
Abstract
There is an increasing focus on the development of cheese with reduced-fat and salt contents, owing to greater consumer focus on the profiling and intake management of nutrients. The current study evaluated the effects of reducing fat and salt, each by 30 and 50%, on the composition, microbiological and biochemical characteristics of Cheddar-style cheese over a 270-day ripening period. Reducing salt from 1.9 to 1.2 or 0.9% (w/w) significantly increased (P < 0.05) moisture, lactic acid, lactic acid-to-protein ratio and water activity and reduced the mean level of residual lactose and starter culture die-off during ripening. Reducing fat from 33 to 22 or 16% (w/w) resulted in significant increases in moisture, protein and lactic acid and reductions in salt-in-moisture, moisture-in-non-fat substances and lactic acid-to-protein ratio. Lactose content, pH and water activity were significantly affected by the interaction of fat and salt. The pH changed during the course of ripening, decreasing in full-fat full-salt cheese, increasing in half-fat reduced-salt and half-fat half-salt cheeses and remaining constant in other cheeses with different fat and salt levels. After 270 days of maturation at 8 °C, the full-fat full-salt cheese had lower concentrations of lactic acid and free amino acids and pH and a higher lactic acid-to-protein ratio than the reduced-fat reduced-salt or half-fat half-salt cheeses. The current results, together with a follow-on communication on the impacts of reducing fat and salt on physical and sensory properties, will provide a knowledge platform which should facilitate the commercial development of reduced-salt, reduced-fat, dry-salted cheeses.
Similar content being viewed by others
1 Introduction
Cheese is a significant food in the diet in the Western World, being an important source of general nutrients and a very rich source of protein and calcium. Apart from this, cheese offers the consumer large diversity in flavour, texture, appearance, cooking properties and usage applications and is extremely versatile as an ingredient in culinary and fast food. Consequently, cheese production continues to expand, from ∼15 to >18 M t between 2005 and 2014 (FAO 2014). The increased global consumption of cheese has been paralleled by more exacting demands for consistency of flavour, functional properties and levels of specific nutrients such as sodium and fat content, driven by an increasing consumer awareness of the role of diet in lifestyle and health. High levels of dietary sodium and fat can adversely affect health, lifestyle and longevity (Flegal et al. 2013; He and MacGregor 2009; Kalogeropoulos et al. 2015). The average daily intake of sodium and fat in the modern western diet is higher than the RDI (UK Department of Health 2011; WHO 2013). Numerous countries including Canada, Finland, Ireland and the UK have national strategies in place to reduce dietary salt intake (Henney et al. 2010).
The levels of salt and fat in cheese differ significantly with variety (e.g. <0.2% salt and <1.8% fat in Quark compared to ∼3.5 and 32.9% for Roquefort), allowing the informed consumer ample choice in respect to sodium and fat intake. Overall, cheese even at relatively high per capita consumption rates (15 kg per annum) contributes <10% of dietary sodium. Nevertheless, cheese is perceived as having high levels of fat and sodium (Johnson et al. 2009). Consequently, manufacturers in pursuit of greater market share continue to develop and improve reduced-fat or reduced-salt variants of the major cheese varieties such as Cheddar. Extensive research has been undertaken to determine the effects of reducing fat content and means to improve the quality of reduced-fat cheese varieties such as Cheddar and Mozzarella (Fenelon and Guinee 2000; Fenelon et al. 2000; Rogers et al. 2009; Rudan et al. 1999; Sánchez-Macías et al. 2010; van Hekken et al. 2007). The key approaches have been reviewed by Johnson et al. (2009) and Møller (2012) and include reducing the volume fraction of the casein network by increasing moisture level or the inclusion of filler materials such as microparticulated whey proteins and hydrocolloid-based materials; reducing the degree of casein cross-linking through reduction in calcium phosphate level; increasing casein hydrolysis; the use of improved starter culture and starter-culture adjuncts, increasing proteolysis and adding fat-derived flavours. Similarly, the effects of reducing salt content have been extensively reported, with most emphasis on full-fat cheese (Arboatti et al. 2014; Czarnacka-Szymani and Jezewska-Zychowicz 2015; Ganesan et al. 2014; Lu and McMahon 2015; Ma et al. 2013; Murtaza et al. 2014; Pastorino et al. 2003; Rulikowska et al. 2013). Apart from the potential risk to safety and microbiological quality (Labrie et al. 2014), a major issue with reduced-salt cheeses is low pH, which is conducive to higher residual coagulant activity, greater hydrolysis of β-casein and an increased risk of bitterness (Guinee and Fox 2004). A recent integrated approach involved the combined effects of an optimized make procedure (normalized moisture content, low proteolytic activity coagulant) and starter cultures (the use of glutamate decarboxylase-negative starter culture and an adjunct Lactobacillius culture with high peptidase and autolytic activities and a capacity to convert citrate to succinic acid) to improve quality in reduced-salt Cheddar (Møller 2012; Møller et al. 2013). Comparatively, few studies have considered the effects of reducing salt in reduced-/low-fat cheese (Paulson et al. 1998; Skeie et al. 2013). Attempts to improve the quality of reduced-sodium cheese have focused mainly on the replacement of sodium chloride with other salts such as potassium chloride or magnesium chloride (Lu and McMahon 2015; McMahon et al. 2014; Thibaudeau et al. 2015).
More recently, there is an increasing commercial interest in the development of hard cheeses with reduced salt and fat levels. As cited above, various studies have considered the separate effects of reducing salt or fat in cheeses. The confounding effects of inter-study differences in cheese type, milk composition, manufacturing procedure and levels of fat or salt reduction make it difficult to discern the interactive effects of fat and salt reduction from published literature. We are unaware of any previous studies having considered the effects of reducing salt and fat in cheeses simultaneously. The current study was undertaken as part of a major national Irish collaboration entitled Cheese Board 2015 (http://www.cheeseboard2015.com) involving cheese manufacturers and institutional research providers to address knowledge gaps in a number of key areas—among them to build technological capacity for the production of reduced fat, reduced salt cheese by pushing the boundaries of Cheddar cheese manufacturing protocols. Hence, the experimental objectives of the present work, i.e. to investigate the effects of the simultaneous incremental reduction of fat and salt, from ∼33 to 16% and from 1.9 to 0.9%, respectively, on the composition and changes in lactose metabolism, water activity and microbiology of Cheddar cheese, made from the same milk under standardized conditions, during ripening. Complementary papers to follow will report on the effect of reducing salt and fat on age-related changes in rheology, proteolysis, lipolysis, volatile compounds and sensory characteristics of these cheeses.
2 Materials and methods
2.1 Standardization and pasteurization of milks
Raw milk was obtained from the winter-calving Moorepark Holstein Friesian herd. Part of the milk was separated at 55 °C to give skim and cream (48% w/w fat). A portion of the skim milk was ultrafiltered to 11 to 14% w/w protein as described by Guinee et al. (2006). Standardized milks with a protein content of 4.0% (w/w) and a protein-to-fat ratio (PFR) of 0.9, 1.62 or 2.65 were prepared by blending skim milk (∼0.07% w/w fat, 3.6% w/w protein), UF retentate (12% w/w protein), raw milk and cream (35% w/w fat) in appropriate quantities. The standardized milks were stored overnight at 8 °C, pasteurized at 72 °C for 15 s, cooled to 31 °C and pumped into 500-L stainless steel, jacketed, cylindrical cheese vats.
2.2 Cheese manufacture
Cheeses with target fat levels of ∼33, 22 and 16% w/w were prepared from the standardized milks (480 kg) as described by Hou et al. (2012). Essentially, the pasteurized milks were inoculated with a starter culture comprising of two Lactococcus lactis strains: subsp. lactis and subsp. cremoris (Chr. Hansen Ireland Ltd, Rohan Industrial Estate, Little Island, Co. Cork, Ireland). After ripening for 30 min at 31 °C, rennet (Chymax Plus, Chr. Hansen Ireland Ltd, 200 IMCU.mL−1), diluted ∼1:10 in de-ionized water, was added at a level of 0.18 mL.kg−1 based on a protein level of 3.4% (w/w). The rennet-induced milk gel was cut at a firmness of 25 Pa, as measured using low-amplitude strain oscillation rheometry (Hou et al. 2012). The curd-whey mixture was cooked to 38 °C, and the whey was drained at pH 6.15. The curd was held at ∼36 °C in jacketed vats until the pH reached 5.35. The curd from each vat was divided into three equal portions, each salted at a level of 2.59% (w/w), 1.50% (w/w) or 0.99% (w/w). The curds and salt were thoroughly mixed, and the salted curds were again manually mixed after 5, 10, 15 and 20 min during the 20-min mellowing period before being moulded, pre-pressed at 0.13 kPa for 30 min and pressed overnight at 2.5 kPa. Cheeses, which ranged in weight from 14.5 to 20 kg, depending on levels of fat and salt, were vacuum-packed and stored at 4 °C for 30 days and 8 °C for the remainder of the 270-day ripening period.
Three replicate cheesemaking trials were undertaken over a 3-week period from January 16 to January 30, 2013. Full-fat (FF, 33%), reduced-fat (RF, 22%) and half-fat (HF, 16%) Cheddar-style cheeses, each with full-salt (FS, 1.9%), reduced-salt (RS, 1.2%) and half-salt (HS, 0.9%) variants, were manufactured in all trials. The resultant cheeses were coded as follows: full-fat full-salt (FFFS), full-fat reduced-salt (FFRS), full-fat half-salt (FFHS), reduced-fat full-salt (RFFS), reduced-fat reduced-salt (RFRS), reduced-fat half-salt (RFHS), half-fat full-salt (HFFS), half-fat reduced-salt (HFRS), half-fat half-salt (HFHS).
2.3 Sampling of cheese
Cheeses were sampled at 1, 14, 30, 90, 150, 210 and 270 days. On each sampling date, a vertical slice, ∼0.5 cm thick, was removed from one of the vertical outside faces of the block and discarded, and a slab (∼6 cm thick, ∼2 kg), including the freshly exposed surface, was taken for analysis. Plug samples (∼10 g), obtained using a sterile cheese trier, were taken for microbiological analyses. Samples were analysed for pH, gross composition, free amino acids and water activity within 48 h. Samples of cheese for lactose and lactic acid analysis were frozen immediately after sampling and, when required for analysis, were subsequently thawed overnight at 4 °C and analysed within 12 h.
2.4 Composition at 14 days
Grated cheese samples were analysed at 14 days in triplicate for protein, using the Kjeldahl method, fat, NaCl, moisture and ash, using standard International Dairy Federation (IDF) methods, as described by Hou et al. (2012). Chesses were ashed at 550 °C and analysed for calcium using standard IDF methods (Guinee et al. 2000). The pH was measured on a cheese slurry prepared from 20 g of cheese and 12 g of distilled H20 after each sampling date.
2.5 SLAB and NSLAB counts
Cheeses were assayed in duplicate for both starter bacteria (SLAB) and non-starter lactic acid bacteria (NSLAB) on days 14, 30, 90 and 150. Initial 1:10 dilutions were prepared by stomaching 10 g of cheese with 90 mL of sterile, 2% (w/v) trisodium citrate for 5 min. SLAB were plated on Lactose M17 agar (Sigma-Aldrich) (Terzaghi and Sandine 1975) and incubated at 30 °C for 3 days, while NSLAB were plated on Lactobacillus selection agar (LBS) (Sigma-Aldrich) with an overlay and incubated aerobically for 5 days at 30 °C (Ragosa et al. 1951).
2.6 Lactose and lactic acid
The concentration of lactose was determined according to Rynne et al. (2007), using Megazyme Lactose and D-Galactose (Rapid) Assay procedure, and that of L (+) and D (−) lactic acid by D-/L-Lactic Acid (Rapid) Assay procedure (Megazyme International Ireland, Bray Business Park, Bray, Co. Wicklow, Ireland). The total lactate was calculated as the sum L (+) and D (−) lactic acid. Each analysis was performed in triplicate.
2.7 FAA concentration
The level of free amino acid (FAA) was measured on a clarified aqueous pH 4.6 soluble extract of the cheese at days 1, 14, 30, 90, 150 and 270, as described by Fenelon et al. (2000). The clarified extract was analysed in triplicate for levels of FAA on a Beckman 6300 High Performance Analyzer (Beckman Instruments Ltd., High Wycombe, UK) using ion-exchange chromatography with postcolumn ninhydrin derivatization and visible colorimetric detection at 570 nm for all amino acids.
2.8 Water activity (a w)
Grated cheese samples (0.5 g) were placed in plastic dishes, covered, stored at 8 °C for 30 min and assayed in duplicate at 25 °C on a Novasina LabMaster AW machine (Novatron Scientific Ltd., Novatron House, 46 London Road, Horsham, West Sussex, RH12 1AY, UK), based on changes in electrolytic resistance. Prior to analysis, the machine was calibrated using a certified Novasina 98% humidity standard (cert no. NS2984-2). The a w of the cheese was measured at 1, 14, 30, 90, 150 and 270 days.
2.9 Statistical analysis
Three replicate cheesemaking trials were undertaken, each with nine treatment cheeses, namely FFFS, FFRS, FFHS, RFFS, RFRS, RFHS, HFFS, HFRS and HFHS. Analysis of variance (ANOVA) was undertaken using the general linear model (GLM) procedure of SAS 9.3 (SAS Institute 2011) to determine the effect of fat or salt on cheese composition variables at 14 days. Tukey’s multiple comparison test was used for paired comparison of treatment means, and the level of significance was determined at P < 0.05.
A split-plot design was used to determine effect of salt or fat and ripening time and their interaction on the specific variables (such as lactose) measured at regular intervals throughout ripening time. Split-plot was also used to determine potential interactive effects of salt and fat on all variables, at different ripening times. Analysis for the split-plot design was carried out using the PROC MIXED procedure of SAS 9.3 (SAS Institute 2011). Statistically significant differences (P < 0.05) between different treatment levels were determined by Tukey’s multiple comparison test. Excel 2010 software was used to calculate the significance of correlations where n is the actual number of data points, and df is the degrees of freedom.
3 Results
3.1 Cheese composition at 14 days
Effect of salt reduction
The compositions of all the cheeses at 14 days are shown in Table 1. Reducing salt content significantly increased the mean levels of moisture, lactic acid and the lactic acid-to-protein ratio (LA:P, an index of the ratio of lactic acid-to-buffering capacity) and reduced the levels of residual lactose in FF, RF and HF cheeses. The level of moisture-in-non-fat substances (MNFS) increased in the RF and HF cheeses, but not in the FF cheese, as the salt content was reduced. Commensurate with the increase in concentration of total lactic acid, the mean pH of the 14-day-old FF, RF and HF cheeses decreased significantly as the salt content was reduced from 1.9 to 1.2 or 0.9%. Linear regression showed significant positive correlations between salt-in-moisture content (S/M) and pH in the FF cheese at all ripening times ≤150 days (r = 0.74 to 0.94, df = 7), in the RF cheese at times ≤30 days (r = 0.78 to 0.89, df = 7) and in the HF cheeses at 1 and 14 days only (r = 0.72 to 0.91, df = 7).
Effect of fat reduction
Reducing the fat content from 33 to 16% (w/w) significantly increased the levels of moisture, protein, lactic acid and calcium and reduced the levels of S/M, MNFS and LA:P in the FS, RS and HS cheeses (Table 1). While the protein content increased significantly on reducing fat from ∼33 to 22% or from 33 to 16%, it did not change significantly on reducing fat from 22 to 16%, reflecting the increasing difficulty in concentrating protein beyond a critical level (44 to 45 g/100.g protein + moisture) during cheese manufacture. The pH of the 14-day-old RS and HS cheese variants increased significantly when the fat content was reduced from 33% to 22 or 16%, unlike the pH of the corresponding FS cheeses which was not affected by fat reduction. The level of residual lactose in the RS and HS cheese variants at 14 days was very low (<0.05%), while that in the FS variant decreased significantly as the fat content was reduced, from ∼0.34% in the FF cheese to ∼0.12% in the HF cheeses.
Analysis of the compositional data at 14 days indicated that lactose content and pH were significantly affected by the interaction of salt and fat (Table 1).
3.2 Changes in SLAB and NSLAB during ripening
The SLAB cell count was significantly affected by salt content and ripening time in the FF, RF and HF cheeses and by the interaction of salt and ripening time in the FF cheese (Fig. 1, Table 2).

Changes in starter culture (SLAB) population in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The SLAB cell count was ∼109.5 cfu.g−1 in all cheeses at day 1. The mean count in the FF, RF and HF cheeses decreased significantly during ripening, with the decrease being the highest in the FS variants and lowest in the HS variants (Fig. 1a–c). Hence, for each fat level, the SLAB population was lowest in the FS variant and highest in the HS variant at 90 and 150 days. Altering fat level did not influence the mean SLAB count in the FS, RS or HS cheese variants (Fig. 1d–f, Table 3). There was an interactive effect of salt and fat on SLAB counts at day 14 but not at any other time (Table 4).
The population of NSLAB in the FF, RF and HF cheeses increased significantly during maturation, from ∼103 to 103.5 cfu.g−1 at day 1 to ~107 to 108 cfu.g−1 at day 150 but was not significantly affected by salt content (Table 2). Similarly, fat level did not influence the mean NSLAB numbers in the FS, RS or HS cheeses (Table 3).
3.3 Changes in lactose during ripening
The effects of fat and salt on the changes in lactose over the course of ripening are shown in Fig. 2 and Tables 2 and 3. The level of lactose in the FF, RF and HF cheeses was significantly affected by salt, ripening time and the interaction of salt and ripening time (Table 2). The levels of lactose at 1 day varied markedly, from ~0.2–0.45% in the FS variants, ≤0.14% in the RS cheeses, to ≤0.05% in the HS cheese variants; by day 270, all cheeses were essentially free of lactose (Table 5).

Changes in levels of residual lactose in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The lactose concentration decreased significantly in the FS and RS cheese variants during ripening (Fig. 2a–c); in contrast, the initial day 1 levels were very low (≤0.05%) in all the HS cheese variants and remained relatively constant during ripening (Fig. 2). Reducing salt content had a significant effect on the mean lactose levels over the 270-day ripening period in the FF, RF and HF cheeses (Fig. 2, Table 2), with the mean level in the FS cheeses being significantly higher than that in the RS or HS variants of the FF, RF and HF cheeses. The magnitude of the difference in lactose between the FS and the RS or HS variants decreased as the fat content of the cheese decreased (Fig. 2). Hence, the effect of the interaction between salt content and ripening time on lactose concentration was significant for the FF, RF and HF cheeses.
Reducing the fat content significantly reduced the mean level of lactose during the 270-day ripening period in the FS cheeses but did not affect the lactose level of the RS or HS cheeses (Fig. 2d–f, Table 3). The more rapid lactose depletion in the RFFS and HFFS cheeses compared to the FFFS cheese reflects a lower initial level of lactose together with a more rapid lactose metabolism associated with a lower S/M content (for a given salt level on a total weight basis). There was an interactive effect between fat content and ripening time in the FS cheese with the difference between the FS and RS or HS variants diminishing with ripening time.
There was an interactive effect of salt and fat on lactose concentration at 14, 30, 90 and 150 days (Table 4), but not at 210 or 270 days (Table 5), by which time residual lactose in the FFFS cheese had decreased to levels of ≤0.06%.
3.4 Changes in lactic during ripening
The concentration of lactic acid was significantly affected by salt and ripening time in the FF, RF and HF cheeses (Fig. 3a–c, Table 2). ANOVA showed that the lactic acid content of all cheeses increased significantly with ripening time and with extent of salt reduction in the FF, RF and HF cheeses. There was an interactive effect between salt and ripening time on the concentration of lactic acid in the HF cheese, with the difference between FS and the HS variants increasing with maturation time.

Changes in levels of total lactic acid in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The lactic acid level in the FS, RS and HS cheeses was significantly affected by fat content and the interaction of fat and ripening time (Fig. 3d–f, Table 3). For each salt level, the mean level of lactic acid over ripening was higher in HF cheese than in the FF or RF cheese which had similar levels.
L (+) and D (−) lactic acid
The concentration of L (+) lactic acid decreased gradually in all cheeses during ripening, while that of D (−) lactic acid simultaneously increased (Fig. 4). The concentration of L (+) lactic acid in the FF, RF and HF cheeses during ripening showed a similar trend to total lactic acid, being highest in the HS and lowest in the FS variant. Similarly, the mean concentration of D (−) lactic acid was highest in the HS and lowest in the FS variant of the FF and RF cheeses but was similar in all three HF cheeses. Hence, the ratio of D (−)-to-L (+) lactic acid increased significantly in all cheeses during ripening but was not affected by salt level.

Changes in the levels of L (+) lactic acid (broken line, filled symbol) and D (−) lactic acid (solid line, open symbol) in half-fat Cheddar-style cheeses. The cheeses, described in Table 1, include full-salt (filled circles, empty circles), reduced-salt (filled squares, empty squares) and half-salt (filled triangles, empty triangles) variants of the half-fat cheese. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The concentration of L (+) lactic acid in the FS, RS and HS cheeses was significantly affected by fat content, while that of D (−) lactic acid was affected by the interaction of fat and ripening time (Table 3). The mean concentration of L (+) lactic acid over the ripening period was highest in the HF cheese and lowest in the FF cheeses. In contrast, fat content did not significantly influence the mean concentration of D (−) lactic acid in the FS, RS or HS cheeses during ripening.
There was no interaction of salt and fat on the concentrations of total, L (+) or D (−) lactic acid (Table 4).
3.5 Changes in pH during ripening
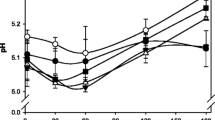
The pH of the FF, RF and HF cheeses was significantly affected by salt content, ripening time and their interaction (Fig. 5, Table 2).

Changes in pH in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The mean pH over the 270-day ripening period decreased significantly in the FFFS cheese, increased in the HFRS and HFHS cheeses and remained constant in the FFRS, FFHS, RFFS, RFRS, RFHS and HFFS cheeses. For each fat level, the pH differences between cheeses with different salt content were highest between 1 to 30 days when the pH of the FS variant was significantly higher than that of the corresponding RS or HS variants, especially in the FF cheese (Fig. 5a–c). The largest differences between cheeses of different salt levels were observed in the FF cheese, with the mean pH of the FFFS cheese over ripening being ~0.2 U higher than the FFHS cheese. Differences in pH between the FS and the RS or HS variants diminished quickly in the RF and HF cheeses. After 270 days, the pH of the FS, RS and HS variants were similar in the FF, RF and HF cheeses (Table 5). Nevertheless, the mean pH at 270 days was highest in the HF cheeses and lowest in the FF cheeses, reflecting a trend similar to that for protein content (Table 1), but opposite to that for LA:P.
The pH of the FS, RS and HS cheese variants was significantly affected by fat content and by the interaction of fat and ripening time in the FS and RS cheeses (Fig. 5d–f, Table 3). The mean pH of the RS and HS cheese variants over the 270-day ripening period increased significantly as the fat content was reduced, while that of the FS variant was unaffected by fat level. At each salt level, the difference in pH between the cheeses of different fat contents increased during maturation. Despite fat level having no effect on the mean pH of the FS cheeses over the ripening period, the pH of the 270-day-old HFFS cheese was ~0.15 pH units higher that of the FFFS cheese.
Salt and fat level interactively affected the pH at 14, 30, 90, 150 and 210 days but not at day 270 when differences in pH between the FS, RS and HS variants at each fat level had, essentially, disappeared (Table 4).
3.6 Changes in FAA content
The concentration of FAA was significantly affected by salt level, ripening time and their interaction in the FF cheese and by ripening time only in the RF and HF cheeses (Fig. 6, Table 2). The levels of FAA increased significantly in all cheeses during ripening, from ≤1000 mg.kg−1 at day 1 to mean values of ~4500 to 6000, 6200 to 6400 and 6500 to 7000 mg.kg−1 in the 270-day-old FF, RF and HF cheeses, respectively, depending on salt content (Table 5).

Changes in the concentration of free amino acids (FAA) in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
Reducing salt content significantly affected the mean level of FAA during ripening in the FF cheese, with the mean concentration in the FFFS cheese variant being significantly higher than that in the FFRS or FFHS variants which were similar (Fig. 6a–c). ANOVA of the data at different ripening times showed that the effect of salt was most pronounced after 150 and 270 days. In contrast, salt level did not influence the mean level of FAA in the RF and HF chesses over ripening.
The mean level of FAA over ripening in the FS, RS and HS cheeses was significantly affected by fat content and the interaction of fat and ripening time (Fig. 6d–f). For each salt level, the mean FAA concentration over ripening was lowest in the FF cheese and highest in the RF or HF cheeses which did not differ significantly. The difference between the FF cheeses and the RF or HF cheeses at each salt level generally increased with ripening time.
There was no interaction between salt and fat on the concentration of FAA at any ripening time (Table 4).
3.7 Changes in a w during ripening
The a w of the FF, RF and HF cheeses was significantly affected by salt level and ripening time (Fig. 7, Table 2). The mean a w of these cheeses decreased significantly during ripening, from ~0.968 to 0.978 in all 1-day-old cheeses to ~0.945 to 0.955 at day 270 depending on salt and fat level (Table 5). The rate of decrease was most rapid between 1 and 14 days.

Changes in water activity (a w) in Cheddar-style cheeses with different fat and salt levels during ripening. The cheeses, described in Table 1, include full-salt (filled circles), reduced-salt (filled squares) and half-salt (filled triangles) variants of full-fat (a), reduced-fat (b) and half-fat (c) cheeses, and full-fat (empty circles), reduced-fat (empty squares) and half-fat (empty triangles) variants of full-salt (d), reduced-salt (e) and half-salt (f) cheeses. Presented values are the means of three replicate trials; error bars represent standard deviations of the mean
The mean a w of the FF, RF and HF cheeses over the 270-day ripening period was significantly affected by salt content (Table 2), with the mean a w of the FS variants of the FF, RF and HF cheeses being significantly lower than those of the corresponding RS or HS variants, which were similar (Fig. 7a–c). The a w of the FS cheese variants was significantly affected by fat content (Fig. 7d–f, Table 3), with the mean a w of the FFFS variant being significantly lower than that of the corresponding RFFS or HFFS cheeses for which the a w was similar.
There was a significant interaction between salt and fat on the a w at 30 and 90 days (Table 4).
4 Discussion
The current study focussed on reducing salt by 30 and 50% in FF (33%), RF (22%) and HF (16%) Cheddar cheeses, in line with an evolving interest in consumer demand and development of reduced-salt, reduced-fat cheeses (Johnson et al. 2009).
Compositionally, reducing salt and fat resulted in additive increases in moisture and lactic acid and a reduction in S/M. However, they had opposite effects on the ratios of moisture-to-protein (as reflected by MNFS) and LA:P, both of which decreased as fat content was reduced. Protein content increased significantly with fat reduction but was unaffected by salt content. The above compositional changes concur with the separate effects of fat and salt reduction reported in previous studies on cheeses such as Cheddar, Mozzarella and Gouda (Arboatti et al. 2014; Czarnacka-Szymani and Jezewska-Zychowicz 2015; Fenelon and Guinee 2000; Kelly et al. 1996; Rulikowska et al. 2013; Tunick et al. 1993). Hence, reducing salt and fat had opposite effects on contents of MNFS and LA:P, which, along with S/M, are considered to be key compositional parameters in controlling Cheddar cheese quality (Lawrence et al. 2004). Both S/M content and LA:P have a major influence on cheese pH (Guinee and Fox 2004).
While all cheeses had similar SLAB counts at 14 days, starter die-off during ripening was more rapid in the FS variant compared to the HS variant in the FF, RF and HF cheeses. The slower decrease in the HS variant during ripening concurs with the findings of Rulikowska et al. (2013) for FF Cheddar and reflects a lower degree of salt-induced starter cell autolysis (Ramírez-Nuñez et al. 2011; Wilkinson et al. 1994). In contrast to SLAB, salt content did not affect the rate of growth or the mean NSLAB counts of the FF, RF or HF cheeses over ripening, which is consistent with the relatively high salt tolerance of NSLAB compared to SLAB (Turner and Thomas 1980).
The relatively high salt and, hence, S/M level, in the FS variant of the FF, RF and HF cheeses resulted in lactose being utilized relatively slowly, especially in the FFFS cheese where significant levels of lactose (~0.15 to 0.45%, w/w) were present at times ≤150 days. The slower lactose metabolism in the FFFS cheese compared to the RFFS and HFFS cheeses reflects its higher S/M, which would be more inhibitory to SLAB metabolism (Thomas and Pearce 1981). The FS variant of the FF, RF and HF cheeses had a relatively low mean level of lactic acid over ripening because of the slow utilization of lactose and the relatively low moisture content (which is the solvent for lactose). Owing to the combined effects of a relatively high S/M and low LA:P, the pH of the FS variant of the FF, RF and HF cheeses decreased relatively slowly during ripening, with the effect being most pronounced in the FS cheese (Fig. 5) where the pH decreased gradually from ~5.35 to 5.20 over 270 days. Similar to the FFFS cheese in the current study, relatively high levels of residual lactose have also been reported by others in full-salt aged Cheddar cheeses with S/M levels of 5.0 to 5.5% (Chou et al. 2003; Hou et al. 2012; Jordan and Cogan 1993). Other studies on full- or reduced-fat Cheddar cheese found that lactose was metabolized rapidly within 1 to 30 days after manufacture (Rynne et al. 2007). Inter-study discrepancies in lactose content probably relate to variations in the salt tolerance of the starter cultures, the lactose content of the cheese milk and the moisture and calcium contents of the cheese (Hou et al. 2012, 2014; Upreti and Metzger 2006). Lactose being a reducing sugar, the persistence of residual lactose in the FFFS cheese, at levels of ~0.15 to 0.45% (w/w) between 1 and 150 days, could contribute to Maillard reactions and browning especially in heated cheese applications such as pizza, where it may be desirable or undesirable depending on brand and types. Nevertheless, the level of residual lactose in the FS cheeses could be reduced through appropriate use of more salt-tolerant starter cultures (Rulikowska 2009) or curd washing (Hou et al. 2012).
Conversely, the low S/M in the HS variants of the FF, RF and HF cheeses resulted in all lactose being effectively fermented to lactic acid by the SLAB during overnight pressing or within the first 1 to 14 days in the RS variants. Such a trend concurs with the results of Irvine and Price (1961) who found that the growth of the starter Lactococci in skim milk was enhanced at S/M levels of 1.0 to 1.5% and inhibited increasingly as S/M was increased from 2 to 5%.
The RS and HS variants of the FF cheese had a lower mean pH over ripening than the corresponding FSFF variant because of their higher contents of moisture and lactic acid and higher LA:P ratio. However, this was not the case for the RF and HF cheeses where the pH of the FS variant did not significantly differ from that of the RS or HS variants, except at times <30 days. The absence of a pH difference between the FS and RS or HS variants of the HF cheeses can be explained partly by the increase in pH of the RS and HS variants versus the constancy of pH in the FS variant during ripening.
In agreement with the results of Fenelon and Guinee (2000), the mean 270-day pH of the FF cheeses (FFFS, FFRS and FFHS) was significantly lower than that of the corresponding RF or HF cheeses despite the higher S/M content of the former (Table 5). The lower pH of the FF cheeses, despite their low level of lactic acid (Fig. 3) relative to the corresponding RF and HF cheeses, may appear surprising based on the findings of previous studies (Hou et al. 2012, 2014; Huffman and Kristoffersen 1984; Shakeel-Ur-Rehman et al. 2004) that show an inverse relationship between pH and lactic acid content. The lower pH of the FF cheeses is likely to be associated with their relatively high LA:P (Hou et al. 2014) and low concentration of FAA. Similarly, previous studies (Fenelon and Guinee 2000; Fenelon et al. 2000) have reported that the pH of half-fat Cheddar cheese increased significantly during ripening (unlike FF Cheddar where the pH remained constant) and that this increase coincided with the development of relatively high concentrations of FAA. The increase in FAA, concomitant with hydrolysis of peptide bonds, results in the generation of −COO− and −NH3 + groups in the cheese environment with the former likely to electrostatically associate with H+ ions. At the LA:P of standard FFFS Cheddar cheese, the protonation of carboxyl and amino groups does not normally lead to an increase in cheese pH because of the excess H+ ions from the lactic acid, which is expected to be fully dissociated in the cheese environment (pKa ∼ 3.86). However, the pH is more likely to increase as the fat content of the cheese is reduced owing to the reduction in LA:P and the increase in concentration of FAA (Fig. 6) which would be more conducive to reducing H+ activity. Hence, while S/M controls the rate or extent of lactose fermentation and, hence, lactic acid content during ripening, the pH of the cheese for a given level of lactic acid depends on the ability of the calcium phosphate para-casein network to buffer the acid and thereby control the pH. Hence, the level of S/M and fat, which is inversely related to the volume fraction of the protein network, interactively affected the pH of the cheeses (Table 4, Fig. 5).
The decline in lactose concentration in the FFFS, RFFS and HFFS cheeses during ripening time (e.g. by 0.43% in the FFFS cheese) coincided with an increase in content of total lactic acid (0.12% w/w). However, the increase in concentration of total lactic acid in the FFFS cheese was lower than expected based on the molar ratio of lactose-to-lactic acid (1:4) and the molecular mass of lactose and lactic acid. This lower-than-expected increase in lactic acid in the FFFS cheese, which is also apparent from the data of previous studies on Cheddar cheese (Chou et al. 2003; Hou et al. 2012; Turner and Thomas 1980), suggests that at least some of the lactose was used by the NSLAB which grew from ~103 cfu.g−1 at day 1 to ~108 cfu.g−1 at day 150. Facultative heterofermentative NSLAB strains (e.g. Lactobacillus casei, Lactobacillus paracasei, Lactobacillus plantarum), which constitute the major NSLAB species in Cheddar cheese, have been found to metabolize lactose to compounds (e.g. acetate, galactose) which were not measured in the current study (Jordan and Cogan 1993; Swearingen et al. 2001). However, even where lactose was utilized quickly (e.g. within the first 14 days in the RS or HS variant of the FF, HF and RF cheeses), NSLAB grew to the same level as in the FS variant where lactose persisted for a longer time. This reflects the ability of NSLAB to utilize substrates in the cheese environment, apart from lactose, to support growth, including amino acids, fatty acids, glycerol or carbohydrates released from glycomacropeptide of κ-casein (galactose, N-acetylgalactosamine, N-acetylneuraminic acid) and from glycoproteins and glycolipids in the milk fat globule membrane (Sgarbi 2012). The increase in lactic acid content during ripening has been reported widely for Cheddar (Chou et al. 2003; Hou et al. 2012; McMahon et al. 2014), despite the lactose being utilized within the first few weeks of ripening in many instances (Chou et al. 2003; Rynne et al. 2007). Similarly, lactic acid increased slowly in the RS and HS variants of the FF, RF and HF cheeses over ripening even though no lactose remained in these cheeses at times ≤14 days. This trend again suggests the metabolism of compounds (apart from lactose) to lactic acid by NSLAB, especially in the RS and HS cheese variants at all fat levels.
The decrease in the a w during ripening from a mean of 0.97 in all cheeses at day 1 to ~0.94 at 270 days is consistent with trends reported previously for Emmental (Rűegg and Blanc 1977) and Cheddar (Hickey et al. 2013). The decrease in a w with ripening time is likely to reflect the concomitant increase in the concentration of FAA, which has been found to be a key factor for the age-related depression of a w in Cheddar cheese with a standard salt content (Hickey et al. 2013). Hence, linear regression analysis indicated a significant inverse relationship (r = 0.71, P < 0.05) between the FAA and a w data for the nine cheeses at the different ripening times in the current study. Nevertheless, the lower mean a w of the FFFS, RFFS and HFFS cheeses over ripening compared to the corresponding FFHS, RFHS and HFHS cheeses concurs with the inverse relationship between salt content and a w in experimental Cheddar cheeses with salt levels in the range 0.5 to 3.0% (w/w) (Murtaza et al. 2014; Rulikowska et al. 2013) and in an assortment of retail cheeses with moisture and S/M levels ranging from 34.8 to 55.1 and 1.45 to 11.1%, respectively (Marcos et al. 1981). The results indicate the predominance of S/M over FAA in controlling the water activity of cheeses where salt content varies to the extent in the current study. The increase in concentration of FAA as fat content was reduced confirms the findings of Fenelon et al. (2000) and is consistent with the higher level of protein. Similarly, the higher level of FAA in the FFFS cheese compared to the FFRS and FFHS cheeses concurs with the more rapid SLAB die-off in the former and confirms the results of studies showing higher starter cell autolysis as S/M is increased in the range 0.2 to 5.0% (Wilkinson et al. 1994). However, the absence of an effect of salt content on FAA in the RF and HF cheeses, despite the more rapid decrease in SLAB in the FS variants, may reflect a lower degree of autolysis of non-viable SLAB cells at the higher pH of these cheeses (RF, HF) compared to the FF cheese (Ramírez-Nuñez et al. 2011).
5 Conclusion
The combined effects of reducing salt and fat on the composition, microbiological and biochemical parameters of Cheddar was investigated in a matrix of nine cheeses, comprising of full-fat (33%, FF), reduced-fat (22%, RF) and half-fat (16%, HF) cheeses, each with full-salt (1.9%, FS), reduced-salt (1.2%, RS) and half-salt (0.9%, HS) variants. Salt and fat had additive effects on moisture, S/M, lactic acid and LA:P and interactive effects on pH, lactose and a w, the magnitude of which changed with ripening time. The current results highlight the dependence of cheese pH on the level of lactose and lactic acid (as influenced by moisture content), S/M which affects the rate of lactose utilization, and the fat level which affects the LA:P and therefore the ability of the calcium phosphate para-casein network to buffer the lactic acid. Hence, in the FF cheeses, residual lactose remaining in the FS variant after pressing was slowly metabolized to lactic acid over 270 days resulting in a slow decrease in pH. In contrast, most lactose was already converted to lactic during pressing in the RS and HS cheeses, and any remaining lactose (<0.14%) was rapidly utilized within 30 days, by which time the pH of these variants had already decreased to its lowest value. The pH changed during ripening to an extent influenced by fat and salt levels, with the pH decreasing in the FS variant of the FF and RF cheeses, remaining essentially constant in the FS variant of the HF cheese and in the RS and HS variants of the FF and RF cheeses, and increasing in the RS and HS variants of the HF cheeses. For each salt content, the mean pH over ripening increased as fat content was reduced, with the effect being more pronounced in the RS and HS cheeses. This effect coincided with the reduction in LA:P and increase in FAA. While salt and fat level did not affect growth of NSLAB or the initial count of SLAB, reducing salt content resulted in lower rates of SLAB die-off and development of FAA during ripening. In contrast, reducing fat content coincided with higher levels of protein and FAA.
The current results highlight the extent of change in key quality parameters such as MNFS, S/M, pH and the LA:P associated with varying degrees of fat and salt reduction (Lawrence et al. 2004). Consequently, they provide a valuable insight on what steps may be taken during manufacture to normalize the quality of reduced-fat, reduced-salt cheeses: e.g. reducing the buffering capacity through milk pre-acidification to reduce the pH of half-fat, half-salt cheese or altering the gel cutting and cooking programmes to reduce moisture in reduced-salt cheeses. Nevertheless, a more complete picture on the impact of the combined effects of fat and salt reduction is warranted prior to the development of appropriate production strategies for Cheddar cheeses with varying degrees of fat and salt reduction. This will be provided in a subsequent communication reporting on the effects of varying fat and salt in the current cheeses on quality aspects including sensory properties of the unheated and heated cheeses, free fatty acids, profiles of free amino acids and volatile compounds, rheology and cooking characteristics.
References
Arboatti AS, Olivares ML, Sabbag NG, Costa SC, Zorrilla SE, Sihufe GA (2014) The influence of sodium chloride reduction on physicochemical, biochemical, rheological and sensory characteristics of Mozzarella cheese. Dairy Sci Technol 94:373–386
Chou YE, Edwards CG, Luedecke LO, Bates MP, Clark S (2003) Nonstartr lactic acid bacteria and aging temperature affect calcium lactate crystallization in cheddar cheese. J Dairy Sci 86:2516–2524
Czarnacka-Szymani J, Jezewska-Zychowicz M (2015) Impact of nutritional information on consumers' acceptance of cheese with reduced sodium chloride content. Int Dairy J 40:47–53
FAO (Food and Agriculture Organization of the United Nations) (2014) FAO statistics division (FAOSTAT), World food and agriculture. http://www.fao.org/docrep/018/i3107e/i3107e.PDF
Fenelon MA, Guinee TP (2000) Primary proeolysis and textural changes during ripening in Cheddar cheeses manufactured to different fat contents. Int Dairy J 10:151–158
Fenelon MA, O'Connor P, Guinee TP (2000) The effect of fat content on the microbiology and proteolysis in cheddar cheese during ripening. J Dairy Sci 83:2173–2183
Flegal KM, Kit BK, Orpana H, Graubard BI (2013) Association of all-cause mortality with overweight and obesity using standard body mass index categories: a systematic review and meta-analysis. JAMA 309:71–82
Ganesan B, Brown K, Irish DA, Brohersen C, McMahon DJ (2014) Manufacture and sensory analysis of reduced- and low-sodium Cheddar and Mozzarella cheeses. J Dairy Sci 97:1970–1982
Guinee TP, Fox PF (2004) Salt in cheese: physical, chemical and biological aspects. Pages 207–259 in Cheese: chemistry, physics and microbiology. General Aspects. Vol. 1. 3rd ed. PF Fox, PLH McSweeney, TM Cogan, and TP Guinee, ed. Academic Press, London, UK
Guinee TP, Auty MAE, Fenelon MA (2000) The effect of fat content on the rheology, microstructure and heat-induced functional characteristics of Cheddar cheese. Int Dairy J 10:277–288
Guinee TP, O’Kennedy BT, Kelly PM (2006) Effect of milk protein standardization using different methods on the composition and yields of cheddar cheese. J Dairy Sci 89:468–482
He FJ, MacGregor GA (2009) A comprehensive review on salt and health and current experience of worldwide salt reduction programmes. J Hum Hypertens 23:363–384
Henney JE, Taylor CL, Boon CS (2010) Strategies to reduce sodium intake in the United States. The National Academies Press, Washington, DC
Hickey DK, Guinee TP, Hou J, Wilkinson MG (2013) Effects of variation in cheese composition and maturation on water activity in Cheddar cheese during ripening. Int Dairy J 30:53–58
Hou J, Hannon JA, McSweeney PLH, Beresford TP, Guinee TP (2012) Effect of curd washing on composition, lactose metabolism, pH, and the growth of non-starter lactic acid bacteria in full-fat Cheddar cheese. Int Dairy J 25:21–28
Hou J, Hannon JA, McSweeney PLH, Beresford TP, Guinee TP (2014) Effect of curd washing on cheese proteolysis, texture, volatile compounds, and sensory grading in full fat Cheddar cheese. Int Dairy J 34:190–198
Huffman LM, Kristoffersen T (1984) Role of lactose in Chedar cheese manufacture and ripening. NZ J Dairy Sci Technol 19:151–162
Irvine DM, Price WV (1961) Influence of salt on the development of acid by lactic acid starters in skim milk and curd submerged in brine. J Dairy Sci 44:243–248
Johnson ME, Kapoor R, McMahon DJ, McCoy DR, Narasimmon RG (2009) Reduction of sodium and fat levels in natural and processed cheeses: scientific and technological aspects. Compr Rev Food Sci Food Saf 8:252–268
Jordan KN, Cogan TM (1993) Identification and growth of non-starter lactic acid bacteria in Irish Cheddar cheese. Irish J Agric Food Res 32:47–55
Kalogeropoulos P, Georgiopoulou VV, Murphy RA, Newman AB, Bauer DC, Harris TB, Yang Z, Applegate WB, Kritchevsky SB (2015) Dietary sodium content, mortality, and risk for cardiovascular events in older adults: the health, aging, and body composition (Health ABC) study. JAMA Intern Med
Kelly M, Fox PF, McSweeney PLH (1996) Effect of salt-in-moisture on proteolysis in Cheddar-type cheese. Milchwissenschaft 51:498–501
Labrie SJ, Dupuis ME, Tremblay DM, Plante PL, Corbeil J, Moineau S (2014) A new Microviridae phage isolated from a failed biotechnological process driven by Escherichia coli. Appl Environ Microbiol 80:6992–7000
Lawrence RC, Gilles J, Creamer LK, Crow VL, Heap HA, Honoré CG, Johnston KA, Samal PK (2004) Cheddar cheese and related dry-salted cheese varieties. Pages 71–102 in Cheese: chemistry, physics and microbiology, Vol 2. 3rd ed. PF Fox, PLH McSweeney, TM Cogan, and TP Guinee, ed. Elesvier Academic Press, Amsterdam
Lu Y, McMahon DJ (2015) Effects of sodium chloride salting and substitution with potassium chloride on whey expulsion of Cheddar cheese. J Dairy Sci 98:78–88
Ma X, James B, Balaban MO, Zhang L, Emanuelsson-Patterson EAC (2013) Quantifying blistering and browning properties of Mozzarella cheese. Part II: cheese with different salt and moisture contents. Food Res Int 54:917–921
Marcos A, Alcala M, Leon F, Fernández-Salguero J, Esteban MA (1981) Water activity and chemical-composition of cheese. J Dairy Sci 64:622–626
McMahon DJ, Oberg CJ, Drake MA, Farkye N, Moyes LV, Arnold MR, Ganesan B, Steele J, Broadbent JR (2014) Effect of sodium, potassium, magnesium, and calcium salt cations on pH, proteolysis, organic acids, and microbial populations during storage of full-fat Cheddar cheese. J Dairy Sci 97:4780–4798
Møller KK (2012) Salt reduction in Cheddar cheese: exploring the potential of lactic acid bacteria and coagulants in restoring flavour and texture. PhD thesis, University of Copenhagen, Denmark
Møller KK, Rattray FP, Ardö Y (2013) Application of selected lactic acid bacteria and coagulant for improving the quality of low-salt Cheddar cheese: chemical, microbiological and rheological evaluation. Int Dairy J 33:163–174
Murtaza MA, Huma N, Sameen A, Murtaza MS, Mahmood S, Mueen-un-Din G, Meraj J (2014) Texture, flavor, and sensory quality of buffalo milk Cheddar cheese as influenced by reducing sodium salt content. J Dairy Sci 97:6700–6707
Pastorino AJ, Hansen CL, McMahon DJ (2003) Effect of salt on structure-function relationships of cheese. J Dairy Sci 86:60–69
Paulson BM, McMahon DJ, Oberg CJ (1998) Influence of sodium chloride on appearance, functionality and protein arrangements in nonfat Mozzarella cheese. J Dairy Sci 81:2053–2064
Ragosa M, Mitchell JA, Wiseman RT (1951) A selective medium for the isolation and determination of oral and faecal lactobacilli. J Bacteriol 62:132–133
Ramírez-Nuñez J, Romero-Medrano R, Nevárez-Moorillón GV, Gutiérrez-Méndez N (2011) Effect of pH and salt gradient on the autolysis of Lactococcus lactis strains. Braz J Microbiol 42:1495–1499
Rogers NR, Drake MA, Daubert CR, McMahon DJ, Bletsch TK, Foegeding EA (2009) The effect of aging on low-fat, reduced-fat, and full-fat Cheddar cheese texture. J Dairy Sci 92:4756–4772
Rudan MA, Barbano DM, Yun JJ, Kindstedt PS (1999) Effect of fat reduction on chemical composition, proteolysis, functionality and yield of Mozzarella cheese. J Dairy Sci 82:661–672
Rűegg VM, Blanc B (1977) Relationships between water activity, water sorption capacity and cheese composition. Milchwissenschaft 32:193–201
Rulikowska A (2009) Investigation of reduced salt content and the impact of starter related differences in salt sensitivity on Cheddar cheese quality. MSc thesis, University of Limerick, Limerick, Ireland
Rulikowska A, Kilcawley KN, Doolan IA, Alonso-Gomez M, Nongonierma AB, Hannon JA, Wilkinson MG (2013) The impact of reduced sodium chloride content on Cheddar cheese quality. Int Dairy J 28:45–55
Rynne NM, Beresford T, Kelly AL, Guinee TP (2007) Effect of milk pasteurisation temperature on age-related changes in lactose metabolism, pH and the growth of non-starter lactic acid bacteria in half-fat Cheddar cheese. Food Chem 100:375–382
Sánchez-Macías D, Fresno M, Moreno-Indias I, Castro N, Morales-delaNuez A, Álvarez S, Argüello A (2010) Physicochemical analysis of full-fat, reduced-fat, and low-fat artisan-style goat cheese. J Dairy Sci 93:3950–3956
SAS Institute (2011) SAS user's guide: statistics, 93rd edn. SAS Inst, Inc, Cary
Sgarbi E (2012) Non starter lactic acid bacteria during cheese ripening: survival, growth and production of molecules potentially involved in aroma formation. PhD. Dissertation, Universita Degli Studi di Parma.
Shakeel-Ur-Rehman, Waldron D, Fox PF (2004) Effect of modifying lactose concentration in cheese curd on proteolysis and in quality of Cheddar cheese. Int Dairy J 14:591–597
Skeie S, Alsethb GM, Østliea H, Abrahamsena RK, Johansena AG, Øyaas J (2013) Improvement of the quality of low-fat cheese using a two-step strategy. Int Dairy J 33:153–162
Swearingen PA, O'Sullivan DJ, Warthesen JJ (2001) Isolation, characterization and influence of native nonstarter lactic acid bacteria on Cheddar cheese quality. J Dairy Sci 84:50–59
Terzaghi BE, Sandine WE (1975) Improved medium for lactic streptococci and their bacteriophages. Appl Environ Microbial 29:807–813
Thibaudeau E, Roy D, St-Gelais D (2015) Production of brine-salted Mozzarella cheese with different ratios of NaCl/KCl. Int Dairy J 40:54–61
Thomas TD, Pearce KN (1981) Influence of sat on lactose fermentation and poteolysis in Cheddar cheese. New Zealand J Dairy Sci Technol 16:253–259
Tunick MH, Malin EL, Smith PW, Shieh JJ, Sullivan BC, Mackey KL, Holsinger VH (1993) Proteolysis and rheology of low-fat and full-fat Mozzarella cheeses prepared from homogenized milk. J Dairy Sci 76:3621–3628
Turner KW, Thomas TD (1980) Lactose fermentation in Cheddar cheese and the effect of salt. New Zealand J Dairy Sci Technol 15:265–276
UK Deparment of Health (2011) Assessment of dietary sodium levels among adults (aged 19–64) in England, 2011. Part 8. http://webarchive.nationalarchives.gov.uk/20130402145952/https://www.wp.dh.gov.uk/transparency/files/2012/06/Sodium-Survey-England-2011_Text_to-DH_FINAL1.pdf
Upreti P, Metzger LE (2006) Influence of calcium and phosphorus, lactose, and salt-to-moisture ratio on Cheddar cheese quality: manufacture and composition. J Dairy Sci 89:420–428
van Hekken DL, Tunick MH, Malin EL, Holsinger VH (2007) Rheology and melt characteristics of low-fat and full fat mozzarella cheese made from microfluidized milk. LWT Food Sci Technol 40:89–98
WHO (2013) New guidance on dietary salt and potassium. http://www.who.int/mediacentre/news/notes/2013/salt_potassium_20130131/en/#.UQqmEiMid2M.twitter
Wilkinson MG, Guinee TP, Fox PF (1994) Factors which may influence the determination of autolysis of starter bacteria during cheddar cheese ripening. Int Dairy J 4:141–160
Acknowledgments
This work was funded by the Department of Agriculture, Fisheries and Food, under the Food Institutional Research Measure with project reference no. 2012219.
Author information
Authors and Affiliations
Corresponding author
Additional information
This paper is part of the Special issue dedicated to the 9th International Cheese Symposium held in Cork, Ireland and organized by Teagasc in collaboration with University College Cork and INRA, 12th & 13th November 2014.
About this article

Cite this article
McCarthy, C.M., Wilkinson, M.G., Kelly, P.M. et al. Effect of salt and fat reduction on the composition, lactose metabolism, water activity and microbiology of Cheddar cheese. Dairy Sci. & Technol. 95, 587–611 (2015). https://doi.org/10.1007/s13594-015-0245-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13594-015-0245-2