
Abstract
Background
Quick and complete recovery of cognitive function after general anesthesia is desirable, particularly for working-age patients. Desflurane is less likely to have long-term effects than older-generation inhalational anesthetics, however, its short-term effects have not been fully investigated. Our objective was to elucidate the short-term effects of desflurane exposure on learning and memory in young adult rats.
Methods
Seven-week old male Sprague–Dawley rats were exposed to air (control), or desflurane at 0.7 or 1.2 minimum alveolar concentration (MAC) for 2 h (day 0). The inhibitory avoidance (IA) test was performed on day 1 to delineate the effects on contextual learning. Separate groups of control and 1.2 MAC desflurane animals underwent the IA test on days 3 and 7 to examine the time-dependent changes. Because the IA test is known to be dependent on the long-term potentiation (LTP) of the hippocampus and the trafficking of the GluR1 subunit of the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor into the synapses, the effects of 1.2 MAC desflurane on these phenomena were evaluated on day 1.
Results
Desflurane at 1.2 MAC, but not 0.7 MAC, significantly decreased the IA latencies on day 1 compared with the control (one-way ANOVA, F [2,48] = 5.974, P = 0.005, post hoc Tukey’s, mean difference [95% confidence interval], control vs. 1.2 MAC, 168 [49.9 to 287], P = 0.004; control vs. 0.7 MAC, 67.5 [− 51.2 to 186], P = 0.362). The latencies were not affected on days 3 and 7 (day 3, control vs. desflurane, P = 0.861; day 7, control vs. desflurane, P > 0.999). Consistently, hippocampal LTP on day 1 was significantly suppressed in the desflurane group compared with the control group (P = 0.006). Moreover, immunoblotting analysis of synaptic GluR1 expression revealed that desflurane exposure significantly suppressed GluR1 delivery to the synapses after IA training.
Conclusion
Exposure to a relatively high concentration of desflurane caused reversible learning and memory impairment in young adult rats associated with suppression of GluR1 delivery to the synapses in the hippocampus.
Similar content being viewed by others


Background
Postoperative cognitive dysfunction (POCD) occurs in not only aged adults but adults of all ages [1], and is known to decrease the levels of daily activity [2], increase mortality, and the risk of leaving the labor market [3]. Although the underlying mechanisms have not been completely elucidated, there is emerging evidence from both clinical trials [4,5,6,7,8,9] and experimental animal studies [10,11,12,13,14,15,16] suggesting the contribution of inhalational anesthetics to POCD.
We previously reported that isoflurane impairs hippocampal learning 7 days after exposure in young adult rats [15]. In contrast to isoflurane, desflurane is reported to induce neither neurotoxicity [16,17,18,19,20,21] nor clinical cognitive dysfunction [4], and is less likely to induce long-term effects on learning and memory in rats [14]. However, regarding cognitive function, these studies investigated the effects 48 h [4] or 1 week or longer [14] after the anesthetic exposure and the shorter-term effects have not been fully studied. Because rapid and complete recovery of cognitive function is highly desirable especially in working-age patients and in ambulatory settings, we considered it meaningful to evaluate the shorter-term effects of desflurane.
The aims of the present study were two-fold. First, we sought to elucidate the short-term (no longer than 7-day) effects of desflurane on learning and memory in young adult rats, by using the inhibitory avoidance (IA) test, a widely used and robust behavioral test of contextual learning. Second, because the IA test is known to be dependent on hippocampal long-term potentiation (LTP), and the synaptic increment of the GluR1 subunit of α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPAR) in the hippocampus [22, 23], we sought to elucidate the effects of desflurane on these phenomena. We previously demonstrated that isoflurane impairs IA learning and LTP, and modulates synaptic GluR1 7 days after the exposure [15]. Our hypothesis was that desflurane would have similar effects, which would occur earlier.
Methods
Subjects
All experiments were conducted with approval from the Institutional Animal Care and Use Committee of the Animal Research Center at the Yokohama City University Graduate School of Medicine, Yokohama, Japan (approval number: F-A-14-038, F-A-17-079). Care and experiments of animals were performed in accordance with the Guidelines for Proper Conduct of Animal Experiments (2006, Science Council of Japan), and the study adhered to the Animal Research: Reporting In Vivo Experiments guidelines.
Male Sprague–Dawley rats (Japan-SLC, Shizuoka, Japan), aged 7 weeks old, weighing 200–270 g (total n = 170) were used in the study. Rats were housed in a temperature- and humidity-controlled room in a 12-h light-dark cycle, with ad libitum water and food. All rats were purchased at least 1 week before they were used.
Anesthesia
On day 0, rats were randomly allocated to the control or desflurane-exposure group for each experiment. Anesthesia was induced and maintained for 2 h in a translucent plastic chamber (length, 30 cm; width, 43 cm; height, 14 cm) with 9.6% or 5.6% desflurane, which corresponds to 1.2 or 0.7 minimum alveolar concentration (MAC) [14, 24], respectively, continuously flushed with a carrier gas consisting of oxygen and nitrogen (FIO2 = 0.40–0.45). Rats were allowed to breathe spontaneously; however CO2 level in the container was maintained at less than 3 mmHg. The fraction of inspiratory and end tidal gas concentration was monitored continuously with Capnomac ULTIMA monitor (Datex, Helsinki, Finland).
Heart rate (HR) and oxygen saturation (SaO2) were measured continuously during anesthesia with a Mouse OX™ Pulse Oximeter (Harvard Apparatus, Holliston, MA, USA). Rectal temperature was maintained at 37 ± 1 °C using a thermostatic bath (36 ± 1 °C).
As for the control group, rats were placed in the plastic chamber and flushed with the same carrier gas for 1 min. They were only exposed for 1 min, with the intention of exposing them to the same stress as that experienced by the desflurane-exposure rats before being anaesthetized.
Physiological variables
Rats (n = 5) of separate groups were anesthetized with 1.2 MAC desflurane for 2 h under the same conditions as described in the previous paragraph, with a catheter inserted into the carotid artery to continuously monitor blood pressure. At the end, blood samples were collected in heparinized syringes and were immediately analyzed using the ABL800 FLEX blood gas analyzer (Radiometer, Tokyo, Japan) to confirm the values of pH, PaCO2, PaO2, and HCO3−. These rats were not used in any further studies.
IA test
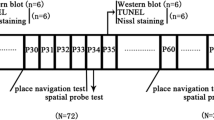
The IA test, a widely used behavioral task to evaluate hippocampus-dependent contextual learning, was first performed to delineate the effect of exposure to desflurane on day 1. Rats were randomly allocated to the control, 0.7 MAC, or 1.2 MAC desflurane-exposure group (n = 17 each), considering the concentration dependency. An identical apparatus (length, 27 cm; width, 45 cm; and height, 25 cm) to that used in the previous study [15], where lighted and dark shock boxes are separated by a trap door and placed in a sound-shielded room, was used. Next, the test was also performed on days 3 and 7 with the control or 1.2 MAC desflurane-exposure group (n = 16 and n = 15 each, respectively) to examine time-dependent changes.
On day 1, the training session was performed. Each rat was initially placed in the lighted box and allowed to explore for 30 s. The trap door was silently opened, and soon after the rat went into the dark box, the door was closed. Subsequently, a scrambled electrical foot shock (1 s, 0.4 mA) was applied via the electrified floor using an SG-1000 shock generator (Melquest, Toyama, Japan). Fifteen seconds later, the rat was returned to its original cage. On day 2, the retention trial was performed. Each rat was again placed in the lighted box, and its latency to enter the dark box with all four paws was measured (maximum cut-off latency of 360 s). Longer retention test latencies are interpreted as better memory. The experimental time line is shown in Fig. 1.

Experimental timeline of IA test
All experiments were performed during the light phase (09:00–14:00) and were recorded with a video camera. The experimenter was blinded to the rat’s allocation group. Each rat was tested only once and euthanized with isoflurane and intraperitoneal pentobarbital after the test.
Electrophysiological recordings
Separate groups of rats were anesthetized with 1.2 MAC desflurane (n = 7) or were handled as controls (n = 5) for brain slice preparation. Prior to decapitation, rats were briefly anesthetized with isoflurane for ethical reason, although brief anesthesia for surgical preparations or euthanasia has been reported to influence brain [25, 26]. Rat brains were rapidly removed and cut out to 300 μm coronal slices containing the dorsal hippocampus (Leika VT1000S; Leika Microsystems, Tokyo, Japan) in ice-cold dissection buffer. Subsequently, slices were transferred to artificial cerebrospinal fluid (ACSF; 22–25 °C), and were incubated at room temperature for 120 min. The dissection buffer and ACSF were bubbled (95% O2, 5% CO2) throughout the procedure.
The prepared brain slice was placed on an 8 × 8 array of planar microelectrodes (MED-P515A, Alpha MED Scientific, Osaka, Japan). Extracellular field excitatory postsynaptic potentials (fEPSPs) of the CA1 area, which were evoked by stimulating Schaffer collaterals, were acquired using the multi-electrode MED64 system (Alpha MED Scientific, Osaka, Japan). After 30 min of test stimulation (half maximal shock, every 60 s) to assess baseline stability, LTP was induced by applying high-frequency stimulation (100 Hz, 1 s), followed by test stimulation for 40 min. The LTP was evaluated by the mean slope of fEPSPs at 36–40 min from the high-frequency stimulation. Slices were continuously perfused with bubbled (95% O2, 5% CO2) ACSF throughout the experiment. Each slice was used only once. Although the experimenter was aware of the rats’ group allocation when performing the recordings, the data analyst was blinded.
Quantitative immunoblotting
Separate groups of rats were anesthetized with 1.2 MAC desflurane (n = 20) or were handled as controls (n = 20) for immunoblot analysis of the AMPAR GluR1 subunit. Half rats of each group were IA-trained and the other half of each group were placed in a sound-shielded room but not IA-trained. Thirty minutes after the IA training or control condition (untrained), hippocampal tissue samples were promptly dissected using ice-cold dissection buffer.
Synaptoneurosomes, which are fractions enriched in synaptic elements, were prepared as previously described [22, 23, 27] . Immunoblotting was performed using primary antibodies against GluR1 (1:1000, Millipore, Temecula, CA, USA) and GAPDH (1:5000, Sigma-Aldrich, St Louis, MO, USA). All immunoblots and densitometry were performed with the experimenter blinded to the groups.
Statistical analyses
The primary outcome was the IA latencies on day 1. The sample size of 17 per group was calculated for the three groups to be compared with a type 1 error of 0.05 and a power of 80% to detect a mean difference of 170 s, assuming the standard deviation to be 150 based on a pilot study where naive rats were IA trained. The normality of distribution was assessed with the Shapiro–Wilk test. For the primary outcome, the data were analyzed using one-way factorial analysis of variance (ANOVA) using desflurane concentration as the one factor followed by post hoc Tukey’s multiple comparison tests. To validate the time-dependent changes, the IA data were analyzed using two-way ANOVA using group and time point (day) as the two factors followed by post hoc Sidak’s multiple comparison tests. The data of the electrophysiological recordings were analyzed using t test. The immunoblot data were analyzed using two-way factorial ANOVA using group and IA training as the two factors followed by post hoc Sidak’s multiple comparison tests. All data are reported as mean ± standard error of mean (SEM). All statistical analyses were performed using the GraphPad PRISM version 6.0 (La Jolla, CA, USA), and P < 0.05 was considered statistically significant.
Results
Physiological variables
Throughout desflurane exposure, HR, SaO2, and the temperatures were within the normal range (data not shown) and arterial blood pressures were stable (Fig. 2). The values of pH, PaCO2, PaO2, and HCO3− were 7.38 ± 0.02, 42.0 ± 2.8, 185.0 ± 11.1, and 24.0 ± 0.5 (mean ± SEM), respectively after 2 h of anesthesia.

Systolic and diastolic arterial blood pressure during desflurane anesthesia. Arterial blood pressures were stable throughout anesthesia. Data are presented as mean ± SEM (n = 5)
IA latencies on day 1 were significantly declined with 1.2 MAC desflurane, but not with 0.7 MAC
The latency to enter the dark box on day 1 after desflurane exposure significantly declined in the 1.2 MAC desflurane group (n = 17, 105 ± 34 s), but not in the 0.7 MAC desflurane group (n = 17, 206 ± 39 s), compared with the control (n = 17, 273 ± 31 s; one-way ANOVA, F [2,48] = 5.974, P = 0.005, post hoc Tukey’s multiple comparison test, mean difference [95% confidence interval], control vs. 1.2 MAC desflurane, 168 [49.9 to 287], P = 0.004; 1.2 MAC desflurane vs. 0.7 MAC desflurane, − 101 [− 220 to 17.7], P = 0.110; control vs. 0.7 MAC desflurane, 67.5 [− 51.2 to 186], P = 0.362; Fig. 3a).

IA latencies. (a) IA latencies on day 1 were significantly declined in the 1.2 MAC desflurane (n = 17, 104 ± 34 s) (post hoc Tukey’s, *P = 0.004), but not in the 0.7 MAC desflurane group (n = 17, 206 ± 39 s) (post hoc Tukey’s, P = 0.362) compared with the control (n = 17, 273 ± 31 s). (b) IA latencies were not declined on day 3 (control [n = 16], 264 ± 38 s; desflurane [n = 16], 229 ± 32 s) and day 7 (control [n = 15], 159 ± 35 s; desflurane [n = 15], 160 ± 44 s) (two-way ANOVA, group, F [1,90] = 5.426, P = 0.022; day, F [2,90] = 0.052, P = 0.417; group x day, F [1,90] = 3.263, P = 0.043, post hoc Sidak’s multiple comparison test, day 3, control vs. desflurane, P = 0.861; day 7, control vs. desflurane, P > 0.999). Each rat was tested only once. The same data are used for day 1 in (a) and (b). Data are presented as mean ± SEM
IA latencies were not declined on days 3 or 7 with 1.2 MAC desflurane
To investigate whether the impairment of learning after exposure to 1.2 MAC desflurane is a long lasting change or a temporary reversible change, we performed IA test on days 3 and 7 after exposure to 1.2 MAC desflurane. The latencies of the desflurane group were not significantly different from those of the controls on days 3 and 7 (day 3, control [n = 16], 264 ± 38 s; desflurane [n = 16], 229 ± 32 s; day 7, control [n = 15], 159 ± 35 s; desflurane [n = 15], 160 ± 44 s, two-way ANOVA, group, F [1,90] = 5.426, P = 0.022; day, F [2,90] = 0.052, P = 0.417; group x day, F [1,90] = 3.263, P = 0.043, post hoc Sidak’s multiple comparison test, day 3, control vs. desflurane, P = 0.861; day 7, control vs. desflurane, P > 0.999) (Fig. 3b).
Collectively, these results showed that exposure to 1.2 MAC concentration of desflurane caused short-term reversible impairments in contextual learning.
Hippocampal LTP was significantly suppressed on day 1 of desflurane exposure
Because contextual learning was impaired on day 1 of desflurane exposure, we assessed the LTP of hippocampal CA1 to confirm the changes in synaptic plasticity. Hippocampal LTP on day 1 was significantly suppressed in the desflurane group (n = 7, 138 ± 4%) compared with the control group (n = 5, 165 ± 8%; P = 0.006; Fig. 4).

Hippocampal LTP was significantly suppressed on day 1 of desflurane exposure. (a) Representative fEPSP traces following the induction of stable LTP. (B) Summary of the fEPSP slopes after applying high-frequency stimulation (100 Hz, 1 s). LTP, induced by the stimulation was significantly suppressed after desflurane exposure (control [n = 5], 165 ± 8%; desflurane [n = 7], 138 ± 4%; *P = 0.006)
Synaptic GluR1 did not increase by IA-training on day 1 of desflurane exposure
IA training is known to induce LTP in the hippocampus by delivering AMPAR to synapse [22, 23]. Because LTP was suppressed with desflurane exposure, we investigated the expression levels of the AMPAR GluR1 subunit on day 1 in both IA-trained and untrained groups. One rat in the IA-trained desflurane group stayed in the light box until the cutoff time of 360 s during the training phase and was not shocked. Thus, this animal was excluded.
There was no difference between the untrained control and untrained desflurane group (125.9 ± 9.2% of untrained control) (group x IA, F [1, 28] = 4.490, P = 0.041; group, F [1, 28] = 0.305, P = 0.585; IA, F [1, 28] = 6.851, P = 0.013; post hoc Sidak’s multiple comparison test, untrained control vs. IA-trained control, P = 0.010; untrained desflurane vs. IA-trained desflurane, P = 1.000; untrained control vs. untrained desflurane, P = 0.326; Fig. 5). After the IA training, the GluR1 protein level was significantly elevated in the control group ([n = 10], 145.9 ± 11.6% of untrained control group [n = 10]). In contrast, the GluR1 protein level was not elevated with IA training in the desflurane group ([n = 9], 103.8 ± 8.9% of untrained desflurane group [n = 10]). These results indicate that desflurane does not alter baseline synaptic GluR1 expression, but suppresses the delivery of GluR1 to hippocampal synaptoneurosomes caused by IA training.

GluR1 was not elevated with IA-training on day 1 of desflurane exposure. (a) Representative immunoblot of GluR1 in synaptoneurosomes. The first three bands were used to create calibration curve. Lanes 1, 5, 9: IA-trained control; lanes 2, 6, 10: IA-trained desflurane; lanes 3, 7, 11: untrained control; lanes 4, 8, 12: untrained desflurane. (b) The amount of GluR1 in synaptoneurosomes was significantly elevated with IA-training in the control (145.9 ± 11.6% of untrained control; post hoc Sidak’s test, *P = 0.003), but not in the desflurane (103.8 ± 8.9% of untrained desflurane; post hoc Sidak’s test, P = 0.927), group
Discussion
The present study has showed that exposure to 1.2 MAC desflurane for 2 h caused impairment of contextual learning on day 1, but not on day 3 or 7. The concentration of desflurane appears important because 0.7 MAC desflurane has no effects on day 1. The impairment in learning on day 1 is accompanied by suppression of hippocampal LTP. Moreover, desflurane inhibits the delivery of AMPAR to synapses produced by IA training without altering baseline synaptic GluR1 expression.
These results are similar to those of isoflurane we demonstrated in our previous study [15]. However the distinct difference is that the learning impairment by desflurane occured and dissipated much earlier than that of isoflurane (day 1 vs. day 7 after exposure). The mechanisms underlying this difference remain unclear, but are unlikely to be explained by the differences in the blood/gas partition coefficients and the resulting differences in the pharmacokinetics of the two anesthetics, because the observed learning impairments occurred long after the anesthetics disappeared from the central nervous system. Several studies have reported that in contrast to isoflurane [20, 21], desflurane does not induce mitochondrial damage [16], apoptosis [17, 18], caspase activation, or amyloid β generation [19] in cells, neurons, or tissues, which suggests that desflurane is less likely than isoflurane to have lasting effects on the central nervous system.
We believe that our finding of desflurane causing learning deficits on day 1 in young adults is relevant, because quick and complete recovery and earlier discharge are required in the ambulatory surgery setting, presumably more so in the working age patients than in older patients. Nearly half of ambulatory surgery patients are reported to resume normal activities on postoperative day 1 [28, 29]. Our results imply that patients may have the risk of cognitive decline on the hours following surgery.
Several clinical studies have compared desflurane with sevoflurane [6, 7, 9], isoflurane [4], or propofol [5, 8] on cognitive function after general anesthesia. Some studies have reported the superiority of desflurane, while other studies have reported no differences, regardless of the time when the neurocognitive tests were performed, as the postoperative tests were performed within 24 h in some studies [6,7,8,9], and after 24 h in others [4,5,6, 8]. Accordingly, the effects of desflurane on cognitive dysfunction in clinical settings remain unclear. The concentration of desflurane may have meaning, as learning was impaired dose-dependently, which is consistent with the previous animal studies [14, 15, 29, 30], and deep anesthesia has been shown to increase postoperative delirium [30, 31], POCD [31, 32], and 1-year mortality [32, 33] clinically.
Our immunoblot analysis has showed that desflurane prevented the increase in synaptoneurosomal GluR1 induced by IA training. This suggests that prior exposure to desflurane interferes with AMPAR trafficking [22], which is known to be essential for the establishment of hippocampal learning. We previously reported that exposure to isoflurane increased GluR1 significantly in the untrained group by inhibiting ubiquitination, a main degradation pathway of GluR1 [15]. By contrast, exposure to desflurane showed no significant differences in GluR1 between the untrained control and desflurane groups. The mechanisms by which desflurane prevents the delivery of AMPAR to synapses are unclear, and as the molecular processes involved in AMPAR trafficking are exceedingly complex [33,34,35], further studies are needed to elucidate the underlying mechanisms by which desflurane prevents the delivery of AMPAR to synapses.
There are limitations to our study. First, we employed only one behavioral test of contextual learning (the IA test), and therefore, did not evaluate other aspects of cognitive function. Second, no group of rats underwent surgery, because we wished to examine the effects of desflurane alone. As inflammation due to surgery is known to cause learning and memory impairment [28, 35], learning deficits caused by desflurane may have been exacerbated and/or prolonged if surgery was superimposed. Third, we examined the AMPAR in the current study because it is known to be critical in learning and memory. However, our results do not preclude the possible role of other receptors such as γ-aminobutyric acid [36] and N-methyl-D-aspartic acid receptors [37], which are known to be the important targets of inhalational anesthetics.
Conclusions
Exposure to 1.2 MAC desflurane for 2 h caused short-term reversible impairment of learning and memory in young adult rats accompanied by suppression of LTP and the delivery of GluR1 to hippocampal synaptoneurosomes. To fully evaluate the short-term effects of inhalational anesthetics on learning and memory, further studies are required in other inhalational anesthetics (e.g. sevoflurane or xenon).
Availability of data and materials
The data of the current study are available from the corresponding author on reasonable request.
Abbreviations
- ACSF:
-
artificial cerebrospinal fluid
- AMPAR:
-
α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor
- ANOVA:
-
analysis of variance
- fEPSPs:
-
extracellular field excitatory postsynaptic potentials
- HR:
-
heart rate
- IA:
-
inhibitory avoidance
- LTP:
-
long-term potentiation
- MAC:
-
minimum alveolar concentration
- POCD:
-
postoperative cognitive dysfunction
- POD:
-
postoperative delirium
- SaO2 :
-
oxygen saturation
- SEM:
-
standard error of mean
References
Monk TG, Weldon BC, Garvan CW, Dede DE, Van Der Aa MT, Heilman KM, et al. Predictors of cognitive dysfunction after major noncardiac surgery. Anesthesiology. 2008;108(1):18–30.
Johnson T, Monk T, Rasmussen LS, Abildstrom H, Houx P, Korttila K, et al. Postoperative cognitive dysfunction in middle-aged patients. Anesthesiology. 2002;96(6):1351–7.
Steinmetz J, Christensen KB, Lund T, Lohse N, Rasmussen LS. Long-term consequences of postoperative cognitive dysfunction. Anesthesiology. 2009;110(3):548–55.
Zhang B, Tian M, Zhen Y, Yue Y, Sherman J, Zheng H, et al. The effects of isoflurane and desflurane on cognitive function in humans. Anesth Analg. 2012;114(2):410–5.
Royse CF, Andrews DT, Newman SN, Stygall J, Williams Z, Pang J, et al. The influence of propofol or desflurane on postoperative cognitive dysfunction in patients undergoing coronary artery bypass surgery. Anaesthesia. 2011;66(6):455–64.
Rörtgen D, Kloos J, Fries M, Grottke O, Rex S, Rossaint R, et al. Comparison of early cognitive function and recovery after desflurane or sevoflurane anaesthesia in the elderly: a double-blinded randomized controlled trial. Br J Anaesth. 2010;104(2):167–74.
Chen X, Zhao M, White PF, Li S, Tang J, Wender RH, et al. The recovery of cognitive function after general anesthesia in elderly patients: a comparison of desflurane and sevoflurane. Anesth Analg. 2001;93(6):1489–94.
Lindqvist M, Schening A, Granstrom A, Bjorne H, Jakobsson JG. Cognitive recovery after ambulatory anaesthesia based on desflurane or propofol: a prospective randomised study. Acta Anaesthesiol Scand. 2014;58(9):1111–20.
Tachibana S, Hayase T, Osuda M, Kazuma S, Yamakage M. Recovery of postoperative cognitive function in elderly patients after a long duration of desflurane anesthesia: a pilot study. J Anesth. 2015;29(4):627–30.
Culley DJ, Baxter M, Yukhananov R, Crosby G. The memory effects of general anesthesia persist for weeks in young and aged rats. Anesth Analg. 2003;96(4):1004–9.
Culley DJ, Baxter MG, Yukhananov R, Crosby G. Long-term impairment of acquisition of a spatial memory task following isoflurane-nitrous oxide anesthesia in rats. Anesthesiology. 2004;100(2):309–14.
Culley DJ, Baxter MG, Crosby CA, Yukhananov R, Crosby G. Impaired acquisition of spatial memory 2 weeks after isoflurane and isoflurane-nitrous oxide anesthesia in aged rats. Anesth Analg. 2004;99(5):1393–7.
Callaway JK, Jones NC, Royse CF. Isoflurane induces cognitive deficits in the Morris water maze task in rats. Eur J Anaesthesiol. 2012;29(5):239–45.
Callaway JK, Jones NC, Royse AG, Royse CF. Memory impairment in rats after desflurane anesthesia is age and dose dependent. J Alzheimers Dis. 2015;44(3):995–1005.
Uchimoto K, Miyazaki T, Kamiya Y, Mihara T, Koyama Y, Taguri M, et al. Isoflurane impairs learning and hippocampal long-term potentiation via the saturation of synaptic plasticity. Anesthesiology. 2014;121(2):302–10.
Zhang Y, Xu Z, Wang H, Dong Y, Shi HN, Culley DJ, et al. Anesthetics isoflurane and desflurane differently affect mitochondrial function, learning, and memory. Ann Neurol. 2012;71(5):687–98.
Loop T, Dovi-Akue D, Frick M, Roesslein M, Egger L, Humar M, et al. Volatile anesthetics induce caspase-dependent, mitochondria-mediated apoptosis in human T lymphocytes in vitro. Anesthesiology. 2005;102(6):1147–57.
Zhang Y, Dong Y, Wu X, Lu Y, Xu Z, Knapp A, et al. The mitochondrial pathway of anesthetic isoflurane-induced apoptosis. J Biol Chem. 2010;285(6):4025–37.
Zhang B, Dong Y, Zhang G, Moir RD, Xia W, Yue Y, et al. The inhalation anesthetic Desflurane induces caspase activation and increases amyloid β-protein levels under hypoxic conditions. J Biol Chem. 2008;283(18):11866–75.
Xie Z, Dong Y, Maeda U, Moir RD, Xia W, Culley DJ, et al. The inhalation anesthetic isoflurane induces a vicious cycle of apoptosis and amyloid β-protein accumulation. J Neurosci Society for Neuroscience. 2007;27(6):1247–54.
Xie Z, Culley DJ, Dong Y, Zhang G, Zhang B, Moir RD, et al. The common inhalation anesthetic isoflurane induces caspase activation and increases amyloid β-protein level in vivo. Ann Neurol. 2008;64(6):618–27.
Whitlock JR, Heynen AJ, Shuler MG, Bear MF. Learning induces long-term potentiation in the hippocampus. Science. 2006;313(5790):1093–7.
Heynen AJ, Quinlan EM, Bae DC, Bear MF. Bidirectional, activity-dependent regulation of glutamate receptors in the adult Hippocampus in vivo. Neuron. 2000;28(2):527–36.
Gong D, Fang Z, Ionescu P, Laster MJ, Terrell RC, Eger EI. Rat strain minimally influences anesthetic and convulsant requirements of inhaled compounds in rats. Anesth Analg. 1998;87(4):963–6.
Staib-Lasarzik I, Kriege O, Timaru-Kast R, Pieter D, Werner C, Engelhard K, et al. Anesthesia for euthanasia influences mRNA expression in healthy mice and after traumatic brain injury. J Neurotrauma. 2014;31(19):1664–71.
Luh C, Gierth K, Timaru-Kast R, Engelhard K, Werner C, Thal SC. Influence of a brief episode of anesthesia during the induction of experimental brain trauma on secondary brain damage and inflammation. PLoS One. 2011;6(5):e19948.
Hollingsworth EB, McNeal ET, Burton JL, Williams RJ, Daly JW, Creveling CR. Biochemical characterization of a filtered synaptoneurosome preparation from Guinea pig cerebral cortex: cyclic adenosine 3′:5′-monophosphate-generating systems, receptors, and enzymes. J Neurosci. 1985;5(8):2240–53.
White PF, Tang J, Wender RH, Yumul R, Stokes OJ, Sloninsky A, et al. Desflurane versus sevoflurane for maintenance of outpatient anesthesia: the effect on early versus late recovery and perioperative coughing. Anesth Analg. 2009;109(2):387–93.
Liu XS, sheng XQ, Zeng QW, Li Q, Liu J, Feng XM, et al. Sevoflurane impairs memory consolidation in rats, possibly through inhibiting phosphorylation of glycogen synthase kinase-3β in the hippocampus. Neurobiol Learn Mem. 2010;94(4):461–7.
Radtke FM, Franck M, Lendner J, Krüger S, Wernecke KD, Spies CD. Monitoring depth of anaesthesia in a randomized trial decreases the rate of postoperative delirium but not postoperative cognitive dysfunction. Br J Anaesth. 2013;110(suppl 1):i98–105.
Chan MTV, Cheng BCP, Lee TMC, Gin T. BIS-guided anesthesia decreases postoperative delirium and cognitive decline. J Neurosurg Anesthesiol. 2013;25(1):33–42.
Monk TG, Saini V, Weldon BC, Sigl JC. Anesthetic management and one-year mortality after noncardiac surgery. Anesth Analg. 2005;100(1):4–10.
Shepherd JD, Huganir RL. The cell biology of synaptic plasticity: AMPA receptor trafficking. Annu Rev Cell Dev Biol. 2007;23(1):613–43.
Henley JM, Barker EA, Glebov OO. Routes, destinations and delays: recent advances in AMPA receptor trafficking. Trends in Neurosci. 2011;34(5):258–68.
Riedel B, Browne K, Silbert B. Cerebral protection: inflammation, endothelial dysfunction, and postoperative cognitive dysfunction. Current Opin Anaesthesiol. 2014;27(1):89–97.
Garcia P, Kolesky S, Jenkins A. General anesthetic actions on GABAA receptors. Curr Neuropharmacol. 2010;8(1):2–9.
Petrenko AB, Yamakura T, Sakimura K, Baba H. Defining the role of NMDA receptors in anesthesia: are we there yet? Vol. 723, Eur J Pharmacol. 2014;723(1):29–37.
Acknowledgements
The authors express their gratitude to Prof. Kenkichi Takase for the valuable comments, Dr. Takahiro Mihara and Dr. Kentaro Tojo for their assistance in editing the manuscript, and Akiko Adachi for technical assistance.
Funding
This work was supported by Grants-in-Aid for Scientific Research (KAKENHI) from the Japan Society for the Promotion of Science (JSPS), Tokyo, Japan.
Grants Number 15 K10517 to Gaku Inagawa, who contributed to the conception of the work, revising the manuscript, and approving the final manuscript.
The funding body played no roles in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study design, A.T. and K.U. collected the data, A.T. analyzed the data and wrote the manuscript, G.I. and T.G. revised the manuscript, and all authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval for this study (approval number: F-A-14-038, F-A-17-079) was provided by the Institutional Animal Care and Use Committee of the Animal Research Center at the Yokohama City University Graduate School of Medicine, Yokohama, Japan.
Consent for publication
Not applicable.
Competing interests
The authors have no competing interests to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Tojo, A., Uchimoto, K., Inagawa, G. et al. Desflurane impairs hippocampal learning on day 1 of exposure: a prospective laboratory study in rats. BMC Anesthesiol 19, 119 (2019). https://doi.org/10.1186/s12871-019-0793-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12871-019-0793-8