
Abstract
Background
Erythromycin A (Er-A) produced by the actinomycete Saccharopolyspora erythraea is an important antibiotic extensively used in human medicine. Dissecting of transcriptional regulators and their target genes associated with erythromycin biosynthesis is crucial to obtain erythromycin overproducer strains through engineering of relevant regulatory elements in S. erythraea.
Results
Here, we identified a TetR family transcriptional regulator (TFR), SACE_5754, negatively controlling erythromycin production. SACE_5754 indirectly repressed the transcription of ery cluster and cannot regulate itself and its adjacent gene SACE_5753. RNA-seq coupled with EMSAs and qRT-PCR was performed to identify the targets of SACE_5754, and confirmed that transcription of SACE_0388 (encoding a pyruvate, water diknase), SACE_3599 (encoding an antibiotic resistance macrolide glycosyltransferase) and SACE_6149 (encoding a FAD-binding monooxygenase) were directly repressed by SACE_5754. A consensus palindromic sequence TYMAGG-n2/n4/n11-KKTKRA (Y: C/T, M: A/C, K: T/G, R: A/G) was proved to be essential for SACE_5754 binding using DNase I footprinting and EMSAs. During the three target genes of SACE_5754, SACE_0388 and SACE_6149 exhibited the positive effect on erythromycin production. Overexpression of either SACE_0388 or SACE_6149 in ∆SACE_5754 further increased the Er-A production. By engineering the industrial strain S. erythraea WB with deletion of SACE_5754 combined with overexpression of either SACE_0388 or SACE_6149, Er-A production in WB∆SACE_5754/pIB139–0388 and WB∆SACE_5754/pIB139–6149 was successively increased by 42 and 30% compared to WB. Co-overexpression of SACE_0388 and SACE_6149 in WB∆SACE_5754 resulted in enhanced Er-A production by 64% relative to WB. In a 5-L fermenter, WB∆SACE_5754/pIB139–0388-6149 produced 4998 mg/L Er-A, a 48% increase over WB.
Conclusion
We have identified a TFR, SACE_5754, as a negative regulator of erythromycin biosynthesis, and engineering of SACE_5754 and its target genes, SACE_0388 and SACE_6149, resulted in enhanced erythromycin production in both wild-type and industrial S. erythraea strains. The strategy demonstrated here may be valuable to facilitate the manipulation of transcriptional regulators and their targets for production improvement of antibiotics in industrial actinomycetes.
Similar content being viewed by others

Background
Erythromycin A (Er-A) is widely used in clinic against pathogenic Gram-positive bacteria, industrially produced by the actinomycete Saccharopolyspora erythraea [1]. As a model of polyketide biosynthesis, erythromycin biosynthesis has been investigated by genetic and biochemical assays for production improvement and structure diversification [2]. At first, two primary metabolic precursors, propionyl-CoA and (2S)-methylmalonyl-CoA units, are enzymatically catalyzed by multifunctional modular PKS to form 6-deoxyerythronolide B (6-dEB) [3]. Next, a series of tailoring reactions including hydroxylation, glycosylation, and methylation take place to produce the final product Er-A. Erythromycin biosynthetic gene cluster (ery cluster) in S. erythraea contains 20 genes arranged in four major polycistronic units [4], but lacks regulatory genes, hampering efforts to improve erythromycin production by engineering relevant regulatory elements.
Traditionally, optimization of medium composition, random mutagenesis and selection have been performed to enhance erythromycin production [5]. Nowadays, rational engineering of the metabolic and regulatory pathways involved in biosynthesis of erythromycin was also an effective way to increase erythromycin production [6, 7]. Since the advent of “Omics” era, the genome-based functional investigations of antibiotic producers shed new lights on systematical understanding of molecular regulation of antibiotic biosynthesis [8]. Especially, transcriptome-driven reverse engineering strategy have been utilized for identifying and manipulating regulatory genes for production improvement of polyketide antibiotics [9]. However, few studies were reported on transcriptome-based target identification of transcriptional regulator associated with antibiotic biosynthesis in actinomycete.
The TetR family transcriptional regulators (TFRs), a class of regulators commonly found in bacteria, participated in diverse cellular processes [10]. In recent years, a variety of TFRs, inside or outside the gene cluster of antibiotic biosynthesis in actinomycete, were identified to regulate the biosynthesis of antibiotics by binding to the promoter region of their target genes [11,12,13,14]. Based on our previous prediction [12], 97 putative TFRs were encoded in the genome of S. erythraea, in which SACE_3986, SACE_7301, SACE_3446 and PccD (SACE_3396) were successively proved to regulate erythromycin biosynthesis [12, 15,16,17]. Nevertheless, additional TFRs associated with erythromycin production in S. erythraea need to be further identified to improve the understanding of regulatory mechanism underlying erythromycin biosynthesis.
Here, we identified a novel TFR, SACE_5754, indirectly repressing the erythromycin biosynthesis. SACE_5754 cannot regulate itself and its adjacent gene, increasing the difficulty to study its targets to understand the molecular mechanism of SACE_5754 for regulating erythromycin biosynthesis. Thereby, this study utilized RNA-seq based transcriptome analysis coupled with qRT-PCR, EMSAs and genetic experiments to identify and characterize SACE_5754’s targets related to erythromycin production. Further engineering SACE_5754 and its target genes significantly increased erythromycin production in both wild-type and industrial S. erythraea strains.
Results
SACE_5754 plays a negative regulatory role in the biosynthesis of erythromycin
Considering the key role of TFRs in antibiotic biosynthesis in actinomycetes, we performed gene inactivation and identified several TFRs involved in erythromycin production in S. erythraea. These contained three TFRs (SACE_3986, SACE_7301 and SACE_3446) previously published and SACE_5754 currently investigated. SACE_5754 contains 624 nucleotides with approximately 22 kDa molecular mass. The location of SACE_5754 and its adjacent genes on the chromosome were shown in Fig. 1a. Blast analysis revealed that SACE_5754 homologs are widely distributed among actinomycetes, suggesting that this TFR may have important biological functions in actinomycetes (Additional file 1: Figure S1a).

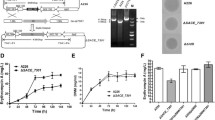
Inactivation of SACE_5754 in S. erythraea A226. a Genetic organization of SACE_5754 and its adjacent genes in S. erythraea A226. b Schematic deletion of SACE_5754 by linearized fragment homologous recombination in S. erythraea A226. c PCR confirmation of the SACE_5754 deletion mutant. Lanes: M, 5000-bp DNA ladder; pUCTSR∆5754, the positive control, 1727 bp amplified from pUCTSR∆5754; A226, the negative control, 624 bp amplified from A226; ∆SACE_5754, 1727 bp amplified from ∆SACE_5754. d. Er-A production in S. erythraea A226 and its derivatives. Mean values of three replicate are shown, with the standard deviation indicated by error bars. *p < 0.05, **p < 0.01, ***p < 0.001, ns, no significant (relative to A226)
SACE_5754 was disrupted with a tsr replacement in S. erythraea A226, generating the ∆SACE_5754 mutant (Fig. 1b and c). Er-A production in ∆SACE_5754 (64 mg/L) was 41% higher than that in strain A226 (45 mg/L). The complemented strain ∆SACE_5754/pIB139–5754 restored the original erythromycin level, suggesting that production enhancement of erythromycin in ∆SACE_5754 was solely due to the SACE_5754 disruption (Fig. 1d). Furthermore, pIB139–5754 and pIB139 was individually introduced into strain A226. The SACE_5754-overexpressed strain A226/pIB139–5754 (31 mg/L) successively exhibited 31 and 28% reduction in Er-A production relative to A226 and A226/ pIB139 (43 mg/L) (Fig. 1d). Deletion and overexpression of SACE_5754 in strain A226 had no effect on cell growth and sporulation, inferring that SACE_5754 was not involved in cell growth or morphological differentiation of S. erythraea (Additional file 1: Figure S1b and 1c).
SACE_5754 indirectly regulates erythromycin biosynthesis
To prove the possibility that SACE_5754 regulates erythromycin production through ery cluster (Fig. 2a), we measured transcripts of ery cluster by qRT-PCR. Results showed that the transcriptional levels of eryAI (SACE_0721, encoding polyketide synthase I), eryBIII (SACE_0731, encoding NDP-4-keto-2, 6-dideoxyhexose 3-C-methyltransferase), ermE (SACE_0733, encoding rRNA methyltransferase) and eryCI (SACE_0734, encoding transaminase) were respectively increased by 4-, 2.7-, 4- and 3- folds compared with A226 (Fig. 2d), indicating SACE_5754 might negatively regulate erythromycin biosynthesis.

SACE_5754 negatively regulates erythromycin production in S. erythraea A226. a Genetic organization of ery cluster, SACE_5754 and its adjacent gene SACE_5753 in S. erythraea A226. b Purification of His6-tagged SACE_5754. c EMSAs of the interaction of probes PeryBVI, PeryAI-BIV, PeryBI-BIII, PeryCI-ermE, PeryK and P5753–5754 with purified His6-SACE_5754 protein. d Effect of SACE_5754 deletion on transcription levels of ery cluster, SACE_5753 and SACE_5754. qRT-PCR was used to quantify the amounts of transcripts in A226 and ΔSACE_5754 cultured for 48 h in liquid R5 medium. Mean values of three replicates are shown, with the standard deviation indicated by error bars.*p < 0.05, **p < 0.01, ***p < 0.001, ns, no significant
To examine whether SACE_5754 might directly regulate transcription of ery cluster, it was expressed in E. coil BL21 (DE3), and was used to examine its affinity to the five regions containing ery promoters with EMSAs (Fig. 2b). Results showed that SACE_5754 did not bind to ery promoters (Fig. 2c), demonstrating that SACE_5754 may regulate erythromycin production by indirectly repressing the expression of ery cluster.
SACE_5754 cannot transcriptionally regulate its own gene and its adjacent gene SACE_5753
Most TFRs are transcriptional regulators to regulate their adjacent genes and/or themselves [6]. To investigate whether SACE_5754 directly binds to its adjacent gene SACE_5753 and to its own promoter regions, His6-SACE_5754 was used to perform EMSAs with DNA fragment P5753–5754, which covers the entire promoter regions of SACE_5753 and SACE_5754, but no obvious gel shift band was detected. Furthermore, we measured transcripts of SACE_5754 and SACE_5753 by qRT-PCR, and found that the transcriptional levels of SACE_5754 and SACE_5753 had no change in A226 and ∆SACE_5754, suggesting that SACE_5754 cannot regulate its own gene and adjacent gene SACE_5753 (Fig. 2d).
Finding SACE_5754 target genes based on transcriptomic data
In order to search for SACE_5754’s targets, transcriptomic comparison between A226 and ∆SACE_5754 was performed by RNA-seq. With the statistic criteria of Foldchange ≥2 and probability ≥0.8, a total of 623 differential expressed genes were identified (440 up-regulated genes and 183 down-regulated genes) in ∆SACE_5754. Herein, promoter regions of 80 genes with a greater than 0.9 probability in the transcriptomic data were tested by EMSAs (Additional file 1: Table S3), and results showed that SACE_5754 could specifically bind to P0388–0389、P3599 and P6148–6149 (Fig. 3a, b and c). Further qRT-PCR analysis found that SACE_0389 and SACE_6148 were not affected by SACE_5754 deletion, but the transcriptional levels of SACE_0388, SACE_3599 and SACE_6149 were increased by 158-, 4.7- and 95- folds, respectively (Fig. 3d). It seemed that SACE_5754 directly represses the transcription of SACE_0388, SACE_3599 and SACE_6149.

SACE_5754 directly represses the expression of SACE_0388, SACE_3599 and SACE_6149. a Genetic organization of candidate target genes of SACE_5754. b EMSAs with SACE_5754 and P0388–0389. c EMSAs with SACE_5754 and P3599. d EMSAs with SACE_5754 and P6148–6149. e Effect of SACE_5754 deletion on transcription levels of related genes. qRT-PCR was used to quantify the amounts of transcripts in A226 and ΔSACE_5754 cultured for 48 h in liquid R5 medium. Mean values of three replicates are shown, with the standard deviation indicated by error bars. *p < 0.05, **p < 0.01, ***p < 0.001, ns, no significant
Identification of the SACE_5754-binding site
In order to precisely determine SACE_5754’s binding site within the intergenic region of SACE_0388 and SACE_0389, a DNase I footprinting experiment was performed, and a 12-bp palindromic sequence AAATCAACACTTGTTGAATTA (termed site A) in P0388–0389 was protected by SACE_5754 (Fig. 4a). To assess the importance of the palindromic sequence for SACE_5754 binding, EMSAs were performed with different probes, named probe 1 (no mutation of site A), probe 2 (with base substitution without the palindromic sequence of site A) and probe 3 (with base substitution within the palindromic sequence of site A) (Fig. 4b). As expected, probe 2 showed the same band shift as probe 1, while probe 3 showed no binding (Fig. 4b), demonstrating that TCAACA-n2-TGTTGA was indispensable for SACE_5754 binding.

Analysis of the SACE_5754-binding sites. a DNase I footprinting assay of SACE_5754-binding site in the P0388–0389. Upper fluorogram: control reaction without protein. Protection regions were acquired with increasing concentrations (0.5 μM and 0.2 μM) of His6-SACE_5754 protein. b SACE_5754 binds to the probe P1and mutated probe P2 and P3 by EMSAs. c DNase I footprinting assay of SACE_5754-binding site in the P3599. Upper fluorogram: control reaction without protein. Protection regions were acquired with increasing concentrations (0.2 μM and 0.5 μM) of His6-SACE_5754 protein. d SACE_5754 binds to the probe P4 and mutated probe P5 and P6 by EMSAs. e DNase I footprinting assay of SACE_5754-binding site in the P6148–6149. Upper fluorogram: control reaction without protein. Protection regions were acquired with increasing concentrations (0.05 μM and 0.2 μM) of His6-SACE_5754 protein. f SACE_5754 binds to the probe P5 and mutated probe P6 and P7 by EMSAs
DNase I footprinting experiments showed that P3599 and P6148–6149 also had 12-bp palindromic sequences protected by SACE_5754, CGTTGAATTCAACACATG TTCAACA (termed site B) and GTTATTCAACATGCGTTGAAAACT (termed site C), respectively (Fig. 4c and e). EMSAs with base substitution mutagenesis showed that the 12-bp palindromic sequences were indispensable for SACE_5754 binding (Fig. 4d and f). Those findings indicated that the palindromic sequence TYMAGG-n2/n4/n11-KKTKRA (Y: C/T, M: A/C, K: T/G, R: A/G) was essential for SACE_5754 binding.
Overexpression of SACE_0388 and SACE_6149 in ∆SACE_5754 improves erythromycin production
In order to explore the relationship of SACE_0388, SACE_3599 and SACE_6149, SACE_5754’s targets, to erythromycin biosynthesis, the ∆SACE_0388, ∆SACE_3599 and ∆SACE_6149 mutants were individually constructed (Additional file 1: Figure S2-S4). Compared with A226, ∆SACE_0388 and ∆SACE_6149 respectively produced lower Er-A by 80 and 60% (Fig. 5a and b); however, ∆SACE_3599 showed no yield change (Fig. 5c). Overexpression of SACE_0388 and SACE_6149 in A226 improved Er-A production by 32 and 28% (Fig. 5a and b). Those findings indicated that SACE_0388 and SACE_6149 had the positive effect on erythromycin production, while SACE_3599 did not affect erythromycin production.

Relationship of SACE_0388, SACE_3599 and SACE_6149 with erythromycin production. a Er-A production in S. erythraea A226 and SACE_0388 mutant strains. b Er-A production in S. erythraea A226 and SACE_6149 mutant strains. c Er-A production in A226, SACE_3599-deleted strains ∆SACE_3599–1, ∆SACE_3599–2 and ∆SACE_3599–3. Mean values of three replicate are shown, with the standard deviation indicated by error bars. *p < 0.05, **p < 0.01, ***p < 0.001, ns, no significant. (relative to A226)
Furthermore, to investigate whether overexpression of SACE_0388 and SACE_6149 in ∆SACE_5754 would further affect Er-A production, pIB139–0388 and pIB139–6149 was introduced into ∆SACE_5754 to obtain ∆SACE_5754/ pIB139–0388 and ∆SACE_5754/pIB139–6149, respectively. Compared with ∆SACE_5754, ∆SACE_5754/ pIB139–0388 and ∆SACE_5754/pIB139–6149 had 83 and 50% increase in the Er-A production, respectively (Fig. 6a).

Using SACE_5754 pathway to improve erythromycin production. a Er-A production in A226 and its derivatives. b Er-A production in S. erythraea WB and its derivatives. Mean values of three replicate are shown, with the standard deviation indicated by error bars. *p < 0.05, **p < 0.01, ***p < 0.001, ns, no significant (relative to A226 or WB). c Time course of Er-A production of WB and WB∆SACE_5754 /pIB139–0388-6149 in a 5 L fermenter
Engineering high-producing industrial S. erythraea WB enhances erythromycin production
To evaluate the possible improvement of erythromycin production in industrial settings, SACE_5754 and its target genes SACE_0388 and SACE_6149 were engineered in a high-yield industrial S. erythraea WB. Firstly, when SACE_5754 was deleted in WB, the obtained WB∆SACE_5754 exhibited production improvement of Er-A by 19% (Fig. 6b). Secondly, either pIB139–0388 or pIB139–6149 was introduced into WB∆SACE_5754. Compared with WB, the resulted WB∆SACE_5754/pIB139–0388 and WB∆SACE_5754/pIB139–6149 improved Er-A production by 42 and 30%, respectively (Fig. 6b). Lastly, co-expression of SACE_0388 and SACE_6149 in WB∆SACE_5754 led to a 64% increase in Er-A production compared to WB (Fig. 6b). Considering further application of the engineered strain in industry, we cultured WB∆SACE_5754/pIB139–0388-6149 in a 5-L fermenter for time course analysis of erythromycin production. The production of Er-A was increased by 48% from 3327 mg/L in WB to 4998 mg/L in WB∆SACE_5754/pIB139–0388-6149 (Fig. 6c).
Discussion
TFRs can be grouped into three types based on their orientation relative to neighboring genes in the genome, which can be used to predict the target genes of TFRs [10]. Type I TFR is divergently oriented to a neighboring transcription unit, as is the case for tetR and tetA, where the intergenic region between the two genes is less than 200 bp [18]. The majority of TFR genes are type I TFRs, which are generally auto-regulatory and also control the expression of adjacent genes [10]. The present work has characterized a novel TFR, SACE_5754, which negatively controls erythromycin biosynthesis in S. erythraea. However, SACE_5754, a type I TFR, cannot regulate itself and SACE_5753, and also didn’t bind to promoter regions of ery cluster, hampering the effort to identify its targets for mechanistic dissection of SACE_5754 regulating erythromycin biosynthesis. In this study, RNA-seq was first used to perform the transcriptomic comparison between the parental strain A226 and the ∆SACE_5754 mutant. Overall, 440 genes were up-regulated while 183 genes were down-regulated in ∆SACE_5754, with most of the genes related to the cellular metabolism in S. erythraea. Further, promoter regions of 80 genes with a greater than 0.9 probability in the RNA-seq data were selected by EMSAs to search for SACE_5754’s targets. Our results proved that SACE_5754 directly repressed the expression of SACE_0388, SACE_3599 and SACE_6149 by binding to the palindromic sequence TYMAGG-n2/n4/n11-KKTKRA (Y: C/T, M: A/C, K: T/G, R: A/G).
SACE_0388, encoding a pyruvate, water diknase, catalyze pyruvate into phosphoenol-pyruvate, which may enter into TCA cycle. Methylmalonyl-CoA can either be filled or drained by methylmalonyl-CoA mutase-catalyzed reaction converting intermediate succinyl-CoA and methylmalonyl-CoA of TCA cycle in S. erythraea [19, 20]. Coupled with transcriptome data, the expression of several genes involved in TCA cycle (citrate synthase, SACE_0069; fumarate reductase iron-sulfur subunit, SACE_1171; malate dehydrogenase, SACE_3674; methylmalonyl-CoA mutase subunit beta, SACE_5638; methylmalonyl-CoA mutase, SACE_5639; succinyl-CoA synthetase subunit alpha, SACE_6668; succinyl-CoA synthetase subunit beta, SACE_6669; phosphoenolpyruvate carboxykinase, SACE_7274) were analyzed by qRT-PCR. Compared to A226, SACE_1171 was transcriptionally down-regulated by 2.4- folds while the transcription of SACE_7274 was up-regulated by 2.0- folds, and other genes was not transcriptionally regulated by SACE_5754 (Additional file 1: Figure S5). These results suggested that SACE_0388 and SACE_ 7274 may carry more phosphoenol-pyruvate to TCA cycle while SACE_1171 may reduce the flow of succinyl-CoA to succinyl but instead converts to methylmalony-CoA metabolite pool, suggesting a possible way of enhancing erythromycin yield. Considering that SACE_0389, encoding a MarR family transcriptional regulator, was not differentially transcribed upon SACE_5754 inactivation, we did not perform gene inactivation of SACE_0389.
SACE_3599 encodes an antibiotic resistance macrolide glycosyltransferase. Glycosyltransferase is an important enzyme that catalyzes the attachment of sugar moieties to acceptor molecules [21]. Deletion of SACE_3599 in A226 didn’t affect erythromycin production, and transcription of SACE_3599 was increased by ~ 4-folds in ∆SACE_5754 relative to that in A226. Recent investigation found that through the regulation of a protein-level modification with overexpression of phosphopantetheinyl transferase genes in 33 Actinomycetes, most of strains displayed novel metabolite profiles [22]. We speculated that that SACE_3599 might be responsible for the biosynthesis of other secondary metabolites, which is worthwhile to do follow-up exploration. SACE_6149, encoding a FAD-binding monooxygenase which has been shown to cover a wide range of different oxygenation reactions [23], may modify some intermediates in erythromycin synthesis. Detailed insights of SACE_6149 into erythromycin production require further investigation. Based on current results, we propose a pathway of regulation of erythromycin production by SACE_5754 in S. erythraea (Additional file 1: Figure S6).
SACE_5754 stimulates erythromycin production by altering the transcription of ery cluster. However, this effect is indirect and SACE_5754’s downstream regulator interacting with promoter regions of ery cluster in S. erythraea remains unknown. Several transcriptional regulators were previously reported to involve in erythromycin biosynthesis in S. erythraea. For example: DasR was required for antibiotic production, pigment biosynthesis, morphological development, chitin and N-acetylglucosamine utilization [24]; SACE_Lrp is an efficient regulator for transporting and catabolizing branched-chain amino acids [6]; TFRs were involved in morphological differentiation, propionyl coenzyme A assimilation and erythromycin production [12, 15,16,17, 25, 26]. Combined with transcriptome data, promoter regions of these transcriptional regulators were selected by EMSAs (Additional file 1: Table S3), and results showed that these transcriptional regulators were not directly regulated by SACE_5754. The direct regulators of ery cluster would be exploring in our studies, for a better understanding of regulatory network of erythromycin production.
Manipulating the regulators and their targets is an efficient way to improve antibiotic production in actinomycetes. In an early study, deletion of the Lrp family transcriptional regulator SACE_Lrp in industrial strain WB enhanced the erythromycin production by 19%, while overexpression of its target gene SACE_5387–5386 in deletion of SACE_Lrp of WB increased the Er-A production by 48% [6]. In this study, deletion of SACE_5754 in the industrial strain WB combined with overexpression of its target genes SACE_0388 or SACE_6149 improved Er-A production by 46 and 34%, respectively. When SACE_0388 and SACE_6149 were further co-expressed in WBΔSACE_5754, Er-A production was increased by 64% compared to WB.
Our present findings providing a strategy for improved erythromycin production based on engineering of SACE_5754 and its target genes may be applied to the augment of other antibiotic produced by actinomycetes in industry that have SACE_5754 homologs.
Conclusions
SACE_5754, a TetR family transcriptional regulator, was identified to negatively regulate erythromycin biosynthesis in S. erythraea. It indirectly repressed the transcription of ery cluster and cannot regulate itself and its adjacent gene. RNA-seq coupled with qRT-PCR, EMSAs and genetic experiments were performed to characterize SACE_5754’s targets associated with erythromycin production. Further engineering of SACE_5754 and its target genes, SACE_0388 and SACE_6149, significantly enhanced erythromycin production relative to its original strain A226. When the combined engineering strategy was carried out in an industrial strain S. erythraea WB, erythromycin production was successively increased by 64% in a flask and by 41% in a fermenter. Our work here presents a new knowledge to understand the regulatory mechanism of erythromycin biosynthesis, and provides a useful strategy to identify and dissect target genes of transcriptional factors for improved erythromycin production.
Methods
Strains, plasmids, and growth conditions
The strains and plasmids used in this work were listed in Additional file 1: Table S1. S. erythraea and its derivatives were grown at 30 °C. Solid R3M medium was used for phenotypic observation and protoplast regeneration [27]. Liquid TSB medium was used for protoplast preparation, seed culture and DNA extraction [28]. The E. coil strain DH5α and BL21 (DE3) were cultured in LB liquid medium or on LB solid plates at 37 °C, used to propagate plasmids for routine cloning and as the host for heterologous protein production, respectively [29].
Gene deletion, complementation, and overexpression
Gene deletion, complementation, and overexpression in S. erythraea were showed as previously described [16] . To construct an SACE_5754 deletion mutant, two 1.5-kb DNA fragments flanking SACE_5754 were prepared from the genomic DNA of strain S. erythraea A226 by PCR using the primer pairs 5754-up-F/R and 5754-down-F/R (Additional file 1: Table S2). The two fragments were digested with HindIII/XbaI and KpnI/EcoRI, ligating into the corresponding sites of pUCTSR [25], generating pUCTSR∆5754. By the homologous recombination with linearized fragments, a 291-bp fragment of the SACE_5754 gene was replaced by thiostrepton resistance gene (tsr) in S. erythraea A226. The desired thiostrepton-resistant mutant, named ∆SACE_5754, was further confirmed by PCR analysis using the primers 5754-C-F/R (Additional file 1: Table S2).
For complementation of SACE_5754 in the ∆SACE_5754 and SACE_5754 overexpression in A226, a 624-bp DNA fragment containing a full-length SACE_5754 was amplified by PCR with the genomic DNA of A226 as a template. The PCR product was cleaved with NdeI/XbaI, and inserted into the corresponding sites of integrative expression plasmid pIB139 [30], yielding pIB139–5754. By PEG-mediated protoplast transformation, pIB139–5754 was introduced into the ∆SACE_5754 mutant and the parental strain A226, respectively. The complemented strain ∆SACE_5754/pIB139–5754 and overexpression strain A226/pIB139–5754 were obtained by apramycin resistance screening and confirmed by PCR analysis with primers Apr-F/R.
In accord with above procedures, we constructed SACE_0388 deletion mutant ∆SACE_0388, complemented of SACE_0388 in the ∆SACE_0388 (∆SACE_0388/pIB139–0388), SACE_0388 overexpression in A226 (A226/pIB139–0388), SACE_3599 deletion mutant ∆SACE_3599, SACE_6149 deletion mutant ∆SACE_6149, complemented of SACE_6149 in the ∆SACE_6149 (∆SACE_6149/pIB139–6149) and SACE_6149 overexpression in A226 (A226/pIB139–6149). All primers used in this work were listed in Additional file 1: Table S2.
For SACE_0388 and SACE_6149 overexpression in the ∆SACE_5754, pIB139–0388 and pIB139–6149 were introduced into ∆SACE_5754 strain to obtain ∆SACE_5754/pIB139–0388 and ∆SACE_5754/pIB139–6149 by PEG-mediated protoplast transformation, respectively. For co-expression of SACE_0388 and SACE_6149 in WB∆SACE_5754, the combined DNA fragment containing PermE* and SACE_6149 from the pIB139–6149 was amplified using the primer pair ermE-F and 6149-C-R. The PCR product was digested with NotI/EcoRV, ligating into the corresponding sites of pIB1390388 to generate pIB139–0388-6149. By PEG-mediated protoplast transformation, pIB139–0388-6149 was introduced into the WB∆SACE_5754. The co-expression SACE_0388 and SACE_6149 in the WB∆SACE_5754 strain WB∆SACE_5754/pIB139–0388-6149 was obtained by apramycin resistance screening and confirmed by PCR analysis with primers Apr-F/R.
Heterologous expression and purification of SACE_5754
The SACE_5754 coding region of 207 amino acids was obtained by PCR using the primers 5754-22b-F and 5754-22b-R (Additional file 1: Table S2). The PCR production was cut with EcoRI/HindIII and cloned into pET22b generating a C-terminal His-tag fusion. The constructed plasmid pET22b-5754 was introduced into E. coli BL21 (DE3), and SACE_5754 expression was induced by 0.5 mM IPTG at 20 °C for18–20 h. The recombinant His6-tagged SACE_5754 was extracted and purified on Ni2+-NTA spin column (Bio-RAD). The quality of the purified protein was estimated by polyacrylamide gel electrophoresis (SDS-PAGE). The purified protein was stored at 4 °C and used for electrophoretic mobility shift assays (EMSAs) and DNase I footprinting assays.
Electrophoretic mobility shift assays (EMSAs)
The EMSAs were performed in the light of previously reported methods [31]. DNA probes were amplified by PCR using the primers listed in Additional file 1: Table S2, and were incubated individually with various concentrations of His6-tagged SACE_5754 in binding-buffer (10 mM Tris (pH 7.5), 5 mM MgCl2, 60 mM KCl, 10 mM DTT, 50 mM EDTA and 10% glycerol) at 30 °C for 10 min in 20 μL reaction mixture. After incubation, the samples were fractionated on 6% native PAGE gels in 1× TAE buffer at 80 mA for 35–45 min.
DNase I footprinting assay
An FAM fluorescence labeling capillary electrophoresis method was used for DNase I footprinting [32]. To identify the binding site of the SACE_5754 protein in the P0388–0389 (the entire SACE_0388-SACE_0389 intergenic region), P3599 (the entire SACE_3599 intergenic region) and P6148–6149 (the entire SACE_6148-SACE_6149 intergenic region), a 261-bp 6-carboxyfluorescein (FAM), a 218-bp 6-carboxyfluorescein (FAM) and a 143-bp 6-carboxyfluorescein (FAM) fluorescence-labeled DNA fragment were amplified by PCR using primers FAM-P0388–0389-F/R, FAM-P3599-F/R and FAM-P6148–6149-F/R (Additional file 1: Table S2), respectively. The labeled DNA fragment (150 ng) and corresponding concentrations of His6-tagged SACE_5754 protein were added to a reaction mixture (final volume 50 μL), and incubated at 30 °C for 10 min in binding buffer. DNase I (1 U/μg; Promega) treatments with various concentrations were performed for 60 s at 25 °C, and terminated by addition of DNase I Stop Solution and heating for 10 min at 65 °C to inactivate the DNase I. DNA samples were analyzed with a 3730 XL DNA Genetic Analyzer (Applied Biosystems) after purification, and data analyses were performed using GeneMarker software program v2.2.
RNA isolation and quantitative real-time PCR assay
Total RNA was isolated, using the TransZol up (Transgen, China), from cultures of S. erythraea A226 and ∆SACE_5754 grown in R5 liquid medium after 2 days. The quality and quantity of RNAs were examined using a microplate reader (BioTek) and confirmed by electrophoresis. The transcription levels of various genes were determined by quantitative real-time PCR analysis as described previously [6] using the primers listed in Additional file 1: Table S2. qRT-PCR was performed on the Applied Biosystems QuantStudio 6 Flex system with Maxima™ SYBR Green/ROX qPCR Master Mix (MBI Fermentas). The hrdB (SACE_1801) gene was used as the internal control, and relative transcription was quantified using a comparative cycle threshold method [33].
Fermentation and measurement of erythromycin
For flask fermentation of S. erythraea and its derivatives, spores were inoculated into 50 ml of TSB seed medium and grown for 2 days. Then, 5 ml seed culture was transferred into 50 ml R5 liquid medium and all fermentation cultures were grown at 220 rpm, 30 °C for 6 days [27]. For bioreactor cultures, S. erythraea WB and its derivatives were cultivated in the industrial medium with a 5-L fermenter (Baoxing, Shanghai, China). Samples (50 ml) were taken every 24 h. Erythromycin was extracted in fermentation culture of the S. erythraea strains as described previously [6]. An Agilent Extend-C18 column (5 μm; 250 × 4.6 mm) was equipped in the Waters Breeze HPLC and was equilibrated with 60% solution A (5 mM ammonia acetate, pH 7.0) and 40% solution B (acetonitrile). An isocratic program was performed at a flow rate of 1.0 mL/min at 25 °C using 2424 ELSD detector.
RNA sequencing
Total RNA was isolated, using the TransZol up (Transgen, China), from cultures of S. erythraea A226 and ∆SACE_5754 grown in R5 liquid medium after 2 days. The quality and quantity of RNAs were examined using a microplate reader (BioTek) and confirmed by electrophoresis. RNA samples of A226 and ∆SACE_5754 were used for RNA sequencing. Library construction and sequencing were performed using Illumina Hiseq™ 2000 at The Beijing Genomics Institution (BGI, Shenzhen, China).The data obtained from the sequencing of the Illumina HiSeq ™ 2000 is called raw reads or raw data, and then the raw reads were subjected to quality control (QC)-controlled to determine if a resequencing step is needed. After raw reads are filtered, the clean reads were aligned to the reference sequence [34]. The original sequence data have been submitted to the database of NCBI Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra/SRP129064) under the accession number SRP129064. For gene expression analysis, the matched reads were calculated and then normalized to probability using Noiseq package method [35]. The significance of the differential expression of genes was defined by bioioformatics service of BGI according to the combination of the absolute criteria of Foldchange ≥2 and probability ≥0.8.
To validate the RNA-seq results, 28 genes annotated with biological functions and the probability from high to low were analyzed by qRT-PCR. Our qRT-PCR measurements for these genes showed similar trends of expression changes estimated from the RNA-seq data (Additional file 1: Figure S7).
Statistical analysis
All data in this study were stated as means ± standard error of mean (SEM), and analysis by Student’s t-test, with *p < 0.05, ** p < 0.01 and ***p < 0.001, ns, no significant.
Abbreviations
- 6-FAM:
-
6-isomer of carboxyfluorescein.
- CoA:
-
Coenzyme A
- EMSA:
-
Electrophoretic mobility shift assay
- Er-A:
-
Erythromycin A
- ery :
-
Erythromycin biosynthetic gene
- IPTG:
-
Isopropylthiogalactoside
- LB:
-
Luria-Bertani medium
- qRT-PCR:
-
Quantitative real-time PCR
- SDS-PAGE:
-
Sodium dodecyl sulfate polyacrylamide gel electrophoresis
- TCA:
-
Tricarboxylic acid
- TFR:
-
TetR family transcriptional regulator
- tsr :
-
Thiostrepton resistance gene
References
Butler MS. Natural products to drugs: natural product-derived compounds in clinical trials. Nat Prod Rep. 2008;25:475–516.
Barajas JF, Blake-Hedges JM, Bailey CB, Curran S, Keasling JD. Engineered polyketides: synergy between protein and host level engineering. Synth Syst Biotechnol. 2017;2:147–66.
Staunton J, Weissman KJ. Polyketide biosynthesis: a millennium review. Nat Prod Rep. 2001;18:380–416.
Reeves AR, English RS, Lampel JS, Post DA, Vanden Boom TJ. Transcriptional organization of the erythromycin biosynthetic gene cluster of Saccharopolyspora erythraea. J Bacteriol. 1999;181:7098–106.
Subathra Devi C, Saini A, Rastogi S, Jemimah Naine S, Mohanasrinivasan V. Strain improvement and optimization studies for enhanced production of erythromycin in bagasse based medium using Saccharopolyspora erythraea MTCC 1103. 3 Biotech. 2015;5:23–31.
Liu J, Chen Y, Wang W, Ren M, Wu P, Wang Y, Li C, Zhang L, Wu H, Weaver DT, Zhang B. Engineering of an Lrp family regulator SACE_Lrp improves erythromycin production in Saccharopolyspora erythraea. Metab Eng. 2017;39:29–37.
Reeves AR, Brikun IA, Cernota WH, Leach BI, Gonzalez MC, Weber JM. Engineering of the methylmalonyl-CoA metabolite node of Saccharopolyspora erythraea for increased erythromycin production. Metab Eng. 2007;9:293–303.
van der Heul HU, Bilyk BL, McDowall KJ, Seipke RF, van Wezel GP. Regulation of antibiotic production in Actinobacteria: new perspectives from the post-genomic era. Nat Prod Rep. 2018;35:575–604.
Nah JH, Kim HJ, Lee HN, Lee MJ, Choi SS, Kim ES. Identification and biotechnological application of novel regulatory genes involved in Streptomyces polyketide overproduction through reverse engineering strategy. Biomed Res Int. 2013;2013:549737.
Cuthbertson L, Nodwell JR. The TetR family of regulators. Microbiol Mol Biol Rev. 2013;77:440–75.
Ray S, Maitra A, Biswas A, Panjikar S, Mondal J, Anand R. Functional insights into the mode of DNA and ligand binding of the TetR family regulator TylP from Streptomyces fradiae. J Biol Chem. 2017;292:15301–11.
Wu H, Wang Y, Yuan L, Mao Y, Wang W, Zhu L, Wu P, Fu C, Müller R, Zhang L, et al. Inactivation of SACE_3446, a TetR family transcriptional regulator, stimulates erythromycin production in Saccharopolyspora erythraea. Synth Syst Biotechnol. 2016;1:39–46.
Lei C, Wang J, Liu Y, Liu X, Zhao G, Wang J. A feedback regulatory model for RifQ-mediated repression of rifamycin export in Amycolatopsis mediterranei. Microb Cell Factories. 2018;17:14.
Xu Y, Ke M, Li J, Tang Y, Wang N, Tan G, Wang Y, Liu R, Bai L, Zhang L, et al. TetR-type regulator SLCG_2919 is a negative regulator of lincomycin biosynthesis in Streptomyces lincolnensis. Appl Environ Microbiol. 2018;85(1). Print 2019 Jan 1.
Wu P, Pan H, Zhang C, Wu H, Yuan L, Huang X, Zhou Y, Ye BC, Weaver DT, Zhang L, Zhang B. SACE_3986, a TetR family transcriptional regulator, negatively controls erythromycin biosynthesis in Saccharopolyspora erythraea. J Ind Microbiol Biotechnol. 2014;41:1159–67.
Wu H, Chen M, Mao Y, Li W, Liu J, Huang X, Zhou Y, Ye BC, Zhang L, Weaver DT, Zhang B. Dissecting and engineering of the TetR family regulator SACE_7301 for enhanced erythromycin production in Saccharopolyspora erythraea. Microb Cell Factories. 2014;13:158.
Xu Z, Liu Y, Ye BC. PccD regulates branched-chain amino acid degradation and exerts a negative effect on erythromycin production in Saccharopolyspora erythraea. Appl Environ Microbiol. 2018;84.
Ahn SK, Cuthbertson L, Nodwell JR. Genome context as a predictive tool for identifying regulatory targets of the TetR family transcriptional regulators. PLoS One. 2012;7:e50562.
Karnicar K, Drobnak I, Petek M, Magdevska V, Horvat J, Vidmar R, Baebler S, Rotter A, Jamnik P, Fujs S, et al. Integrated omics approaches provide strategies for rapid erythromycin yield increase in Saccharopolyspora erythraea. Microb Cell Factories. 2016;15:93.
Xu JY, Xu Z, Zhou Y, Ye BC. Lysine Malonylome may affect the central metabolism and erythromycin biosynthesis pathway in Saccharopolyspora erythraea. J Proteome Res. 2016;15:1685–701.
Borisova SA, Liu HW. Characterization of glycosyltransferase DesVII and its auxiliary partner protein DesVIII in the methymycin/picromycin biosynthetic pathway. Biochemistry. 2010;49:8071–84.
Zhang B, Tian W, Wang S, Yan X, Jia X, Pierens GK, Chen W, Ma H, Deng Z, Qu X. Activation of natural products biosynthetic pathways via a protein modification level regulation. ACS Chem Biol. 2017;12:1732–6.
van Berkel WJ, Kamerbeek NM, Fraaije MW. Flavoprotein monooxygenases, a diverse class of oxidative biocatalysts. J Biotechnol. 2006;124:670–89.
Liao CH, Xu Y, Rigali S, Ye BC. DasR is a pleiotropic regulator required for antibiotic production, pigment biosynthesis, and morphological development in Saccharopolyspora erythraea. Appl Microbiol Biotechnol. 2015;99:10215–24.
Han S, Song P, Ren T, Huang X, Cao C, Zhang B. Identification of SACE_7040, a member of TetR family related to the morphological differentiation of Saccharopolyspora erythraea. Curr Microbiol. 2011;63:121–5.
Yin X, Xu X, Wu H, Yuan L, Huang X, Zhang B. SACE_0012, a TetR-family transcriptional regulator, affects the morphogenesis of Saccharopolyspora erythraea. Curr Microbiol. 2013;67:647–51.
Kieser T, Buttner M, Chater K, DA H. Practical Streptomyces Genetics. Norwich: The John Innes Foundation; 2000.
Summers RG, Donadio S, Staver MJ, Wendt-Pienkowski E, Hutchinson CR, Katz L. Sequencing and mutagenesis of genes from the erythromycin biosynthetic gene cluster of Saccharopolyspora erythraea that are involved in L-mycarose and D-desosamine production. Microbiology. 1997;143(Pt 10):3251–62.
Sambrook J. RussellDW: Molecular cloning: a laboratory manual. 3rd ed. New York: Cold Spring Harbor Laboratory; 2001.
Wu H, Mao Y, Chen M, Pan H, Huang X, Ren M, Wu H, Li J, Xu Z, Yuan H, et al. Capturing the target genes of BldD in Saccharopolyspora erythraea using improved genomic SELEX method. Appl Microbiol Biotechnol. 2015;99:2683–92.
Hellman LM, Fried MG. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat Protoc. 2007;2:1849–61.
Zianni M, Tessanne K, Merighi M, Laguna R, Tabita FR. Identification of the DNA bases of a DNase I footprint by the use of dye primer sequencing on an automated capillary DNA analysis instrument. J Biomol Tech. 2006;17:103–13.
Livak K, Schmittgenb T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT. Methods. 2001;25:402–8.
Oliynyk M, Samborskyy M, Lester JB, Mironenko T, Scott N, Dickens S, Haydock SF, Leadlay PF. Complete genome sequence of the erythromycin-producing bacterium Saccharopolyspora erythraea NRRL23338. Nat Biotechnol. 2007;25:447–53.
Tarazona S, Garcia-Alcalde F, Dopazo J, Ferrer A, Conesa A. Differential expression in RNA-seq: a matter of depth. Genome Res. 2011;21:2213–23.
Acknowledgements
Not applicable
Funding
This work was supported by the National Natural Science Foundation of China (31300081, 31570074 and 31600064), the National Program on Key Basic Research Project (973 programs, 2013CB734000), Open Fund for Discipline Construction from Institute of Physical Science and Information Technology at Anhui University, and Anhui Provincial Natural Science Foundation (1708085QC49).
Availability of data and materials
The original RNA-seq data have been submitted to the database of NCBI Sequence Read Archive (https://www.ncbi.nlm.nih.gov/sra/SRP129064) under the accession number SRP129064.
Author information
Authors and Affiliations
Contributions
BZ and LZ supervised this study. BZ, LZ, HW and ZC conceived of the study, involved in study design, data interpretation and drafted the manuscript. HW, ZC, CZ, WZ, JN, HF and YC performed the experiments and data analysis. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Table S1. Strains and plasmids used in this study. Table S2. Primers used in this study. Table S3. Putative target genes of SACE_5754 were screened by EMSAs. Figure S1. Analyses of SACE_5754 homologs and comparison of growth rates and morphological differentiation in S. erythraea A226 and relevant strains. Figure S2. Inactivation of SACE_0388 in S. erythraea A226. Figure S3. Inactivation of SACE_3599 in S. erythraea A226. Figure S4. Inactivation of SACE_6149 in S. erythraea A226. Figure S5. Effects of SACE_5754 disruption on transcriptional levels of related genes involving in TCA cycle. Figure S6. Possible regulatory pathway of SACE_5754 on erythromycin biosynthesis in S. erythraea. Figure S7. qRT-PCR analysis of the accuracy of transcriptome analysis. (DOCX 968 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wu, H., Chu, Z., Zhang, W. et al. Transcriptome-guided target identification of the TetR-like regulator SACE_5754 and engineered overproduction of erythromycin in Saccharopolyspora erythraea. J Biol Eng 13, 11 (2019). https://doi.org/10.1186/s13036-018-0135-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13036-018-0135-2