.A., Ali, H.B., El-Ashry, Z.M. et al. Karyotype variation and biochemical analysis of five Vicia species. Bull Natl Res Cent 44, 91 (2020). https://doi.org/10.1186/s42269-020-00347-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s42269-020-00347-3