
Abstract
Background
Only a few studies have addressed the relationship between toll-like receptors 2 and 4 (TLR2 and TLR4) and the production of local and systemic cytokines in response to physical exercise, and they have produced conflicting results. We aimed to determine whether acute and chronic exercise outcomes are associated with changes in TLR2 and TLR4 expression and signaling and if so, the mechanisms that connect them.
Methods
PubMed database were consulted. This systematic review selected 39 articles, 26 involving humans and 13 based on rodents.
Results
In acute resistance exercise studies, 75% reported a decrease in TLR4 or TLR2 expression and 25% did not find differences. For chronic resistance exercise studies, 67% reported a reduction of expression and 33% did not find differences. Studies of both types reported reductions in pro-inflammatory cytokines. In acute aerobic exercise studies, 40% revealed a decline in the expression of the receptors, 7% reported no significant difference, 40% showed an increase, and 13% did not evaluate their expression. Fifty-eight percent of studies of chronic aerobic exercise revealed a reduction in expression, 17% did not find a difference, and 25% reported increases; they also suggested that the expression of the receptors might be correlated with that of inflammatory cytokines. In studies on combined exercise, 50% reported a decline in receptors expression and 50% did not find a difference.
Conclusions
The majority of the articles (54%) link different types of exercise to a decline in TLR4 and TLR2 expression. However, aerobic exercise may induce inflammations through its influence on these receptor pathways. Higher levels of inflammation were seen in acute sessions (40%) than regular sessions (25%).
Similar content being viewed by others

Key Points
-
It is known that regular exercise acts as an anti-inflammatory agent by down-regulating TLR4 in immune cells. Paradoxically, acute, extended, or intense exercise can be harmful to the immune system.
-
The molecular mechanisms by which various types of physical exercise modulate the TLR2 and TLR4 pathways are still not fully understood.
-
Physical exercise reduced the expression of TLR2 and TLR4. However, aerobic exercise is potentially inflammatory when compared with resistance exercise.
Background
The connections between lifestyle factors and health have been the subject of intense research, partly motivated by alarming changes in the health landscape of industrialized societies. One clear trend is that moderate exercise benefits health in many ways, while extremes of too little or excessive exercise have been linked to chronic diseases. Many of these have an immune component—individuals with very sedentary lifestyles often fall prey to low-grade chronic inflammations [1,2,3,4]. Over the long term, this condition can lead to type 2 diabetes, cardiovascular diseases, particular types of cancer, chronic respiratory diseases, and other serious health problems. Physicians have called this constellation a worldwide epidemic [5]. The immune system can also be disrupted by excessive exercise. While progress has been made, there remain many gaps in our understanding of the mechanisms that connect the types and amounts of a person’s activity to immune responses and disease.
The prevalence of inflammations suggests a logical point of departure for such studies. Inflammation involves complex interactions at the molecular and cellular levels that can arise in any vascular tissue as a result of traumatic, infectious, post-ischemic, toxic, or autoimmune injuries [6]. Toll-like receptors play a role in many of these conditions; they are known to make significant contributions to obesity [7, 8], type 2 diabetes [9], non-alcoholic steatosis [10], cardiovascular disease [11, 12], cerebral ischemia [13, 14], Alzheimer’s disease [15], rheumatoid arthritis [16], and other diseases. This review examined recent work that suggests they also help modulate the effects of different levels of physical activity on states of health and disease.
TLRs are type I transmembrane proteins involved in both innate and adaptive immune system responses [17, 18]. These receptors mediate the recognition of pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (DAMPs)—specific molecules released by damaged or necrotic cells [18, 19]. The immune activities of TLRs are generally modulated through signaling via the NF-kB pathway. Responses begin with the stimulation of the receptor by an external signal. This alters the cytoplasmic regions of TLRs, which contain Toll/interleukin-1 (IL-1) receptor (TIR) domains. Stimulation causes these domains to recruit adaptor proteins in a process that ultimately activates the nuclear transcription factor NF-kB [17]. This releases NF-kB for transport to the cell nucleus, where it triggers the transcription of cytokines including IL-1β, IL-6, and IL-8 interleukins; TNF-α [20,21,22]; and other elements [23] that play key roles in the immune system responses. Alongside cytokines, NF-kB induces the expression of growth factors and other molecules involved in stress response, cell proliferation, and cell cycle progression [24,25,26].
TLRs are expressed in the immune cells including macrophages, dendritic cells (DCs), B cells, and specific types of T cells. They are also present in non-immune cells such as fibroblasts and epithelial cells [27] and in the tissues of the ovary, prostate, placenta, testicles, lungs, liver, and skeletal muscle [28].
The toll-like receptors TLR2 and TLR4 have received particular attention due to their ability to identify molecular patterns exhibited by several invasive pathogens [18]. They also seem to play an important role in the anti-inflammatory effects observed in physically active individuals [29]. Regular exercise has been determined to have anti-inflammatory effects [2, 29,30,31,32,33,34] by downregulating TLR4 in the immune cells. A bit paradoxically, at the other end of the activity spectrum, acute, extended, or intense exercise can have a negative impact on the immune system [35,36,37,38,39,40,41,42]. But the molecular mechanisms by which exercise modulates the TLR2 and TLR4 pathways are still not fully understood.
One plausible link comes from the demonstration that TLR2 and TLR4 are activated by the extracellular non-esterified fatty acids (NEFAs). Concentrations of extracellular NEFAs undergo transient increases during aerobic exercise (AE). If levels are chronically elevated, however, TLRs may induce the production of pro-inflammatory cytokines in macrophages, adipocytes, liver, and skeletal muscle cells. This suggests that the receptors may participate in the development of insulin resistance [43]. Yet, they also have protective effects against insulin resistance, which may be explained by the down-regulation of TLR expression that occurs during physical exercise [43].
Here, this review investigated the existing literature on the inflammatory and anti-inflammatory effects of different types of physical exercise with a focus on systematically collecting connections to TLR2 and TLR4 modulation and signaling. To accomplish this, the results were divided into single sessions of acute exercise and chronic exercise, based on periodicity. Additionally, this review identified key biomarkers and analyzed the combined TLR2 and TLR4 responses to markers involved in the process of inflammation process, including anti- and pro-inflammatory cytokines, adaptor proteins, and the transcription factor NF-kB.
Inflammatory Effects of Physical Exercise
Analyzing the modulation of inflammation patterns permits insights into specific underlying physiological mechanisms. As a controllable model of stress, physical exercise is a good tool to analyze inflammatory responses [44].
Physical exercise permits the control of variables related to activity such as volume, intensity, frequency, and duration. These factors have led to its adoption as a good strategy to study alterations that occur due to inflammations caused by stress and their implications for health [45,46,47]. Local and systemic cytokine production in response to physical exercise resembles the cytokine response to infections, trauma, and sepsis [44, 45, 48]. There is evidence that very strenuous physical exercise can cause substantial tissue damage and initiate an inflammatory reaction and excessive immunosuppression, in a way that highly resembles features observed in clinical sepsis [49]. However, trauma, infection, and septic complications can produce an uncontrollable inflammatory response with long-term detrimental or fatal consequences. In physical exercise, although the inflammatory cascade has obvious similarities, the response appears to be limited [44].
Usually, the process of inflammation has an overall positive effect on the organism. Short-term, acute inflammation allows the body to survive progressive tissue destruction by promoting healing [50, 51]. On the other hand, if destruction and repair are not properly coordinated, inflammation may lead to persistent tissue damage. The mechanisms by which acute inflammation starts and develops are well understood, but little is known about the causes of chronic inflammation and its association with molecular and cellular pathways [51].
A comparison can also be made between chronic inflammation and strenuous physical exercise in which pro-inflammatory pathways seem to be activated [38, 41, 52]. In response to heavy exercise, inflammation stimulates tissue monocyte production, and platelet hyperactivity promotes fibrinogen biosynthesis and induces the formation of the microparticle and the accumulation of erythrocytes to trigger a prothrombotic state. In fact, vigorous aerobic exercise may be atherogenic and atherothrombotic due to the overproduction of mitochondrial-free radicals in the skeletal and myocardial muscle. On the other hand, both moderate AE and low-load resistance exercise (RE) may reduce inflammation and improve fibrinolysis. [52].
An elegant study [53] found associations between all causes of mortality and doses of jogging. Light and moderate joggers had a lower mortality than sedentary non-joggers, while there was no significant statistical difference between mortality in strenuous joggers and the sedentary group. In this analysis, high running loads in sports such as marathons, ultramarathons, triathlons, and long high-intensity bike rides can cause negative effects such as acute inflammations; in the long term, these activities may lead to chronic inflammation, irregular fibrosis formation, alterations in the size of the cardiac chambers, and atrial fibrillation [54]. Moreover, long-distance runners may have increased levels of atherosclerosis and coronary disease due to constant training throughout the year [54]. In atherosclerosis, the endothelial permeability is increased by the oxidative damage that promotes the entry of lipoproteins in the subendothelial space, resulting in inflammation [55]. When the lipoproteins are oxidative, they interact with TLR4 in particular and promote cardiovascular disease [56].
According to the American College of Sports Medicine (ACSM) and the American Heart Association [57], the minimum recommendation for physical exercise for adults and seniors aiming to avoid chronic disease is 30 min of moderate aerobic activity per day, five times a week; 20 min per day of intense activity, three times a week; or a combination of moderate and vigorous activity. These guidelines also suggest that high loads of AE may be necessary for some groups to prevent a transition to an estimation that they are overweight or a diagnosis of obesity. However, they also recommend limiting vigorous physical training to 60 min a day, for a weekly total of no more than 5 h, including 1 to 2 days without high-intensity exercise per week [58, 59]. Strenuous AE has been shown to induce an excess of reactive oxygen species (ROS) [60]; can modulate TLR4 signal transduction at many levels [61]; stimulate pro-inflammatory transcription factors such as NF-kB, AP-1, and Nrf2 [62, 63]; and promote inflammation [64].
NADPH oxidase 4 (NOX4), involved in redox signaling in vascular cells, has direct interactions with TLR4 in both for the generation of endogenous and exogenous ROS-mediated by LPS and the activation of NF-kB [65]. In addition, high levels of ROS in the muscles can provoke a hyperactivation of the innate immune system in cells such as macrophages and neutrophils [66], and it leads to the production of several peroxides and aldehydes that are potentially toxic to the cells [67], also affecting T cell polarization and contributing to pro-inflammatory cytokine secretion [68]. It is already known that ROS production and neutrophil counts change in athletes involved in activities such as running, jumping, throwing, combined events (triathlon, heptathlon, and decathlon), swimming, cycling, and soccer, but only high-intensity exercise induces oxidative damage in lymphocytes [69]. In contrast, moderate-intensity AE stimulates the combat of excessive ROS by maintaining redox balance in the muscle [70]. A study [71] of soccer players showed a significant correlation between leukocyte ROS production and creatine kinase (CK) values, considered a qualitative marker for microtrauma skeletal muscle.
In fact, the physiological effects of strenuous AE, for example, participation in triathlons, include a large increase in CK, C-reactive protein (CRP), cortisol, and aldosterone and a decrease in testosterone levels [72]. Moreover, after strenuous exercise, increased levels of LPS may trigger an increase in the production of pro-inflammatory cytokines [73,74,75,76]. Long periods of AE [72] or short acute sessions of strenuous physical exercise [41] can disturb homeostasis and enhance inflammation. Consistent with this, Rodrigues-Miguelez et al. [39] found an increase in TLR4 and pro-inflammatory cytokines such as TNF-α and IL-1β in acute AE sessions; however, the effects were reversed with regular training in reasonable doses.
TNF-α represents a group of peptides that are released into the bloodstream in response to the endotoxin stimulation during infectious processes. TNF-α has a catabolic effect [77] and plays a role in the loss of muscle mass that usually appears in chronic diseases such as rheumatoid arthritis and cancer [78]. TNF-α genesis in low-grade systemic inflammation is thought to occur mainly in the adipose tissue [79,80,81]. Furthermore, systemic inflammation and high concentrations of pro-inflammatory cytokines act on the hypothalamic-pituitary-adrenal axis and can increase serum concentrations of cortisol [82, 83]. Physical exercise and nutrition modulate the cortisol response. Variables such as intensity, lactate accumulation, total volume, and resting period determine the level of cortisol released to stimulate glycogenolysis and gluconeogenesis [84, 85]. Moderate- to high-intensity exercise can cause increases in circulating levels of cortisol. On the other hand, low-intensity exercise (40% VO2max) reduces circulating levels of cortisol [84]. In the study by Lira et al. [76], TLR-4 and NF-kBp65 were increased in animals from both groups (overtraining and resting after overtraining). Additionally, a decrease in the performance and an increase in the production of corticosterone and endotoxin were observed in overtraining groups compared to both control and trained groups, indicating that chronically high levels of plasma cortisol can increase inflammation in the epididymal adipose tissue.
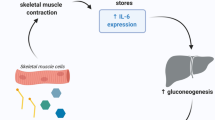
Thereby, an excess of physical (blood cortisol levels) and oxidative stress (intracellular ROS accumulation) can generate temporary immune dysfunction [86]. In contrast, physical exercise at moderate intensities regulates the immune system and reduces oxidative stress [87]. Figure 1 presents a simplified comparison of some mechanisms that can be activated by strenuous physical exercise and by regular exercise performed at moderate intensity.

Signaling involving TLR2 and TLR4 in strenuous and moderate aerobic exercise. Excess physical exercise increases LPS levels and contributes to TLR2, TLR4, and NF-kB upregulation. As a consequence, there is an increase in circulating pro-inflammatory cytokines. Stimuli of exercise stress transmit nerve impulses to the brain, raising the levels of counter-regulatory hormones such as cortisol. Accordingly, high mitochondrial oxidative stress induced by strenuous aerobic exercise causes excessive intracellular ROS formation that also upregulates NF-kB expression, intensifying the acute inflammation state. Under these excessive stress conditions, adaptive immunity can be triggered by the increase in costimulatory molecules in antigen-presenting cells, thus activating T cells. In contrast, the regular physical exercise of moderate intensity reduces LPS, TLR2, TLR4, and NF-kB expression. Under these conditions, NF-kB does not translocate to the cell nucleus. Instead, the anti-inflammatory pathway PI3K/AKT/mTOR is activated, promoting the production of anti-inflammatory cytokines such as IL-10 that inactivate TNF-α. Physical exercise at a moderate intensity also has a compensatory effect against the exacerbated production of reactive oxygen and nitrogen species responsible for the oxidative damage. Elevated production of IGF-1 is observed after exercise. IGF-1 provides an anti-inflammatory effect on the skeletal muscle cells, reducing the expression of the pro-inflammatory cytokines through a decrease of TLR4 expression
Anti-inflammatory Effects of Physical Exercise
It is well known that regular physical exercise has anti-inflammatory effects [8, 29,30,31, 88,89,90,91,92,93]. Therefore, regular physical exercise, as well as a physically active lifestyle, may be useful as a treatment for a range of chronic diseases and conditions characterized by low-grade systemic inflammation [3, 94].
However, the link between physical exercise and TLRs is still a matter of debate. Although the pro-inflammatory effects of TLR2 and TLR4 signaling have been well studied, anti-inflammatory responses due to the activation of these receptors are still not fully understood [95]. For this reason, this article will briefly address a number of molecules that act directly during the processes of adaptation to physical exercise—including hormones, myokines, and chemical molecules such as ROS.
The skeletal muscle can function as an endocrine organ due to its production of growth hormones and cytokines known as myokines, which are induced by an exercise stimulus [96, 97]. One of the best-known exercise-induced adaptations [98, 99] is an increase in circulating levels of insulin-like growth factor 1 (IGF-1). Elevated levels of circulating IGF-1 have been observed after exercise, probably in response to hepatic secretion stimulated by growth hormone (GH) [85].
The first evidence that IGF-1 is a potent modulator of TLR4 (protein expression) in the skeletal muscles was provided by Lee [31]. The author demonstrated that IGF-1 stimulation had anti-inflammatory effects on the skeletal muscle and suppressed TLR4 signaling. Treatment with IGF-1 attenuated the amounts of endogenous IL-6 and TNF-α, indicating that IGF-1 had an anti-inflammatory effect on the skeletal muscle cells by reducing the expression of pro-inflammatory cytokines under baseline conditions through a down-regulation of the expression of TLR4. This led to a hypothesis that cells with low levels of TLR4 are less responsive to ligands that stimulate endogenous inflammation, such as the heat shock protein, and thus contribute to a lower basal response of pro-inflammatory cytokines [31]. In addition to the anti-inflammatory effects of IGF-1, regular AE promotes the remodeling of mitochondrial networks with significant improvements in both the quality and quantity of the mitochondria [100]. This results in positive changes in the respiratory capacity and oxygen extraction of trained subjects [100, 101].
Likewise, there is an increase in angiogenesis, the formation of new capillaries from pre-existing ones. High levels of VEGF—resulting from endurance training—offer favorable conditions for an increase in the density of the muscle capillaries [100]. Furthermore, a moderate level of AE reduces pro-atherogenic cytokines such as TNF-α and IFN-γ and simultaneously increases atheroprotective cytokines such as IL-4, IL-10, and TGF-β [102].
The anti-inflammatory effects of regular exercise might be mediated by a reduction of visceral fat mass followed by a decline in the release of adipocytokines, as well by the anti-inflammatory environment induced by exercise [103]. This environment consists of three variables: cortisol and adrenaline release from suprarenal glands, an increase in the production and release of IL-6 and other myokines from skeletal muscle, and a decrease in amounts of TLR (cell surface protein and mRNA expression) - in monocytes and macrophages, and as a consequence, the inhibition of the release of pro-inflammatory cytokines [103].
In fact, there is evidence that exercise is responsible for reducing the expression of these receptors at both mRNA expression and protein levels [2, 29, 30, 32, 93]. In diet-induced obesity rats (DIO), both acute aerobic exercise (AAE) and chronic aerobic exercise (CAE) led to a significant suppression of the TLR4 signaling pathway in liver, muscle, and adipose tissue, reduced LPS in serum, and improved insulin signaling [9]. However, the anti-inflammatory responses induced by TLR4 activation have not been characterized as clearly. In contrast to TLR4 pro-inflammatory signaling at the cell surface, TLR4 signaling from endosomal compartments induces the secretion of the anti-inflammatory cytokine IL-10 [95].
During physical exercise, a transient increase in IL-6 in circulation appears to be responsible for a further increase in the levels of circulating anti-inflammatory cytokines such as IL-10 and IL-1ra [104,105,106]; this also stimulates the release of cortisol from the adrenal glands [106]. Increases in IL-6 levels during exercise are transient and return to resting levels usually within 1 h after exercise [107]. This phenomenon may occur because IL-6 production is modulated by the glycogen content in muscles [108], which function as an energy sensor [97].
The anti-inflammatory effects of TLR2 and TLR4 during exercise are mediated by the PI3K/AKT/mTOR pathway after an activation of adaptor proteins, leading to the production of IL-10 (Fig. 1) [95], an anti-inflammatory cytokine produced by Th1 cells, monocytes, and macrophages that is present in higher concentrations after physical exercise and acts as a potent inhibitor of pro-inflammatory cytokines [109, 106].
IL-10/IL-10R signaling is mediated by the activation of the JAK/STAT pathway through the phosphorylation of the Tyk2/JAK1 tyrosine, which results in the activation of STAT3 [110]. This mechanism is independent of the toll-like pathway. An analysis of the IL-10/TNF-α ratio is often used as an indicator of inflammatory conditions [32, 111]. This is evidence that IL-10 acts as a natural antagonist of TNF-α and is able to inhibit NF-κβ signaling [110, 112], as shown in Fig. 1.
Methods
This review consulted the PubMed database in a search involving seven keywords: “exercise,” “training,” “physical activity,” “TLR,” “TLR2,” “TLR4,” and “toll-like,” To cross-reference the words, 12 groups were created to link terms associated with exercise (“exercise,” “training,” “physical activity”) to toll-like terms (“TLR,” “TLR2,” “TLR4,” and “toll-like”), building groups formed from two individual keywords linked by the Boolean operator “AND.” This produced groups organized as follows: group 1: “exercise” and “TLR”; group 2: “exercise” and “TLR2”; group 3: “exercise” and “TLR4”; group 4: “exercise” and “toll-like”; group 5: “training” and “TLR”; group 6: “training” and “TLR2”; group 7: “training” and “TLR4”; group 8: “training” and “toll-like”; group 9: “physical activity” and “TLR”; group 10: “physical activity” and “TLR2”; group 11: “physical activity” and “TLR4”; and group 12: “physical activity” and “toll-like.”
Only studies carried out directly in animal models (human, rat, and mouse) were included. For scientific substantiation, 119 scientific articles were also consulted in addition to the 39 studies which met the criteria of eligibility for this review.
Criteria which excluded articles from this review, described in Table 1, fell into categories as follows: non-English articles; literature reviews; articles that did not cover Toll-like receptors (TLRs); articles studying TLRs other than TLR2 and TLR4; articles without exercise protocols; experimental articles that did not use humans, mice, or rats; and finally, articles that involved diet, supplementation, or drugs. To do so, codes to link the eligibility criteria of all of the items found in the search were created.
Initially, 1385 articles were found. After an update, the search ended up with 1548 articles from the PubMed database. The updated search was carried out in October 2015. The search group distribution can be seen in Table 2. Figure 2 shows a flowchart of the article selection process, as well as how the articles were linked to the search theme. The total number of articles found and the distribution of the excluded articles are also carefully detailed.

Literature search flowchart
Results and Discussion
To investigate the roles of TLR2 and TLR4 behavior in the inflammatory and anti-inflammatory effects of exercise, the results were distributed according to the type of exercise (resistance, aerobic, and combined) and frequency of training (acute or chronic), taking the exclusion criteria into account.
Considering the total of 39 studies that met the eligibility requirements for this review, 28 articles were based on the samples from a disease-free setting and 11 samples related to a disease. Three articles studied the effects of exercise and TLR2 and TLR4 on obesity [8, 113, 114], one on pre-diabetes [115], one on low back pain [116], two on cerebral ischemia [13, 14], one on pulmonary inflammation [117], one on Alzheimer’s disease [15], one on chronic fatigue syndrome [36], and one on multiple sclerosis and fibromyalgia [118].
As shown in Table 3, 21 of the 39 eligible articles (54%) showed a reduction in TLR4 and/or TLR2 at the levels of both cell surface protein and mRNA expression, 7 (18%) did not show statistically significant differences, 2 articles (5%) did not test TLR4 and/or TLR2 expression but were included in this review for the evaluation of downstream targets of the receptor pathways, and 9 articles (23%) reported an increase in TLR2 and/or TLR4 (gene expression or protein levels) after AE sessions.
The results were also analyzed by subgroups and divided according to the type and frequency of training (Table 3 and Fig. 3). For chronic resistance exercise (CRE), four articles (67%) reported a reduction of TLR4 and/or TLR2 expression and two (33%) did not show any significant change. For acute resistance exercise (ARE), three articles (75%) revealed a decrease in the expression of these receptors and one study (25%) failed to find a significant difference. For CAE, seven articles (58%) reported a reduction in TLR4 and/or TLR2 expression, two studies (17%) did not find a significant difference, and three articles (25%) found an increase in the expression of TLR4 and/or TLR2. For AAE, six experiments (40%) showed a decrease, one (7%) did not show any difference, six (40%) reported an increase, and two articles (13%) tested neither TLR2 nor TLR4 expression. Regarding combined exercise (CE), one study (50%) reported a reduction in the expression of the receptors and one study (50%) revealed no significant difference.

Effects of exercise on TLR2 and TLR4. For chronic resistance exercise, 67% of studies reported a reduction of TLR4 and/or TLR2 expression. For acute resistance exercise, 75% of studies revealed a decrease in the expression of these receptors. For chronic aerobic exercise, 58% of studies reported a reduction in TLR4 and/or TLR2 expression and 25% found an increase in the expression of TLR4 and/or TLR2. For acute aerobic exercise, 40% of studies showed a decrease and 40% reported an increase. Regarding combined exercise, 50% of studies reported a reduction in the expression of the receptors
Resistance Exercise and Inflammation
Six articles that studied TLR4 and/or TLR2 behavior with CRE were identified (Table 4). Two studies found a reduction of TLR4 and TLR2 in terms of protein expression [92, 119], two revealed a decrease in mRNA expression [116, 120], and two did not find a statistically significant difference [8, 121]. Three articles [29, 88, 122] showed reductions in the protein and gene expression of TLR4 after an ARE session, and one article [123] did not show a significant difference in TLR2 (protein levels), as shown in Table 5. This systematic review showed that resistance exercise (RE), whether acute or chronic, could act as a regulator of inflammation. In this subset of the literature, we observed no increases in the expression of TLR4 and/or TLR2 or pro-inflammatory cytokines after exercise.
Some studies [30, 124, 125] corroborate the results of this review and suggest that CRE may have anti-inflammatory effects. In contrast, ARE may stimulate changes in metabolic demand and promote inflammatory responses, whose occurrences is fundamentally determined by the exercise protocol [126, 127]. In this analysis, ARE transiently increases circulating levels of CK and pro-inflammatory cytokines, e.g., TNF [126] and IL1β [127]. Some studies that were not eligible for this review [128, 129] have shown that ARE induced microdamage in the skeletal muscle, along with an increase in inflammation markers such as IL-6, IL-8, monocyte chemotactic protein-1 (MCP-1), CK, and CRP when performed at high levels of stress.
The ten eligible studies of CRE and ARE [8, 29, 88, 92, 116, 119,120,121,122,123], tested different frequencies, intensities, and durations of exercise, none of these methods, however, produced changes in levels of TLR2 and/or TLR4. In these studies, intensities ranged from 60 to 80% of 1 RM with a gradual increase [119], or 6–14 RM [121]. In one study [92], the training volume followed a criterion of progression. Another study [120] used 80, 90, and 95% of maximal volitional strength capacity (MVSC), with low training volume as the criterion.
Regarding the inflammation markers that were subjected to the analysis here, neither acute nor chronic RE increased levels of pro-inflammatory cytokines such as TNF-α or IL-6. Eight studies tested TNF-α, and the majority [8, 88, 92, 116, 120, 122] found a significant decline of this cytokine. Two studies [29, 119] found no difference in this marker. Four studies analyzed levels of IL-6 after RE. Two studies [8, 116] found a drop in levels, but no significant difference appeared in the studies by Zanchi et al. [120] and McFarlin et al. [29].
The results showed that the RE protocols for both chronic and acute training adopted by the authors did not generate a pro-inflammatory response. Instead, three studies analyzed by this review [92, 119, 120] established an inverse relationship between the TLR2 and TLR4 receptors and IL-10. In the five studies that investigated IL-10 with RE, four [8, 92, 119, 120] found an increase in this marker and one study found no significant difference [121]. It is known that IL-10 levels are higher after chronic exercise, and this anti-inflammatory cytokine acts as a natural TNF-α antagonist [106, 109].
Aerobic Exercise and Inflammation
A total of 12 articles verified that TLR4 and TLR2 undergo changes in response to CAE (Table 6). Four studies verified a significant decrease in TLR4 and/or TLR2 [13, 15, 76, 115] in terms of protein levels, two studies [117, 130] showed reductions in mRNA expression, and one indicated decreases at both the gene and protein level [14]. Two studies [74, 114] revealed an increase in TLR4 and/or TLR2 (gene and protein), one study reported increased mRNA expression [131], and two studies [113, 132] did not find any significant difference in TLR4 expression.
In 15 studies, a relationship between AAE and TLR2 and/or TLR4 was identified (Table 7). Three studies [133,134,135] found a significant reduction of TLR4 and/or TLR2 (protein levels), and two revealed a decrease in mRNA expression [136, 137]. Four studies [35, 39, 40, 42] found an increase in the protein levels of these receptors, and two studies [36, 37] increased mRNA expression. One study did not find a significant difference [138], and one study reported a significant decline in TLR4 (mRNA expression) in multiple sclerosis but found no difference in cases of fibromyalgia [118]. Two studies [139, 140] did not analyze TLR2 or TLR4 expression.
As demonstrated by the results from the analysis of TLR2 and TLR4 behavior, this review showed that in 23% of all of the articles that were analyzed, AE was associated with increases in inflammation. These results differ from previous studies that tested the expression of these receptors in RE. Ten months of CAE was more effective than strength and flexibility exercises in reducing inflammatory markers such as CRP, IL-6, and IL-18 in the elderly [141].
Most studies found that CAE reduced the levels of TLR2 and/or TLR4 [13,14,15, 76, 115, 117, 130]. However, the major immunological benefits came with exercise performed at a moderate intensity [13,14,15, 76, 117, 130, 132]. On the other hand, Zheng et al. [131] observed an increase in TLR2 (gene expression) and inflammatory cytokines such as TNF-α and IL-6 in the regular moderate intensity exercise group (badminton), with or without stimulation from microbial antigens. However, cytokine levels were suppressed after non-microbial antigen stimulation. The authors attributed this result to possible improvements in the body’s resistance to invasion by pathogens in response to regular exercise, indicating that an increase of these receptors does not necessarily indicate a negative impact on health, though further research is still needed to address this possibility.
The chronic low-grade inflammatory profile (CLIP) is a common feature of the normal aging process, and it is also involved in the pathogenesis of several age-related diseases [142]. CLIP has already been recognized as a factor that plays a causative role in the development of sarcopenia. TNF-α and IL-6 are the most commonly reported inflammatory parameters in these studies [143]. Additionally, human aging is associated with metabolic endotoxemia and high levels of signaling of the RST4-NFkB-MAPK pathway in the muscle. These factors may play a role in the types of insulin resistance mediated by aging and muscle loss [74]. In this analysis, Ghosh et al. [74] observed an increase in TLR4 (mRNA and protein levels) in older people but not in younger participants. The study examined people engaged in a progressive regime of the intensity and volume of training, ranging from 65 to 80% of VO2max, and an increase in the duration and number of sessions. Their results provide evidence that higher LPS flow in the elderly can play a critical role in age-related sarcopenia and insulin resistance.
Studies that did not fit our criteria [54, 58, 144, 59, 145] suggested that CAE performed under conditions of high stress leads to inflammation in participants of all ages. They observed that long-distance runners might have increased levels of atherosclerosis and coronary heart diseases due to a training regime that went uninterrupted over many years [54]. Additionally, endotoxemia was found in 68% of athletes after a long-distance triathlon, and LPS levels were associated with higher levels of CRP [75]. A recent study showed that 24 h of continuous ultramarathon activity resulted in a higher level of LPS and increased levels of circulating pro-inflammatory cytokines [146]. In fact, prolonged intense physical exercise leads to elevated concentrations of counter-regulatory hormones in plasma such as cortisol and catecholamines related to low immunity [147]. In addition, high levels of muscle oxidative stress lead to an excessive production of ROS and inflammation [60]. In contrast, regular moderate physical exercise can compensate for oxidative stress [148].
Short acute sessions of physical exercise may disturb homeostasis and increase inflammation [41], as verified by some of the articles reviewed here [35, 37, 39, 40, 42]. With the exception of the study by Light et al. [36], which tested an AAE protocol at moderate intensity and in samples obtained from individuals with disease, studies based on different strenuous exercise protocols consistently led to increases in TLR4, TLR2, and pro-inflammatory cytokines [35, 37, 39, 40, 42]. Rodrigues-Migueles et al. [39] found an increase in TLR4 (protein) and pro-inflammatory cytokines in AAE sessions. However, all of these effects were extinguished by CAE through a weekly exercise protocol of increasing intensity and duration.
In studies which reported increases in TLR2, TLR4, and pro-inflammatory cytokines after acute sessions, IL-10 was tested in only three experiments, all of which revealed a significant increase in the expression of this cytokine [36, 37, 118]. This was probably caused by a transient increase in IL-6 which then led to a subsequent increase in levels of IL-10 [104, 106]. However, other studies [133,134,135] indicated that AAE had beneficial effects, as observed through a decline in terms of protein levels of TLR2 and/or TLR4 and at the mRNA expression [118, 137]. Radom-Aizik et al. [137] verified that AAE not only prevents the normal effects of aging in terms of atherosclerosis but also reduces its symptoms in a manner that promotes cardiovascular health despite the global stress response that is generally evoked by this activity.
One exception is a study by Liao et al. [136], which showed a reduction in TLR4 (gene expression), but also showed an increase in inflammatory responses as exhibited by high levels of TNF-α, NF-kB, and LPS. The reason for the down-regulation of TLR4 is not clear, but the authors believe that this may be related to high levels of ROS. Here, from our review of the literature, we suggest that increases in circulating LPS and an excessive generation of ROS are the main actors in the acute inflammatory process generated by excessive AE. However, more studies are needed to complete the mechanistic picture that links these effects and other aspects of inflammatory responses in AE.
Combined Exercise and Inflammation
Only two studies [93, 149] relating TLR2 and/or TLR4 to CE (combining aerobic and resistance exercises in single sessions) were found. One study [93] demonstrated a significant decline in TLR4, and the other [149] did not find a difference in TLR4 (Table 8).
The Timmerman et al. [149] study analyzed the response of 12 weeks of exercise training on the part of aged, physically inactive subjects who performed AE for 20 min and RE for 30 min. No significant differences in TLR4 (protein expression) were found in the trained group compared to the controls, but a decline in TNF-α was observed. Stewart et al. [93] compared CE effects in adult and aged participants and showed a significant decline in TLR4 as well as IL-6 in the physically inactive groups compared to controls; however, levels of TLR2 were not significantly changed.
Another experiment [150] verified a decline in CRP in both trained and active control groups and concluded that AE and RE may be applied in the same session as a potential therapeutic intervention for adults and aged individuals to avoid some chronic diseases. Therefore, this review suggests that AE and RE in combination protect against the negative effects of AE.
Exercise, Disease, and Inflammation
The majority of the studies eligible for this review show that both AE [13,14,15, 113,114,115, 117] and RE [8, 116] can act as excellent auxiliary treatments for chronic disease. However, we found no article that tested ARE in samples from patients with diseases.
One of the important features of obesity-induced inflammation is a phenotypic change in the populations of macrophages and T cells present in the adipose tissue. This is reflected in levels of the production of anti- and pro-inflammatory cytokines [151]. It has been suggested that free saturated fatty acids can induce inflammation through the activation of macrophages, TLR2, and TLR4 in the adipose tissue, culminating in the activation of NF-kB and an increased expression of pro-inflammatory cytokines such as TNF-α or IL-6 [7, 9, 151].
The study by Phillips et al. [8] in post-menopausal obese women showed that CRE did not decrease TLR4 in terms of mRNA expression but reduced inflammatory markers such as TNF-α and IL-6. In another study related to obesity, 10 days of either moderate (MICT) or high intensity (HIIT) CAE in inactive overweight women promoted improvements in glucose control and cardiorespiratory capacity and a decrease in TLR2 and TLR4 (protein content) [115].
Most studies in this review that tested the levels of TLR2 and/or TLR4 receptors in a disease context used moderate load protocols, with the exception of the study by Nickel et al. [114], which studied marathon runners and found an increase in the mRNA expression and protein levels of these receptors. In this study, TLR2 was significantly increased in lean-non-elite athletes when compared to the obese-non-elite and lean-elite groups, and TLR4 increased in all groups in response to exercise. However, levels of the systemic cytokines TNF-α and IL-6 remained stable. Interestingly, oxidized low-density lipoprotein (oxLDL) levels in obese athletes were reduced and associated with higher adiponectin levels, in contrast to increased levels of oxLDL found in the group of lean-elite athletes [114]. This can be understood from the fact that TLR4 plays a crucial role in cellular responses to oxLDL exposure and the activation of NF-κB [152, 153]. Wang et al. [152] showed that the activation of the TLR4/NF-κB signaling pathway was a potential mechanism for oxLDL-induced apoptosis in cardiomyocytes.
Higher levels of this low-density lipoprotein (LDL) are usually associated with an increased risk for atherosclerosis [114], and marathon runners may, in fact, have increased levels of atherosclerosis [54]. LDL, when modified by enzymes such as phospholipases, gives rise to oxidized low-density lipoprotein (oxLDL), which contributes to the formation and progression of atherosclerotic plaques [152, 154]. oxLDL is known to be immunogenic and activates endothelial cells, monocytes, macrophages, and T cells [155]. Furthermore, oxLDL is toxic at higher concentrations and thus could be a cause of cell death in lesions [156]. The plasma level of oxLDL was shown to be a predictor of mortality in patients with chronic congestive heart failure [157] and induced severe cell damage in ventricular myocytes [158].
This review also found articles that generally analyzed TLR2 and/or TLR4 expression in relation to other diseases. The study by Zwagerman et al. [14], for example, found that in addition to reduced levels of TLR4 (gene and protein), CAE reduced the frequency of cerebral infarction. Another study [36] analyzed chronic fatigue syndrome in acute AE sessions at moderate intensity for 25 min. In addition to an increase in the mRNA expression of TLR4 and pro-inflammatory cytokines, symptoms such as pain and physical and mental fatigue became worse after exercise, suggesting a dysregulation of the immune and sympathetic nervous systems.
Conclusions
This is the first systematic review of the literature that addresses the roles of TLR2 and TLR4 receptors in various types of exercise. Our main finding is evidence for an accentuation in the inflammatory processes orchestrated by these receptors in both AAE and CAE. The results also suggest that the expression of the receptors is correlated with that of anti- and pro-inflammatory cytokines. Taken together, these data open new perspectives for studies aimed at a better understanding of the response of inflammatory processes to physical exercise.
An analysis of the pathways involving TLR2 and TLR4 reveal something about the way specific types of physical exercise are related to differences in the types of inflammatory responses they stimulate. The results indicate that AE is potentially inflammatory; a smaller number of studies revealed that acute exercise has anti-inflammatory effects, compared to studies of chronic exercise.
Our analysis showed that in RE, TLR2 and TLR4 expression and signaling adopt an anti-inflammatory pattern. Studies that met our criteria for inclusion indicated that acute or chronic sessions reduced TLRs as well as inflammatory cytokines, particularly TNF-α, and promoted increases in IL-10, which can be considered a beneficial adaptation for both healthy people and those affected by certain diseases.
The same results were obtained when differences in the populations and intensities of exercise were taken into account. This indicates that RE can be broadly used to prevent or minimize the potentially deleterious effects of TLR expression and that the intensity can be manipulated to achieve other goals, such as increasing body strength, without a loss of benefits vis-à-vis the overall inflammatory profile.
For AE, the intensity of exercise is a crucial factor—better responses were achieved under moderate intensities. But overall, whether the effects of AE will be positive or negative depends on a person’s other physiological characteristics, so they must be taken into account.
Generally, CE seems to be a good choice in most situations due to its positive effects on TLR expression and signaling. In other words, the possible negative “side effects” of AE can be overcome through the positive impact of RE. This combination of training strategies appears to improve a person’s general inflammatory profile while maintaining the cardiovascular and metabolic benefits of AE. In most cases, this leads to better adaptations. But because the number of studies addressing the effects of TLR2 and TLR4 in CE is very small, further research is needed for both amateurs and elite athletes.
Change history
31 January 2018
The original article [1] mistakenly omits a grant acknowledgement; thus, the authors would like to acknowledge that the original article was supported by FAPESP 2015/20082-7.
Abbreviations
- AAE:
-
Acute aerobic exercise
- AE:
-
Aerobic exercises
- ARE:
-
Acute resistance exercise
- Arg1:
-
Arginase-1
- CAE:
-
Chronic aerobic exercise
- CE:
-
Combined exercise
- CK:
-
Creatine kinase
- CRE:
-
Chronic resistance exercise
- CRP:
-
C-reactive protein
- DAMPs:
-
Damage-associated molecular patterns
- IGF-1:
-
Insulin-like growth factor 1
- LPS:
-
Lipopolysaccharides
- MAPK:
-
Mitogen-activated protein kinase
- PAMPS:
-
Pathogen-associated molecular patterns
- RE:
-
Resistance exercise
- ROS:
-
Reactive oxygen species
- TLR:
-
Toll-like receptor
- TLR2:
-
Toll-like receptor 2
- TLR4:
-
Toll-like receptor 4
- TNF-α:
-
Tumor necrosis factor alpha
- VEGF:
-
Vascular endothelial growth factor
References
Bermudez EA, Rifai N, Buring J, Manson JE, Ridker PM. Interrelationships among circulating interleukin-6, C-reactive protein, and traditional cardiovascular risk factors in women. Arterioscler Thromb Vasc Biol. 2002;22(10):1668–73.
Gleeson M, McFarlin B, Flynn M. Exercise and Toll-like receptors. Exerc Immunol Rev. 2006;12:34–53.
McFarlin BK, Flynn MG, Campbell WW, Craig BA, Robinson JP, Stewart LK, et al. Physical activity status, but not age, influences inflammatory biomarkers and Toll-like receptor 4. J Gerontol Ser A Biol Med Sci. 2006;61(4):388–93.
Scheidt-Nave C, Bismar H, Leidig-Bruckner G, Woitge H, Seibel MJ, Ziegler R, et al. Serum interleukin 6 is a major predictor of bone loss in women specific to the first decade past menopause. J Clin Endocrinol Metab. 2001;86(5):2032–42. https://doi.org/10.1210/jcem.86.5.7445.
Daar AS, Singer PA, Persad DL, Pramming SK, Matthews DR, Beaglehole R, et al. Grand challenges in chronic non-communicable diseases. Nature. 2007;450(7169):494–6. https://doi.org/10.1038/450494a.
Nathan C. Points of control in inflammation. Nature. 2002;420(6917):846–52. https://doi.org/10.1038/nature01320.
Fessler MB, Rudel LL, Brown JM. Toll-like receptor signaling links dietary fatty acids to the metabolic syndrome. Curr Opin Lipidol. 2009;20(5):379–85. https://doi.org/10.1097/MOL.0b013e32832fa5c4.
Phillips MD, Patrizi RM, Cheek DJ, Wooten JS, Barbee JJ, Mitchell JB. Resistance training reduces subclinical inflammation in obese, postmenopausal women. Med Sci Sports Exerc. 2012;44(11):2099–110. https://doi.org/10.1249/MSS.0b013e3182644984.
Oliveira AG, Carvalho BM, Tobar N, Ropelle ER, Pauli JR, Bagarolli RA, et al. Physical exercise reduces circulating lipopolysaccharide and TLR4 activation and improves insulin signaling in tissues of DIO rats. Diabetes. 2011;60(3):784–96. https://doi.org/10.2337/db09-1907.
Miura K, Ohnishi H. Role of gut microbiota and Toll-like receptors in nonalcoholic fatty liver disease. World J Gastroenterol. 2014;20(23):7381–91. https://doi.org/10.3748/wjg.v20.i23.7381.
Dybdahl B, Wahba A, Lien E, Flo TH, Waage A, Qureshi N, et al. Inflammatory response after open heart surgery: release of heat-shock protein 70 and signaling through toll-like receptor-4. Circulation. 2002;105(6):685–90.
Laberge MA, Moore KJ, Freeman MW. Atherosclerosis and innate immune signaling. Ann Med. 2005;37(2):130–40. https://doi.org/10.1080/07853890510007304.
Ma Y, He M, Qiang L. Exercise therapy downregulates the overexpression of TLR4, TLR2, MyD88 and NF-κB after cerebral ischemia in rats. Int J Mol Sci. 2013;14(2):3718–33. https://doi.org/10.3390/ijms14023718.
Zwagerman N, Plumlee C, Guthikonda M, Ding Y. Toll-like receptor-4 and cytokine cascade in stroke after exercise. Neurol Res. 2010;32(2):123–6. https://doi.org/10.1179/016164109x12464612122812.
Choi DH, Kwon IS, Koo JH, Jang YC, Kang EB, Byun JE, et al. The effect of treadmill exercise on inflammatory responses in rat model of streptozotocin-induced experimental dementia of Alzheimer’s type. Journal of exercise nutrition & biochemistry. 2014;18(2):225–33. https://doi.org/10.5717/jenb.2014.18.2.225.
Pierer M, Rethage J, Seibl R, Lauener R, Brentano F, Wagner U, et al. Chemokine secretion of rheumatoid arthritis synovial fibroblasts stimulated by Toll-like receptor 2 ligands. Journal of immunology (Baltimore, Md: 1950). 2004;172(2):1256–65.
Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11(5):373–84. https://doi.org/10.1038/ni.1863.
Mukherjee S, Karmakar S, Babu SP. TLR2 and TLR4 mediated host immune responses in major infectious diseases: a review. Braz J Infect Dis. 2016;20(2):193–204. https://doi.org/10.1016/j.bjid.2015.10.011.
Kawai T, Akira S. Toll-like receptors and their crosstalk with other innate receptors in infection and immunity. Immunity. 2011;34(5):637–50. https://doi.org/10.1016/j.immuni.2011.05.006.
Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392(6673):245–52. https://doi.org/10.1038/32588.
Means TK, Golenbock DT, Fenton MJ. The biology of Toll-like receptors. Cytokine Growth Factor Rev. 2000;11(3):219–32.
Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol. 2001;1(2):135–45. https://doi.org/10.1038/35100529.
Cristofaro P, Opal SM. Role of Toll-like receptors in infection and immunity: clinical implications. Drugs. 2006;66(1):15–29.
Ghosh S, May MJ, Kopp EB, NF-kappa B. Rel proteins: evolutionarily conserved mediators of immune responses. Annu Rev Immunol. 1998;16:225–60. https://doi.org/10.1146/annurev.immunol.16.1.225.
Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999;18(49):6853–66. https://doi.org/10.1038/sj.onc.1203239.
Siebenlist U, Franzoso G, Brown K. Structure, regulation and function of NF-kappa B. Annu Rev Cell Biol. 1994;10:405–55. https://doi.org/10.1146/annurev.cb.10.110194.002201.
Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124(4):783–801. https://doi.org/10.1016/j.cell.2006.02.015.
Chang ZL. Important aspects of Toll-like receptors, ligands and their signaling pathways. Inflamm Res. 2010;59(10):791–808. https://doi.org/10.1007/s00011-010-0208-2.
McFarlin BK, Flynn MG, Campbell WW, Stewart LK, Timmerman KL. TLR4 is lower in resistance-trained older women and related to inflammatory cytokines. Med Sci Sports Exerc. 2004;36(11):1876–83.
Flynn MG, McFarlin BK, Phillips MD, Stewart LK, Timmerman KL. Toll-like receptor 4 and CD14 mRNA expression are lower in resistive exercise-trained elderly women. J Appl Physiol. 2003;95(5):1833–42. https://doi.org/10.1152/japplphysiol.00359.2003.
Lee WJ. IGF-I exerts an anti-inflammatory effect on skeletal muscle cells through down-regulation of TLR4 signaling. Immune network. 2011;11(4):223–6. https://doi.org/10.4110/in.2011.11.4.223.
Petersen AM, Pedersen BK. The anti-inflammatory effect of exercise. J Appl Physiol. 2005;98(4):1154–62. https://doi.org/10.1152/japplphysiol.00164.2004.
Taaffe DR, Harris TB, Ferrucci L, Rowe J, Seeman TE. Cross-sectional and prospective relationships of interleukin-6 and C-reactive protein with physical performance in elderly persons: MacArthur studies of successful aging. J Gerontol A Biol Sci Med Sci. 2000;55(12):M709–15.
Walsh NP, Gleeson M, Pyne DB, Nieman DC, Dhabhar FS, Shephard RJ, et al. Position statement. Part two: maintaining immune health. Exerc Immunol Rev. 2011;17:64–103.
Booth S, Florida-James GD, McFarlin BK, Spielmann G, O'Connor DP, Simpson RJ. The impact of acute strenuous exercise on TLR2, TLR4 and HLA.DR expression on human blood monocytes induced by autologous serum. Eur J Appl Physiol. 2010;110(6):1259–68. https://doi.org/10.1007/s00421-010-1616-2.
Light AR, White AT, Hughen RW, Light KC. Moderate exercise increases expression for sensory, adrenergic, and immune genes in chronic fatigue syndrome patients but not in normal subjects. J Pain. 2009;10(10):1099–112. https://doi.org/10.1016/j.jpain.2009.06.003.
Neubauer O, Sabapathy S, Lazarus R, Jowett JBM, Desbrow B, Peake JM, et al. Transcriptome analysis of neutrophils after endurance exercise reveals novel signaling mechanisms in the immune response to physiological stress. J Appl Physiol. 2013;114(12):1677–88. https://doi.org/10.1152/japplphysiol.00143.2013.
Nieman DC. Exercise, infection, and immunity. Int J Sports Med. 1994;15(Suppl 3):S131–41. https://doi.org/10.1055/s-2007-1021128.
Rodriguez-Miguelez P, Lima-Cabello E, Martinez-Florez S, Almar M, Cuevas MJ, Gonzalez-Gallego J. Hypoxia-inducible factor-1 modulates the expression of vascular endothelial growth factor and endothelial nitric oxide synthase induced by eccentric exercise. J Appl Physiol. 2015;118(8):1075–83. https://doi.org/10.1152/japplphysiol.00780.2014.
Rosa JC, Lira FS, Eguchi R, Pimentel GD, Venancio DP, Cunha CA, et al. Exhaustive exercise increases inflammatory response via Toll like receptor-4 and NF-kappaBp65 pathway in rat adipose tissue. J Cell Physiol. 2011;226(6):1604–7. https://doi.org/10.1002/jcp.22490.
Simpson RJ, Bosch JA. Special issue on exercise immunology: current perspectives on aging, health and extreme performance. Brain Behav Immun. 2014;39:1–7. https://doi.org/10.1016/j.bbi.2014.03.006.
Zbinden-Foncea H, Raymackers JM, Deldicque L, Renard P, Francaux M. TLR2 and TLR4 activate p38 MAPK and JNK during endurance exercise in skeletal muscle. Med Sci Sports Exerc. 2012;44(8):1463–72. https://doi.org/10.1249/MSS.0b013e31824e0d5d.
Francaux M. Toll-like receptor signalling induced by endurance exercise. Applied physiology, nutrition, and metabolism = Physiologie appliquee, nutrition et metabolisme. 2009;34(3):454–8. https://doi.org/10.1139/h09-036.
Shek PN, Shephard RJ. Physical exercise as a human model of limited inflammatory response. Can J Physiol Pharmacol. 1998;76(5):589–97.
Moldoveanu AI, Shephard RJ, Shek PN. The cytokine response to physical activity and training. Sports Med. 2001;31(2):115–44. https://doi.org/10.2165/00007256-200131020-00004.
Nakajima T, Kurano M, Hasegawa T, Takano H, Iida H, Yasuda T, et al. Pentraxin3 and high-sensitive C-reactive protein are independent inflammatory markers released during high-intensity exercise. Eur J Appl Physiol. 2010;110(5):905–13. https://doi.org/10.1007/s00421-010-1572-x.
Okutsu M, Suzuki K, Ishijima T, Peake J, Higuchi M. The effects of acute exercise-induced cortisol on CCR2 expression on human monocytes. Brain Behav Immun. 2008;22(7):1066–71. https://doi.org/10.1016/j.bbi.2008.03.006.
Pedersen BK, Ostrowski K, Rohde T, Bruunsgaard H. The cytokine response to strenuous exercise. Can J Physiol Pharmacol. 1998;76(5):505–11.
Shephard RJ, Shek PN. Immune responses to inflammation and trauma: a physical training model. Can J Physiol Pharmacol. 1998;76(5):469–72.
Jeon J-W, Ha U-H, Paek S-H. In vitro inflammation inhibition model based on semi-continuous Toll-like receptor biosensing. PLoS One. 2014;9(8):e105212. https://doi.org/10.1371/journal.pone.0105212.
Weiss U. Inflammation. Nature. 2008;454(7203):427.
Chen YW, Apostolakis S, Lip GY. Exercise-induced changes in inflammatory processes: implications for thrombogenesis in cardiovascular disease. Ann Med. 2014;46(7):439–55. https://doi.org/10.3109/07853890.2014.927713.
Schnohr P, O’Keefe JH, Marott JL, Lange P, Jensen GB. Dose of jogging and long-term mortality: the Copenhagen City Heart Study. J Am Coll Cardiol. 2015;65(5):411–9. https://doi.org/10.1016/j.jacc.2014.11.023.
Lavie CJ, Lee DC, Sui X, Arena R, O’Keefe JH, Church TS, et al. Effects of running on chronic diseases and cardiovascular and all-cause mortality. Mayo Clin Proc. 2015;90(11):1541–52. https://doi.org/10.1016/j.mayocp.2015.08.001.
Sima AV, Stancu CS, Simionescu M. Vascular endothelium in atherosclerosis. Cell Tissue Res. 2009;335(1):191–203. https://doi.org/10.1007/s00441-008-0678-5.
Bjorkbacka H. Multiple roles of Toll-like receptor signaling in atherosclerosis. Curr Opin Lipidol. 2006;17(5):527–33. https://doi.org/10.1097/01.mol.0000245258.25387.ec.
Haskell WL, Lee IM, Pate RR, Powell KE, Blair SN, Franklin BA, et al. Physical activity and public health: updated recommendation for adults from the American College of Sports Medicine and the American Heart Association. Med Sci Sports Exerc. 2007;39(8):1423–34. https://doi.org/10.1249/mss.0b013e3180616b27.
Lavie CJ, O’Keefe JH, Sallis RE. Exercise and the heart—the harm of too little and too much. Current sports medicine reports. 2015;14(2):104–9. https://doi.org/10.1249/jsr.0000000000000134.
O’Keefe JH, Patil HR, Lavie CJ, Magalski A, Vogel RA, McCullough PA. Potential adverse cardiovascular effects from excessive endurance exercise. Mayo Clin Proc. 2012;87(6):587–95. https://doi.org/10.1016/j.mayocp.2012.04.005.
Lee S, Kim M, Lim W, Kim T, Kang C. Strenuous exercise induces mitochondrial damage in skeletal muscle of old mice. Biochem Biophys Res Commun. 2015;461(2):354–60. https://doi.org/10.1016/j.bbrc.2015.04.038.
Kong X, Thimmulappa R, Kombairaju P, Biswal S. NADPH oxidase-dependent reactive oxygen species mediate amplified TLR4 signaling and sepsis-induced mortality in Nrf2-deficient mice. Journal of immunology (Baltimore, Md: 1950). 2010;185(1):569–77. https://doi.org/10.4049/jimmunol.0902315.
Imhoff BR, Hansen JM. Extracellular redox status regulates Nrf2 activation through mitochondrial reactive oxygen species. The Biochemical journal. 2009;424(3):491–500. https://doi.org/10.1042/bj20091286.
Sen CK, Packer L. Antioxidant and redox regulation of gene transcription. FASEB J. 1996;10(7):709–20.
Moriuchi H, Moriuchi M, Fauci AS. Nuclear factor-kappa B potently up-regulates the promoter activity of RANTES, a chemokine that blocks HIV infection. Journal of immunology (Baltimore, Md: 1950). 1997;158(7):3483–91.
Park HS, Jung HY, Park EY, Kim J, Lee WJ, Bae YS. Cutting edge: direct interaction of TLR4 with NAD(P)H oxidase 4 isozyme is essential for lipopolysaccharide-induced production of reactive oxygen species and activation of NF-kappa B. Journal of immunology (Baltimore, Md: 1950). 2004;173(6):3589–93.
Lorne E, Zmijewski JW, Zhao X, Liu G, Tsuruta Y, Park Y-J, et al. Role of extracellular superoxide in neutrophil activation: interactions between xanthine oxidase and TLR4 induce proinflammatory cytokine production. Am J Physiol Cell Physiol. 2008;294(4):C985–C93. https://doi.org/10.1152/ajpcell.00454.2007.
Alessio HM, Goldfarb AH, Cutler RG. MDA content increases in fast- and slow-twitch skeletal muscle with intensity of exercise in a rat. Am J Phys. 1988;255(6 Pt 1):C874–7.
Jackson SH, Devadas S, Kwon J, Pinto LA, Williams MS. T cells express a phagocyte-type NADPH oxidase that is activated after T cell receptor stimulation. Nat Immunol. 2004;5(8):818–27. https://doi.org/10.1038/ni1096.
Mougios V. Reference intervals for serum creatine kinase in athletes. Br J Sports Med. 2007;41(10):674–8. https://doi.org/10.1136/bjsm.2006.034041.
Mangner N, Linke A, Oberbach A, Kullnick Y, Gielen S, Sandri M, et al. Exercise training prevents TNF-α induced loss of force in the diaphragm of mice. PLoS One. 2013;8(1):e52274. https://doi.org/10.1371/journal.pone.0052274.
Becatti M, Mannucci A, Barygina V, Mascherini G, Emmi G, Silvestri E, et al. Redox status alterations during the competitive season in elite soccer players: focus on peripheral leukocyte-derived ROS. Intern Emerg Med. 2017; https://doi.org/10.1007/s11739-017-1653-5.
Baur DA, Bach CW, Hyder WJ, Ormsbee MJ. Fluid retention, muscle damage, and altered body composition at the Ultraman Triathlon. Eur J Appl Physiol. 2016;116(3):447–58. https://doi.org/10.1007/s00421-015-3291-9.
Bosenberg AT, Brock-Utne JG, Gaffin SL, Wells MT, Blake GT. Strenuous exercise causes systemic endotoxemia. J Appl Physiol. 1988;65(1):106–8.
Ghosh S, Lertwattanarak R, Garduno Jde J, Galeana JJ, Li J, Zamarripa F, et al. Elevated muscle TLR4 expression and metabolic endotoxemia in human aging. J Gerontol A Biol Sci Med Sci. 2015;70(2):232–46. https://doi.org/10.1093/gerona/glu067.
Jeukendrup AE, Vet-Joop K, Sturk A, Stegen JH, Senden J, Saris WH, et al. Relationship between gastro-intestinal complaints and endotoxaemia, cytokine release and the acute-phase reaction during and after a long-distance triathlon in highly trained men. Clinical science (London, England: 1979). 2000;98(1):47–55.
Lira FS, Rosa JC, Pimentel GD, Tarini VA, Arida RM, Faloppa F, et al. Inflammation and adipose tissue: effects of progressive load training in rats. Lipids Health Dis. 2010;9:109. https://doi.org/10.1186/1476-511x-9-109.
Beutler B, Mahoney J, Le Trang N, Pekala P, Cerami A. Purification of cachectin, a lipoprotein lipase-suppressing hormone secreted by endotoxin-induced RAW 264.7 cells. J Exp Med. 1985;161(5):984–95.
Espat NJ, Copeland EM, Moldawer LL. Tumor necrosis factor and cachexia: a current perspective. Surg Oncol. 1994;3(5):255–62.
Coppack SW. Pro-inflammatory cytokines and adipose tissue. Proc Nutr Soc. 2001;60(3):349–56.
Hotamisligil GS. Inflammation and metabolic disorders. Nature. 2006;444(7121):860–7. https://doi.org/10.1038/nature05485.
Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-alpha: direct role in obesity-linked insulin resistance. Science (New York, NY). 1993;259(5091):87–91.
Lakier SL. Overtraining, excessive exercise, and altered immunity: is this a T helper-1 versus T helper-2 lymphocyte response? Sports medicine (Auckland, NZ). 2003;33(5):347–64.
Smith LL. Tissue trauma: the underlying cause of overtraining syndrome? Journal of strength and conditioning research / National Strength & Conditioning Association. 2004;18(1):185–93.
Hill EE, Zack E, Battaglini C, Viru M, Viru A, Hackney AC. Exercise and circulating cortisol levels: the intensity threshold effect. J Endocrinol Investig. 2008;31(7):587–91. https://doi.org/10.1007/bf03345606.
Kraemer WJ, Ratamess NA. Hormonal responses and adaptations to resistance exercise and training. Sports medicine (Auckland, NZ). 2005;35(4):339–61.
Jin CH, Paik IY, Kwak YS, Jee YS, Kim JY. Exhaustive submaximal endurance and resistance exercises induce temporary immunosuppression via physical and oxidative stress. Journal of exercise rehabilitation. 2015;11(4):198–203. 10.12965/jer.150221.
Radak Z, Chung HY, Koltai E, Taylor AW, Goto S. Exercise, oxidative stress and hormesis. Ageing Res Rev. 2008;7(1):34–42. https://doi.org/10.1016/j.arr.2007.04.004.
Fernandez-Gonzalo R, De Paz JA, Rodriguez-Miguelez P, Cuevas MJ, Gonzalez-Gallego J. Effects of eccentric exercise on Toll-like receptor 4 signaling pathway in peripheral blood mononuclear cells. J Appl Physiol. 2012;112(12):2011–8. https://doi.org/10.1152/japplphysiol.01499.2011.
Ford ES. Does exercise reduce inflammation? Physical activity and C-reactive protein among U.S. adults. Epidemiology (Cambridge, Mass). 2002;13(5):561–8. https://doi.org/10.1097/01.ede.0000023965.92535.c0.
Giannopoulou I, Fernhall B, Carhart R, Weinstock RS, Baynard T, Figueroa A, et al. Effects of diet and/or exercise on the adipocytokine and inflammatory cytokine levels of postmenopausal women with type 2 diabetes. Metab Clin Exp. 2005;54(7):866–75. https://doi.org/10.1016/j.metabol.2005.01.033.
Oberbach A, Tonjes A, Kloting N, Fasshauer M, Kratzsch J, Busse MW, et al. Effect of a 4 week physical training program on plasma concentrations of inflammatory markers in patients with abnormal glucose tolerance. European journal of endocrinology / European Federation of Endocrine Societies. 2006;154(4):577–85. https://doi.org/10.1530/eje.1.02127.
Rodriguez-Miguelez P, Fernandez-Gonzalo R, Collado PS, Almar M, Martinez-Florez S, de Paz JA, et al. Whole-body vibration improves the anti-inflammatory status in elderly subjects through Toll-like receptor 2 and 4 signaling pathways. Mech Ageing Dev. 2015;150:12–9. https://doi.org/10.1016/j.mad.2015.08.002.
Stewart LK, Flynn MG, Campbell WW, Craig BA, Robinson JP, McFarlin BK, et al. Influence of exercise training and age on CD14+ cell-surface expression of toll-like receptor 2 and 4. Brain Behav Immun. 2005;19(5):389–97. https://doi.org/10.1016/j.bbi.2005.04.003.
Pedersen BK, Saltin B. Evidence for prescribing exercise as therapy in chronic disease. Scand J Med Sci Sports. 2006;16(Suppl 1):3–63. https://doi.org/10.1111/j.1600-0838.2006.00520.x.
Li J, Lee DSW, Madrenas J. Evolving bacterial envelopes and plasticity of TLR2-dependent responses: basic research and translational opportunities. Front Immunol. 2013;4 https://doi.org/10.3389/fimmu.2013.00347.
Febbraio MA, Pedersen BK. Contraction-induced myokine production and release: is skeletal muscle an endocrine organ? Exerc Sport Sci Rev. 2005;33(3):114–9.
Pedersen BK, Steensberg A, Fischer C, Keller C, Keller P, Plomgaard P, et al. Searching for the exercise factor: is IL-6 a candidate? J Muscle Res Cell Motil. 2003;24(2–3):113–9.
de Rezende Gomes M, Santana de Oliveira Pires I, Alves de Castro I, Tirapegui J. Effect of moderate physical exercise on plasma and tissue levels of insulin-like growth factor-1 in adult rats. Nutr Res. 2004;24(7):555–64. https://doi.org/10.1016/j.nutres.2004.04.003.
Eliakim A, Nemet D. Exercise training, physical fitness and the growth hormone-insulin-like growth factor-1 axis and cytokine balance. Medicine and sport science. 2010;55:128–40. https://doi.org/10.1159/000321977.
Booth FW, Ruegsegger GN, Toedebusch RG, Yan Z. Endurance exercise and the regulation of skeletal muscle metabolism. Prog Mol Biol Transl Sci. 2015;135:129–51. https://doi.org/10.1016/bs.pmbts.2015.07.016.
Egan B, Zierath JR. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013;17(2):162–84. https://doi.org/10.1016/j.cmet.2012.12.012.
Smith J, Dykes R, Douglas JE, Krishnaswamy G, Berk S. Long-term exercise and atherogenic activity of blood mononuclear cells in persons at risk of developing ischemic heart disease. JAMA. 1999;281(18):1722–7. https://doi.org/10.1001/jama.281.18.1722.
Gleeson M, Bishop NC, Stensel DJ, Lindley MR, Mastana SS, Nimmo MA. The anti-inflammatory effects of exercise: mechanisms and implications for the prevention and treatment of disease. Nat Rev Immunol. 2011;11(9):607–15. https://doi.org/10.1038/nri3041.
Ostrowski K, Rohde T, Asp S, Schjerling P, Pedersen BK. Pro- and anti-inflammatory cytokine balance in strenuous exercise in humans. J Physiol. 1999;515(Pt 1):287–91.
Ostrowski K, Schjerling P, Pedersen BK. Physical activity and plasma interleukin-6 in humans—effect of intensity of exercise. Eur J Appl Physiol. 2000;83(6):512–5. https://doi.org/10.1007/s004210000312.
Steensberg A, Fischer CP, Keller C, Moller K, Pedersen BK. IL-6 enhances plasma IL-1ra, IL-10, and cortisol in humans. Am J Physiol Endocrinol Metab. 2003;285(2):E433–7. https://doi.org/10.1152/ajpendo.00074.2003.
Meckel Y, Eliakim A, Seraev M, Zaldivar F, Cooper DM, Sagiv M, et al. The effect of a brief sprint interval exercise on growth factors and inflammatory mediators. Journal of strength and conditioning research/National Strength and Conditioning Association. 2009;23(1):225–30. https://doi.org/10.1519/JSC.0b013e3181876a9a.
Pedersen BK, Fischer CP. Beneficial health effects of exercise—the role of IL-6 as a myokine. Trends Pharmacol Sci. 2007;28(4):152–6. https://doi.org/10.1016/j.tips.2007.02.002.
Petersen AM, Pedersen BK. The role of IL-6 in mediating the anti-inflammatory effects of exercise. J Physiol Pharmacol. 2006;57(Suppl 10):43–51.
Hubo M, Trinschek B, Kryczanowsky F, Tuettenberg A, Steinbrink K, Jonuleit H. Costimulatory molecules on immunogenic versus tolerogenic human dendritic cells. Front Immunol. 2013;4 https://doi.org/10.3389/fimmu.2013.00082.
Lira FS, Rosa JC, Zanchi NE, Yamashita AS, Lopes RD, Lopes AC, et al. Regulation of inflammation in the adipose tissue in cancer cachexia: effect of exercise. Cell Biochem Funct. 2009;27(2):71–5. https://doi.org/10.1002/cbf.1540.
Schottelius AJ, Mayo MW, Sartor RB, Baldwin AS Jr. Interleukin-10 signaling blocks inhibitor of kappaB kinase activity and nuclear factor kappaB DNA binding. J Biol Chem. 1999;274(45):31868–74.
Nickel T, Emslander I, Sisic Z, David R, Schmaderer C, Marx N, et al. Modulation of dendritic cells and Toll-like receptors by marathon running. Eur J Appl Physiol. 2012;112(5):1699–708. https://doi.org/10.1007/s00421-011-2140-8.
Nickel T, Hanssen H, Emslander I, Drexel V, Hertel G, Schmidt-Trucksass A, et al. Immunomodulatory effects of aerobic training in obesity. Mediat Inflamm. 2011;2011:308965. https://doi.org/10.1155/2011/308965.
Robinson E, Durrer C, Simtchouk S, Jung ME, Bourne JE, Voth E, et al. Short-term high-intensity interval and moderate-intensity continuous training reduce leukocyte TLR4 in inactive adults at elevated risk of type 2 diabetes. J Appl Physiol. 2015;119(5):508–16. https://doi.org/10.1152/japplphysiol.00334.2015.
Cheng YY, Kao CL, Ma HI, Hung CH, Wang CT, Liu DH, et al. SIRT1-related inhibition of pro-inflammatory responses and oxidative stress are involved in the mechanism of nonspecific low back pain relief after exercise through modulation of Toll-like receptor 4. J Biochem. 2015;158(4):299–308. https://doi.org/10.1093/jb/mvv041.
Fashi M, Agha Alinejad H, Asilian Mahabadi H. The effect of aerobic exercise in ambient particulate matter on lung tissue inflammation and lung cancer. Iranian journal of cancer prevention. 2015;8(3):e2333. 10.17795/ijcp2333.
White AT, Light AR, Hughen RW, Vanhaitsma TA, Light KC. Differences in metabolite-detecting, adrenergic, and immune gene expression after moderate exercise in patients with chronic fatigue syndrome, patients with multiple sclerosis, and healthy controls. Psychosom Med. 2012;74(1):46–54. https://doi.org/10.1097/PSY.0b013e31824152ed.
Rodriguez-Miguelez P, Fernandez-Gonzalo R, Almar M, Mejias Y, Rivas A, de Paz JA, et al. Role of Toll-like receptor 2 and 4 signaling pathways on the inflammatory response to resistance training in elderly subjects. Age (Dordr). 2014;36(6):9734. https://doi.org/10.1007/s11357-014-9734-0.
Zanchi NE, Lira FS, de Siqueira Filho MA, Rosa JC, de Oliveira Carvalho CR, Seelaender M, et al. Chronic low frequency/low volume resistance training reduces pro-inflammatory cytokine protein levels and TLR4 mRNA in rat skeletal muscle. Eur J Appl Physiol. 2010;109(6):1095–102. https://doi.org/10.1007/s00421-010-1456-0.
Prestes J, da Cunha Nascimento D, Tibana RA, Teixeira TG, Vieira DC, Tajra V, et al. Understanding the individual responsiveness to resistance training periodization. Age (Dordr). 2015;37(3):9793. https://doi.org/10.1007/s11357-015-9793-x.
Fernandez-Gonzalo R, De Paz JA, Rodriguez-Miguelez P, Cuevas MJ, Gonzalez-Gallego J. TLR4-mediated blunting of inflammatory responses to eccentric exercise in young women. 2014;2014:479395. doi:https://doi.org/10.1155/2014/479395.
Millard A-L, Valli PV, Stussi G, Mueller NJ, Yung GP, Seebach JD. Brief exercise increases peripheral blood NK cell counts without immediate functional changes, but impairs their responses to ex vivo stimulation. Front Immunol. 2013; https://doi.org/10.3389/fimmu.2013.00125.
Greiwe JS, Cheng B, Rubin DC, Yarasheski KE, Semenkovich CF. Resistance exercise decreases skeletal muscle tumor necrosis factor alpha in frail elderly humans. FASEB J. 2001;15(2):475–82. https://doi.org/10.1096/fj.00-0274com.
Phillips MD, Flynn MG, McFarlin BK, Stewart LK, Timmerman KL. Resistance training at eight-repetition maximum reduces the inflammatory milieu in elderly women. Med Sci Sports Exerc. 2010;42(2):314–25. https://doi.org/10.1249/MSS.0b013e3181b11ab7.
Fatouros I, Chatzinikolaou A, Paltoglou G, Petridou A, Avloniti A, Jamurtas A, et al. Acute resistance exercise results in catecholaminergic rather than hypothalamic-pituitary-adrenal axis stimulation during exercise in young men. Stress (Amsterdam, Netherlands). 2010;13(6):461–8. https://doi.org/10.3109/10253891003743432.
Przybyla B, Gurley C, Harvey JF, Bearden E, Kortebein P, Evans WJ, et al. Aging alters macrophage properties in human skeletal muscle both at rest and in response to acute resistance exercise. Exp Gerontol. 2006;41(3):320–7. https://doi.org/10.1016/j.exger.2005.12.007.
Barnes JN, Trombold JR, Dhindsa M, Lin HF, Tanaka H. Arterial stiffening following eccentric exercise-induced muscle damage. J Appl Physiol. 2010;109(4):1102–8. https://doi.org/10.1152/japplphysiol.00548.2010.
Stupka N, Tarnopolsky MA, Yardley NJ, Phillips SM. Cellular adaptation to repeated eccentric exercise-induced muscle damage. J Appl Physiol. 2001;91(4):1669–78.
Jun JK, Lee WL, Park HG, Lee SK, Jeong SH, Lee YR. Moderate intensity exercise inhibits macrophage infiltration and attenuates adipocyte inflammation in ovariectomized rats. Journal of exercise nutrition and biochemistry. 2014;18(1):119–27. https://doi.org/10.5717/jenb.2014.18.1.119.
Zheng Q, Cui G, Chen J, Gao H, Wei Y, Uede T, et al. Regular exercise enhances the immune response against microbial antigens through up-regulation of Toll-like receptor signaling pathways. Cell Physiol Biochem. 2015;37(2):735–46. https://doi.org/10.1159/000430391.
Holland AM, Hyatt HW, Smuder AJ, Sollanek KJ, Morton AB, Roberts MD, et al. Influence of endurance exercise training on antioxidant enzymes, tight junction proteins, and inflammatory markers in the rat ileum. BMC research notes. 2015;8:514. https://doi.org/10.1186/s13104-015-1500-6.
Lancaster GI, Khan Q, Drysdale P, Wallace F, Jeukendrup AE, Drayson MT, et al. The physiological regulation of Toll-like receptor expression and function in humans. J Physiol. 2005;563(Pt 3):945–55. https://doi.org/10.1113/jphysiol.2004.081224.
Oliveira M, Gleeson M. The influence of prolonged cycling on monocyte Toll-like receptor 2 and 4 expression in healthy men. Eur J Appl Physiol. 2010;109(2):251–7. https://doi.org/10.1007/s00421-009-1350-9.
Simpson RJ, McFarlin BK, McSporran C, Spielmann G, Hartaigh B, Guy K. Toll-like receptor expression on classic and pro-inflammatory blood monocytes after acute exercise in humans. Brain Behav Immun. 2009;23(2):232–9. https://doi.org/10.1016/j.bbi.2008.09.013.
Liao P, Zhou J, Ji LL, Zhang Y. Eccentric contraction induces inflammatory responses in rat skeletal muscle: role of tumor necrosis factor-alpha. American Journal of Physiology Regulatory, Integrative and Comparative Physiology. 2010;298(3):R599–607. https://doi.org/10.1152/ajpregu.00480.2009.
Radom-Aizik S, Zaldivar FP Jr, Haddad F, Cooper DM. Impact of brief exercise on circulating monocyte gene and microRNA expression: implications for atherosclerotic vascular disease. Brain Behav Immun. 2014;39:121–9. https://doi.org/10.1016/j.bbi.2014.01.003.
Tanaka Y, Kawanishi N, Shiva D, Tsutsumi N, Uchida M, Kitamura H, et al. Exhaustive exercise reduces tumor necrosis factor-alpha production in response to lipopolysaccharide in mice. Neuroimmunomodulation. 2010;17(4):279–86. https://doi.org/10.1159/000290044.
Li H, Geib RW. Exploring the use of five color flow cytometry to examine the effect of acute tai chi practice on pro inflammatory monocyte subtypes—biomed 2013. Biomed Sci Instrum. 2013;49:209–15.
Ortega E, Hinchado MD, Martin-Cordero L, Asea A. The effect of stress-inducible extracellular Hsp72 on human neutrophil chemotaxis: a role during acute intense exercise. Stress (Amsterdam, Netherlands). 2009;12(3):240–9. https://doi.org/10.1080/10253890802309853.
Kohut ML, McCann DA, Russell DW, Konopka DN, Cunnick JE, Franke WD, et al. Aerobic exercise, but not flexibility/resistance exercise, reduces serum IL-18, CRP, and IL-6 independent of beta-blockers, BMI, and psychosocial factors in older adults. Brain Behav Immun. 2006;20(3):201–9. https://doi.org/10.1016/j.bbi.2005.12.002.
Tilstra JS, Clauson CL, Niedernhofer LJ, Robbins PD. NF-kappaB in Aging and Disease. Aging and disease. 2011;2(6):449–65.
Beyer I, Mets T, Bautmans I. Chronic low-grade inflammation and age-related sarcopenia. Current Opinion in Clinical Nutrition and Metabolic Care. 2012;15(1):12–22. https://doi.org/10.1097/MCO.0b013e32834dd297.
McCullough PA, Lavie CJ. Coronary artery plaque and cardiotoxicity as a result of extreme endurance exercise. Mo Med. 2014;111:9Y94.
O’Keefe JH, Lavie CJ, Guazzi M. Part 1: potential dangers of extreme endurance exercise: how much is too much? Part 2: screening of school-age athletes. Prog Cardiovasc Dis. 2015;57(4):396–405. https://doi.org/10.1016/j.pcad.2014.11.004.
Gill SK, Hankey J, Wright A, Marczak S, Hemming K, Allerton DM, et al. The impact of a 24-h ultra-marathon on circulatory endotoxin and cytokine profile. Int J Sports Med. 2015;36(8):688–95. https://doi.org/10.1055/s-0034-1398535.
Pedersen BK, Bruunsgaard H, Jensen M, Krzywkowski K, Ostrowski K. Exercise and immune function: effect of ageing and nutrition. Proc Nutr Soc. 1999;58(3):733–42.
Radak Z, Taylor AW, Ohno H, Goto S. Adaptation to exercise-induced oxidative stress: from muscle to brain. Exerc Immunol Rev. 2001;7:90–107.
Timmerman KL, Flynn MG, Coen PM, Markofski MM, Pence BD. Exercise training-induced lowering of inflammatory (CD14+CD16+) monocytes: a role in the anti-inflammatory influence of exercise? J Leukoc Biol. 2008;84(5):1271–8. https://doi.org/10.1189/jlb.0408244.
Stewart LK, Flynn MG, Campbell WW, Craig BA, Robinson JP, Timmerman KL, et al. The influence of exercise training on inflammatory cytokines and C-reactive protein. Med Sci Sports Exerc. 2007;39(10):1714–9. https://doi.org/10.1249/mss.0b013e31811ece1c.
Johnson AR, Milner JJ, Makowski L. The inflammation highway: metabolism accelerates inflammatory traffic in obesity. Immunol Rev. 2012;249(1):218–38. https://doi.org/10.1111/j.1600-065X.2012.01151.x.
Wang X, Sun Y, Yang H, Lu Y, Li L. Oxidized low-density lipoprotein induces apoptosis in cultured neonatal rat cardiomyocytes by modulating the TLR4/NF-κB pathway. Sci Rep. 2016;6:27866. https://doi.org/10.1038/srep27866.
Yang K, Zhang XJ, Cao LJ, Liu XH, Liu ZH, Wang XQ, et al. Toll-like receptor 4 mediates inflammatory cytokine secretion in smooth muscle cells induced by oxidized low-density lipoprotein. PLoS One. 2014;9(4):e95935. https://doi.org/10.1371/journal.pone.0095935.
Frostegård J. Immunity, atherosclerosis and cardiovascular disease. BMC Med. 2013;11:117. https://doi.org/10.1186/1741-7015-11-117.
Frostegård J, Zhang Y, Sun J, Yan K, Liu A. Oxidized low-density lipoprotein (OxLDL)-treated dendritic cells promote activation of T cells in human atherosclerotic plaque and blood, which is repressed by Statins: microRNA let-7c is integral to the effect. J Am Heart Assoc. 2016;5(9) https://doi.org/10.1161/jaha.116.003976.
Lizard G, Monier S, Cordelet C, Gesquière L, Deckert V, Gueldry S, et al. Characterization and Comparison of the Mode of Cell Death, Apoptosis Versus Necrosis, Induced by 7β-hydroxycholesterol and 7-ketocholesterol in the cells of the vascular wall. Arterioscler Thromb Vasc Biol. 1999;19(5):1190–200. https://doi.org/10.1161/01.atv.19.5.1190.
Tsutsui T, Tsutamoto T, Wada A, Maeda K, Mabuchi N, Hayashi M, et al. Plasma oxidized low-density lipoprotein as a prognostic predictor in patients with chronic congestive heart failure. J Am Coll Cardiol. 2002;39(6):957–62. https://doi.org/10.1016/s0735-1097(02)01721-7.
Zorn-Pauly K, Schaffer P, Pelzmann B, Bernhart E, Wei G, Lang P, et al. Oxidized LDL induces ventricular myocyte damage and abnormal electrical activity—role of lipid hydroperoxides. Cardiovasc Res. 2005;66(1):74–83. https://doi.org/10.1016/j.cardiores.2004.12.009.
Acknowledgments
We thank Russ Hodge for assisting in writing the manuscript.
Availability of Data and Materials
Not applicable.
Funding
The authors have received no payment or services from a third party for any aspect of the submitted work.
Author information
Authors and Affiliations
Contributions
PAMC: substantial contributions to the conception; design and drafting of the work; survey of the literature; preparation of tables and creation of figures; analysis; interpretation of data; critical review. MFG: contributions to the conception; review of tables; analysis; critical review. JSH: contributed to the analysis; English translation; critical review. FHO: contributions to the conception; analysis; critical review. RCA: contributions to the conception; analysis, interpretation of data; critical review. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Authors’ Information
PAMC: Ph.D. Student in Medicine (Nephrology) Program at Federal University of São Paulo, Department of Biophysics at Federal University of São Paulo, and Laboratory of Exercise Genetics and Metabolism at Federal University of São Paulo (UNIFESP), São Paulo, SP, Brazil with a collaborative period in Berlin, Germany at Max-Delbrück-Centrum für Molekulare Medizin; Master of Physical Education at São Judas Tadeu University, São Paulo, SP, Brazil. MFG: Ph.D. Student in Molecular Biology Program at Federal University of São Paulo, Department of Biophysics at Federal University of São Paulo, and Laboratory of Exercise Genetics and Metabolism at Federal University of São Paulo (UNIFESP), São Paulo, SP, Brazil; Master of Molecular Biology at Federal University of São Paulo, São Paulo, SP, Brazil. JSH: Master Degree Student in Neurology/Neuroscience Program at Federal University of São Paulo and Exercise Neurophysiology Laboratory at Federal University of São Paulo (UNIFESP), São Paulo, SP, Brazil; Graduation in Physical Education at University of Santos (UNIMES), São Paulo, SP, Brazil. FHO: Ph.D. Student in Medicine (Nephrology) Program at Federal University of São Paulo, Department of Biophysics at Federal University of São Paulo, and Laboratory of Exercise Genetics and Metabolism at Federal University of São Paulo (UNIFESP), São Paulo, SP, Brazil; Master of Health Sciences at Faculty of Medical Sciences of Santa Casa (FCMSCSP), São Paulo, SP, Brazil. RCA: Post-doctorate in Biological Sciences at Federal University of São Paulo (UNIFESP), SP, Brazil; Ph.D. in Biological Sciences (Molecular Biology) at Federal University of São Paulo (UNIFESP), SP, Brazil with a collaborative period in Berlin, Germany at Max-Delbrück-Centrum für Molekulare Medizin; Master of Biological Sciences (Physiology and Pharmacology) at Federal University of Minas Gerais (UFMG), MG, Brazil. He is currently a professor of the Biophysics Department of the Federal University of São Paulo (EPM). He has an experience in the field of Biophysics, working mainly on the following subjects: physical exercise, obesity, kinins, transgenic animals, kallikrein, inflammation, and blood pressure.
Ethics Approval and Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
Paula Andréa Malveira Cavalcante, Marcos Fernandes Gregnani, Jessica Salles Henrique, Fábio Henrique Ornellas, and Ronaldo Carvalho Araújo declare that they have no conflicts of interest related to this manuscript.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional information
A correction to this article is available online at https://doi.org/10.1186/s40798-018-0120-9.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Cavalcante, P.A.M., Gregnani, M.F., Henrique, J.S. et al. Aerobic but not Resistance Exercise Can Induce Inflammatory Pathways via Toll-Like 2 and 4: a Systematic Review. Sports Med - Open 3, 42 (2017). https://doi.org/10.1186/s40798-017-0111-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40798-017-0111-2