
Abstract
Tree-related microhabitats (TreMs) have been promoted as indicators of forest biodiversity and to guide conservation practices. Ensuring the provision of diverse TreMs in the long term is crucial for the survival of many forest-dwelling species. Yet, this task is challenging in the absence of information regarding TreM dynamics. We analysed the temporal development of TreMs on 11,569 living trees in temperate European forests. To identify drivers of change in TreM abundance and richness over a period of 3–12 years, we estimated the rates of TreM persistence and loss events at the tree-level using survival analysis methods: persistence was characterised by consistency and increment events (when TreM numbers were maintained or increased) and loss was defined as a reduction in TreM numbers or their disappearance. Stratified Cox proportional hazards models were fitted for different TreM groups. Our study revealed a highly dynamic TreM development on living habitat trees, particularly on large trees. While specific TreMs are prone to disappearing, irrespective of tree species or TreM groups, total TreM richness persists over a 12-year period. TreMs such as crown deadwood, epiphytes or woodpecker cavities are prone to decrease in the long term. However, large trees were more likely to maintain a certain degree of TreM richness. Increasing diameters resulted in high persistence rates in seven TreM groups and concomitantly low loss rates in four of them (exposed sap- and heartwood, concavities). Selecting habitat trees based on TreMs should consider the likelihood of TreMs being lost over time, to ensure the long-term provision of microhabitats for associated species.
Graphical Abstract

Similar content being viewed by others
Avoid common mistakes on your manuscript.
Highlights
-
Assemblages of microhabitats remain diverse over time, especially on large trees.
-
However, certain microhabitats may disappear over time.
-
Retention of functionally different trees may secure the continuity of microhabitats.
Introduction
The retention of deadwood and habitat trees during silvicultural operations mitigates biodiversity loss in forests (Gustafsson and others 2012, 2020; Pastur and others 2020; Muys and others 2022). Following forest disturbance, biological legacies can provide a life-boating function for species associated with their structures and substrate. The retention of habitat trees (large, old, living or dead trees with microhabitats, Bütler and others 2013) aims to ensure both spatial and temporal habitat continuity that cannot be provided by deadwood (Franklin and others 2000, 2012). Tree-related microhabitats (TreMs) are often used to identify and select high-quality habitat trees and are considered useful indicators of biodiversity in European and North American forests (Asbeck and others 2021a, b; Martin and others 2022; Larrieu and others 2022; Spînu and others 2022). TreMs are defined as “distinct, well-delineated structures occurring on living or standing dead trees, that constitute a particular and essential substrate or life site for species during at least a part of their life cycle to develop, feed, shelter or breed” (Larrieu and others 2018). Some TreMs support forest-dwelling birds, rodents, bats, carnivores and invertebrates (for example, rot-holes: Gouix and Brustel 2012; cavities: Bunnell and others 2013; Remm and Lõhmus 2011). Certain TreMs are critical for the survival of specialist species (for example, dendrotelmata for tree-hole breeding mosquitos: Kitching 1917; buttress cavities for salamander species: Basile and others 2017; tree hollows for the click and hermit beetles: Ranius and others 2002; Svensson and others 2004). Based on morphological characteristics and associated taxa, a typology of TreMs has been proposed (Larrieu and others 2018, Appendix 1). This typology contains 15 different groups such as exposed sap- and heartwood or concavities, which comprise 47, more precise, types as stem and limb breakage, dendrotelms. Tree dimensions, vitality status, species, age and stand management, as well as forest conditions and forest type are all important drivers of TreM occurrence and diversity (Michel and Winter 2009; Vuidot and others 2011; Winter and others 2015; Paillet and others 2017; Asbeck and others 2019; Larrieu and others 2014; Kozak and others 2023). Large, old broadleaved and/or dead standing trees in temperate forest commonly support the richest and most abundant TreMs.
TreM inventories are now included regularly in biodiversity surveys (for example, the Index of Biodiversity Potential in France: Larrieu and Gonin 2008; Gosselin and Larrieu 2020; Germany: Zeller and others 2022), in research projects (Martin and others 2022), and educational exercises (Cosyns and others 2020; Joa and Schraml 2020) and are promoted in integrative forest management guidelines for the designation of habitat trees. Yet we have a limited understanding of the long-term implications of habitat tree selections based on TreMs.
Considering the temporal dynamics of TreMs, how their persistence and loss may affect habitat provisioning is crucial. Some TreMs are only a transitory feature of trees and thus represent ephemeral resource patches (Finn 2001) and others can persist for decades or even centuries. For example, bark pockets last until the piece of bark providing shelter drops off whereas dead branches might stay in the crown until they have sufficiently decayed to break off (Basham 1991). Likewise, large rot-holes may persist as long as the habitat tree (Larrieu and others 2022), while the life of some living TreMs, such as fruiting bodies of saprotrophic agarics is no longer than a few days. The processes and events affecting TreM dynamics can be tree or stand specific. They can occur continuously (for example, tree senescence accompanied by the capacity to compartmentalize wounds and decay), regularly (for example, silvicultural operations) or occasionally (for example, natural disturbances). While wood decay and breakage are processes occurring on all trees as they age, these events can happen at different rates as influenced by tree- and stand level attributes (Basham 1991; Mäkinen 2002; Schütz and others 2006; Kahl and others 2017; Zemlerová and others 2023). For instance, insect galleries might occur often during the lifetime of a tree and can evolve into foraging cavities that subsequently might become rot-holes. TreMs such as exposed sap- and heartwood cannot form multiple times on a tree without causing its death (for example, a dead tree top or broken main fork). TreMs might persist longer on slow-growing hardwoods with low rates of decay and breakage in comparison to many conifers and fast-growing tree species (Kõrkjas and others 2021). On the latter, TreMs usually form quicker and probably at higher rates (Kõrkjas and others 2021; Spînu and others 2023). Additionally, depending on TreM size and the tree’s capacity to inhibit the decaying fungus and form new wood, sealing the scars that are associated with certain TreMs, some tree species can fully overgrow TreMs after a few years or decades (Shigo 1984; Stoffel and Perret 2006; Smith 2015). While we do have some information on how different TreMs may develop in principle, there is little quantitative information on temporal dynamics of TreM that can guide conservation management. Given the variability in TreM dynamics, ensuring a constant, rich and diverse supply of TreMs to aid the populations of taxa dependent on them can be challenging for forest practitioners (Courbaud and others 2022; Larrieu and others 2022).
While some studies have investigated the persistence of tree cavities, hollows or wounds affecting timber quality (Edworthy and others 2012; Lindenmayer and Wood 2010; Wesołowski and others 2011, 2012; Tavankar 2017, 2022), there is a lack of comprehensive longitudinal studies on the temporal dynamics of TreMs (Puverel and others 2019). This gap may be attributed to the scarcity of repeated inventories that assess TreMs on individual trees. Longitudinal data can help to understand the development of TreMs over time, which can inform an effective selection of habitat trees that ensures continuous provision of ecological niches.
Substantial progress in understanding temporal dynamics of TreMs has been made through a method developed by Courbaud and others (2017). TreM formation rates were estimated based on TreM presence on living trees of different diameters. Studies using this adaptation of the survival and reliability theory, where increasing diameters in different trees are thought to represent tree development over time, indicated that TreMs accumulate during tree growth (Courbaud and others 2017, 2022; Jahed and others 2020). However, the relationship between diameter, age and TreM occurrence may not be as direct and linear, as assumed, as both variables were shown to have distinct effects on TreM dynamics (Kozak and others 2023). This approach also assumes that TreMs never disappear from trees and trees never die, which does not reflect the reality of ecological processes. To overcome the limitations of this design in the evaluation of time-dependent outcomes, it is crucial to complement it with studies that have repeatedly recorded TreMs on the same trees (Lindenmayer and others 2011; Courbaud and others 2017; Asbeck and others 2023).
In this study, we merged different datasets with longitudinal TreM surveys from several regions in Europe. Following previous efforts (Courbaud and others 2017, 2020), we initiated a comprehensive investigation into the temporal development of microhabitats on living habitat trees, more likely to persist over the course of multiple inventories than dead trees. Once formed on a living tree, a TreM can either remain or be lost over a certain time interval. To quantify these two main trends, we assessed changes in TreM richness for each TreM group over time. The change was defined as one of the following four event types, which are exclusive (do not happen at the same time on one individual tree):
(1) TreM increase (an increase in TreMs), (2) TreM consistency (no change occurs over time). These two event types were summarized as TreM persistence; (3) TreM reduction (a decrease in TreMs), (4) TreM disappearance (the final number of TreMs is zero). These two event types were summarized as TreM loss.
We hypothesized that:
-
a.
On living trees, the processes of increase and consistency outweigh reduction and disappearance events of TreMs, as TreMs are expected to accumulate on trees over time.
-
b.
Temporal TreM dynamics (persistence and loss rates) differ among tree species and across TreM groups. Owing to the growth dynamics and low decay compartmentalization capacity of many European broadleaved species, we expect them to have a greater TreM turnover than in conifers (with both persistence and loss events occurring more frequently). Moreover, certain TreMs such as exposed sapwood and crown deadwood are probably short-lived and can decrease over time, while rot-holes, exposed sap- and heartwood are expected to persist over repeated inventories.
-
c.
Temporal dynamics of TreMs are influenced by presence of certain TreMs at the time of the first survey, such as fresh exudates, insect galleries. These TreMs may indicate tree vitality and thus the potential for development of subsequent TreMs.
-
d.
Tree size is a significant driver of TreM dynamics and rates of both TreM persistence and TreM loss are highest on large trees.
Material and Methods
Data Origin and Harmonization of TreM Classifications for Living Trees
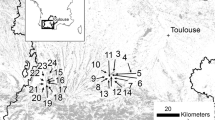
Our study focused on the temporal TreM development on trees which were alive at the time of two subsequent surveys, with repeated TreM observations on 11,569 living trees belonging to 30 tree species. Longitudinal TreM data were more common on living trees than dead ones (Appendix 2a). In addition, TreM profiles differ significantly between dead and living trees (Spînu and others 2022) and targeting the latter ones facilitated a focused data interpretation and reduced possible errors from the data harmonization. The surveys concerned all trees, whether or not they bore TreMs initially. We used the availability of unique datasets with repeated TreM surveys for an extensive study on temporal TreM development. These data were collected by three research and natural area management groups from different regions in Europe: France—Cateau and others 2024; Germany—Spînu and others 2022; Slovakia/Romania—Kozak and others 2023 (Figure 1). Each group used its own TreM description and field protocols (for examples, different size thresholds) and older inventories were carried out only at TreM group level. Despite this, we were able to harmonize the data at the level of TreM groups based on a common typology (Larrieu and others 2018). We only retained TreM types that appeared in each classification and assigned them to 15 broader TreM groups proposed by Larrieu and others (2018), following a similar approach as other previous studies (Asbeck and others 2021b; Courbaud and others 2022). The TreM groups included: woodpecker cavities, rot-holes, insect galleries and bore holes, concavities, exposed sapwood and heartwood, crown deadwood, twig tangles, burrs and cankers, perennial and ephemeral fungal fruiting bodies, epiphytic or parasitic crypto- and phanerogams, nests, microsoils and fresh exudates. Specifics of the study areas, individual classifications and corresponding TreM groups are provided in Appendix 1.

Location of the study areas.
The study plots were located in both managed and unmanaged mature stands, within a wide range of forest types. Most of the studied Central European sites were not affected by major disturbances, for example no major windthrows occurred between 2005 and 2020 in France. The rates of tree mortality in the period between the surveys in Germany were low in comparison with other European regions (Spînu 2023). In general terms, disturbances play an important role in shaping the structure and composition of primary forests from the Carpathian region, but few sites have been disturbed over the period covering the surveys used in our analysis (Janda and others 2017; Meigs and others 2018; Zemlerová and others 2023).
Owing to the small number of repeated observations in most TreM groups and of some tree species, the analyses could only be carried out at the level of tree species category (broadleaves—57.6% and conifers—42.4%), but not for individual tree species. Details on all tree species and an overview of the distribution of TreM groups at tree level, for both surveys can be found in Appendix 2 and 3).
TreMs were inventoried between 2006 and 2021, with each living tree being surveyed twice within that interval. The time interval between the first and the second survey, later called survey period, ranged between 3 and 12 years and differed between the different regions and studies (Table 1). The diameters at breast height (DBH at 1.3 m) of habitat trees varied between 6 and 129 cm for broadleaves and 8 and 142 cm for conifers. The trees were located within a wide altitudinal range between 120 and 2018 m.a.s.l. European beech (Fagus sylvatica (L.), 33.9%), Norway spruce (Picea abies (L.), 19.5%) and silver fir (Abies alba (Mill.), 17.6%) were the main species. The main characteristics of TreM groups and corresponding living habitat trees are summarized in Table 2.
We conducted several data analyses to understand how temporal dynamics of TreMs on living habitat trees change over the course of a 12-year period. Firstly, we used a random forest analysis to describe overall TreM patterns. By means of survival analysis methods, we analyse the dynamics of TreMs on habitat trees to address hypotheses a, b and d. Additionally, a second random forest analysis was conducted to assess whether changes in TreM development were related variables describing living habitat trees at the first survey, such as DBH, altitude, and presence of specific TreM groups, in line with hypotheses c and d.
Description of the TreM Data
Habitat trees were defined as trees exhibiting the presence of at least one TreM at the first survey. The number of habitat trees was 4145, representing 35% of the total number of 11,569 living trees included in the study. The TreM counts followed a right-skewed distribution in each TreM group. The richness of TreMs for each tree and survey was calculated as the sum of TreM groups present on a tree (for example, crown deadwood, exposed sapwood, cavities). To gain a first impression of the data, we drew alluvial plots to visualise the number of TreMs at each survey. Additionally, a random forest analysis was conducted to assess whether the presence of certain TreM groups predicted the overall TreM richness at both surveys (Breiman 2001). The independent variables considered in the analysis included tree DBH, altitude and presence of TreM groups. For the first survey, the predicted variables were log-transformed to improve the fit. However, log-transformation was not possible for the second survey due to presence of zero counts. We ran 1000 bootstrap samples and optimized the number of splits for each random forest on habitat trees, stratified by survey. This non-parametric approach allowed for the modelling of non-linear effects and complex interactions between predictors without making prior assumptions about the data. To assess the quality of predictions, we report variable importance lists, percentages of explained variance, and prediction errors. For this purpose, variables were ranked in terms of importance based on their impact in predicting the outcome (Appendices 5, 6). A variable was considered more important than another, if its removal from the prediction yielded an error larger than errors obtained through the removal of other variables. We conducted an additional random forest analysis to assess whether the change in TreM development related to variables describing living habitat trees at the first survey, such as DBH, altitude, and frequency of specific TreM groups. Similar to the methods used to describe the TreM data, we identified those TreM groups that predicted certain changes in TreM richness. Here, change was defined as the difference in TreM richness between the surveys.
Prediction and Modelling of TreM Dynamics
We used survival analysis methods to analyse the dynamics of TreMs on habitat trees, for both total TreM richness, as well as separately for each TreM group. The focus of our study was on habitat trees, trees which exhibited at the first survey at least one TreM (to model the presence of multiple TreM groups by means of TreM richness) or a specific TreM group (to model at each TreM group level). We defined two main patterns of TreM development, consisting of mutually exclusive events that can be referred to as competing risks. First, TreM persistence encompassed events where the number of TreMs remained the same (consistency events) or increased over time (increment events). Second, TreM loss represented events where the number of TreMs decreased (reduction events), potentially resulting in complete absence (disappearance events).
Further, we defined the time-to-event as the time of occurrence of an event and the type represents one of the four development events (consistency, increment, reduction, disappearance). The outcome was the combination of time-to-event (denoted by T) and type of event (denoted by D) variables, further referred to as (T, D). The independent variables were tree DBH, altitude and the TreMs count at first survey (jointly denoted by Z and referred as the covariate vector). Our aim was to model and estimate the type-specific cumulative incidence functions of (T, D):
where P stands for probability and j stands for any event type out of the four events described above. The standard way of estimating is through the type-specific hazard rates:
We defined the survival function in the following way:
We fitted Cox proportional hazards models to each type-specific hazard. For a tree with the covariate vector Z, the type-j specific hazard was modelled as:
where \({\lambda }_{j,0}\)(t) is the baseline type-specific hazard and the vector \({\beta }_{j}\) represents the covariates effects on type-j hazard. Λ(u) represents the cumulative hazard function for the jth event at the time u. Tied events were dealt with by means of the Breslow correction.
Owing to the low number of burrs and cankers, insect galleries, and nests in the data set (< 20 observations), fitting Cox proportional hazards models was not adequate for these TreM groups. All four types of events were recorded only in two TreM groups: crown deadwood and fresh exudates. Other TreM groups exhibited no or a low number TreM reduction event. If the number of events was too low (< 20), the analysis was carried out at the level of the two main development patterns, TreM persistence and loss.
To overcome the issue of non-proportionality, we fitted stratified Cox proportional models at the hazard level. This was based on an exploratory analysis using the non-parametric Kaplan–Meier estimator, stratified by either tree categories or research sites. This approach implied that no effect was estimated for the stratifying variables. Instead, the difference in the data source (for example, survey intervals) or between tree categories (broadleaves, conifers) was measured at the baseline survival level by allowing the estimation of survival curves to mold to each stratum (Appendix 4). Since twig tangles and fresh exudates were not recorded in older surveys, their average study interval of 3 years was the shortest of all TreM groups. The exact occurrence of events on a tree was likely not recorded at the exact time of occurrence but at the end of the inventory interval, during which it occurred. The scope of the analysis was to observe general patterns of change, as our data and methods do not allow for the identification of the point in time when a change occurred.
All analyses were performed in R version 4.2.0 (R Core team 2021), using the randomForest (Liaw and Wiener 2002), survival (Therneau 2023) packages and represented graphically through theggplot2 (Wickham 2016) package.
Results
Data Description
The top predictors for the richness of TreM groups at the first survey were fresh exudates, microsoils, crown deadwood, concavities, exposed sap- and heartwood, and annual fungi, in that order. At the second survey, the important predictors were annual fungi, exposed sapwood only, crown deadwood, perennial fungi, epiphytes, exposed sap- and heartwood and twig tangles. The percentage of explained variance for this random forest analysis reached 97.7% for the TreM richness in the first survey, and 96.3% in the second survey (Appendix 5). While some TreMs remained significant predictors of richness over time (crown deadwood, exposed sap heartwood, annual and perennial fungi), the ranking of other TreM groups decreased (microsoils, fresh exudates, concavities), being replaced by others TreM groups, such as exposed sapwood or epiphytes (Appendix 6). This observation was in line with the patterns provided by alluvial plots, which showed that these top predictors were also the TreM groups with highest frequency of occurrence (Appendix 3). The differences in TreM occurrence between surveys indicated a trend of loss at the tree level for most TreM groups (Appendix 3).
Changes in TreM Richness at the Tree-Level
Throughout the study period, the incidences of TreM persistence events (increase and consistency) and loss events (reduction and disappearance) increased to reach a plateau (Figure 2, Appendix 7). The TreM richness showed similar cumulative incidence rates for persistence and loss events, irrespective of tree species group. Approximately 30% of the living habitat trees experienced consistency or increase events. Loss events accounted for approximately 40% of all events, while persistence represented 60% (Figure 2). The positive difference in frequency between persistence and loss events was higher in broadleaves than in conifers, with a 20% difference in cumulative incidence by the end of the survey period.

Cumulative incidence rates for all types of all events in the richness of TreMs at both type and group level, for each habitat tree since the first survey. Persistence events (increment, consistency) are marked in dark and light orange, while loss events (reduction, disappearance) are indicated in light and dark blue.
Temporal Changes at the TreM Group-Level
In contrast to TreM richness, losses represented the vast majority of events observed within each TreM groups, both on conifer and broadleaved habitat trees (Figure 3). Reduction and disappearance events exhibited the highest cumulative incidence rates throughout the survey period and showed the fastest increase in rates over time. On average, there was a cumulative incidence of over 70% in the sample population, irrespective of tree category, to experience a loss in a TreM group by the end of the survey period. Although the cumulative rates of loss varied among TreM groups, ranging from the lowest values for concavities (52%) to the highest for twig tangles (88%), losses were consistently observed in all groups. Crown deadwood, perennial fungi, and fresh exudates were the least likely to be lost on conifers (Table 3, Figure 3).

Cumulative incidence rates for all types of events in different TreM groups, since the first survey. Dashed lines represent events combined at the level of the two main development patterns due to the low number of recordings at event type level. Persistence processes (increment and consistency events) are represented in orange shades. Loss processes (reduction and disappearance events) are shown in blue shades.
Increment and consistency events were generally rare for most TreM groups, with a cumulative incidence of less than 10% and 20% respectively (Table 3, Figure 3) by the end of the survey period. The only TreM group that showed a slightly higher rate of increment were concavities in broadleaves, with an incidence rate of 40% by the end of the survey period. However, woodpecker cavities, rot-holes, exposed sap- and heartwood (broadleaved trees), perennial fungi (conifers), epiphytes, and concavities (both broadleaves and conifers) were groups that primarily experienced consistency, with an approximate incidence rate of 20% (Table 3, Figure 3).
Sufficient sample sizes to distinguish reduction and disappearance events were available for two groups, namely crown deadwood and fresh exudates. The disappearance events showed incidence rates almost six times higher than TreM reduction by the end of survey period (Table 3, Figure 3).
Drivers of TreM Development
The results of the stratified Cox proportional hazard models indicated that the presence of certain TreMs at the first survey had an impact on hazard rates for five TreM groups. Specifically, the occurrence of exposed sap- and heartwood, crown deadwood, rot-holes, fresh exudates, and twig tangles at the beginning of the study increased the hazard rates for loss events. On the other hand, the presence of fresh exudates at the first survey had a negative effect on consistency and reduction events (Table 3). The second random forest analysis further supported the importance of rot-holes, fresh exudates, and crown deadwood as predictors of the temporal changes in TreM richness (Appendix 6).
Tree DBH was significantly associated with at least one type of event in most TreM groups, except for woodpecker cavities, epiphytes, and crown deadwood (Table 3). Increasing tree DBH was associated with higher rates of persistence events (mostly increment) in seven TreM groups and overall TreM richness, while simultaneously leading to lower rates of loss events in three of those groups. The rates of both persistence and loss events increased with tree DBH for twig tangles and overall TreM richness. Tree DBH emerged as an important driver for loss events of exposed sap- and heartwood, perennial fungi, concavities, fresh exudates, and twig tangles; with a negative effect in the first four categories and a positive effect in the last one (Table 3).
Altitude was associated with at least one type of event in four TreM groups and overall TreM richness. Higher altitude was linked to lower rates of loss in woodpecker cavities, exposed sap- and heartwood, epiphytes, and to lower rates of persistence for rot-holes. All events related to TreM richness showed lower rates with increasing altitude (Table 3).
Discussion
We studied the temporal dynamics of TreMs on a dataset comprising 11,569 living habitat trees distributed over four countries in Europe. Our study shows that TreM development on living habitat trees is highly dynamic, particularly on large trees, as tree dimension emerged as a driver of both TreM loss and persistence. The results indicate that, while TreM richness can persist in the long term, specific TreMs are prone to disappear on individual trees, irrespective of tree species or TreM group. Additionally, the presence of rot-holes, fresh exudates and crown deadwood on living trees could serve as indicators of subsequent changes in their TreM profile.
Balance Between TreM Persistence and Loss at the Tree-Level
We showed that the overall TreM richness remained stable over a period of 12 years at least. The processes of increment and consistency did not outweigh reduction and disappearance of TreMs, regardless of the tree species (Figure 2). However, broadleaved trees exhibited relatively higher rates of TreM increment compared to their loss. The resulting increase can be attributed to the morphological and wood characteristics. Species such as F. sylvatica (the dominant broadleaved species in our study) have a hard xylem protected by a thin bark, in comparison to P. abies thicker bark that protects a more flexible, softer xylem. This makes them more sensitive to mechanical impacts such as those caused by rockfalls, and lower capacity to seal wounds compared to species such as P. abies (Trappmann and Stoffel 2013). Additionally, the wood of F. sylvatica decays faster than that of P. abies (Kahl and others 2017). Thus, once formed, TreMs that are related to injuries or facilitate wood decay may lead to the subsequent formation of new TreMs. For instance, the formation rate of TreMs on F. sylvatica was found to be twice as high compared to conifers such as A. alba (Courbaud and others 2017).
However, the changes in the TreM data between surveys indicated a general balance between the formation of new TreMs and the loss of existing ones. For example, losses in fresh exudates and concavities were compensated by gains in exposed sapwood and epiphytes. The occurrence of living epiphytes can increase over time as a tree ages and its surface area expands, especially under favourable growth and dispersal conditions for bryophytes or lichens (Lie and others 2009; Asbeck and others 2021a, b; Kõrkjas and others 2021). Crown deadwood and exposed sap- and heartwood, which include dead branches of large diameters, dead tops, cracks and fissures, were present in similar shares at both surveys. The longevity of dead branches on decay-resistant tree species such as P. menziesii (Mirb.) Franco and P. sylvestris has been observed in previous studies. Records show that indeed dead branches of those species can persist in the canopy for up to 10, and 6 years, respectively (Kahl and others 2017; Mäkinen 2002). Dead branches may persist even longer in broadleaved species such as Quercus, yet such data is scarce. TreMs associated with exposed sap- and heartwood, such as broken tops or forks are less likely to disappear or occur repeatedly on a living tree without causing its death (Kõrkjas and others 2021).
Certain TreM Groups are Prone to Decline or Disappear in the Long Term
Our results suggest that all TreM groups on living habitat trees are susceptible to long-term loss, regardless of tree species. The patterns of loss varied in frequency and incidence rates, but by the end of the study, more than half of the habitat trees experienced losses in their TreM groups. The lowest rates of loss were found among crown deadwood, perennial fungi, and exposed sapwood on living conifers. This observation may be attributed to new occurrences of those TreMs, as responses of conifers to stress caused by recent episodes of drought events and bark beetle outbreaks (Senf and others 2018; Jakoby and others 2019). This pattern can also explain why TreMs such as crown deadwood, fresh exudates, and rot-holes were observed to predict future changes in TreM richness at the tree level. Crown dieback, increased resin production and bark loss are indicators of declining tree vitality triggered by stress associated with recent extreme drought and subsequent bark-beetle outbreaks (Bouget and others 2019). Often, trees infested by bark beetles are colonised by perennial fungi such as Fomitopsis pinicola (Vogel and others 2017). These changes in tree vitality, characterised by elevated rates of wood decay and breakage, likely have an impact on TreM composition profile.
However, the underlying causes of apparent TreM loss are likely to vary among different TreM groups (see Table 4). Previous studies have examined the temporal development of woodpecker cavities, rot-holes, fire scars, as well as TreMs that affect timber quality (for example, Wesołowski 2011, 2012; Edworthy and others 2012; Tavankar and others 2017, 2019). The drivers of the temporal changes, particularly loss, across all TreM groups remain to be identified. While the loss of certain TreMs might seem straightforward and intuitive (for example, structures that naturally fall off or decay over time, such as crown deadwood or ephemeral fungi), the drivers behind loss processes can be diverse. For instance, the changes in TreMs linked to presence and activities of specific forest-dwelling species could be manifold (for example, woodpecker cavities or rot-holes). Cavities utilised by vertebrates, for example, are rich in nitrogen and attract invertebrate colonization, which may further excavate galleries, provide access for other organisms, and eventually lead to wood breakage and decay (Gibbons and Lindenmayer 2002; Siiton and Jonsson 2012). Concurrently, tree defence mechanisms may compartmentalise injuries by inhibiting wood-decaying fungi and sealing wounds, often associated with TreMs (Smith 2015). All the subsequent processes can alter the decay dynamics and shape the development of TreMs, eventually leading to their decline or disappearance.
Cavities have been reported to last approximately 10 years (Edworthy and others 2012; Edworthy and Martin 2013), with slightly shorter rates for excavated cavities than for non-excavated ones (Wesołowski 2011, 2012). Our findings align with these observations, as approximately 60% of the broadleaved trees and 80% conifers experienced loss of woodpecker cavities over the 12-year study period. The loss of woodpecker cavities on living trees could be attributed to several biological drivers, such as breakage of branches and trunks with cavities, destruction by predators, decay and collapse of the walls, closure through growth of callus tissues (Wesołowski and Martin 2018). Similar causes can lead to loss of other TreMs too. For example, while large wounds may persist for up to 17 years, exposed wood patches and wounds of smaller sizes may be sealed successfully after 10 years depending on the extent of xylem and phloem injury (Vasaitis and others 2012; Tavankar and others 2017, 2019). The masking of wounds can contribute to the loss of TreMs, with a rate varying among tree species, with P. abies trees compartmentalising and closing scars at a faster rate than A. alba or F. sylvatica (Stoffel and others 2005; Trappmann and Stoffel 2013). A detailed overview of possible drivers of TreM loss among all groups is presented in Table 4.
Larger Trees have Highly Dynamic TreM Development Profile
Tree size played a significant role in the TreM development over the course of 3–12 years. In accordance with previous studies (Asbeck and others 2019; Paillet and others 2019; Courbaud and others 2022; Kozak and others 2023), our results support the notion that TreM composition becomes more diverse on trees during their growth. This phenomenon is attributed to the accelerated formation of new TreMs together with the accumulation of TreMs over time (Paillet and others 2019; Asbeck and others 2021a, b; Courbaud and others 2022). In our study, tree DBH appeared as an important driver of both TreM loss and persistence. The stability of TreMs on large trees was evident, as indicated by higher persistence rates of half of the TreM groups compared to smaller trees. Specifically, large trees lose less exposed sap- and heartwood, perennial fungi, concavities, and fresh exudates. This can be attributed to the ability of large trees to withstand breakage from wind or snow more successfully than smaller trees (Bragg and others 2002), resulting in fewer losses of TreMs specific to breakage processes (stem and limb breakage, cracks and splits). The presence of fresh exudates and perennial fungi on large trees may be linked to their susceptibility to stress induced by recent droughts and subsequent colonization by wood decaying fungi (Das and others 2016; Pfeifer and others 2011). However, the dynamics of TreMs on very large and old trees, reaching the stage of senescence, might show contrasting patterns due to advanced decay and breakage, important processes involved in TreM formation and drivers of TreM persistence (Edworthy and others 2012; Kõrkjas and others 2021). In our study, it was not possible to describe TreM dynamics on very large, old trees (over 80 cm in DBH) only, of which we had only a limited number in the data set.
Study Limitations and Research Needs
Our study allowed us to capture the manifold events involved in TreM development on living habitat trees across different European forest regions. However, our findings may be complemented by future studies, at finer temporal scale. Our analysis was likely influenced by different survey intervals as the re-survey of the TreMs was conducted to align with specific project objectives rather than capturing changes in TreMs. Thus, we could not record precisely the moment in time when an event occurred. The variations in survey periods from 3 to 12 years also limited site-specific comparisons and the evaluation of the effect of stand management, forest types, and growing conditions on TreM dynamics. Additionally, our analysis could capture only the dynamics of TreM groups. The stability in the TreM richness suggests increases in numbers of individual TreMs within each TreM group. A targeted TreM study, dedicated to quantify temporal changes in TreMsat tree species level, could better capture the underlying causes of TreM persistence and loss. The use of different, older typologies and subsequent simplifications as well as the observer bias are likely sources of inaccuracies in our study (Paillet and others 2015). Nowadays, harmonized TreM typologies are widely available (Kraus and others 2016; Larrieu and others 2018). Thus, future studies may be able to provide more accurate and detailed insights into the development of specific TreMs and allow better understanding of how TreMs evolve, how they influence each other and develop into other TreMs. While the length of this study was sufficient to observe changes in many TreMs, longer periods between the surveys would allow the study of longer-lived microhabitats, such as rot-holes and exposed heartwood structures of large dimensions. Such information can further be implemented in dynamic forest models (Courbaud and others 2022).
We described TreM loss and persistence only on living habitat trees, exhibiting at least one TreM in the first survey. The decision to work only with living trees improved the statistical power by avoiding problems of zero-inflated data. However, we could not capture the formation of TreMs on individuals with no TreMs, nor describe how TreMs evolve after tree death. Certain TreMs, such as broken stems, forks, hollows, large areas or bark loss can affect tree vitality and potentially lead to subsequent tree mortality (Siitonen 2012). Because TreM composition and diversity differ between living and dead trees (Paillet and others 2019; Spînu and others 2022), we suggest further investigations on the dynamics of individual TreMs after tree death. This is particularly important in the context of increasing forest disturbances and high tree mortality rates (Seidl and others 2017), which shape the occurrence of microhabitats on trees (Zemlerová and others 2023), and likely their temporal development.
Our data contained recorded losses in certain TreM groups that are unlikely to disappear naturally or increase over time on living trees, such as large rot-holes, stem and limb breakages, dead tops. It is thus essential to acknowledge the possibility of observer errors, which could have resulted in incorrect assessments of losses or increments in TreM occurrence. Certain TreMs might have been overlooked during the initial or subsequent survey, leading to inaccuracies in our data.
In addition, while certain TreMs may be easily recovered due to quick and frequent formation, or even through artificial restoration, the loss of others such as large rot-holes, holds more significant implications for the species they host and need suitable management solutions. However, because we studied TreM dynamics only at the individual tree level, we cannot say how the overall provision of different microhabitats develops over time at the level of forest stands, where TreM dynamics are shaped by tree growth, changes in tree vitality and disturbances.
The retention of old, large trees with abundant and diverse TreMs has been recognised as a high-priority conservation strategy for enhancing the conservation value of European forests (Larsen and others 2022). However, such trees are often scarce in managed European forests and there may be a need to retain trees without TreMs and allow for TreM development over time (Larrieu and Cabanettes 2012; Asbeck and others 2019; Vandekerkhove and others 2018). Simulation studies have also suggested that retention of trees with no TreMs, particularly of short-lived tree species, can contribute to maintaining a continuous long-lasting habitat pool (Fan and others 2004; Courbaud and others 2017). For future studies, it would be valuable to investigate the changes in TreMs on trees with various attributes, including different species, senescence stages, presence of decay, or other factors related to TreM development (Edworthy and others 2012; Larrieu and others 2022).
Conclusions and Implications for Future Forest Management
So far, the selection of habitat trees has focused on their current TreM occurrence without quantifying the impact of this practice for future habitat provisioning. Our findings show that although TreM richness was persistent at the tree level, TreMs in all groups were susceptible to disappearing in the long term. We therefore suggest a diversified approach to habitat tree selection in order to secure a continuous supply of resources in the future. One key suggestion is to prioritise selection of trees with rare TreMs and additionally retain functionally different trees that provide diverse TreMs (for example, different tree species or vitality status). Biodiversity conservation can be promoted from the early stages of forest development and trees with unusual shapes or defects that would normally be removed, could be retained and allowed to develop into habitat trees. Further, the high rates of TreM losses over a 12-year period suggest that revisiting sites at regular intervals and monitoring TreM development of habitat trees might aid conservation efforts. Such practices are time consuming though and should be encouraged through financial incentives for retention of habitat trees.
Data availability
Data analysed during this study are provided in full within the published article.
References
Asbeck T, Großmann J, Paillet Y, Winiger N, Bauhus J. 2021a. The Use of Tree-Related Microhabitats as Forest Biodiversity Indicators and to Guide Integrated Forest Management. Current Forestry Reports. https://doi.org/10.1007/s40725-020-00132-5.
Asbeck T, Pyttel P, Frey J, Bauhus J. 2019. Predicting abundance and diversity of tree-related microhabitats in Central European montane forests from common forest attributes. Forest Ecology and Management 432:400–408. https://doi.org/10.1016/j.foreco.2018.09.043.
Asbeck T, Kozák D, Spinu AP, Mikoláš M, Zemlerová V, Svoboda M. 2021b. Tree-related microhabitats follow similar patterns but are more diverse in primary compared to managed temperate mountain forests. Ecosystems. https://doi.org/10.1007/s10021-021-00681-1.
Asbeck T, Benneter A, Huber A, Margaritis D, Buse J, Popa F, Pyttel P, Förschler M, Gärtner S, Bauhus J. 2023. Enhancing structural complexity: An experiment conducted in the Black Forest National Park, Germany. Ecology and Evolution 13(1):e9732. https://doi.org/10.1002/ece3.9732.
Basham JT. Stem decay in living trees in Ontario’s forests: A users’ compendium and guide. Information report No. O-X-408 (Technical Report) | ETDEWEB. (1991). Canada
Basile M, Romano A, Costa A, Posillico M, Scinti Roger D, Crisci A, Raimondi R, Altea T, Garfì V, Santopuoli G, Marchetti M, Salvidio S, De Cinti B, Matteucci G. 2017. Seasonality and microhabitat selection in a forest-dwelling salamander. The Science of Nature 104(9–10):80. https://doi.org/10.1007/s00114-017-1500-6.
Bässler C, Cadotte MW, Beudert B, Heibl C, Blaschke M, Bradtka JH, Langbehn T, Werth S, Müller J. 2016. Contrasting patterns of lichen functional diversity and species richness across an elevation gradient. Ecography 39(7):689–698. https://doi.org/10.1111/ecog.01789.
Bilgili E, Coskuner KA, Baysal I, Ozturk M, Usta Y, Eroglu M, Norton D. 2020. The distribution of pine mistletoe (Viscum album ssp. austriacum) in Scots pine (Pinus sylvestris) forests: from stand to tree level. Scandinavian Journal of Forest Research, 35(1–2), pp. 20–28. https://doi.org/10.1080/02827581.2020.1729402
Bragg DC, Shelton MG, Zeide B. 2003. Impacts and management implications of ice storms on forests in the southern United States. Forest Ecology and Management 186(1):99–123. https://doi.org/10.1016/S0378-1127(03)00230-5.
Breiman L. 2001. Random Forests. Machine Learning 45:5–32.
Bunnell FL. 2013. Sustaining Cavity-Using Species: Patterns of Cavity Use and Implications to Forest Management. ISRN Forestry 2013:1–33. https://doi.org/10.1155/2013/457698.
Buse J, Ranius T, Assmann T. 2008. An endangered longhorn beetle associated with old oaks and its possible role as an ecosystem engineer. Conservation Biology 22(2):329–337. https://doi.org/10.1111/j.1523-1739.2007.00880.x.
Bütler R, Lachat T, Larrieu L, Paillet, Y. 2013. Habitat trees: key elements for forest biodiversity. In Kraus D, Krumm F, Eds. Integrative approaches as an opportunity for the conservation of forest biodiversity. Joensuu: European Forest Institute. p 284.
Cateau, E., Debaive, N., Drapier, N., Chantreau, F., Gilg, O., Laroche, F., Morin, X., Demets, V., Pimenta, R., Thompson, L., Paillet, Y. 2024. Tree inventory data from permanent plots in French forest reserves. Ecology. (Accepted)
Cockle KL, Martin K, Bodrati A. 2017. Persistence and loss of tree cavities used by birds in the subtropical Atlantic Forest. Forest Ecology and Management 384:200–207. https://doi.org/10.1016/j.foreco.2016.10.052.
Cornwell WK, Cornelissen JH, Allison SD, Bauhus J, Eggleton P, Preston CM, Scarff F, Weedon JT, Wirth C, Zanne AE. 2009. Plant traits and wood fates across the globe: rotted, burned, or consumed? Global Change Biology, 15(10), pp. 2431–2449. https://doi.org/10.1111/j.1365-2486.2009.01916.x
Cosyns H, Joa B, Mikoleit R, Krumm F, Schuck A, Winkel G, Schulz T. 2020. Resolving the trade-off between production and biodiversity conservation in integrated forest management: Comparing tree selection practices of foresters and conservationists. Biodiversity and Conservation 29(13):3717–3737. https://doi.org/10.1007/s10531-020-02046-x.
Courbaud B, Larrieu L, Kozak D, Kraus D, Lachat T, Ladet S, Müller J, Paillet Y, Sagheb-Talebi K, Schuck A, Stillhard J, Svoboda M, Zudin S. 2022. Factors influencing the rate of formation of tree-related microhabitats and implications for biodiversity conservation and forest management. Journal of Applied Ecology 1365–2664:14068. https://doi.org/10.1111/1365-2664.14068.
Courbaud B, Pupin C, Letort A, Cabanettes A, Larrieu L. 2017. Modelling the probability of microhabitat formation on trees using cross-sectional data. Methods in Ecology and Evolution 8(10):1347–1359. https://doi.org/10.1111/2041-210X.12773.
Das AJ, Stephenson NL, Davis KP. 2016. Why do trees die? Characterizing the drivers of background tree mortality. Ecology 97(10):2616–2627. https://doi.org/10.1002/ecy.1497.
Edworthy AB, Wiebe KL, Martin K. 2012. Survival analysis of a critical resource for cavity-nesting communities: Patterns of tree cavity longevity. Ecological Applications 22(6):1733–1742. https://doi.org/10.1890/11-1594.1.
Edworthy AB, Martin K. 2014. Long-term dynamics of the characteristics of tree cavities used for nesting by vertebrates. Forest Ecology and Management 334:122–128. https://doi.org/10.1016/j.foreco.2014.09.001.
Evans J. 1985. The control of epicormic branches. In Advances in Practical Arboriculture. D. Patch (ed.). Forestry Commission Bulletin 65. HMSO, London, pp. 115–120.
Fan Z, Shifley SR, Thompson FR, Larsen DR. 2004. Simulated cavity tree dynamics under alternative timber harvest regimes. Forest Ecology and Management 193(3):399–412. https://doi.org/10.1016/j.foreco.2004.02.008.
Fedrowitz K, Koricheva J, Baker SC, Lindenmayer DB, Palik B, Rosenvald R, Beese W, Franklin JF, Kouki J, Macdonald E, Messier C, Sverdrup-Thygeson A, Gustafsson L. 2014. REVIEW: Can retention forestry help conserve biodiversity? A Meta-Analysis. Journal of Applied Ecology 51(6):1669–1679. https://doi.org/10.1111/1365-2664.12289.
Finn JA. 2001. Ephemeral resource patches as model systems for diversity-function experiments. Oikos 92(2):363–366.
Franklin JF, Lindenmayer D, MacMahon JA, McKee A, Magnuson J, Perry DA, Waide R, Foster D. 2000. Threads of Continuity. There are immense differences between even-aged silvicultural disturbances (especially clearcutting) and natural disturbances, such as windthrow, wildfire, and even volcanic eruptions. Conservation in Practice, 1(1), 8–17. https://doi.org/10.1111/j.1526-4629.2000.tb00155.x
Fritz Ö, Heilmann-Clausen J. 2010. Rot holes create key microhabitats for epiphytic lichens and bryophytes on beech (Fagus sylvatica). Biol. Conserv. 143(4):1008–1016. https://doi.org/10.1016/j.biocon.2010.01.016.
Gibbons P, Lindenmayer D. 2002. Tree hollows and wildlife conservation in Australia. Clayton: CSIRO Publishing.
Gosselin F, Larrieu L. 2020. Developing and using statistical tools to estimate observer effect for ordered class data: The case of the IBP (Index of Biodiversity Potential). Ecol. Indic. 110:105884. https://doi.org/10.1016/j.ecolind.2019.105884.
Gossner MM. 2018. A three year study of the phenology of insect larvae (Coleoptera, Diptera) in water-filled tree holes in the canopy of a beech tree. Eur J Entomol. https://doi.org/10.14411/eje.2018.052.
Gouix N, Brustel H. 2012. Emergence trap, a new method to survey Limoniscus violaceus (Coleoptera: Elateridae) from hollow trees. Biodivers. Conserv. 21(2):421–436. https://doi.org/10.1007/s10531-011-0190-1.
Günther E, Hellmann M. 2005. Development and new tenants of holes of spotted woodpeckers (Dendrocopos) in the ‘“Swift-forest”’ in the Harz Mountains (Sachsen-Anhalt). Results of twenty years investigations of the use of natural tree holes. Orn. Jber. Mus. Heineanum 23:103–122. in German.
Gustafsson L, Baker SC, Bauhus J, Beese WJ, Brodie A, Kouki J, Lindenmayer DB, Lõhmus A, Pastur GM, Messier C, Neyland M, Palik B, Sverdrup-Thygeson A, Volney WJA, Wayne A, Franklin JF. 2012. Retention forestry to maintain multifunctional forests: a world perspective. BioScience 62(7):633–645. https://doi.org/10.1525/bio.2012.62.7.6.
Gustafsson L, Bauhus J, Asbeck T, Augustynczik ALD, Basile M, Frey J, Gutzat F, Hanewinkel M, Helbach J, Jonker M, Knuff A, Messier C, Penner J, Pyttel P, Reif A, Storch F, Winiger N, Winkel G, Yousefpour R, Storch I. 2020. Retention as an integrated biodiversity conservation approach for continuous-cover forestry in Europe. Ambio 49:85–97. https://doi.org/10.1007/s13280-019-01190-1.
He X, He KS, Hyvönen J. 2016. Will bryophytes survive in a warming world? Perspect. Plant Ecol. Evol. Syst. 19:49–60. https://doi.org/10.1016/j.ppees.2016.02.005.
Jahed RR, Kavousi MR, Farashiani ME, Sagheb-Talebi K, Babanezhad M, Courbaud B, Larrieu L. 2020. A comparison of the formation rates and composition of tree-related microhabitats in beech-dominated primeval Carpathian and Hyrcanian forests. Forests 11(2):144. https://doi.org/10.3390/f11020144.
Jakoby O, Lischke H, Wermelinger B. 2019. Climate change alters elevational phenology patterns of the European spruce bark beetle (Ips typographus). Global Chang. Biol. 25(12):4048–4063. https://doi.org/10.1111/gcb.14766.
Janda P, Trotsiuk V, Mikoláš M, Bače R, Nagel TA, Seidl R, Svoboda M. 2017. The historical disturbance regime of mountain Norway spruce forests in the Western Carpathians and its influence on current forest structure and composition. For. Ecol. Manag. 388:67–78. https://doi.org/10.1016/j.foreco.2016.08.014.
Joa B, Schraml, U. 2020. Conservation practiced by private forest owners in Southwest Germany–the role of values, perceptions and local forest knowledge. Forest Policy Econ. 115:102141. https://doi.org/10.1016/j.forpol.2020.102141.
Kahl T, Arnstadt T, Baber K, Bässler C, Bauhus J, Gossner MM. 2017. Wood decay rates of 13 temperate tree species in relation to wood properties, enzyme activities and organismic diversities. For. Ecol. Manag. 391:86–95. https://doi.org/10.1016/j.foreco.2017.02.012.
Kitching RL. 1971. An ecological study of water-filled tree-holes and their position in the woodland ecosystem. J. Anim. Ecol. 40(2):281. https://doi.org/10.2307/3247.
Kõrkjas M, Remm L, Lõhmus A. 2021. Development rates and persistence of the microhabitats initiated by disease and injuries in live trees: a review. For. Ecol. Manag. 482:118833. https://doi.org/10.1016/j.foreco.2020.118833.
Kozák D, Svitok M, Zemlerová V, Mikoláš M, Lachat T, Larrieu L, Svoboda M. 2023. Importance of conserving large and old trees to continuity of tree-related microhabitats. Conserv Biol. https://doi.org/10.1111/cobi.14066.
Kraus D, Bütler R, Krumm F, Lachat T, Larrieu L, Mergner U, Paillet Y, Rydkvist T, Schuck A, Winter S. 2016. Catalogue Tree-Microhabitats Reference-Field-List. Integrate+ Technical Paper. http://www.integrateplus.org/uploads/images/Mediacenter/Catalogue_Tree-Microhabitats_Reference-Field-List_EN.pdf
Larrieu L, Gonin P. 2008. L’indice de biodiversité potentielle (ibp): une méthode simple et rapide pour évaluer la biodiversité potentielle des peuplements forestiers. Revue Forestière Française. https://doi.org/10.4267/2042/28373.
Larrieu L, Cabanettes A. 2012. Species, live status, and diameter are important tree features for diversity and abundance of tree microhabitats in subnatural montane beech–fir forests. Can. J. for. Res. 42(8):1433–1445. https://doi.org/10.1139/x2012-077.
Larrieu L, Cabanettes A, Brin A, Bouget C, Deconchat M. 2014. Tree microhabitats at the stand scale in montane beech–fir forests: practical information for taxa conservation in forestry. Eur. J. for. Res. 133(2):355–367. https://doi.org/10.1007/s10342-013-0767-1.
Larrieu L, Paillet Y, Winter S, Bütler R, Kraus D, Krumm F, Lachat T, Michel AK, Regnery B, Vandekerkhove K. 2018. Tree related microhabitats in temperate and Mediterranean European forests: a hierarchical typology for inventory standardization. Ecol. Indic. 84:194–207. https://doi.org/10.1016/j.ecolind.2017.08.051.
Larrieu L, Courbaud B, Drénou C, Goulard M, Bütler R, Kozák D, Kraus D, Krumm F, Lachat T, Müller J, Paillet Y, Schuck A, Stillhard J, Svoboda M, Vandekerkhove K. 2022. Perspectives: key factors determining the presence of tree-related microhabitats: a synthesis of potential factors at site, stand and tree scales, with perspectives for further research. For. Ecol. Manag. 515:120235. https://doi.org/10.1016/j.foreco.2022.120235.
Larsen JB, Angelstam P, Bauhus J, Carvalho JF, Diaci J, Dobrowolska D, Gazda A, Gustafsson L, Krumm F, Knoke T, Konczal A, Kuuluvainen T, Mason B, Motta R, Pötzelsberger E, Rigling A, Schuck A. 2022. Closer-to-nature forest management (from science to policy) [from science to policy]. Eur for Inst. https://doi.org/10.36333/fs12.
Liaw A, Wiener M. 2002. Classification and regression by randomForest. R News 2(3):18–22.
Lie MH, Arup U, Grytnes JA, Ohlson M. 2009. The importance of host tree age, size and growth rate as determinants of epiphytic lichen diversity in boreal spruce forests. Biodivers. Conserv. 18(13):3579–3596. https://doi.org/10.1007/s10531-009-9661-z.
Lindenmayer DB, Laurance WF, Franklin JF. 2012. Global decline in large old trees. Science 338(6112):1305. https://doi.org/10.1126/science.1231070.
Lindenmayer DB, Laurance WF, Franklin JF, Likens GE, Banks SC, Blanchard W, Gibbons P, Ikin K, Blair D, McBurney L, Manning AD, Stein JAR. 2014. New policies for old trees: averting a global crisis in a keystone ecological structure. Conserv. Lett. 7(1):61–69. https://doi.org/10.1111/conl.12013.
Lindenmayer DB, Wood J, McBurney L, Michael D, Crane M, MacGregor C, Montague-Drake R, Gibbons P, Banks SC. 2011. Cross-sectional vs. longitudinal research: a case study of trees with hollows and marsupials in Australian forests. Ecol. Monogr. 81(4):557–580. https://doi.org/10.1890/11-0279.1.
Lindenmayer DB, Wood JT. 2010. Long-term patterns in the decay, collapse, and abundance of trees with hollows in the mountain ash (Eucalyptus regnans) forests of Victoria, southeastern Australia. Can. J. for. Res. 40(1):48–54. https://doi.org/10.1139/X09-185.
Mäkinen H. 2002. Effect of stand density on the branch development of silver birch (Betula pendula Roth) in central Finland. Trees 16(4):346–353. https://doi.org/10.1007/s00468-002-0162-x.
Martin M, Paillet Y, Larrieu L, Kern CC, Raymond P, Drapeau P, Fenton NJ. 2022. Tree-related microhabitats are promising yet underused tools for biodiversity and nature conservation: a systematic review for international perspectives. Front. for. Glob. Chang. 5:818474. https://doi.org/10.3389/ffgc.2022.818474.
Martin M, Raymond P, Boucher Y. 2021. Influence of individual tree characteristics, spatial structure and logging history on tree-related microhabitat occurrence in North American hardwood forests. For. Ecosyst. 8(1):27. https://doi.org/10.1186/s40663-021-00305-z.
Martínez Pastur GJ, Vanha-Majamaa I, Franklin JF. 2020. Ecological perspectives on variable retention forestry. Ecol. Process. 9(1):12. https://doi.org/10.1186/s13717-020-0215-3.
Menkis A, Redr D, Bengtsson V, Hedin J, Niklasson M, Nordén B, Dahlberg A. 2022. Endophytes dominate fungal communities in six-year-old veteranisation wounds in living oak trunks. Fungal Ecol. 59:101020. https://doi.org/10.1016/j.funeco.2020.101020.
Metzler B, Hecht U, Nill M, Brüchert F, Fink S, Kohnle U. 2012. Comparing Norway spruce and silver fir regarding impact of bark wounds. For. Ecol. Manag. 274:99–107. https://doi.org/10.1016/j.foreco.2012.02.016.
Meyer W, Meyer B. 2001. Construction and use of Black Woodpecker Dryocopus martius holes in Thuringia/Germany. Abh. Ber. Mus. Heineanum 5:121–131. in German.
Michel AK, Winter S. 2009. Tree microhabitat structures as indicators of biodiversity in Douglas-fir forests of different stand ages and management histories in the Pacific Northwest, U.S.A. For. Ecol. Manag. 257(6):1453–1464. https://doi.org/10.1016/j.foreco.2008.11.027.
Micó E. 2018. Saproxylic insects in tree hollows. In: Ulyshen MD, Ed. Saproxylic insects: diversity, ecology and conservation, . Berlin: Springer. pp 693–727. https://doi.org/10.1007/978-3-319-75937-1_21.
Millington WF, Chaney WR. 1973. Shedding of shoots and branches. In: Kozlowski TT, Ed. Shedding of plant parts. New York: Academic Press. pp 149–204.
Muys B, Angelstam P, Bauhus J, Bouriaud L, Jactel H, Kraigher H, Müller J, Pettorelli N, Pötzelsberger E, Primmer E, Svoboda M, Thorsen BJ, Van Meerbeek K, European Forest Institute. 2022. Forest biodiversity in Europe (From Science to Policy) [From Science to Policy]. Eur for Inst. https://doi.org/10.36333/fs13.
Nascimbene J, Casazza G, Benesperi R, Catalano I, Cataldo D, Grillo M, Isocrono D, Matteucci E, Ongaro S, Potenza G, Puntillo D. 2016. Climate change fosters the decline of epiphytic Lobaria species in Italy. Biol. Conserv. 201:377–384. https://doi.org/10.1016/j.biocon.2016.08.003.
Nykänen ML, Broadgate M, Kellomäki S, Peltola H, Quine C. 1997. Factors affecting snow damage of trees with particular reference to European conditions. Silva Fennica. https://doi.org/10.14214/sf.a8519.
O’Hara KL. 2007. Pruning wounds and occlusion: a long-standing conundrum in forestry. J. For. 105(3):131–138. https://doi.org/10.1093/jof/105.3.131.
Paillet Y, Coutadeur P, Vuidot A, Archaux F, Gosselin F. 2015. Strong observer effect on tree microhabitats inventories: a case study in a French lowland forest. Ecol. Indic. 49:14–23. https://doi.org/10.1016/j.ecolind.2014.08.023.
Paillet Y, Archaux F, Boulanger V, Debaive N, Fuhr M, Gilg O, Gosselin F, Guilbert E. 2017. Snags and large trees drive higher tree microhabitat densities in strict forest reserves. For. Ecol. Manag. 389:176–186. https://doi.org/10.1016/j.foreco.2016.12.014.
Paillet Y, Debaive N, Archaux F, Cateau E, Gilg O, Guilbert E. 2019. Nothing else matters? Tree diameter and living status have more effects than biogeoclimatic context on microhabitat number and occurrence: an analysis in French forest reserves. PLoS ONE 14(5):e0216500. https://doi.org/10.1371/journal.pone.0216500.
Pfeifer EM, Hicke JA, Meddens AJH. 2011. Observations and modeling of aboveground tree carbon stocks and fluxes following a bark beetle outbreak in the western United States. Global Chang. Biol. 17(1):339–350. https://doi.org/10.1111/j.1365-2486.2010.02226.x.
Puettman KJ, Coates KD, Messier C. 2009. A critique of silviculture: managing for complexity. Washington, D.C.: Island Press. p 188p.
Pouska V, Lepš J, Svoboda M, Lepšová A. 2011. How do log characteristics influence the occurrence of wood fungi in a mountain spruce forest? Fungal Ecol. 4:201–209. https://doi.org/10.1016/j.funeco.2010.11.004.
Puverel C, Abourachid A, Böhmer C, Leban JM, Svoboda M, Paillet Y. 2019. This is my spot: what are the characteristics of the trees excavated by the Black Woodpecker? A case study in two managed French forests. For. Ecol. Manag. 453:117621. https://doi.org/10.1016/j.foreco.2019.117621.
Ranius T. 2002. Osmoderma eremita as an indicator of species richness of beetles in tree hollows. Biodivers. Conserv. 11(5):931–941. https://doi.org/10.1023/A:1015364020043.
R Core Team. 2021. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing.
Remm J, Lõhmus A. 2011. Tree cavities in forests—The broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 262(4):579–585. https://doi.org/10.1016/j.foreco.2011.04.028.
Schütz JP, Götz M, Schmid W, Mandallaz D. 2006. Vulnerability of spruce (Picea abies) and beech (Fagus sylvatica) forest stands to storms and consequences for silviculture. Eur. J. for. Res. 125(3):291–302. https://doi.org/10.1007/s10342-006-0111-0.
Seidl R, Schelhaas MJ, Lexer MJ. 2011. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 17(9):2842–2852. https://doi.org/10.1111/j.1365-2486.2011.02452.x.
Senf C, Pflugmacher D, Zhiqiang Y, Sebald J, Knorn J, Neumann M, Hostert P, Seidl R. 2018. Canopy mortality has doubled in Europe’s temperate forests over the last three decades. Nat. Commun. 9(1):4978. https://doi.org/10.1038/s41467-018-07539-6.
Shigo AL. 1984. Compartmentalization: a conceptual framework for how trees grow and defend themselves. Annu. Rev. Phytopathol. 22:189–214.
Smith KT. 2015. Compartmentalization, resource allocation, and wood quality. Curr. for. Rep. 1(1):8–15.
Spielmann M, Bücking W, Quadt V, Krumm F. 2013. Integration of nature protection in forest policy in Baden-Württemberg (Germany). INTEGRATE country report. EFICENT-OEF. Freiburg.
Spînu AP, Asbeck T, Bauhus J. 2022. Combined retention of large living and dead trees can improve provision of tree-related microhabitats in Central European montane forests. Eur. J. for. Res. 141(6):1105–1120. https://doi.org/10.1007/s10342-022-01493-1.
Spînu AP, Mysiak W, Bauhus J, Bielak K, Niklasson M. 2023. Pioneer tree species accelerate restoration of tree-related microhabitats in 50-year-old reserves of Białowieża Forest, Poland. Ecol. Evol. 13(7):e10238. https://doi.org/10.1002/ece3.10238.
Spînu AP, 2023. Quality and temporal development of habitat trees and their microhabitats in temperate European forests. Thesis submitted in partial fulfilment of the requirements of the degree Doctor rer. nat. of the Faculty of Environment and Natural Resources, University of Freiburg, Freiburg im Breisgau, Germany. https://freidok.uni-freiburg.de/fedora/objects/freidok:241287/datastreams/FILE1/content
Stoffel M. 2005. Assessing the vertical distribution and visibility of rockfall scars in trees (reviewed paper). Schweiz. z. Fur Forstwes. 156(6):195–199. https://doi.org/10.3188/szf.2005.0195.
Stoffel M, Perret S. 2006. Reconstructing past rockfall activity with tree rings: some methodological considerations. Dendrochronologia 24(1):1–15. https://doi.org/10.1016/j.dendro.2006.04.001.
Siitonen J. 2012. Microhabitats. In: Stokland J, Siitonen J, Jonsson BG, Eds. Biodiversity in dead wood. Cambridge: Cambridge University Press.
Svensson GP, Larsson MC, Hedin J. 2004. Attraction of the larval predator Elater ferrugineus to the sex pheromone of its prey, Osmoderma eremita, and its implication for conservation biology. J. Chem. Ecol. 30(2):353–363. https://doi.org/10.1023/b:joec.0000017982.51642.8c.
Tavankar F, Ezzati S, Latterini F, Lo Monaco A, Venanzi R, Picchio R. 2022. Assessment of wound recovery and radial growth 10 years after forest operations in hardwood stands. Forests 13(9):1393. https://doi.org/10.3390/f13091393.
Tavankar F, Picchio R, Nikooy M, Lo Monaco A, Venanzi R, Bodaghi AI. 2017. Healing rate of logging wounds on broadleaf trees in Hyrcanian forest with some technological implications. Drewno. Prace Naukowe. Doniesienia. Komunikaty 60:199. https://doi.org/10.12841/wood.1644-3985.200.05.
Therneau T. 2023. A Package for Survival Analysis in R. R package version 3.5–5. https://CRAN.R-project.org/package=survival
Trappmann D, Stoffel M. 2013. Counting scars on tree stems to assess rockfall hazards: a low effort approach, but how reliable? Geomorphology 180–181:180–186. https://doi.org/10.1016/j.geomorph.2012.10.009.
Vandekerkhove K, Vanhellemont M, Vrška T, Meyer P, Tabaku V, Thomaes A, Verheyen K. 2018. Very large trees in a lowland old-growth beech (Fagus sylvatica L.) forest: density, size, growth and spatial patterns in comparison to reference sites in Europe. For. Ecol. Manag. 417:1–17. https://doi.org/10.1016/j.foreco.2018.02.033.
Vasaitis R, Bakys R, Vasiliauskas A. 2012. Discoloration and associated fungi in stems of silver birch (Betula pendula Roth.) following logging damage. For. Pathol. 42(5):387–392. https://doi.org/10.1111/j.1439-0329.2012.00771.x.
Vogel S, Alvarez B, Bässler C, Müller J, Thorn S. 2017. The Red-belted Bracket (Fomitopsis pinicola ) colonizes spruce trees early after bark beetle attack and persists. Fungal Ecol. 27:182–188. https://doi.org/10.1016/j.funeco.2016.12.007.
Vuidot A, Paillet Y, Archaux F, Gosselin F. 2011. Influence of tree characteristics and forest management on tree microhabitats. Biol. Conserv. 144(1):441–450. https://doi.org/10.1016/j.biocon.2010.09.030.
Wesołowski T. 2011. “Lifespan” of woodpecker-made holes in a primeval temperate forest: a thirty year study. For. Ecol. Manag. 262(9):1846–1852. https://doi.org/10.1016/j.foreco.2011.08.001.
Wesołowski T, Martin K. 2018. Tree holes and hole nesting birds in European and North American forests. In: Mikusiński G, Roberge J-M, Fuller RJ, Eds. Ecology and conservation of European forest birds. Cambridge: Cambridge University Press. pp 79–134. https://doi.org/10.1017/9781139680363.
Wickham H. 2016. ggplot2: Elegant graphics for data analysis. Springer-Verlag New York. ISBN 978-3-319-24277-4. https://ggplot2.tidyverse.org
Winter S, Höfler J, Michel AK, Böck A, Ankerst DP. 2015. Association of tree and plot characteristics with microhabitat formation in European beech and Douglas-fir forests. Eur. J. for. Res. 134(2):335–347. https://doi.org/10.1007/s10342-014-0855-x.
Zekhuis M. 2018. Ivy can go up the tree! Vakblad Natuur Bos Landschap 15(5):12–16.
Zeller L, Baumann C, Gonin P, Heidrich L, Keye C, Konrad F, Larrieu L, Meyer P, Sennhenn-Reulen H, Müller J, Schall P, Ammer C. 2022. Index of biodiversity potential (IBP) versus direct species monitoring in temperate forests. Ecol. Indic. 136:108692. https://doi.org/10.1016/j.ecolind.2022.108692.
Zemlerová L, Trotsiuk V, Morrissey RC, Bače R, Mikoláš M, Svoboda M. 2017. Old trees as a key source of epiphytic lichen persistence and spatial distribution in mountain Norway spruce forests. Biodivers. Conserv. 26(8):1943–1958. https://doi.org/10.1007/s10531-017-1338-4.
Zemlerová V, Kozák D, Mikoláš M, Svitok M, Bače R, Smyčková M, Svoboda M. 2023. Natural disturbances are essential determinants of tree-related microhabitat availability in temperate forests. Ecosystems. https://doi.org/10.1007/s10021-023-00830-8.
Acknowledgements
We are grateful to the field assistants who collected data and to the managers of forest districts and reserves, who made this study possible. The data collected in France were part of “Protocole de Suivi des Réserves Forestières” (PSDRF), ONF-RNF. We would like to thank Cătălina Munteanu, Michael Wohlwend, Radek Bače and Katarína Markuljaková for suggestions on the manuscript and research support.
Funding
Open Access funding enabled and organized by Projekt DEAL. The study was funded by the Germany Research Foundation (DFG) as part of the Research Training Group GRK 2123/2 ConFoBi and by Czech Science Foundation (Grand GACR 22–31322S). Our study brings together authors from a number of different countries, including scientists based or born in the regions where the study was carried out.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare there are no competing interests.
Additional information
Author contributions
A. P. Spînu and J. Bauhus developed the study design; A. P. Spînu, T. Asbeck, D. Kozak, M. Svoboda, M. Mikoláš, Y. Paillet collected the data and contributed to the interpretation of the results. A. P Spînu and A. Nicolaie carried out the data analysis based on the analytical methodology designed by A. Nicolaie. A. P. Spînu was in charge of the overall direction and planning, manuscript writing with significant input from J. Bauhus. All authors contributed to the final manuscript.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Spînu, A.P., Nicolaie, M.A., Asbeck, T. et al. Temporal Development of Microhabitats on Living Habitat Trees in Temperate European Forests. Ecosystems (2024). https://doi.org/10.1007/s10021-024-00915-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10021-024-00915-y