
Abstract
Background
Systemic lupus erythematosus (SLE) is a remarkably heterogeneous autoimmune disease. Despite tremendous efforts, our knowledge of serum protein patterns in severe SLE phenotypes is still limited. We investigated the serum protein pattern of SLE, with special emphasis on irreversible organ damage and active lupus nephritis (LN) as assessed by renal Systemic Lupus Erythematosus Disease Activity Index.
Methods
We used proximity extension immunoassay (PEA, Proseek Multiplex, Olink) to assess the serum levels of ninety-two inflammation-related proteins in Czech patients with SLE (n = 75) and age-matched healthy control subjects (n = 23). Subgroup analysis was carried out on the basis of organ damage (with/without, 42/33) and biopsy-proven LN (with/without, 27/48; active LN, n = 13; inactive LN, n = 14).
Results
Of thirty deregulated proteins between SLE and the healthy controls (P corr < 0.05), the top upregulated proteins in SLE were sirtuin 2, interleukin 18 (IL18), and caspase 8 (P corr < 0.0006). Of these, sirtuin 2 and caspase 8 had not yet been reported with SLE. Elevated levels of IL8, CCL2/MCP1, CCL11, and MMP10 (P corr < 0.05) were detected in patients with organ damage for which the serum levels of CCL11 and MMP10 were particularly informative in organ damage prediction. Comparing patients based on LN, elevated levels of CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8, sIL18R1, bNGF, and GDNF (P corr < 0.05) were detected in active LN. Except GDNF, all LN-associated markers showed usefulness in prediction of active renal disease.
Conclusions
This highly sensitive PEA analysis identified the serum pattern of SLE, organ damage, and active LN, with many novel candidate proteins detected. Their exact role and suitability as biomarkers in SLE deserve further investigation.
Similar content being viewed by others


Background
Systemic lupus erythematosus (SLE) is a serious, complex, multi-system autoimmune rheumatic disease with significant variability in the phenotypes and severity of the disease. The greatest challenges continue to be the prevention and management of irreversible organ damage and active lupus nephritis (LN), one of the most feared phenotypes in SLE.
Organ damage is a primary outcome in SLE, which is accrued not only during the disease course, but also by therapy itself [1]. Early damage is more likely to be linked to active inflammation, while late irreversible damage is often attributable to the side effects of drugs and especially to chronic and cumulative corticosteroid exposure [2]. The Systemic Lupus International Collaborating Clinics/American College of Rheumatology SLICC/ACR Damage Index (SDI), divided into 38 items grouped in 12 organ systems, is a valid measure of irreversible organ damage in SLE [1]. Despite improvement in the survival of SLE patients in recent decades, significantly higher morbidity and mortality are reported in patients developing irreversible organ damage [1]. The patterns of organ damage vary among populations [3,4,5], but the musculoskeletal, cardiovascular, and renal systems are those most frequently affected [6]. Nowadays, prevention of irreversible damage is a major goal in the management of SLE patients and identification of the key molecules involved in the pathogenesis of organ damage is needed.
Lupus nephritis is a major manifestation associated with higher morbidity and mortality of SLE patients [7]. It has a considerable influence on treatment decisions, as well as long-term outcomes. The effective treatment of LN requires a correct diagnosis, timely intervention, and early treatment of any disease relapse. Renal biopsy is still the gold standard for diagnosis and deciding on therapy in LN but its invasive nature prevents it from being used repetitively in many cases [8]. Traditional clinical parameters such as proteinuria, glomerular filtration rate, urine sediments, anti-dsDNA antibodies, and complement levels are not sensitive or specific enough to detect activity and early relapse of LN [9, 10]. Novel serum and urinary biomarkers such as cytokines and chemokines CCL2 [11], CCL3, CCL5 [12], IL17 [11], BLyS, APRIL [13], growth factor TGFβ [11] and others (TWEAK [14], IGFBP2 [15], OPG [16]) have recently been nominated for diagnosis and monitoring of LN. Although intensively investigated [17, 18], only a few biomarkers have been assessed for prediction of renal activity or prognosis. Identification of novel and reliable biomarkers or their combinations for LN reflecting also disease activity is, therefore, highly desirable.
In this study we aimed to assess the serum protein pattern of SLE using a highly sensitive multiplex proximity extension immunoassay (PEA) on 92 inflammation-related proteins. Special emphasis was given to serum patterns associated with irreversible organ damage and LN reflecting the renal disease activity and their usefulness in the prediction of these severe phenotypes.
Methods
Study population and materials
Serum samples were obtained from 75 Czech SLE patients; all enrolled patients fulfilled the ACR classification criteria [19]. The samples were aliquoted and stored at − 80 °C until further use. Organ damage was assessed by means of the SDI damage index (Systemic Lupus International Collaborating Clinics/American College of Rheumatology Damage Index) [1] and disease activity was evaluated by means of SLEDAI (Systemic Lupus Erythematosus Disease Activity Index) [20]. Subgroups were formed on the basis of (1) the SDI (SDI = 0, n = 33; SDI = 1, n = 17; SDI ≥ 2, n = 25), (2) the biopsy-proven presence of LN (no LN, n = 48; LN, n = 27), and (3) the renal SLEDAI within LN subgroup, where renal SLEDAI score of ≥ 4 was taken as an indicator of active LN (inactive LN, n = 14; active LN, n = 13). The renal SLEDAI consists of the four renal parameters: hematuria, pyuria, proteinuria, and urinary casts [20]. The mean of LN duration in active LN patients was 7 years (range 0–19 years) and in inactive LN patients 8 years (range 1–18 years). The demographic and clinical features are described in Table 1. The age-matched control group of healthy subjects comprised 23 medical staff members (mean age 40, range 26–73, female/male 15/8), who gave statements about their health status and excluded any medication used for SLE treatment (corticosteroids, antimalarials, immunosuppressant drugs). The patients and control subjects provided written informed consent about the usage of peripheral blood for the purpose of this study, which was approved by the ethics committee of the University Hospital and Palacky University Olomouc.
Proximity extension immunoassay (PEA)
The serum levels of 92 inflammation-related proteins were simultaneously measured by a PEA using the Proseek Multiplex Inflammation kit I (Olink Bioscience, Sweden) according to the manufacturer’s recommendation. Briefly, each analyte is recognized by a pair of oligonucleotide-labelled antibodies and when binding to their correct targets, they give rise to reporter amplicons which are amplified and quantified by microfluidic-based real-time PCR (BioMark™ HD System, Fluidigm Corporation). The data obtained is normalized and used for the relative quantification of the concentration of each analyte [26, 27]. The PEA kits offer the same level of performance as ELISA and comparable sensitivity to standard ELISA kits with much less sample and a higher dynamic range. For a panel description see Additional file 1: Table S1; for the sensitivity and specificity parameters of the PEA analysis see [26, 27].
Statistics
All statistical analyses were performed on linearized data (linear ddCq) for each analyte. Statistical tests (Mann–Whitney–Wilcoxon test, Benjamini–Hochberg correction, Spearman correlations, Receiver Operating Characteristic (ROC) curve analysis, and Bayesian probability model) were performed using the R statistical software with the Caret package (http://www.r-project.org/; http://topepo.github.io/caret/index.html). The P value for each protein was adjusted for multiple comparisons using the False Discovery Rate by the Benjamini–Hochberg procedure. P corr value < 0.05 was considered significant.
Results
Protein pattern of SLE
In order to assess the serum protein fingerprint associated with SLE, we compared the serum protein levels obtained by PEA immunoassay in the SLE patients and healthy controls. Of 92 biomarkers that were analyzed, the levels of 14 analytes (IL1A, IL2, sIL2RB, IL4, IL5, IL13, IL20, sIL20RA, IL33, TSLP, ARTN, TNF, LIF, NRTN) were below the limit of detection in our sample set and therefore they were excluded from further analysis. Comparing SLE and the controls, 29 proteins were upregulated and sDNER downregulated in SLE (P corr < 0.05; Table 2a, Additional file 1: Table S2). The distribution of the serum levels of top-upregulated proteins (sirtuin 2, IL18, caspase 8, sCD40/sTNFRSF5, sSLAMF1, sTNFRSF9, axin 1, sulfotransferase 1A1, STAMBP, CCL19/MIP-3ß, IL10, and CCL4/MIP-1β; P corr < 0.003) is shown in Fig. 1. For the serum protein pattern associated with SLE and the changes in protein levels between SLE and the controls for top-deregulated analytes see Figs. 2a and 3a.

Distribution of serum levels for top-deregulated proteins between healthy controls and SLE. Group means are indicated by horizontal bars, error bars indicate 95% CI; P corr values after multiple corrections are stated

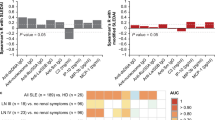
Protein serum fingerprints associated with a SLE, b organ damage, and c active lupus nephritis (LN). Fingerprints are presented as FC (foldchange of group medians) of serum levels of all deregulated serum proteins between particular groups (P corr < 0.05)

Changes in protein levels for top-deregulated analytes between a SLE and controls, b patients with/without organ damage, and c patients with active lupus nephritis and without lupus nephritis (no LN). Changes are presented as percentage of changes between group medians of particular groups
Because of the suggested central role of the IFN pathway in SLE pathogenesis by promoting feedback loops progressively disrupting peripheral immune tolerance and driving disease activity [28, 29], we investigated the IFN protein “signature” of nine IFN-regulated cytokines. Because of the reported association of an increased IFN gene expression “signature” with disease activity [28, 29], we performed correlation analysis among the protein levels of IFN-regulated chemokines and disease activity as assessed by SLEDAI. The analysis revealed elevation of six IFN-regulated cytokines (IL6, CCL2/MCP1, CCL3/MIP-1α, sCD40, CXCL11, and CCL19; P corr ≤ 0.01) in SLE and three (CCL8/MCP2, CXCL9, and CXCL10) did not reach significance (P corr > 0.05). Interestingly, only a mild positive correlation (r = 0.25, P = 0.03; Additional file 1: Table S3) was observed between the levels of IFN-regulated chemokines and disease activity as assessed by SLEDAI. Disease activity assessed by SLEDAI correlated better with the following analytes: IL8, GDNF, CX3CL1/fractalkine (r ≥ 0.403, P ≤ 0.0003), and CCL7/MCP3, IL15RA, VEGFA, and MMP10 (r ≥ 0.355, P ≤ 0.002; Additional file 1: Table S3).
Protein pattern of organ damage
To obtain the protein pattern associated with organ damage, we compared the serum patterns from SLE patients with/without organ damage and subgroups according to the SDI (SDI ≥ 2/SDI = 1/SDI = 0).
The distribution of damaged organs in our patient group and reported cohorts is shown in Additional file 1: Figure S1, Table S4. In the patients with organ damage (SDI ≥ 1), elevated serum levels of IL8, CCL2, IL6, CCL11/eotaxin, FGF21, MMP10, IL18, CCL3, FGF5, and FGF23 (P corr < 0.05) were detected (Table 2b, Fig. 4). The serum protein pattern associated with organ damage and the changes in protein levels between SLE patients with/without organ damage are shown in Figs. 2b and 3b. Although the serum level of CCL11 did not differ between the controls and SLE patients as a whole, the patients with organ damage had higher levels of CCL11 in comparison to those with no organ damage, as well as to the control group (Additional file 1: Figure S2a). We did not observe differences in serum protein pattern between patients with SDI = 1 and SDI ≥ 2 (data not shown).

Distribution of serum levels of proteins distinguishing SLE patients with/without organ damage. Group means are indicated by horizontal bars, error bars indicate 95% CI; P corr values for differences after multiple corrections are stated
Among organ damage associated analytes, the cumulative dose of glucocorticoids correlated positively with levels of IL8, CCL11 (r ≥ 0.326, P ≤ 0.004), CCL2 and MMP10 (r ≥ 0.249, P < 0.05; Additional file 1: Table S3). Additionaly, cumulative dose of glucocorticoids correlated with BDNF, CCL25, CXCL1, GDNF, IL17C, sADA, sCDCP1, sIL18R1, sSCF, and sTGFA (P < 0.05; Additional file 1: Table S3). Moreover, IL8 (r = 0.416, P = 0.0002), MMP10 (r = 0.355, P = 0.002), CCL2, and CCL11 (r ≥ 0.261, P ≤ 0.02; Additional file 1: Table S3) correlated positively with disease activity. In line with other reports, a higher cumulative dosage of glucocorticoids was registered in the patients with SDI ≥ 1 (mean of 30.6 g, min–max 2.6–79.2 g) compared with those without damage (12.8, 0–54.0). Regarding association of disease duration and serum levels of studied proteins, we observed only mild association for CCL11 (r = 0.230, P = 0.047). The disease duration in SLE patients correlated with SDI (r = 0.298, P = 0.009).
Protein pattern of active lupus nephritis and other clinical subsets of SLE
To investigate the serum patterns associated with active LN, we compared subgroups of SLE patients with/without biopsy-proven LN and subgroups of patients with LN classified by the renal SLEDAI as active (renal SLEDAI ≥ 4) or inactive renal disease at the day of sampling. Moreover, we assessed serum patterns associated with other clinical subsets of SLE as neurological, hematological, cardiovascular, skin and musculoskeletal involvements, antiphospholipid syndrome, and renal disorder.
The analysis in biopsy-proven LN patients with active renal disease revealed elevated protein levels of CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8, sIL18R1, bNGF, and GDNF compared to those without LN (Table 2c, Fig. 5). Although the serum levels of GDNF did not differ between the control group and SLE as a whole, its level was enhanced in the patients with LN in comparison to those without LN and the control group (Additional file 1: Figure S2b). The serum protein pattern associated with active LN and the changes in protein levels between the SLE patients without LN and active LN are shown in Figs. 2c and 3c.

Distribution of serum levels of proteins distinguishing SLE patients without lupus nephritis (LN), with inactive lupus nephritis (inactive LN) and active lupus nephritis (active LN). Group means are indicated by horizontal bars, error bars indicate 95% CI
When LN patients with active renal disease was compared to inactive LN subgroup, elevation of sIL15RA, CSF1, bNGF, sIL18R1, sCD40, sCX3CL1, and caspase 8 (P < 0.05, Table 2d, Fig. 5), but not GDNF, in active LN patients was observed.
In the other studied clinical subsets no differences in the serum pattern were detected. The subanalysis confirmed that no candidate biomarker for SLE, organ damage and/or LN are influenced by the gender (data not shown).
Identification of patients with a high probability of organ damage and active lupus nephritis
To investigate the utility of the serum levels of phenotype-associated proteins for the identification of patients with a high probability of severe phenotypes, we constructed probability plots for phenotype-associated proteins based on a Bayesian statistical approach. Additionally, we constructed ROC curves for the proteins associated with organ damage and active LN.
In organ damage, the best predictive model was observed for the serum levels of CCL11 and MMP10, followed by CCL2, whereas IL6 and IL8 were not informative (Fig. 6). Higher serum levels of CCL11 and MMP10 correspond to a higher probability of organ damage. For the analytes associated with organ damage, the ROC curve analysis showed that the area under the curve (AUC) of IL8, CCL2, IL6, CCL11, FGF21, MMP10, IL18, CCL3, FGF5, and FGF23 was 0.784, 0.738, 0.731, 0.727, 0.723, 0.706, 0.697, 0.691, 0.689, and 0.676, respectively (Additional file 1: Figure S3a; for sensitivity, specificity, and other parameters see Additional file 1: Table S5a).

Probability plots of serum analytes associated with organ damage in SLE patients. The grey curve represents a simulated model based on the individual patient serum levels and the black line represents overall trend calculated by the Bayesian statistical approach. The increasing overall trend the higher probability of organ damage. Higher serum levels of CCL11 and MMP10 correspond to higher probability of organ damage, lower serum levels of these analytes to lower probability of organ damage. IL8 and IL6 serum levels were not informative for organ damage prediction
In active LN, the best predictive value was observed for CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8, and sIL18R1 (Fig. 7). Higher serum levels of all these analytes correspond to a higher probability of the presence of active LN. For the analytes associated with active LN, the ROC curve analysis showed that the AUC of CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8, sIL18R1, bNGF, and GDNF were 0.873, 0.857, 0.854, 0.832, 0.798, 0.783, 0.780, and 0.778, respectively (Additional file 1: Figure S3b, Table S5b). Moreover, we observed great sensitivity and specificity for proteins sIL15RA (AUC: 0.879, sensitivity: 100%, specificity: 64.3%), CSF1 (0.813, 84.6, 78.6), sIL18R1 (0.810, 84.6, 78.6), and bNGF (0.805, 69.2, 100) showing good discrimination between active and inactive renal disease in LN patient subgroup (Additional file 1: Figure S3c, Table S5c). Inactive LN patients do not differ from patients without LN, except for GDNF (Fig. 5), suggesting that serum GDNF level remains elevated even when LN is inactive.

Probability plots of serum analytes associated with active lupus nephritis (LN) in SLE patients. The grey curve represents a simulated model based on the individual patient serum levels and the black line represents overall trend calculated by the Bayesian statistical approach. The increasing overall trend the higher probability of active LN. Higher serum levels correspond to higher probability of active LN, lower serum levels of these analytes to lower probability of active LN. The best predictive value was observed for CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8 (CASP8), and sIL18R1
All nominated biomarkers associated with organ damage and active LN showed better discrimination ability in our cohort than the classical markers (Additional file 1: Table S6). The only exception was proteinuria (AUC 0.869), one of the criteria for renal SLEDAI classification.
Discussion
Using innovative highly sensitive multiplex PEA analysis on 92 inflammation-related proteins, we identified the serum protein pattern associated with SLE, with many proteins not yet reported in this disease. Moreover, we identified the serum patterns associated with irreversible organ damage and active LN and identified proteins showing utility for the identification of patients at risk of these severe disease manifestations.
This serum protein study in SLE patients revealed the deregulation of 30 proteins in SLE. The majority of the upregulated proteins were known inflammatory mediators: IL6, IL10 [30], IL18 [31], CX3CL1 [32], CCL2 [33], CCL3, CCL7, CCL19 [34], and FGF23 [35] already reported in SLE previously. Interestingly, the most upregulated proteins—sirtuin 2 and caspase 8—were not associated with SLE or even with any autoimmune disease. However, recent reports in animal models and cell lines support their involvement in inflammation and autoimmunity. Regarding sirtuin 2, macrophages expressing this protein produced more iNOS/NO upon LPS stimulation than those with depleted sirtuin 2 [36]. This result was also confirmed in vivo, where WT mice responded to LPS by increased NO levels and a higher amount of M1-macrophages compared to sirtuin 2 KO mice [36]. Elevated sirtuin 2 also contributed to prolonged hypoinflammation in a septic murine model [37]. Regarding caspase 8, a protein widely recognized for its role in apoptosis, recent reports identify this enzyme as a crucial regulator of inflammation through NFκB activation and cleavage of pro-IL1β and/or pro-IL18, similarly to caspase 1 [38, 39]. These observations lead us to suggest that caspase 8 may also promote autoimmunity by stimulating IL17 production by T cells, as shown for caspase 1 [40]. Moreover, the therapeutic potential of caspase 8 is supported by the observation of attenuated retinal ischemic damage resulting from the inhibition of caspase 8, resulting in the blockade of IL1β production [41]. However, there is evidence about the pleiotropic effects of sirtuin 2 and caspase 8, and thus future studies on their role in SLE are needed.
Further highly upregulated proteins, IL18 and sulfotransferase 1A1, were already reported in autoimmunity. An elevated IL18 serum level was reported in SLE [42], especially in LN patients [43, 44]. Regarding sulfotransferase 1A1, higher activity was found in autoimmune thyroid disease glands compared to normal thyroids [45], but no information yet exists in SLE. Interestingly, we did not detect any elevation of the serum level of the previously reported SLE-associated factor TWEAK and IFNγ [46, 47]. Despite the reported association of the IFN gene expression “signature” with disease activity in SLE [28, 29], we did not confirm either elevated levels of the IFN-regulated chemokines CCL8, CXCL9, CXCL10 or strong correlation of the IFN protein “signature” with disease activity at the protein level in the sera of our patients. Our observation is in line with others [28], thus supporting the opinion that cytokine levels in serum are a less sensitive readout for activation of the IFN pathway than the gene expression “signature”.
Despite tremendous efforts, the greatest challenges still remain in the management of SLE patients with severe organ damage and active LN. Thus, there is a need to identify novel biomarkers that will better facilitate the assessment of organ involvement and disease activity. In our study, SLE patients with organ damage had elevated serum levels of IL8, CCL2, IL6, CCL11, FGF21, MMP10, IL18, CCL3, FGF5, and FGF23 compared to those without organ damage. Of these, enhanced levels of CCL11, MMP10, and CCL2 were informative for the identification of patients with organ damage. Importantly, CCL11, MMP10, and CCL2 also correlated with disease activity. The elevation of the chemokine CCL11 was already associated with damage to various organs, as shown in idiopathic retroperitoneal fibrosis [48] and liver cirrhosis patients [49]. Moreover, in murine models of lung fibrosis [50], as well as of eosinophilic myocarditis [51], the blockade of the CCL11-CCR3 pathway prevented organ damage. Similarly, MMP10 was linked to renal damage [52] and tissue destruction in arthritis [53]. Elevation of MMP10 was already reported in SLE patients [54] and in a murine LN model with glomerulonephritis [55]. Another protein associated with organ damage, CCL2, was already reported in kidney damage in lupus murine models [56] and in SLE patients with irreversible renal damage [57]. Although IL6 and IL8, cytokines involved in the pathogenesis of SLE, were also enhanced in our patients with organ damage, our analysis did not support their predictive value for this severe phenotype. The usefulness of CCL11, MMP10, and CCL2 as biomarkers or possible treatment targets needs to be elucidated in future studies.
Lupus nephritis is considered another challenging SLE phenotype from the point of view of its prediction and preemptive diagnostics. Renal biopsy is still the gold standard to assess the renal involvement of SLE and its severity and pathological category [8]. The search for non-invasive biomarkers in serum and urine reflecting the renal disease activity is therefore a major focus of interest. Our serum protein analysis in LN patients with active renal disease revealed upregulated levels of CSF1, sIL15RA, sCD40, sCX3CL1, caspase 8, sIL18R1, bNGF, and GDNF compared to those without LN. All these markers showed excellent discrimination for active LN, significantly better than the classical markers as shown by us and others [9, 10]. Moreover, we observed good discrimination between active and inactive renal disease in LN patient subgroup for all markers, except for GDNF. Apart from caspase 8 and sIL15RA, emerging evidence of the active involvement of these proteins in LN already exists. Regarding CSF1, elevated serum levels in patients with SLE were shown to reflect kidney histopathology and to predict renal disease activity [58]. Moreover, CSF1 deficiency protected against LN in murine models [59]. Enhanced protein and gene expression of IL15RA was detected in leucocytes from SLE patients [60, 61], probably as a results of hydroxymethylation in promoter region of this gene in SLE [61]. There is also evidence about the crucial role of the CD40-CD40L system in the development, progression and outcome of SLE [62]. Enhanced CD40L protein level was detected in sera from SLE patients [62, 63] as well as class III and IV LN and other inflammatory renal diseases [64]. Moreover, CD40 gene silencing reduced the progression of experimental LN [65]. Regarding sCX3CL1, elevated expression was reported in proliferative LN [66] and the administration of a CX3CL1 antagonist to mice delayed the initiation and ameliorated the progression of LN [67]. Also enhanced expression of IL18R1 has already been reported in SLE patients [68] as well as in peripheral plasmacytoid DCs in active LN patients [69]. Similarly, increased levels of NGF, a complex of 3 subunits—aNGF, bNGF, and gNGF, has been reported in the sera of SLE patients [70] and various renal disorders [71]. Regarding GDNF, a high expression of this protein was detected in renal biopsies from patients with proteinuric nephropathy [72] and increased plasma levels of GDNF were reported in patients with chronic renal diseases [73]. This mesangial autocrine growth factor was shown to play a pivotal role in mesangial cell proliferation, which is essential for the progression of various glomerular diseases [74]. Our study did not confirm IL18 as a useful biomarker to assess the activity of renal disease, as reported by others [42, 43]. On the other hand, our results nominated spectrum of novel biomarkers of renal involvement for further confirmation studies.
Althougth relatively high sensitivity and specificity was obtained for each individual marker in our LN and organ damage subgroups, we believe that using rather a panel of multiple biomarkers and/or combination with other clinical and laboratory parameters would be an appropriate approach in the identification of patients with these severe manifestations.
Conclusions
This exploratory study revealed many novel proteins associated with SLE for future immunopathogenesis studies, as well as nominating candidate biomarkers for irreversible organ damage and active lupus nephritis. Future studies on larger cohorts with well-defined phenotypes as well as the longitudinal follow-up during disease development are needed to prove the suitability of these proteins or their combinations as biomarkers for organ damage and lupus nephritis, with special emphasis on disease activity.
Abbreviations
- 4E-BP1:
-
eukaryotic translation initiation factor 4E-binding protein 1
- aNGF:
-
alpha-nerve growth factor
- APRIL:
-
a proliferation-inducing ligand
- ARTN:
-
artemin
- AXIN1:
-
axin 1
- BLyS:
-
B lymphocyte stimulator
- BDNF:
-
brain-derived neurotrophic factor
- bNGF:
-
beta-nerve growth factor
- CASP8:
-
caspase 8
- CCL2:
-
C-C motif chemokine ligand 2, monocyte chemotactic protein 1
- CCL3:
-
C-C motif chemokine ligand 3, macrophage inflammatory protein 1-alpha
- CCL4:
-
C-C motif chemokine ligand 4, macrophage inflammatory protein 1-beta
- CCL7:
-
C-C motif chemokine ligand 7, monocyte chemotactic protein 3
- CCL8:
-
C-C motif chemokine ligand 8, monocyte chemotactic protein 2
- CCL11:
-
C-C motif chemokine ligand 11, eotaxin-1
- CCL13:
-
C-C motif chemokine ligand 13, monocyte chemotactic protein 4
- CCL19:
-
C-C motif chemokine ligand 19, macrophage inflammatory protein 3-beta
- CCL20:
-
C-C motif chemokine ligand 20, macrophage inflammatory protein 3-alpha
- CCL23:
-
C-C motif chemokine ligand 23, macrophage inflammatory protein 3
- CCL25:
-
C-C motif chemokine ligand 25
- CCL28:
-
C-C motif chemokine ligand 28
- CD40L:
-
cluster of differentiation 40 ligand
- CCR3:
-
C-C chemokine receptor type 3
- CSF1:
-
macrophage colony-stimulating factor 1
- CST5:
-
cystatin D
- CXCL1:
-
C-X-C motif chemokine ligand 1
- CXCL5:
-
C-X-C motif chemokine ligand 5
- CXCL6:
-
C-X-C motif chemokine ligand 6
- CXCL9:
-
C-X-C motif chemokine ligand 9
- CXCL10:
-
C-X-C motif chemokine ligand 10
- CXCL11:
-
C-X-C motif chemokine ligand 11
- EN.RAGE:
-
extracellular newly identified receptor for advanced glycation end-products binding protein, protein S100-A12
- FGF5:
-
fibroblast growth factor 5
- FGF19:
-
fibroblast growth factor 19
- FGF21:
-
fibroblast growth factor 21
- FGF23:
-
fibroblast growth factor 23
- gNGF:
-
gamma-nerve growth factor
- GDNF:
-
glial cell line-derived neurotrophic factor
- IFN:
-
interferon
- IFNG:
-
interferon gamma
- IGFBP2:
-
insulin like growth factor binding protein 2
- IL1A:
-
interleukin-1 alpha
- IL1B:
-
interleukin-1 beta
- IL2:
-
interleukin-2
- IL4:
-
interleukin-4
- IL5:
-
interleukin-5
- IL6:
-
interleukin-6
- IL7:
-
interleukin-7
- IL8:
-
interleukin-8
- IL10:
-
interleukin-10
- IL12B:
-
interleukin-12 beta
- IL13:
-
interleukin-13
- IL17:
-
interleukin-17
- IL17A:
-
interleukin-17A
- IL17C:
-
interleukin-17C
- IL18:
-
interleukin-18
- IL20:
-
interleukin-20
- IL24:
-
interleukin-24
- IL33:
-
interleukin-33
- iNOS/NO:
-
inducible nitric oxide synthase/nitrogen oxide
- KO:
-
knock out
- LAP.TGFB1:
-
latency-associated peptide transforming growth factor beta-1
- LIF:
-
leukemia inhibitory factor
- LN:
-
lupus nephritis
- LPS:
-
lipopolysaccharide
- MMP1:
-
matrix metalloproteinase-1
- MMP10:
-
matrix metalloproteinase-10
- NFκB:
-
nuclear factor kappa B
- NGF:
-
nerve growth factor
- NRTN:
-
neurturin
- NT3:
-
neurotrophin-3
- OSM:
-
oncostatin-M
- PCR:
-
polymerase chain reaction
- PEA:
-
proximity extension immunoassay
- ROC:
-
receiver operating characteristic
- sADA:
-
adenosine deaminase, soluble
- sCD244:
-
natural killer cell receptor 2B4, soluble
- sCD40:
-
cluster of differentiation 40, tumor necrosis factor receptor superfamily member 5, soluble
- sCD5:
-
cluster of differentiation 5, soluble
- sCD6:
-
cluster of differentiation 6, soluble
- sCDCP1:
-
CUB domain-containing protein 1, soluble
- sCX3CL1:
-
C-X3-C motif chemokine ligand 1, fractalkine, soluble
- SDI:
-
Systemic Lupus International Collaborating Clinics/American College of Rheumatology Damage Index
- sDNER:
-
delta and notch-like epidermal growth factor-related receptor, soluble
- sFlt3L:
-
Fms-related tyrosine kinase 3 ligand, soluble
- sHGF:
-
hepatocyte growth factor
- sIL10RA:
-
interleukin-10 receptor subunit alpha, soluble
- sIL10RB:
-
interleukin-10 receptor subunit beta, soluble
- sIL15RA:
-
interleukin-15 receptor subunit alpha, soluble
- sIL18R1:
-
interleukin-18 receptor 1, soluble
- sIL20RA:
-
interleukin-20 receptor subunit alpha, soluble
- sIL22RA1:
-
interleukin-22 receptor subunit alpha-1, soluble
- sIL2RB:
-
interleukin-2 receptor subunit beta, soluble
- SIRT2:
-
sirtuin 2
- SLE:
-
systemic lupus erythematosus
- SLEDAI:
-
Systemic Lupus Erythematosus Disease Activity Index
- sLIFR:
-
leukemia inhibitory factor receptor, soluble
- sOPG:
-
osteoprotegerin, soluble
- sPDL1:
-
programmed cell death 1 ligand 1, soluble
- sSCF:
-
stem cell factor, soluble
- sSLAMF1:
-
signaling lymphocytic activation molecule 1, soluble
- ST1A1:
-
sulfotransferase 1A1
- STAMBP:
-
signal transducing adaptor molecule-binding protein
- sTGFA:
-
transforming growth factor alpha, soluble
- sTNFB:
-
tumor necrosis factor-beta, soluble
- sTNFRSF9:
-
tumor necrosis factor receptor superfamily member 9, soluble
- sTNFSF14:
-
tumor necrosis factor ligand superfamily member 14, soluble
- sTRAIL:
-
tumor necrosis factor-related apoptosis-inducing ligand, soluble
- sTRANCE:
-
tumor necrosis factor-related activation-induced cytokine, soluble
- sTWEAK:
-
tumor necrosis factor ligand superfamily member 12, soluble
- TGFβ:
-
transforming growth factor beta
- TNF:
-
tumor necrosis factor
- TSLP:
-
thymic stromal lymphopoietin
- uPA:
-
urokinase-type plasminogen activator
- VEGFA:
-
vascular endothelial growth factor A
- WT:
-
wild type
References
Gladman DD, Goldsmith CH, Urowitz MB, Bacon P, Fortin P, Ginzler E, et al. The Systemic Lupus International Collaborating Clinics/American College of Rheumatology (SLICC/ACR) damage index for systemic lupus erythematosus international comparison. J Rheumatol. 2000;27:373–6.
Nossent J, Cikes N, Kiss E, Marchesoni A, Nassonova V, Mosca M, et al. Current causes of death in systemic lupus erythematosus in Europe, 2000–2004: relation to disease activity and damage accrual. Lupus. 2007;16:309–17.
Chambers SA, Allen E, Rahman A, Isenberg D. Damage and mortality in a group of British patients with systemic lupus erythematosus followed up for over 10 years. Rheumatology (Oxford). 2009;48:673–5.
Mok CC, Ho CT, Wong RW, Lau CS. Damage accrual in southern Chinese patients with systemic lupus erythematosus. J Rheumatol. 2003;30:1513–9.
Danila MI, Pons-Estel GJ, Zhang J, Vilá LM, Reveille JD, Alarcón GS. Renal damage is the most important predictor of mortality within the damage index: data from LUMINA LXIV, a multiethnic US cohort. Rheumatology (Oxford). 2009;48:542–5.
Doria A, Gatto M, Zen M, Iaccarino L, Punzi L. Optimizing outcome in SLE: treating-to-target and definition of treatment goals. Autoimmun Rev. 2014;13:770–7.
Cameron JS. Lupus nephritis. J Am Soc Nephrol. 1999;10:413–24.
Misra R, Gupta R. Biomarkers in lupus nephritis. Int J Rheum Dis. 2015;18:219–32.
Balow JE. Clinical presentation and monitoring of lupus nephritis. Lupus. 2005;14:25–30.
Enghard P, Riemekasten G. Immunology and the diagnosis of lupus nephritis. Lupus. 2009;18:287–90.
Susianti H, Iriane VM, Dharmanata S, Handono K, Widijanti A, Gunawan A, et al. Analysis of urinary TGF-b1, MCP-1, NGAL, and IL-17 as biomarkers for lupus nephritis. Pathophysiology. 2015;22:65–71.
Kulkarni O, Anders HJ. Chemokines in lupus nephritis. Front Biosci. 2008;13:3312–20.
López P, Rodríguez-Carrio J, Caminal-Montero L, Mozo L, Suárez A. A pathogenic IFNα, BLyS and IL-17 axis in Systemic Lupus Erythematosus patients. Sci Rep. 2016;6:20651.
Schwartz N, Rubinstein T, Burkly LC, Collins CE, Blanco I, Su L, et al. Urinary TWEAK as a biomarker of lupus nephritis: a multicenter cohort study. Arthritis Res Ther. 2009;11:R143.
Ding H, Kharboutli M, Saxena R, Wu T. Insulin-like growth factor binding protein-2 as a novel biomarker for disease activity and renal pathology changes in lupus nephritis. Clin Exp Immunol. 2016;184:11–8.
Abdallah E, El-Shishtawy S, Sherif N, Abdelwahab MA. Diagnostic performance of urinary osteoprotegrin as a novel biomarker for early detection of lupus nephritis activity. Life Sci J. 2015;12:75–81.
Li Y, Fang X, Li QZ. Biomarker profiling for lupus nephritis. Genomics Proteomics Bioinform. 2013;11:158–65.
Benjachat T, Tongyoo P, Tantivitayakul P, Somparn P, Hirankarn N, Prom-On S, et al. Biomarkers for refractory Lupus nephritis: a microarray study of kidney tissue. Int J Mol Sci. 2015;16:14276–90.
Hochberg MC. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 1997;40:1725.
Bombardier C, Gladman DD, Urowitz MB, Caron D, Chang CH. Derivation of the SLEDAI. A disease activity index for lupus patients. Arthritis Rheum. 1992;35:630–40.
The American College of Rheumatology nomenclature and case definitions for neuropsychiatric lupus syndromes. Arthritis Rheum. 1999;42:599–608.
Gillian JN, Sontheimer RD. Skin manifestations of SLE. Clin Rheum Dis. 1981;8:207–18.
Miakis S, Lockshin MD, Atsumi T, Branch DW, Brey RL, Cervera R, et al. International consensus statement on an update of the classification criteria for definite antiphospholipid syndrome (APS). J Thromb Haemost. 2006;4:295–306.
Singh S, Wu T, Xie C, Vanarsa K, Han J, Mahajan T, et al. Urine VCAM-1 as a marker of renal pathology activity index in lupus nephritis. Arthritis Res Ther. 2012;14:R164.
Xuejing Z, Jiazhen T, Jun L, Xiangqing X, Shuguang Y, Fuyou L. Urinary TWEAK level as a marker of lupus nephritis activity in 46 cases. J Biomed Biotechnol. 2012;2012:359647.
Assarsson E, Lundberg M, Holmquist G, Björkesten J, Thorsen SB, Ekman D, et al. Homogenous 96-plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS ONE. 2014;9:e95192.
Schneiderova P, Pika T, Gajdos P, Fillerova R, Kromer P, Kudelka M, et al. Serum protein fingerprinting by PEA immunoassay coupled with a pattern-recognition algorithms distinguishes MGUS and multiple myeloma. Oncotarget. 2016 doi:10.18632/oncotarget.11242. [Epub ahead of print].
Baechler EC, Batliwalla FM, Karypis G, Gaffney PM, Ortmann WA, Espe KJ, et al. Interferon-inducible gene expression signature in peripheral blood cells of patients with severe lupus. Proc Natl Acad Sci USA. 2003;100:2610–5.
Feng X, Huang J, Liu Y, Xiao L, Wang D, Hua B, et al. Identification of interferon-inducible genes as diagnostic biomarker for systemic lupus erythematosus. Clin Rheumatol. 2015;34:71–9.
Gröndal G, Gunnarsson I, Rönnelid J, Rogberg S, Klareskog L, Lundberg I. Cytokine production, serum levels and disease activity in systemic lupus erythematosus. Clin Exp Rheumatol. 2000;18:565–70.
Favilli F, Anzilotti C, Martinelli L, Quattroni P, De Martino S, Pratesi F, et al. IL-18 activity in systemic lupus erythematosus. Ann N Y Acad Sci. 2009;1173:301–9.
Yajima N, Kasama T, Isozaki T, Odai T, Matsunawa M, Negishi M, et al. Elevated levels of soluble fractalkine in active systemic lupus erythematosus: potential involvement in neuropsychiatric manifestations. Arthritis Rheum. 2005;52:1670–5.
Hrycek E, Franek A, Błaszczak E, Dworak J, Hrycek A. Serum levels of selected chemokines in systemic lupus erythematosus patients. Rheumatol Int. 2013;33:2423–7.
Bauer JW, Baechler EC, Petri M, Batliwalla FM, Crawford D, Ortmann WA, et al. Elevated serum levels of interferon-regulated chemokines are biomarkers for active human systemic lupus erythematosus. PLoS Med. 2006;3:e491.
Masi L, Cavalli L, Falcini F, Franceschelli F, Leoncini G, Fossi C, et al. P8-measurement of fibroblast growth factor-23 (FGF23) in the serum of patients affected by juvenile systemic lupus erythematosus: a possible marker of kidney damage. Clin Cases Miner Bone Metab. 2010;7:214.
Lee AS, Jung YJ, Kim D, Nguyen-Thanh T, Kang KP, Lee S, et al. SIRT2 ameliorates lipopolysaccharide-induced inflammation in macrophages. Biochem Biophys Res Commun. 2014;450:1363–9.
Wang X, Buechler NL, Martin A, Wells J, Yoza B, McCall CE, et al. Sirtuin-2 regulates sepsis inflammation in ob/ob mice. PLoS ONE. 2016;11:e0160431.
Lemmers B, Salmena L, Bidère N, Su H, Matysiak-Zablocki E, Murakami K, et al. Essential role for caspase-8 in Toll-like receptors and NF-κB signaling. J Biol Chem. 2007;282:7416–23.
Gurung P, Anand PK, Malireddi RK, Vande Walle L, Van Opdenbosch N, Dillon CP, et al. FADD and caspase-8 mediate priming and activation of the canonical and noncanonical Nlrp3 inflammasomes. J Immunol. 2014;192:1835–46.
Lalor SJ, Dungan LS, Sutton CE, Basdeo SA, Fletcher JM, Mills KH. Caspase-1-processed cytokines IL-1beta and IL-18 promote IL-17 production by gammadelta and CD4 T cells that mediate autoimmunity. J Immunol. 2011;186:5738–48.
Chi W, Li F, Chen H, Wang Y, Zhu Y, Yang X, et al. Caspase-8 promotes NLRP1/NLRP3 inflammasome activation and IL-1β production in acute glaucoma. Proc Natl Acad Sci USA. 2014;111:11181–6.
Sahebari M, Rezaieyazdi Z, Nakhjavani MJ, Hatef M, Mahmoudi M, Akhlaghi S. Correlation between serum concentrations of soluble Fas (CD95/Apo-1) and IL-18 in patients with systemic lupus erythematosus. Rheumatol Int. 2012;32:601–6.
Mohsen MA, Abdel Karim SA, Abbas TM, Amin M. Serum interleukin-18 levels in patients with systemic lupus erythematosus: relation with disease activity and lupus nephritis. Egypt Rheumatol. 2013;35:45–51.
Calvani N, Richards HB, Tucci M, Pannarale G, Silvestris F. Up-regulation of IL-18 and predominance of a Th1 immune response is a hallmark of lupus nephritis. Clin Exp Immunol. 2004;138:171–8.
Ebmeier CC, Anderson RJ. Human thyroid phenol sulfotransferase enzymes 1A1 and 1A3: activities in normal and diseased thyroid glands, and inhibition by thyroid hormones and phytoestrogens. J Clin Endocrinol Metab. 2004;89:5597–605.
Choe JY, Kim SK. Serum TWEAK as a biomarker for disease activity of systemic lupus erythematosus. Inflamm Res. 2016;65:479–88.
Al-Janadi M, Al-Balla S, Al-Dalaan A, Raziuddin S. Cytokine profile in systemic lupus erythematosus, rheumatoid arthritis and other rheumatic diseases. J Clin Immunol. 1993;13:58–67.
Mangieri D, Corradi D, Martorana D, Malerba G, Palmisano A, Libri I, et al. Eotaxin/CCL11 in idiopathic retroperitoneal fibrosis. Nephrol Dial Transpl. 2012;27:3875–84.
Tacke F, Trautwein C, Yagmur E, Hellerbrand C, Wiest R, Brenner DA, et al. Up-regulated eotaxin plasma levels in chronic liver disease patients indicate hepatic inflammation, advanced fibrosis and adverse clinical course. J Gastroenterol Hepatol. 2007;22:1256–64.
Huaux F, Gharaee-Kermani M, Liu T, Morel V, McGarry B, Ullenbruch M, et al. Role of Eotaxin-1 (CCL11) and CC chemokine receptor 3 (CCR3) in bleomycin-induced lung injury and fibrosis. Am J Pathol. 2005;167:1485–96.
Diny NL, Hou X, Barin JG, Chen G, Talor MV, Schaub J, et al. Macrophages and cardiac fibroblasts are the main producers of eotaxins and regulate eosinophil trafficking to the heart. Eur J Immunol. 2016; doi: 10.1002/eji.201646557. [Epub ahead of print].
Friese RS, Rao F, Khandrika S, Thomas B, Ziegler MG, Schmid-Schönbein GW, et al. Matrix metalloproteinases: discrete elevations in essential hypertension and hypertensive end-stage renal disease. Clin Exp Hypertens. 2009;31:521–33.
Barksby HE, Milner JM, Patterson AM, Peake NJ, Hui W, Robson T, et al. Matrix metalloproteinase 10 promotion of collagenolysis via procollagenase activation: implications for cartilage degradation in arthritis. Arthritis Rheum. 2006;54:3244–53.
Wu T, Ding H, Han J, Arriens C, Wei C, Han W, et al. Antibody-array-based proteomic screening of serum markers in systemic lupus erythematosus: a discovery study. J Proteome Res. 2016;15:2102–14.
Wang H, Bagavant H, Deshmukh U. Glomerular transcriptional profiles reveal the candidate biomarkers diagnostic for the progression of lupus nephritis from acute to chronic stages. Arthritis Rheum. 2008;58:S317.
Tesch GH, Maifert S, Schwarting A, Rollins BJ, Kelley VR. Monocyte chemoattractant protein 1-dependent leukocytic infiltrates are responsible for autoimmune disease in MRL-Fas(lpr) mice. J Exp Med. 1999;190:1813–24.
Chan RW, Lai FM, Li EK, Tam L, Chow K, Lai K, et al. Intrarenal cytokine gene expression in lupus nephritis. Ann Rheum Dis. 2007;66:886–92.
Menke J, Amann K, Cavagna L, Blettner M, Weinmann A, Schwarting A, et al. Colony-stimulating factor-1: a potential biomarker for lupus nephritis. J Am Soc Nephrol. 2015;26:379–89.
Menke J, Rabacal WA, Byrne KT, Iwata Y, Schwartz MM, Stanley ER, et al. Circulating CSF-1 promotes monocyte and macrophage phenotypes that enhance lupus nephritis. J Am Soc Nephrol. 2009;20:2581–92.
Baranda L, de la Fuente H, Layseca-Espinosa E, Portales-Perez D, Nino-Moreno P, Valencia-Pacheco G, et al. IL-15 and IL-15R in leucocytes from patients with systemic lupus erythematosus. Rheumatology. 2005;44:1507–13.
Zhao M, Wang J, Liao W, Li D, Li M, Wu H, et al. Increased 5-hydroxymethylcytosine in CD4(+) T cells in systemic lupus erythematosus. J Autoimmun. 2016;69:64–73.
Goules A, Tzioufas AG, Manousakis MN, Kirou KA, Crow MK, Routsias JG. Elevated levels of soluble CD40 ligand (sCD40L) in serum of patients with systemic autoimmune diseases. J Autoimmun. 2006;26:165–71.
Ciferska H, Horak P, Hermanova Z, Ordeltova M, Zadrazil J, Tichy T, et al. The levels of sCD30 and of sCD40L in a group of patients with systemic lupus erythematodes and their diagnostic value. Clin Rheumatol. 2007;26:723–8.
Yellin MJ, D’Agati V, Parkinson G, Han AS, Szema A, Baum D, et al. Immunohistologic analysis of renal CD40 and CD40L expression in lupus nephritis and other glomerulonephritides. Arthritis Rheum. 1997;40:124–34.
Ripoll E, Merino A, Herrero-Fresneda I, Aran JM, Goma M, Bolanos N, et al. CD40 gene silencing reduces the progression of experimental lupus nephritis modulating local milieu and systemic mechanisms. PLoS ONE. 2013;8:e65068.
Nakatani K, Yoshimoto S, Iwano M, Asai O, Samejima K, Sakan H, et al. Fractalkine expression and CD16 + monocyte accumulation in glomerular lesions: association with their severity and diversity in lupus models. Am J Physiol Renal Physiol. 2010;299:F207–16.
Inoue A, Hasegawa H, Kohno M, Ito MR, Terada M, Imai T, et al. Antagonist of fractalkine (CX3CL1) delays the initiation and ameliorates the progression of lupus nephritis in MRL/lpr mice. Arthritis Rheum. 2005;52:1522–33.
Han GM, Chen SL, Shen N, Ye S, Bao CD, Gu YY. Analysis of gene expression profiles in human systemic lupus erythematosus using oligonucleotide microarray. Genes Immun. 2003;4:177–86.
Tucci M, Quatraro C, Lombardi L, Pellegrino C, Dammacco F, Silvestris F. Glomerular accumulation of plasmacytoid dendritic cells in active lupus nephritis: role of interleukin-18. Arthritis Rheum. 2008;58:251–62.
Bracci-Laudiero L, Aloe L, Levi-Montalcini R, Galeazzi M, Schilter D, Scully JL, et al. Increased levels of NGF in sera of systemic lupus erythematosus patients. Neuroreport. 1993;4:563–5.
Antonucci MT, Bonofiglio R, Papalia T, Caruso F, Caroleo MC, Mancuso D, et al. Nerve growth factor and its monocyte receptors are affected in kidney disease. Nephron Clin Pract. 2009;111:c21–8.
Morigi M, Locatelli M, Rota C, Buelli S, Corna D, Rizzo P, et al. A previously unrecognized role of C3a in proteinuric progressive nephropathy. Sci Rep. 2016;6:28445.
Onodera H, Nagata T, Kanazawa M, Taguma Y, Itoyama Y. Increased plasma GDNF levels in patients with chronic renal diseases. Nephrol Dial Transplant. 1999;14:1604–5.
Kalechman Y, Sredni B, Weinstein T, Freidkin I, Tobar A, Albeck M, et al. Production of the Novel Mesangial Autocrine Growth Factors GDNF and IL-10 Is Regulated by the Immunomodulator AS101. J Am Soc Nephrol. 2003;14:620–30.
Authors’ contributions
All authors contributed substantially to the conception and design, analysis and interpretation of data, drafting the article or revising it critically for important intellectual content. Final approval of the version to be published was given by all the authors. All authors read and approved the final manuscript.
Acknowledgements
We apologize to many authors whose important works could not be cited due to space limitations. We thank Martin Radvansky and Sarka Zehnalova for their kind help with preparing figures for Additional file.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All data generated or analysed during this study are included in this published article and its additional file.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Patients and control subjects provided written informed consent about the usage of peripheral blood for the purpose of this study which was approved by the ethics committee of University Hospital and Palacky University Olomouc.
Funding
Funding was obtained from the Ministry of Health of Czech Republic (MZ CR VES15-28659A).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Petrackova, A., Smrzova, A., Gajdos, P. et al. Serum protein pattern associated with organ damage and lupus nephritis in systemic lupus erythematosus revealed by PEA immunoassay. Clin Proteom 14, 32 (2017). https://doi.org/10.1186/s12014-017-9167-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12014-017-9167-8