
Abstract
Background
The bicomponent leukotoxins and the pyrogenic toxin superantigens (PTSAgs) are important virulence factors of Staphylococcus aureus. It is necessary to survey the prevalence and expression of these toxin-encoding genes for understanding the possible pathogenic capacity of S. aureus to cause disease.
Methods
Five leukotoxin genes and thirteen PTSAg determinants were detected for 177 S. aureus isolates from blood (n = 88) and wound (n = 89) infections by Polymerase Chain Reaction (PCR). The expression of leukotoxin ED (lukED) was determined by quantitative real-time PCR (qRT-PCR). The genetic backgrounds of isolates were analyzed by Staphylococcal Cassette Chromosome mec (SCCmec) typing (for methicillin-resistant S. aureus isolates), Pulsed-Field Gel Electrophoresis (PFGE), accessory gene regulator (agr) typing and Multilocus Sequence Typing (MLST, for representative isolates based on PFGE type) methods.
Results
99.4% (176/177) isolates contained at least one of leukotoxin genes. Among them, 94.9% (168/177), 81.4% (144/177) and 67.8% (120/177) isolates harbored hlgBC, lukED and lukAB, respectively. Compared to leukotoxin genes, there was a relatively lower overall prevalence of PTSAg genes [99.4% versus 72.9% (129/177), P < 0.001], and they were organized in 59 patterns, with the most common combination of the egc cluster with or without other PTSAg genes. Genetic analysis showed the distributions of certain toxin genes were associated with the genetic backgrounds of isolates. The egc cluster was a common feature of CC5 isolates, among which ST5 and ST764 isolates harbored more PTSAg genes. The lukED was not present in ST398 isolates, and its expression was quite different among isolates. No significant correlations were observed between the lukED expression levels of strains and the ST or agr types.
Conclusions
The present study elucidated the distribution of leukotoxin and PTSAg genes and the expression of lukED in blood and wound isolates, and analyzed the relationship between them with genetic characteristics of isolates. These data improve the current understanding of the possible pathogenicity of S. aureus.
Similar content being viewed by others


Background
Staphylococcus aureus (S. aureus) is a serious pathogen that causes various clinical infections with considerable morbidity and mortality due to its capability to produce different virulence factors [1]. Among these virulence factors, the bicomponent leukotoxins and the pyrogenic toxin superantigens (PTSAgs) have attracted great attention for their ability to destruct the membranes of host cells or regulate the immune responses by activating immune cells abnormally [2,3,4,5].
In S. aureus, seven leukotoxins have been identified. Panton-Valentine leukocidin (PVL), gamma (γ)-hemolysin (HlgAB and HlgBC), leukotoxin ED (LukED), and leukotoxin AB/GH (LukAB/GH) are found in isolates associated with human infections [2, 3]. Leucocidin MFʹ (LukMFʹ) and leucocidin PQ (LukPQ) are only detected in strains from zoonotic infections [6, 7]. Except for pvl (encoding PVL), the data on the overall prevalence of leukotoxin family in clinical S. aureus isolates are very limited in China, especially that of the recently identified LukAB, which is the only leukotoxin known to enhance the survival of S. aureus [2, 3, 8].
Previous epidemiological data showed that lukED existence is widespread among S. aureus isolates [9], and this toxin has an important role in S. aureus bloodstream infection, impetigo and antibiotic-associated diarrhea [3, 10]. Said-Salim et al. [11], Boakes et al. [12] and Yu et al. [13] reported that the production of PVL differentiated from strain to strain, and this difference is associated with the severity of specific infections (such as skin and soft tissue infections, SSTIs). Then, is the expression pattern of lukED among clinical S. aureus isolates the same as that of PVL? Until now, no data can be used to clarify this question.
S. aureus can also secrete an array of pyrogenic toxin superantigens (PTSAgs), including toxic shock syndrome toxin-1 (TSST-1), staphylococcal enterotoxins (SEs), and SE-like toxins. PTSAgs are able to activate T-cells and antigen-presenting cells (APCs) to release proinflammatory cytokines, increase sensitivity to bacterial lipopolysaccharide (LPS) [14], and are associated with some diseases, such as toxic shock syndrome, food poisoning and allergic syndromes [15]. Study also indicates that PTSAgs can play a major role in the pathophysiological mechanism of sepsis [16]. Therefore, it is required to get better understanding of the PTSAg genes distribution in S. aureus isolates from clinical samples.
In this study, we conducted a retrospective study to determine the distribution of genes encoding leukotoxins and PTSAgs among clinical S. aureus isolates obtained from blood and wounds. Subsequently, we analyzed the genetic characteristics of these isolates, and the expression of lukED in some lukED-positive strains. Furthermore, the relationship between genetic backgrounds and the carriage of virulence genes, or the expression level of lukED was assessed.
Methods
Study design and strain identification
Between June 2014 and October 2016, a total of 177 non-duplicate clinical S. aureus isolates (88 strains from blood and 89 isolates from wounds) were separated from six hospitals in, China, namely, Shanghai General Hospital (23 blood isolates and 61 wound isolates, from June 2015 to June 2016), Ruijin Hospital (33 blood isolates, from July 2015 to December 2015), Renji Hospital (15 blood isolates, from January 2016 to March 2016), Shanghai Sixth People′s Hospital (17 blood isolates and 9 wound isolates, from June 2015 to March 2016), Shanghai People′s Hospital of Putuo District (6 wound isolates, from January 2015 to February 2015), and The General Hospital of Lishui City, Zhejiang Province (13 wound isolates, June 2014 to August 2014) (Fig. 1). All of the isolates were identified by VITEK microbiology analyser (bioMérieux, Marcy l′ Etoile, France). Methicillin resistance was confirmed by disk diffusion test with a 30 μg cefoxitin disk (Oxoid, Basingstoke, UK) [17] and Polymerase Chain Reaction (PCR) amplification of mecA and mecC genes (the primers were listed in Additional file 1) [18, 19]. S. aureus Newman was used as a control standard for the amplification and expression of lukED. The quality control strain of disk test was ATCC25923. Methicillin-resistant S. aureus (MRSA) strains NCTC10442, N315, 85/2082, JCSC4744 and D12 [20] were utilized as reference strains for SCCmec type I, II, III, IV and V, respectively.

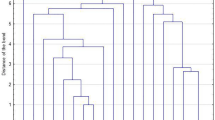
PFGE-based dendrogram showing the genetic relationships of 177 S. aureus isolates. PFGE cluster was assigned by ≥ 80% genetic similarity. The hospital origin, source, methicillin resistance, SCCmec type, ST type, clonal complex, agr type, toxin gene content and the expression of lukED of the isolates were showed. NT, not typeable; wo, wound; bl, blood; egc, seg-sei-sem-sen-seo. mecA +, methicillin-resistant Staphylococcus aureus; mecA-, methicillin-susceptible Staphylococcus aureus; PTSAg, pyrogenic toxin superantigen; lukED expression, the fold change of lukED expression level compared to that of Newman strain, A, Shanghai General Hospital; B, Ruijin Hospital; C, Renji Hospital; D, Shanghai Sixth People’s Hospital; E, Shanghai People’s Hospital of Putuo District; F, The General Hospital of Lishui City, Zhejiang Province
DNA extraction
Suspensions of S. aureus cultures were incubated with lysostaphin (Sangon, Shanghai, China) at 37 °C for 30 min. Then the genomic DNA was extracted from each isolate using TIANamp Bacteria DNA Kit (Tiangen, Beijing, China) following the manufacturer’s instructions, and used as an amplification template for PCR.
Detection of leukotoxin and PTSAg genes
All isolates were screened for the presence of genes encoding leukotoxins (pvl, lukAB, lukED, hlgCB, lukM) and PTSAgs (sea-see, seg-sej, sem-seo and tst) by PCR [21,22,23,24]. Primers used for the amplification of the toxin genes were listed in Additional file 1. One randomly picked PCR product for each gene was sequenced to verify the certainty of target fragment.
Genotyping methods
SCCmec typing
Staphylococcal Cassette Chromosome mec (SCCmec) type I-V were determined by multiplex PCR using the primers (see Additional file 1) derived from the published sequences for MRSA isolates [25].
agr typing
PCR amplification of the accessory gene regulatory (agr) alleles (allele I to IV) was carried out by a previously described method [26] using the primers shown in Additional file 1.
PFGE typing
SmaI-Pulsed-Field Gel Electrophoresis (PFGE) analysis was performed for all isolates to understand their homology as described by McDougal et al. [27]. The patterns of DNA fingerprint were analyzed using BioNumerics sofeware7.0. Percent similarities were identified as described previously [27]. The cluster cutoff was set at 80% similarity.
MLST typing
Multilocus Sequence Typing (MLST) was performed by PCR amplification and sequencing of internal fragments of seven housekeeping genes (arcC, aroE, glpF, gmk, pta, tpi and yqiL) on representative isolates of each PFGE type as previously outlined by Enright et al. [28]. Sequence types (STs) were determined through the MLST website for S. aureus (http://saureus.mlst.net/). The eBURST v.3 [29] was used to classify the related STs into clonal complexes (CCs).
Expression level of lukED at different growth phases
Two clinical S. aureus isolates chosen randomly and well-characterized strain Newman were selected to explore the expression variation of lukED at early, middle and late exponential growth phases at 37 °C in Tryptic Soy Broth (TSB) (Oxoid, Basingstoke, UK). Total RNA was extracted by TaKaRa MiniBEST Universal RNA Extraction Kit (Takara, Dalian, China) and then used for the synthesis of cDNA with reverse transcription Enzyme Mix (Takara, Dalian, China). The expression of lukED was detected by quantitative real-time PCR (qRT-PCR) using SYBR Premix Ex Taq (Takara, Dalian, China) on a 7500 Real-Time PCR system (ABI Biosystems, Cary, USA), as described by previous study [30]. The gene 16s rRNAwas selected as an endogenous control. Specific primers were listed in Additional file 1. The mRNA product of lukED in each strain was standardized to strain Newman. 2-∆∆CT method was applied to analyze the relative expression of each strain.
Transcription level of lukED among isolates with different genetic characteristics
The randomly selected lukED-positive S. aureus isolates [identified further by matrix-assisted laser desorption/ ionization-time of flight mass spectrometry (MALDI-TOF MS, Microflex LT, Bruker Daltonik, Bremen, Germany)] with various PFGE-agr-ST types were normally grown to the post-exponential phase of growth at 37 °C in TSB. The expression of lukED was performed by qRT-PCR as described above.
Statistical analysis
Categorical variables were tested for differences between groups using the Pearson’s Chi-square test or Fisher’s exact test. Kruskal-Wallis test was applied to compare the lukED expression level of S. aureus with different genetic backgrounds. Statistical analysis was computed using IBM SPSS Statistics, Version 23.0 (IBM Corp., Armonk, USA). P values less than 0.05 (two-tailed) were considered to be statistically significant.
Results
MRSA identification and SCCmec typing
Among 177 S. aureus isolates, 47 (26.6%) were identified as MRSA (Fig. 1). The prevalence of MRSA isolates was similar in both blood (31.8%, 28/88) and wound isolates (21.3%, 19/89) (P = 0.149). Four SCCmec types (type II to V) were identified among 39 (83.0%, 39/47) MRSA isolates. An additional 8 (17.0%, 8/47) isolates could not be typed. SCCmec II was the predominant type (40.4%, 19/47), followed by SCCmec IV (25.6%, 12/47) with subtypes IVa (10 isolates) and IVd (2 isolates), SCCmec V (12.8%, 6/47) and SCCmec III (4.3%, 2/47). Within MRSA isolates from blood, SCCmec II was the most prevalent (50%, 14/28), followed by SCCmec IV (32%, 9/28), whereas SCCmec V (32%, 6/19) was the most common in wound MRSA isolates.
agr typing
Four agr types were identified in 168 isolates, and an additional 9 isolates (5 from blood and 4 from wounds) were untypable (Fig. 1). The agr I was the predominant type (63.3%, 112/177), followed by agr II (21.5%, 38/177) and agr III (9.0%, 16/177). Only two isolates from wounds were found to carry agr IV.
PFGE
The PFGE analysis discriminated 169 S. aureus isolates into 50 clonal types, indicating the relatedness of S. aureus isolates from six hospitals was relatively diverse (Fig. 1). Six isolates from wounds and two isolates from blood (consisted of one MRSA and seven MSSA) could not be typed using SmaI restriction enzyme. Overall, the most prevalent clonal type, type 16 (11.9%, 21/177), was identified in 10 blood isolates and 11 wound isolates. The second most frequently type was type 17 (8.4%, 15/177), and was found in 5 strains from blood and 10 strains from wounds, followed by type 33 (7.9%, 14/177), and type 20 and 21 (5.7%, 10/177 each). The remaining 45 types were showed in Fig. 1.
MLST
MLST was performed on 72 representative isolates of each PFGE pattern (randomly selected) and 8 non-PFGE typeable isolates. As shown in Fig. 1, a total of 27 distinct STs were identified. The most common ST type was ST59 (11.3%, 9/80), with 7 strains from blood and 2 from wounds, followed by ST188 and ST398 (10.0%, 8/80 each). Interestingly, 8 non-PFGE typeable isolates were all identified as ST398, including 2 blood isolates and 6 wound isolates (1 MRSA and 7 MSSA). The STs comprised of six isolates (7.5%, 6/80 each) were ST7 and ST764. ST5 and ST630 were found in five isolates (6.3%, 5/80), respectively. In MRSA and MSSA isolates, the most frequently detected ST types were ST59 and ST188, respectively. Moreover, ST59 and ST398 were the predominant types in blood and wound isolates, respectively. By eBURST analysis, CC5 was the most common clone (17.5%, 14/80), followed by CC59 (11.3%, 9/80), CC188 and CC398 (10%, 8/80 each) (Fig. 1).
Distribution of leukotoxin determinants
The data on the prevalence of leukotoxin genes were summarized in Table 1 and Fig. 1. Overall, hlgBC was the most prevalent (94.9%, 168/177), followed by lukED (81.4%, 144/177), lukAB (67.8%, 120/177) and pvl (7.9%, 14/177). All isolates were lukM negative. No significant difference was observed with respect to the overall possession of leukotoxin genes between blood and wound isolates.
With the exception of one MRSA (PFGE1-MRSA-SCCmec NT-agr NT) without toxin gene detected from blood, a total of 176 isolates (99.4%) harbored 446 leukotoxin genes forming 10 distinct leukotoxin gene profiles (Fig. 1). The most common combination was lukED, lukAB plus hlgBC (52.5%, 93/177), followed by lukED plus hlgBC (18.1%, 32/177) and lukAB plus hlgBC (9.6%, 17/177). Among all isolates detected, eight (4 blood and 4 wound isolates, 4.5%, 8/177) harbored the most leukotoxin genes content (lukED, lukAB, hlgBC and pvl), including 3 MRSA (SCCmec IV) and 5 MSSA. Moreover, 13 isolates (7.3%, 13/177) were only hlgBC positive and 5 isolates (2.8%, 5/177) harbored only lukED. The average possession of leukotoxin genes between MRSA and MSSA (2.40 versus 2.56 per strain) was similar, whereas, an analysis on single leukotoxin gene revealed that lukED was significantly more common in MSSA than in MRSA (86.2% versus 68.1%, P = 0.006).
Further analysis indicated that the lukED gene was more frequently possessed by isolates from agr II group than by those from agr I group (94.6% versus 76.8%, P = 0.039), or mainly existed in ST188, ST7, ST764 and ST5 isolates, but not found in ST398 isolates (Table 2). Moreover, ST630 harbored the most frequency of lukAB (100.0%) compared to other STs, and no pvl was detected in ST7, ST764, ST5 and ST630 (Table 2).
Prevalence of PTSAg genes
The distribution of the PTSAg genes was shown in Table 1 and Fig. 1. The seg exhibited a relatively high prevalence (36.7%, 65/177), followed by sei (29.4%, 52/177), sem (28.8%, 51/177), seb (28.2%, 50/177), sen (22.6%, 40/177) and seo (21.5%, 38/177). The positive rates of the other PTSAg genes (sea, sec, sed, seh, sej and tst) were ranged from 7.3 to 18.6%, and no see was found. Statistical analysis showed sen (31.8%, 28/88 versus 13.5%, 12/89, P = 0.001) and seo (28.4%, 25/88 versus 14.6%, 13/89, P = 0.025) were more common in blood isolates than in wound isolates.
129 (72.9%, 129/177) isolates could be detected for possessing at least one of PTSAg genes, of which 41 (31.8%, 41/129, 28 from blood and 13 from wounds) harbored five or more PTSAg genes (high virulence gene content) (Fig. 1). The combination of the complete enterotoxin gene cluster egc (seg, sei, sem, sen, seo), coexisting with or without other PTSAg genes, was the most prevalent enterotoxin gene pattern, presenting in 22 blood isolates and 9 wound isolates (25.0%, 22/88 versus 10.1%, 9/89; P = 0.011). Moreover, the sed-sej combination was only presented in 4.0% (7/177) isolates.
Association between agr groups or major STs and PTSAg genes distribution was displayed in Table 2. The sec was markedly more common in agr II (P = 0.003) and agr III isolates (P = 0.014) than in agr I isolates. The seh was detected more often in isolates with agr III than in those with agr I and agr II (P < 0.001 each). Compared to agr I (P ≤ 0.039) and agr III isolates (P ≤ 0.007), the agr II isolates contain more the seb, seg, sei, sem, sen and seo. The most prevalent egc cluster was more commonly found in isolates belonging to agr II than those with agr I (34.2%, 13/38 versus 13.4%, 15/112, P = 0.004), or present commonly in ST5 (60.0%) and ST764 (50.0%) isolates, but was completely absent in ST59 and ST630 isolates. Additionally, we observed that agr II isolates contained higher number of enterotoxin genes (mean, 4.2) than agr I (mean, 1.7) and agr III isolates (mean, 2.0). ST5 and ST764 isolates carried higher frequency of PTSAg genes (mean, 6 and 5, respectively) compared to other STs. The seb was mainly present in ST59 strains (77.8%), followed by ST764 (66.7%) and ST188 (50.0%). In addition, the sec (100.0%) and tst (80.0%) were mainly found in ST5 strains. No enterotoxin gene was detected in ST630 isolates.
Expression level of lukED among isolates with various genetic background
In the present study, our data confirmed the S. aureus isolates produced the highest expression amount of lukED at the late exponential growth phase (data not shown). Therefore, we chose this stage to study the expression of lukED in clinical isolates. Of the 63 lukED-positive S. aureus isolates typed by MLST, 46 selected randomly were grown to the late exponential growth phase for analyzing the mRNA expression of lukED. As shown in Fig. 1, an up to 260-fold difference was observed between the highest and the lowest lukED expressions strains, with the exception of two isolates expressing extremely low level of lukED [26× and 28×, both were PFGE39-agrI-CC20 (ST20/ST1281) blood isolates]. Compared to lukED expression level of strain Newman, 54.3% (25/46) isolates was 2 folds higher, whereas 41.3% (19/46) isolates was 2 folds lower (Fig. 1). Of note, two strains from wounds with the highest transcriptional levels of lukED all belonged to PFGE16-agrI-ST7, which were 23.0 (isolate 123) and 11.5 (isolate 752b) folds higher than that of Newman strain, respectively (Fig. 1). However, no significant differences were observed in the overall expression levels of lukED among the major ST types (ST5, ST7, ST188, ST630 and ST764, P = 0.072) or agr groups (agr I, agr II and agr III, P = 0.718) (Fig. 1).
Discussion
Blood and wound S. aureus infections are common clinical diseases. Therefore, we investigated some major toxin (such as leukotoxin and PTSAg) genes existence for getting insight into the potential pathogenic ability of S. aureus from the two kinds of samples.
Although previous studies reported that each member of the leukotoxins has its distinct role in the pathogenesis of S. aureus by both in vitro and in vivo investigation [10, 31,32,33,34,35], to the best of our knowledge, this is the first study on the overall prevalence of this toxin family among clinical S. aureus isolates in China. Our data showed the prevalent rates of hlgBC (94.9%) and lukED (81.4%) were similar to those of previous reports [33, 36,37,38,39,40,41]. The lukAB, whose distribution is unknown due to lack of investigation in a large number of clinical strains, was carried by 67.8% of our isolates. There is a varying carriage of pvl among MRSA, ranging from 2.3 to 50.7% in China [42,43,44,45]. In the present study, a relatively low prevalence of pvl-positive isolates (7.9%) was found, which was in agreement with our previous data (6.6%) [46].
Regarding the PTSAg genes, Dramann et al. [5] has reviewed that approximately 80% of clinical S. aureus isolates carry an average of 5 to 6 genes, and the gene profiles varied remarkably among S. aureus strains. In this study, the overall positive rate of PTSAg genes was 72.9%, and a total of 59 PTSAg gene combinations were observed. However, due to only 13 PTSAg genes detected here, a much lower average carriage (mean, 3.3, 420/129) was found in the PTSAg gene-positive strains. A study from China showed sea was the most prevalent enterotoxin gene (41.53%) in S. aureus isolates from bacteraemia [47]. However, the positive rate of sea was only 15.9% in our blood strains, and had no significant difference between the strains from blood and wounds (15.9% versus 21.3%) (Table 1). This discrepancy is most likely caused by the difference in genetic backgrounds of strains [47, 48]. Although enterotoxin gene cluster egc has no connection with life-threatening infections, the possession of this operon may be conducive to the colonization of S. aureus and function in certain infections [49,50,51]. The total prevalence rate of 17.5% for the intact egc in this study (Fig. 1) resembled the results observed by Xie et al. [52] and Chao et al. [53]. However, we found a significant difference (24.7% versus 10.2%, P = 0.011) of this gene cluster carriage in our isolates from blood and wounds (Fig. 1), which indicated this cluster might link to the origins of isolates. Usually, sed-sej is located on plasmid pIB485, and the coexistence of both genes has been reported in some studies [53, 54]. Here, the fixed combination was only detected in 4% isolates, and 10.7% (19/177) isolates possessed sej or sed (Fig. 1). This uncharacterized combination of toxin genes indicated the diversity of yet-undescribed variants of mobile genetic element (MGE).
Previous studies presented that CC clones of S. aureus often display different toxin gene patterns [48, 53, 55]. For example, the toxin locus of lukED was present in CC1, CC5 and CC7 etc., but completely absent from CC22, CC30 and CC398 etc. [53, 55]. In this study, the distribution of lukED in CC isolates was basically in line with the previous reports, except 1 isolate with CC22 and 1 isolate with CC30 (Fig. 1). Previous data indicated that the lukED is located on a mobile pathogenicity island, vSaβ [5]. Therefore, we speculated the isolates with CC22 and CC30 obtained the lukED through the horizontal transfer of the vSaβ. Apart from lukED negative, ST398 isolates also harbor fewer PTSAg genes [56, 57]. This phenomenon was confirmed by our data in Fig. 1. Previous data indicated that the egc cluster was a common feature of CC5 isolates [58, 59]. The same phenomenon was found in our isolates (Fig. 1). γ-hemolysin, a core genome-encoded leukotoxin, is highly conserved [3], and therefore can be detected in nearly all our S. aureus strains. Although lukAB is also located in the core genome, its locus is often disrupted by the insertion of a prophage [3]. This may explain the relatively lower prevalence of lukAB among each S. aureus lineage in this study, compared to that of γ-hemolysin-encoding gene hlgCB (Table 2, Fig. 1). Particular association had been observed between LukED-producing strains and agr II, as well as for TSST-1 and agr III isolates [21, 36]. However, our data only showed the correlation between lukED and agr II isolates. Besides, Fig. 1 displayed the PTSAg genes were preferable more common in agr II isolates. These carriage differences of toxin genes among isolates with different genetic backgrounds might be related to the heterogeneous nature of the infections and patients. In this study, the total number of isolates is not particularly large, which leaded to a relative small proportion (26.6%, 47/177) of MRSA strains and a few MRSA clones (Table 1 and Fig. 1). The asymmetrical distribution may influence the objective distribution of virulence determinants. This is one of the limitations of the present study.
The expression differences of immune evasion genes among strains may have vital influence for the pathogenesis of bacteria [55]. Previous studies exhibited that LukED plays an essential role in S. aureus infections [3, 10]. Therefore, we detected the transcription level of this leukotoxin in clinical isolates. The results of qRT-PCR revealed a marked strain-to-strain variation in lukED mRNA transcription, even in isolates with the same genetic characteristics. Consistent with the study on the production of PVL [12], we observed that agr types did not affect the lukED expressions significantly (Fig. 1). In addition, no remarkable associations were observed between major STs and the expression of this gene. Because only a limited number of ST or agr type isolates were included in this study, the different expressions of lukED attributed to various ST types or agr groups can’t be ruled out. And more comprehensive investigations of abundant isolates are needed to explore the association between lukED expression and different genetic background. In order to better verify the toxin’s role in bacterial pathogenesis, it is very important to study the relationship between the toxin expression level and the disease severity. In this study, we conducted a retrospective investigation designed only to understand the expression of lukED in S. aureus isolates from blood and wounds. If the correlation of the expression levels of lukED with the severity of infectious diseases is evaluated, it will provide a more convincing evidence to elucidate the function of LukED in the pathogenesis of S. aureus. This will be the research interest of the future study.
Conclusions
In summary, this work exhibits the prevalence of leukotoxin and partial PTSAg genes in clinical S. aureus isolates from blood and wound in eastern region hospitals of China. Genotypic analysis illustrates a high genetic diversity of these isolates, and certain toxin genes, such as lukED and the egc cluster, may be lineage specific. In particular, S. aureus isolates belonging to ST764 and ST5, as well as agr II, were likely to harbor more PTSAg genes, which may suggest an alarming situation of infected patients. The different expression of lukED was found in clinical isolates, however, the association of this difference with the genetic backgrounds of isolates needs to be further unraveled.
Abbreviations
- agr :
-
Accessory gene regulator
- CC:
-
Clonal complex
- CCs:
-
Clonal complexes
- HlgAB and HlgBC:
-
Gamma (γ)-hemolysin
- LPS:
-
Lipopolysaccharide
- LukAB/GH:
-
Leukotoxin AB/GH
- LukED:
-
Leukotoxin ED
- LukMFʹ:
-
Leucocidin MFʹ
- LukPQ:
-
Leucocidin PQ
- MGE:
-
Mobile genetic element
- MLST:
-
Multilocus sequence typing
- MRSA:
-
Methicillin-resistant Staphylococcus aureus
- PCR:
-
Polymerase chain reaction
- PFGE:
-
Pulsed-field gel electrophoresis
- PTSAgs:
-
Pyrogenic toxin superantigens
- PVL:
-
Panton-Valentine leucocidin
- qRT-PCR:
-
Quantitative real-time PCR
- S. aureus :
-
Staphylococcus aureus
- SCCmec :
-
Staphylococcal cassette chromosome mec
- SEs:
-
Staphylococcal enterotoxins
- STs:
-
Sequence types
- TSST-1:
-
Toxic shock syndrome toxin-1
References
Benfield T, Espersen F, Frimodt-Moller N, Jensen AG, Larsen AR, Pallesen LV, et al. Increasing incidence but decreasing in-hospital mortality of adult Staphylococcus aureus bacteraemia between 1981 and 2000. Clin Microbiol Infect. 2007;13(3):257–63.
Yoong P, Torres VJ. The effects of Staphylococcus aureus leukotoxins on the host: cell lysis and beyond. Curr Opin Microbiol. 2013;16(1):63–9.
Alonzo F 3rd, Torres VJ. The bicomponent pore-forming leucocidins of Staphylococcus aureus. Microbiol Mol Biol Rev. 2014;78(2):199–230.
Spaulding AR, Salgado-Pabon W, Kohler PL, Horswill AR, Leung DY, Schlievert PM. Staphylococcal and streptococcal superantigen exotoxins. Clin Microbiol Rev. 2013;26(3):422–47.
Grumann D, Nubel U, Broker BM. Staphylococcus aureus toxins--their functions and geetics. Infect Genet Evol. 2014;21:583–92.
Koop G, Vrieling M, Storisteanu DM, Lok LS, Monie T, van Wigcheren G, et al. Identification of lukPQ, a novel, equid-adapted leukocidin of Staphylococcus aureus. Sci Rep. 2017;7:40660.
Spaan AN, van Strijp JAG, Torres VJ. Leukocidins: staphylococcal bi-component pore-forming toxins find their receptors. Nat Rev Microbiol. 2017;15(7):435–47.
Dumont AL, Nygaard TK, Watkins RL, Smith A, Kozhaya L, Kreiswirth BN, et al. Characterization of a new cytotoxin that contributes to Staphylococcus aureus pathogenesis. Mol Microbiol. 2011;79(3):814–25.
Seilie ES, Bubeck WJ. Staphylococcus aureus pore-forming toxins: the interface of pathogen and host complexity. Semin Cell Dev Biol. 2017;72:101–16.
Alonzo F 3rd, Benson MA, Chen J, Novick RP, Shopsin B, Torres VJ. Staphylococcus aureus leucocidin ED contributes to systemic infection by targeting neutrophils and promoting bacterial growth in vivo. Mol Microbiol. 2012;83(2):423–35.
Said-Salim B, Mathema B, Braughton K, Davis S, Sinsimer D, Eisner W, et al. Differential distribution and expression of Panton-valentine leucocidin among community-acquired methicillin-resistant Staphylococcus aureus strains. J Clin Microbiol. 2005;43(7):3373–9.
Boakes E, Kearns AM, Badiou C, Lina G, Hill RL, Ellington MJ. Do differences in Panton-valentine leukocidin production among international methicillin-resistant Staphylococcus aureus clones affect disease presentation and severity? J Clin Microbiol. 2012;50(5):1773–6.
Yu F, Liu Y, Xu Y, Shang Y, Lou D, Qin Z, et al. Expression of Panton-valentine leukocidin mRNA among Staphylococcus aureus isolates associates with specific clinical presentations. PLoS One. 2013;8(12):e83368.
Stach CS, Herrera A, Schlievert PM. Staphylococcal superantigens interact with multiple host receptors to cause serious diseases. Immunol Res. 2014;59(1–3):177–81.
Argudin MA, Mendoza MC, Vazquez F, Rodicio MR. Exotoxin gene backgrounds in bloodstream and wound Staphylococcus aureus isolates from geriatric patients attending a long-term care Spanish hospital. J Med Microbiol. 2011;60(Pt 11):1605–12.
Azuma K, Koike K, Kobayashi T, Mochizuki T, Mashiko K, Yamamoto Y. Detection of circulating superantigens in an intensive care unit population. Int J Infect Dis. 2004;8(5):292–8.
Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing, twenty-fifth informational supplement; M100-S25. Wayne: Clin Lab Stand Institute; 2015.
Bignardi GE, Woodford N, Chapman A, Johnson AP, Speller DC. Detection of the mec-A gene and phenotypic detection of resistance in Staphylococcus aureus isolates with borderline or low-level methicillin resistance. J Antimicrob Chemother. 1996;37(1):53–63.
Ganesan A, Crawford K, Mende K, Murray CK, Lloyd B, Ellis M, et al. Evaluation for a novel methicillin resistance (mecC) homologue in methicillin-resistant Staphylococcus aureus isolates obtained from injured military personnel. J Clin Microbiol. 2013;51(9):3073–5.
Han LZ, Ho PL, Ni YX, Zhang H, Jiang YQ, Chu HQ, et al. Panton-Valentine leukocidin-positive MRSA, Shanghai. Emerg Infect Dis. 2010;16(4):731–3.
Jarraud S, Mougel C, Thioulouse J, Lina G, Meugnier H, Forey F, et al. Relationships between Staphylococcus aureus genetic background, virulence factors, agr groups (alleles), and human disease. Infect Immun. 2002;70(2):631–41.
Lina G, Piemont Y, Godail-Gamot F, Bes M, Peter MO, Gauduchon V, et al. Involvement of Panton-valentine leukocidin-producing Staphylococcus aureus in primary skin infections and pneumonia. Clin Infect Dis. 1999;29(5):1128–32.
Monday SR, Bohach GA. Use of multiplex PCR to detect classical and newly described pyrogenic toxin genes in staphylococcal isolates. J Clin Microbiol. 1999;37(10):3411–4.
McLauchlin J, Narayanan GL, Mithani V, O'Neill G. The detection of enterotoxins and toxic shock syndrome toxin genes in Staphylococcus aureus by polymerase chain reaction. J Food Prot. 2000;63(4):479–88.
Zhang K, McClure JA, Elsayed S, Louie T, Conly JM. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J Clin Microbiol. 2005;43(10):5026–33.
Lina G, Boutite F, Tristan A, Bes M, Etienne J, Vandenesch F. Bacterial competition for human nasal cavity colonization: role of staphylococcal agr alleles. Appl Environ Microbiol. 2003;69(1):18–23.
McDougal LK, Steward CD, Killgore GE, Chaitram JM, McAllister SK, Tenover FC. Pulsed-field gel electrophoresis typing of oxacillin-resistant Staphylococcus aureus isolates from the United States: establishing a national database. J Clin Microbiol. 2003;41(11):5113–20.
Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–15.
Feil EJ, Li BC, Aanensen DM, Hanage WP, Spratt BG. Eburst: inferring patterns of evolutionary descent among clusters of related bacterial genotypes from multilocus sequence typing data. J Bacteriol. 2004;186(5):1518–30.
Balasubramanian D, Ohneck EA, Chapman J, Weiss A, Kim MK, Reyes-Robles T, et al. Staphylococcus aureus coordinates leukocidin expression and pathogenesis by sensing metabolic fluxes via RpiRc. MBio. 2016;7(3):e00818–6.
DuMont AL, Yoong P, Day CJ, Alonzo F 3rd, McDonald WH, Jennings MP, et al. Staphylococcus aureus lukAB cytotoxin kills human neutrophils by targeting the CD11b subunit of the integrin mac-1. Proc Natl Acad Sci U S A. 2013;110(26):10794–9.
Reyes-Robles T, Alonzo F 3rd, Kozhaya L, Lacy DB, Unutmaz D, Torres VJ. Staphylococcus aureus leukotoxin ED targets the chemokine receptors CXCR1 and CXCR2 to kill leukocytes and promote infection. Cell Host Microbe. 2013;14(4):453–9.
Spaan AN, Vrieling M, Wallet P, Badiou C, Reyes-Robles T, Ohneck EA, et al. The staphylococcal toxins gamma-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat Commun. 2014;5:5438.
Melehani JH, James DB, DuMont AL, Torres VJ, Duncan JA. Staphylococcus aureus leukocidin a/B(lukAB) kills human monocytes via host NLRP3 and ASC when extracellular, but not intracellular. PLoS Pathog. 2015;11(6):e1004970.
Diep BA, Le VT, Visram ZC, Rouha H, Stulik L, Dip EC, et al. Improved protection in a rabbit model of community-associated methicillin-resistant Staphylococcus aureus necrotizing pneumonia upon neutralization of leukocidins in addition to alpha-hemolysin. Antimicrob Agents Chemother. 2016;60(10):6333–40.
von Eiff C, Friedrich AW, Peters G, Becker K. Prevalence of genes encoding for members of the staphylococcal leukotoxin family among clinical isolates of Staphylococcus aureus. Diagn Microbiol Infect Dis. 2004;49(3):157–62.
Gravet A, Colin DA, Keller D, Girardot R, Monteil H, Prevost G. Characterization of a novel structural member, lukE-lukD, of the bi-component staphylococcal leucotoxins family. FEBS Lett. 1998;436(2):202–8.
Gravet A, Rondeau M, Harf-Monteil C, Grunenberger F, Monteil H, Scheftel JM, Prevost G. Predominant Staphylococcus aureus isolated from antibiotic-associated diarrhea is clinically relevant and produces enterotoxin a and the bicomponent toxin lukE-lukD. J Clin Microbiol. 1999;37(12):4012–9.
Gravet A, Couppie P, Meunier O, Clyti E, Moreau B, Pradinaud R, et al. Staphylococcus aureus isolated in cases of impetigo produces both epidermolysin a or B and lukE-lukD in 78% of 131 retrospective and prospective cases. J Clin Microbiol. 2001;39(12):4349–56.
Morinaga N, Kaihou Y, Noda M. Purification, cloning and characterization of variant lukE-lukD with strong leukocidal activity of staphylococcal bi-component leukotoxin family. Microbiol Immunol. 2003;47(1):81–90.
Diep BA, Carleton HA, Chang RF, Sensabaugh GF, Perdreau-Remington F. Roles of 34 virulence genes in the evolution of hospital- and community-associated strains of methicillin-resistant Staphylococcus aureus. J Infect Dis. 2006;193(11):1495–503.
Yu F, Chen Z, Liu C, Zhang X, Lin X, Chi S, et al. Prevalence of Staphylococcus aureus carrying Panton-valentine leukocidin genes among isolates from hospitalised patients in China. Clin Microbiol Infect. 2008;14(4):381–4.
Liu Y, Wang H, Du N, Shen E, Chen H, Niu J, et al. Molecular evidence for spread of two major methicillin-resistant Staphylococcus aureus clones with a unique geographic distribution in Chinese hospitals. Antimicrob Agents Chemother. 2009;53(2):512–8.
Bakthavatchalam YD, Nabarro LEB, Ralph R, Veeraraghavan B. Diagnosis and management of panton-valentine leukocidin toxin associated Staphylococcus aureus infection: an update. Virulence. 2017;7:0.
Qin Y, Wen F, Zheng Y, Zhao R, Hu Q, Zhang R. Antimicrobial resistance and molecular characteristics of methicillin-resistant Staphylococcus aureus isolates from child patients of high-risk wards in Shenzhen, China. Jpn J Infect Dis. 2017;70(5):479–84.
Zhao H, Hu F, Jin S, Xu X, Zou Y, Ding B, et al. Typing of Panton-valentine leukocidin-encoding phages and lukSF-PV gene sequence variation in Staphylococcus aureus from China. Front Microbiol. 2016;7:1200.
He W, Chen H, Zhao C, Zhang F, Li H, Wang Q, et al. Population structure and characterisation of Staphylococcus aureus from bacteraemia at multiple hospitals in China: association between antimicrobial resistance, toxin genes and genotypes. Int J Antimicrob Agents. 2013;42(3):211–9.
Yu F, Li T, Huang X, Xie J, Xu Y, Tu J, et al. Virulence gene profiling and molecular characterization of hospital-acquired Staphylococcus aureus isolates associated with bloodstream infection. Diagn Microbiol Infect Dis. 2012;74(4):363–8.
van Belkum A, Melles DC, Snijders SV, van Leeuwen WB, Wertheim HF, Nouwen JL, et al. Clonal distribution and differential occurrence of the enterotoxin gene cluster, egc, in carriage- versus bacteremia-associated isolates of Staphylococcus aureus. J Clin Microbiol. 2006;44(4):1555–7.
Morgan WR, Caldwell MD, Brady JM, Stemper ME, Reed KD, Shukla SK. Necrotizing fasciitis due to a methicillin-sensitive Staphylococcus aureus isolate harboring an enterotoxin gene cluster. J Clin Microbiol. 2007;45(2):668–71.
Liu Q, Han L, Li B, Sun J, Ni Y. Virulence characteristic and MLST-agr genetic background of high-level mupirocin-resistant, MRSA isolates from shanghai and Wenzhou, China. PloS one. 2012;7(5):e37005. https://doi.org/10.1371/journal.pone.0037005.
Xie Y, He Y, Gehring A, Hu Y, Li Q, Tu SI, et al. Genotypes and toxin gene profiles of Staphylococcus aureus clinical isolates from China. PLoS One. 2011;6(12):e28276.
Chao G, Bao G, Cao Y, Yan W, Wang Y, Zhang X, et al. Prevalence and diversity of enterotoxin genes with genetic background of Staphylococcus aureus isolates from different origins in China. Int J Food Microbiol. 2015;211:142–7.
Becker K, Friedrich AW, Lubritz G, Weilert M, Peters G, Von Eiff C. Prevalence of genes encoding pyrogenic toxin superantigens and exfoliative toxins among strains of Staphylococcus aureus isolated from blood and nasal specimens. J Clin Microbiol. 2003;41(4):1434–9.
McCarthy AJ, Lindsay JA. Staphylococcus aureus innate immune evasion is lineage-specific: a bioinfomatics study. Infect Genet Evol. 2013;19:7–14.
Kadlec K, Ehricht R, Monecke S, Steinacker U, Kaspar H, Mankertz J, et al. Diversity of antimicrobial resistance pheno- and genotypes of methicillin-resistant Staphylococcus aureus ST398 from diseased swine. J Antimicrob Chemother. 2009;64(6):1156–64.
Fessler A, Scott C, Kadlec K, Ehricht R, Monecke S, Schwarz S. Characterization of methicillin-resistant Staphylococcus aureus ST398 from cases of bovine mastitis. J Antimicrob Chemother. 2010;65(4):619–25.
Fessler AT, Kadlec K, Hassel M, Hauschild T, Eidam C, Ehricht R, et al. Characterization of methicillin-resistant Staphylococcus aureus isolates from food and food products of poultry origin in Germany. Appl Environ Microbiol. 2011;77(20):7151–7.
Monecke S, Coombs G, Shore AC, Coleman DC, Akpaka P, Borg M, et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS One. 2011;6(4):e17936.
Acknowledgements
The authors wish to thank Lizhong Han, Min Li, Jin Tang, Jinwei Huang and Jianmin Ren for providing the clinical S. aureus isolates from Shanghai City and Zhejiang Province.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No.81772247 and No.81371872).
Availability of data and materials
All the data on which the conclusions of this study rely are presented in Fig. 1, Table 1 and Table 2.
Author information
Authors and Affiliations
Contributions
QL designed and conceived the investigation; HZ, FH, XX, SJ, HY and FG gathered the clinical isolates; CH, SX and HZ carried out the experiments; CH, SX, FH, XX, SJ, HY and FG collected the data and did the statistical analysis. CH, SX and QL wrote the paper. CH and QL revised the manuscript. All authors critically reviewed the manuscript and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This is a retrospective study. The strains used here were all preserved strains taken as part of routine care from clinical laboratories. Since no animal or human data were used in this study, the ethics committee of the Shanghai General Hospital ruled that no formal ethics approval was required in this particular case.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Sequences of primers used for PCR in this study. (DOCX 41 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
He, C., Xu, S., Zhao, H. et al. Leukotoxin and pyrogenic toxin Superantigen gene backgrounds in bloodstream and wound Staphylococcus aureus isolates from eastern region of China. BMC Infect Dis 18, 395 (2018). https://doi.org/10.1186/s12879-018-3297-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-018-3297-0