
Abstract
Background
Vast areas of lowland neotropical forest have regenerated after initially being cleared for agricultural purposes. The ecological value of regenerating second growth to forest-dwelling birds may largely depend on the age of the forest, associated vegetative structure, and when it is capable of sustaining avian demographics similar to those found in pristine forest.
Methods
To determine the influence of second growth age on bird demography, we estimated the annual survival of six central Amazonian bird species residing in pristine forest, a single 100 and a single 10 ha forest fragment, taking into consideration age of the surrounding matrix (i.e. regenerating forest adjacent to each fragment) as an explanatory variable.
Results
Study species exhibited three responses: arboreal, flocking and ant-following insectivores (Willisornis poecilinotus, Thamnomanes ardesiacus and Pithys albifrons) showed declines in survival associated with fragmentation followed by an increase in survival after 5 years of matrix regeneration. Conversely, Percnostola rufifrons, a gap-specialist, showed elevated survival in response to fragmentation followed by a decline after 5 years of regeneration. Lastly, facultative flocking and frugivore species (Glyphorynchus spirurus and Dixiphia pipra, respectively) showed no response to adjacent clearing and subsequent regeneration.
Conclusions
Our results in association with previous studies confirm that the value of regenerating forest surrounding habitat patches is dependent on two factors: ecological guild of the species in question and second growth age. Given the rapid increase in survival following succession, we suggest that the ecological value of young tropical forest should not be based solely on a contemporary snapshot, but rather, on the future value of mature second growth as well.
Similar content being viewed by others


Background
Bird populations in tropical forest fragments are often considered dynamic (i.e. subject to frequent turnover) relative to their counterparts in pristine forest (Laurance et al. 2011). The dynamic nature of bird populations in forest fragments is in part governed by the condition of the surrounding habitat or matrix where presence of corridors, history of land-use, matrix age, and distance to source populations can determine the balance between extinction and persistence in forest fragments (Renjifo 2001; Ewers and Didham 2006; Stouffer and Bierregaard 2007; Sodhi et al. 2011; Wolfe et al. 2015). Matrix-mediated changes in bird populations within forest fragments can occur rapidly, or unfold over longer periods of time, as documented in the central Amazon (Stouffer et al. 2011; Wolfe et al. 2015).
At the Biological Dynamics of Forest Fragmentation Project (BDFFP) near Manaus, Brazil, the relationship between bird populations within fragments and the surrounding matrix was measured over a 30-year period, where the fragmentation of pristine forest led to a rapid increase in bird capture rates, indicative of higher abundance following isolation (Bierregaard and Lovejoy 1989). Increased capture rates probably reflect a “crowding effect” where birds sought refuge from deforestation by moving into remnant habitat patches (Bierregaard et al. 1992; Debinski and Holt 2000). After several months of elevated bird abundance, the number of captures in forest fragments declined, culminating in the disappearance of several species from fragments (Bierregaard and Lovejoy 1989; Bierregaard et al. 1992). The pattern of rapid increase in abundance followed by decline within fragments suggests that a formerly open system had become closed after clearing, thereby eliminating dispersal and expediting local extinction events. After years of matrix regeneration, many of the previously absent species not only returned to the fragments (Stouffer et al. 2011), but also established themselves within adjacent second growth (Wolfe et al. 2015). Once established, these pioneering bird populations in fragments and second growth may be subject to source-sink dynamics where vulnerable species periodically perish and are subsequently replaced by individuals from higher quality habitats with higher survival and reproductive surpluses (Sodhi et al. 2011). Clearly, the transition of Amazonian forest fragments from depauperate patches to potential sources may principally depend on the condition of the surrounding matrix (Renjifo 2001; Ewers and Didham 2006; Stouffer and Bierregaard 2007; Kennedy et al. 2010). The dynamic role of a developing matrix revealed at the BDFFP suggests that bird survival in fragments is likely mediated by the matrix as well. Therefore, exploring critical links between the survival of tropical birds within forest fragments and the condition of the surrounding matrix should be an essential focus of conservation biology in tropical forests.
To date, few studies have examined avian survival in forest fragments as a function of the condition of the surrounding matrix. One such study found that Ovenbird (Seiurus aurocapillus) apparent annual survival in Canada was considerably lower (34%) in small fragments (> 15 ha) in an agricultural landscape than in forestry fragments (56%) and continuous forest (62%) (Bayne and Hobson 2002). Low apparent survival may have been driven by heightened dispersal out of the agricultural landscape, where birds incapable of successfully nesting left the area (Bayne and Hobson 2002). Additionally, differences in apparent survival between treatments suggest that edge effects, or changes in population dynamics at the boundary of two habitats, associated with a low-contrast forestry matrix did not yield as negative of an impact as those associated with a high-contrast agricultural matrix (Bayne and Hobson 2002). Clearly, not all edge effects are equal: amount of contrast between matrix and habitat yield an asymmetrical influence on avian survival within forest fragments (Bayne and Hobson 2002). The asymmetrical influence of edge effects associated with fragmented landscapes has been implicated in the decline of plant and animal populations throughout the globe (Debinski and Holt 2000). Thus, birds in fragments surrounded by a young matrix may be subject to the deleterious synergy of edge effects and dispersal limitation relative to fragments surrounded by mature matrix (Gilpin and Soulé 1986).
Deleterious edge effects and dispersal limitation may be particularly acute in tropical systems where insectivorous and flocking birds are often more sedentary, closely tied to forest-like characteristics, and sometimes incapable of dispersing long distances (Stouffer and Bierregaard 1995; Laurance et al. 2004; Sodhi et al. 2004; Moore et al. 2008). These sensitive tropical birds may avoid edges comprised of young second growth where heightened exposure to wind, sun, increased air temperature, increased soil temperature and storm damage can result in the death of trees they rely upon, as well as modifications to the microclimates they prefer (Saunders et al. 1991; Camargo and Kapos 1995; Murcia 1995; Laurance and Curran 2008). The cumulative threat of edge-induced mortality is meaningful; approximately half of the world’s tropical forests are degraded and in some stage of regeneration (Wright 2005). Thus, assessing the influence of matrix age and associated edge effects on tropical bird survival within habitat fragments is of critical importance. Here we used long-term capture data from a 10 and 100 ha forest fragment in the central Amazon to test the following hypothesis and prediction: if age and associated vegetative structure of the adjacent matrix influences survival of sensitive avian foraging guilds (flocking, arboreal insectivores and ant-followers), then survival estimates of sensitive foraging guilds will be positively associated with matrix age.
Methods
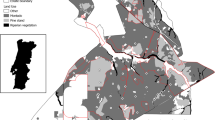
The study was conducted in terra firme Amazonian rainforest at the BDFFP, approximately 80 km north of Manaus, Brazil (Fig. 1). The BDFFP consists of 1, 10 and 100 ha forest fragments as part of the largest and longest operating landscape fragmentation experiment on Earth (for a history of the project see Bierregaard 2001). Forests at BDFFP are characterized by nutrient poor soils, diverse tree and canopy assemblages where the most abundant species of trees occur within several families: Lecythidaceae, Fabaceae, Sapotaceae and Burseraceae (Rankin-de-Mérona et al. 1992). Here, we used the BDFFP’s long-term bird banding database that encompasses data collected within fragments from 1979, before isolation occurred, through 1993. Mist-nets (12 m, 36 mm mesh), with the bottom trammel set at ground level, were used to capture birds. The number of nets differed among fragments; a single line of 16 nets was used in 10 ha fragments, and three single lines of 16 nets were used in 100 ha fragments. We considered one complete sample a single day of netting that began at 06:00 until 14:00 h. All captured birds except hummingbirds were banded with uniquely numbered aluminum bands, and then processed to record weight, age, sex, wing chord, and body and flight feather molt. To ascertain the effects of matrix condition on avian survival, we only used data from fragments if sampling occurred during three discrete time periods: (1) pre-isolation, (2) less than 5 years of matrix regeneration, and (3) more than 5 years of matrix regeneration. Fragments that were only sampled during two time periods, pre-isolation and less than 5 years of matrix regeneration, were not used in the analysis. The amount of time between individual sampling events varied over time; fragments were generally sampled at least every 6 weeks during the course of this study from 1979 through 1992, although fragments were sampled more often for about 1 year before and after initial isolation (Stouffer et al. 2006). In this study, we only used data from two fragments, a single 10 ha (Cidade Powell) and 100 ha (Porto Alegre) fragment, that met our stringent data selection criteria based on: (1) at least three samples per year, (2) capture data from pre-isolation, < 5-year post-isolation, and > 5-year post-isolation periods, and (3) enough capture data for model convergence. We recognize that limiting bird captures to only two forest fragments reduced the exportability of our results; however, to reduce data heterogeneity, subsequent overdispersion, and promote model convergence, we prioritized data quality (relying on two fragments) over quantity (including additional fragments with poorer quality data).

Map of the Biological Dynamics of Forest Fragmentation Project (BDFFP) detailing “ranch areas” that were used in the project which include areas of continuous and secondary forest, and experimental forest fragments. The map also details history of fragment isolation from 1979 to 2011, location of access roads, and location of historic and contemporary mistnet locations. For our analysis, we used pre-and-post isolation bird capture data from a single 10 ha (Esteio) and 100 ha (Porto Alegre) fragment
We chose six study species based on their diverse ecological, dietary guilds (Table 1) and high number of captures/recaptures to ensure that we could examine the effects of matrix condition on survival. Three of our study species are characterized as a flock obligate, arboreal insectivore, or obligate ant-follower (Thamnomanes ardesiacus, Willisornis poecilonotus, and Pithys albifrons, respectively), ecological guilds associated with population decline when subjected to forest fragmentation and degradation (Stouffer and Bierregaard 2007; Stouffer et al. 2011; Wolfe et al. 2015). Conversely, our other three study species, Dixiphia pipra, Glyphorynchus spirurus, Percnostola rufifrons, are associated with frugivory, facultatively joining flocks, and forest gaps, respectively, ecological guilds associated with resilience to forest fragmentation and degradation (Stouffer and Bierregaard 2007; Stouffer et al. 2011; Powell et al. 2015; Wolfe et al. 2015). Between 1979 and 2008, we captured 57,891 birds representing 183 species at BDFFP. Out of the 57,891 individuals captured, 18,584 were individuals from one of our six study species, representing approximately 32% of the avian community documented via mist-net capture. Capture histories for each species were constructed by pooling all samples within a calendar year into a single occasion then grouping birds by fragment and transients. We defined transients as individuals that were only captured once within a single year and residents were birds captured more than once within a single year. Transient models can account for survival deflation because of floater individuals moving through the study area (Pradel et al. 1997). We used Cormack-Jolly-Seber models in Program MARK (White and Burnham 1999) to estimate annual survival for each species where apparent survival (“ϕ”) and recapture probability (“p”) parameters jointly varied by time (“t” or year), time independence (“.” or no effect of year), group [“g”, which included fragment and transients (individuals separated into two groups: captured only once within a year, or captured more than once within a year)], and group-time interactions (g × t). To determine the influence of matrix condition on study species’ survival, we formulated linear time trends within Program MARK’s design matrix representative of two stages of matrix succession; time trends were calculated for each fragment’s unique isolation and re-isolation history: (1) pre-isolation, (2) less than 5 years of matrix regeneration, and (3) more than 5 years of matrix regeneration. We associated time trends (representative of three levels of matrix growth) with the grouping variable (g-matrix) for the survival parameter only. Thus, each study species had a total of eight candidate a priori models (Table 1).
We evaluated model fit two ways: (1) testing for trap happiness/shyness, and (2) quantifying over-dispersion. Program RELEASE goodness-of-fit (GOF) was used to test for trap happy/shy individuals in program MARK (Cooch and White 2018). Program Release performs two separate GOF tests (three if comparing groups, which we did not): test 2 measures the probability that an individual known to be alive at occasion (i) is captured again depending on whether it was marked at or before occasion (i) and test 3 measures whether, among those animals captured again, does when they were seen depend on whether they were marked on or before occasion (i) (Cooch and White 2018). Importantly, program RELEASE results are not unequivocal, and failure to reject the null hypothesis may reflect a lack of power to detect meaningful differences in net shyness. We calculated the over-dispersion factor (ĉ) for each species by dividing the deviance of each species’ global model by the deviance estimated via a boot-strapping goodness-of-fit routine (using 1000 iterations). Top models were selected if they were at least two Akaike information criterion (AICc) values less than, and had fewer parameters relative to the next most competitive model (Arnold 2010). Taxonomy follows Remsen et al. (2011).
Results
From 1979‒1993 we recorded 1857 captures in the two experimental fragments representing 932 individual birds of the six study species (Table 1). The gap specialist, Percnostola rufifrons, was the least frequently captured species while the single frugivore and arboreal insectivore, Dixiphia pipra and Willisornis poecilonotus, were the most commonly captured species. Program RELEASE yielded non-significant values (p < 0.05) for models associated with each study species, indicating no identifiable trap shyness. Additionally, models associated with each species yielded ĉ < 2, indicative of adequate overdispersion. All six study species exhibited nearly the same top competitive model which included a grouping variable (fragment and transiency) and matrix condition (time trend representative of three stages of matrix regeneration) associated with annual survival (Table 1). Recapture probabilities in each top model included a group and time interaction (g × t) except for Willisornis poecilonotus which yielded a time dependent recapture probability parameter (t). Top models were unequivocal (all ∆AICc > 28) (Table 1) and ∆AICc > 128 from the null model (Table 1).
Our six study species exhibited three responses to matrix clearing and subsequent regeneration. The arboreal insectivore (Willisornis poecilonotus), obligate ant-follower (Pithys albifrons) and obligate flocking species (Thamnomanes ardesiacus) all suffered a decline in apparent survival within the first 5 years after isolation, followed by a marginal recovery in the following 5-year period of matrix regeneration in both the 100 ha and 10 ha fragments (Table 2). Conversely, the flock drop-out (Glyphorynchus spirurus) and frugivore (Dixiphia pipra) showed virtually no change over the course of the study (Table 2). The gap specialist (Percnostola rufifrons) responded positively to isolation as demonstrated by an increase in survival within the first 5 years after isolation, followed by a slight decrease in apparent survival during the subsequent 5-year period of matrix regeneration in both the 100 ha and 10 ha fragments (Table 2).
Discussion
Bird populations in forest fragments are often subject to source-sink dynamics (Renjifo 2001; Ewers and Didham 2006) where the condition of the surrounding matrix may tip the balance in favor of survival and reproductive surplus or population decline (Ewers and Didham 2006; Sodhi et al. 2011). In this study we demonstrated that apparent survival of tropical birds residing in forest fragments was closely associated with the condition of the adjacent matrix. As predicted, the flock obligate, arboreal insectivore and obligate ant-follower species (Pithys albifrons, Thamnomanes ardesiacus and Willisornis poecilonotus) all exhibited reductions in apparent survival within the first five years after initial isolation (Fig. 2). The reduction in apparent survival coincided with their decreased abundance within fragments (Bierregaard and Lovejoy 1989). Considering these forest-dwelling insectivores are averse to crossing roads and clearings (Stouffer and Bierregaard 1995) we suspect that documented decreases in apparent survival do not represent dispersal events through a barren matrix, but rather suggests reduction in actual survival. Potential reductions in survival may be a direct result of high-contrast edge effects where tree mortality, changes in insect communities, and changes in ambient soil and air temperature wielded negative forces on Thamnomanes ardesiacus, Pithys albifrons, and Willisornis poecilonotus residing in forest fragments.

Apparent survival estimates with standard errors for three central Amazonian bird species captured in a 10 ha fragment. Survival varies as a function of age of the surrounding matrix. Foraging guild affiliation is denoted as follows: (In, Fob—Insectivore, Flock Obligate; (In, Ar)—Insectivore, Arboreal; (In, Anf)—Insectivore, Ant-follower
Although young matrix was associated with lower survival among the flock obligate, arboreal insectivore and obligate ant follower species (Thamnomanes ardesiacus, Willisornis poecilonotus, and Pithys albifrons), we found the reverse effect with Percnostola rufifrons, the single gap specialist (Fig. 3). Percnostola rufifrons exhibited an increase in survival during the first five years after isolation. Being a gap specialist, Percnostola rufifrons has acquired evolutionary adaptations to life on the edge. For example, Percnostola rufifrons is the only known Neotropical passerine to exhibit a third molt (the presupplemental molt), within their first year of life (Johnson and Wolfe 2014, 2017); the insertion of an extra molt may mitigate the rigors of living in a potentially hostile environment by expediting the replacement of degraded plumage. Other adaptations associated with Percnostola rufifrons that have yet to be documented may also include an ability to forage on a diversity of arthropod communities associated with regenerating forest, edge, and mature forest. In addition to the three study species that exhibited a decrease and one that exhibited an increase in survival relative to the condition of the surrounding matrix, two other species, Glyphorynchus spirurus and Dixiphia pipra, appeared to show only a negligible increase in survival when subjected to a young matrix; this finding supports previous work suggesting that Glyphorynchus spirurus does not avoid edge habitat (Powell et al. 2015).

Apparent survival estimates with standard errors for three central Amazonian bird species captured in a 10 ha fragment. Survival varies as a function of age of the surrounding matrix. Foraging guild affiliation is denoted as follows: (In, Fld)—Insectivore, Flock-dropout; (In, Ga)—Insectivore, Gap; (Fr, Co)—Frugivore, Core
We assert that the increase in survival for the gap specialist (Percnostola rufifrons) represents a real increase in survival associated with edge and regenerating forest habitats, or maybe a lack of competitors. We also assert that the static response of Glyphorynchus spirurus and Dixiphia pipra represents the resiliency of these species to fragmented forests. Young regenerating tropical forest is dynamic and subject to more sun and varying levels of soil moisture than primary forest (Wolfe et al. 2014). The combination of increased solar exposure and oscillating periods of soil moisture can elevate primary productivity (Brown and Lugo 1990) in regenerating tropical forest resulting in more insect and fruit resources for birds not adverse to foraging along edge (e.g. Glyphorynchus spirurus, Dixiphia pipra and Percnostola rufifrons). These same regenerating tropical forests may also be water limited and highly unstable during droughts when fruits and insects become scarce (Wolfe et al. 2015). Although our study indicated that Glyphorynchus spirurus, Dixiphia pipra and Percnostola rufifrons may exhibit higher apparent survival near edge, previous studies suggest that bird populations in regenerating forest may be subject to boom-bust cycles as a result of precipitation regimes (Wolfe et al. 2015; Ryder and Sillett 2016). We suggest that Glyphorynchus spirurus, Dixiphia pipra and Percnostola rufifrons may be adapted to living with unstable resources unlike sensitive species, Pithys albifrons, Thamnomanes ardesiacus and Willisornis poecilonotus, which may be averse to foraging along edge and reliant on more stable resources found in continuous forest.
The influence of matrix age on the apparent survival of birds residing in Amazonian forest fragments is variable where young matrix was detrimental to forest dwelling species, beneficial to a gap specialist and inconsequential to others. Insectivorous birds that rely upon mature forest are of special conservation concern (Şekercioḡlu et al. 2002), and our study suggests that young regenerating matrix (> 5 years) has less ecological value than mature matrix for these sensitive species. However, each of the sensitive study species (Pithys albifrons, Thamnomanes ardesiacus and Willisornis poecilonotus) were found to readily disperse through and occupy older (25 years old) second growth (see Wolfe et al. 2015). Thus, young regenerating matrixes should be viewed as having ecological value for certain species, while hosting unrealized potential for others.
Numerous hypotheses have been developed and tested—often equivocally—attempting to explain the sensitivity of insectivores and mixed-species flocks to forest degradation. These hypotheses include niche breadth, food scarcity, physiological constraints, visual constraints, increased nest failure, and acoustic and movement constraints (Canaday 1996; Stratford and Robinson 2005; Moore et al. 2008; Robinson and Sherry 2012; Rutt et al. 2019). Our study in association with previous findings (Stouffer et al. 2011; Powell et al. 2015; Wolfe et al. 2015), suggests that many sensitive birds residing in forest fragments will first exhibit increased metapopulation connectivity as the matrix regenerates, followed by the maturation of available territories within the matrix itself. Thus, the conservation value of tropical forest must not be solely based on a contemporary snapshot of available habitat, but rather the future ecological value of regenerating forest in human-dominated landscapes.
Availability of data and materials
Data used in this study are archived at the Landbird Monitoring Network of the Americas (LaMNA; http://www.klamathbird.org/lamna/).
References
Arnold TW. Uninformative parameters and model selection using Akaike’s information criterion. J Wildl Manage. 2010;74:1175–8.
Bayne EM, Hobson KA. Apparent survival of male Ovenbirds in fragmented and forested boreal landscapes. Ecology. 2002;83:1307–16.
Bierregaard R Jr. Lessons from Amazonia: the ecology and conservation of a fragmented forest. New Haven: Yale University Press; 2001.
Bierregaard RO Jr, Lovejoy TE. Effects of forest fragmentation on Amazonian understory bird communities. Acta Amaz. 1989;19:215–41.
Bierregaard RO Jr, Lovejoy TE, Kapos V, Dos Santos AA, Hutchings RW. The biological dynamics of tropical rainforest fragments. Bioscience. 1992;42:859–66.
Brown S, Lugo AE. Tropical secondary forests. J Trop Ecol. 1990;6:1–32.
Camargo JLC, Kapos V. Complex edge effects on soil moisture and microclimate in central Amazonian forest. J Trop Ecol. 1995;11:205–21.
Canaday C. Loss of insectivorous birds along a gradient of human impact in Amazonia. Biol Conserv. 1996;77:63–77.
Cooch E, White G. Program MARK: a gentle introduction. 2018. http://www.phidot.org/software/mark/docs/book. Accessed Jan 2018.
Debinski DM, Holt RD. A survey and overview of habitat fragmentation experiments. Conserv Biol. 2000;14:342–55.
Ewers RM, Didham RK. Confounding factors in the detection of species responses to habitat fragmentation. Biol Rev. 2006;81:117–42.
Gilpin ME, Soulé ME. Minimum viable populations: processes of species extinctions. In: Soulé ME, editor. Conservation biology: The science of scarcity and diversity. Sunderland: Sinauer Associates; 1986. p. 19–34.
Johnson EI, Wolfe JD. Thamnophilidae (antbird) molt strategies in a central amazonian rainforest. Wilson J Ornithol. 2014;26:451–62.
Johnson EI, Wolfe JD. Molt in Neotropical birds: life history and aging criteria. Boca Raton: CRC Press; 2017.
Kennedy CM, Marra PP, Fagan WF, Neel MC. Landscape matrix and species traits mediate responses of Neotropical resident birds to forest fragmentation in Jamaica. Ecol Monogr. 2010;80:651–69.
Laurance WF, Camargo JLC, Luizao RCC, Laurance SG, Pimm SL, Bruna EM, et al. The fate of Amazonian forest fragments: a 32-year investigation. Biol Conserv. 2011;144:56–67.
Laurance WF, Curran TJ. Impacts of wind disturbance on fragmented tropical forests: a review and synthesis. Austral Ecol. 2008;33:399–408.
Laurance SG, Stouffer PC, Laurance WF. Effects of road clearings on movement patterns of understory rainforest birds in central Amazonia. Conserv Biol. 2004;18:1099–109.
Moore RP, Robinson WD, Lovette IJ, Robinson TR. Experimental evidence for extreme dispersal limitation in tropical forest birds. Ecol Lett. 2008;11:960–8.
Murcia C. Edge effects in fragmented forests: implications for conservation. Trends Ecol Evol. 1995;10:58–62.
Powell LL, Zurita G, Wolfe JD, Johnson EI, Stouffer PC. Changes in habitat use at rain forest edges through succession: a case study of understory birds in the Brazilian Amazon. Biotropica. 2015;47:723–32.
Pradel R, Hines JE, Lebreton JD, Nichols JD. Capture-recapture survival models taking account of transients. Biometrics. 1997;71:60–72.
Rankin-de-Mérona JM, Prance GT, Hutchings RW, Silva MFD, Rodrigues WA, Uehling ME. Preliminary results of a large-scale tree inventory of upland rain forest in the Central Amazon. Acta Amazonica. 1992;22:493–534.
Remsen Jr JV, Cadena CD, Jaramillo A, Nores M, Pacheco JF, Robbins MB, et al. A classification of the bird species of South America. American Ornithologists’ Union. 2011. http://www.museum.lsu.edu/~Remsen/SACCBaseline.html. Accessed 20 Jan 2014.
Renjifo LM. Effect of natural and anthropogenic landscape matrices on the abundance of subandean bird species. Ecol Appl. 2001;11:14–31.
Robinson WD, Sherry TW. Mechanisms of avian population decline and species loss in tropical forest fragments. J Ornithol. 2012;153:141–52.
Rutt CL, Midway SR, Jirinec V, Wolfe JD, Stouffer PC. Examining the microclimate hypothesis in Amazonian birds: indirect tests of the ‘visual constraints’ mechanism. Oikos. 2019;6:798–810.
Ryder TB, Sillett TS. Climate, demography and lek stability in an Amazonian bird. P Roy Soc B-Biol Sci. 2016;283:20152314.
Saunders DA, Hobbs RJ, Margules CR. Biological consequences of ecosystem fragmentation: a review. Conserv Biol. 1991;5:18–32.
Şekercioḡlu ÇH, Ehrlich PR, Daily GC, Aygen D, Goehring D, Sandí RF. Disappearance of insectivorous birds from tropical forest fragments. Proc Natl Acad Sci USA. 2002;99:263–7.
Sodhi NS, Liow LH, Bazzaz FA. Avian extinctions from tropical and subtropical forests. Annu Rev Ecol Evol Syst. 2004;35:323–45.
Sodhi NS, Sekercioglu CH, Barlow J, Robinson SK. Conservation of tropical birds. New York: Wiley; 2011.
Stouffer PC, Johnson EI, Bierregaard RO Jr, Lovejoy TE. Understory bird communities in Amazonian rainforest fragments: species turnover through 25 years post-isolation in recovering landscapes. PLoS ONE. 2011;6:e20543.
Stouffer PC, Bierregaard RO Jr. Recovery potential of understory bird communities in Amazonian rainforest fragments. Braz J Ornith. 2007;15:219–29.
Stouffer PC, Bierregaard RO Jr. Use of Amazonian forest fragments by understory insectivorous birds. Ecology. 1995;76:2429–45.
Stouffer PC, Bierregaard RO Jr, Strong C, Lovejoy TE. Long-term landscape change and bird abundance in Amazonian rainforest fragments. Conserv Biol. 2006;20:1212–23.
Stratford JA, Robinson WD. Gulliver travels to the fragmented tropics: geographic variation in mechanisms of avian extinction. Front Ecol Environ. 2005;3:85–92.
White GC, Burnham KP. Program MARK: survival estimation from populations of marked animals. Bird Study. 1999;46:S120–39.
Wolfe JD, Johnson MD, Ralph CJ. Do birds select habitat or food resources? Nearctic-neotropic migrants in Northeastern Costa Rica. PLoS ONE. 2014;9:e86221.
Wolfe JD, Ralph CJ, Elizondo P. Demographic response of a tropical bird to the El Niño southern oscillation in mature and young forest in Costa Rica. Oecologia. 2015. https://doi.org/10.1007/s00442-015-3256-z.
Wright SJ. Tropical forests in a changing environment. Trends Ecol Evol. 2005;20:553–60.
Acknowledgements
Thanks to the multitude of banders, assistants and mateiros who helped collect the data. Logistical support from the Biological Dynamics of Forest Fragmentation Project, Brazil’s Instituto Nacional de Pesquisas da Amazônia, and the Smithsonian Institution made this research possible. Thanks to committee members and field collaborators: B. Williamson, R. Brumfield and G. Ferraz. This is publication 783 of the Biological Dynamics of Forest Fragments Project Technical Series.
Funding
Funding was provided by a US National Science Foundation (LTREB 0545491) grant and American Ornithologsts’ Union research grant.
Author information
Authors and Affiliations
Contributions
Project design was developed by ROB and TEL. Data collection was conducted by ROB, TEL, and PCS. JDW conducted the analysis. JDW, PCS, and DAL drafted the manuscript, and all authors shared in editing and revising the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research complies with the current laws of Brazil and the United States of America, and the research was approved under a Louisiana State University Institutional Animal Care and Use Committee Permit to PCS.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wolfe, J.D., Stouffer, P.C., Bierregaard, R.O. et al. Effects of a regenerating matrix on the survival of birds in tropical forest fragments. Avian Res 11, 8 (2020). https://doi.org/10.1186/s40657-020-00193-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-020-00193-x