
Abstract
Dengue fever is one of the most important vector-borne diseases. It is transmitted by Aedes Stegomyia aegypti, and one of the most effective strategies to combat the disease is the reduction of exposure to bites of these mosquitoes. In this paper, we present a game-theoretical model in which individuals choose their own level of protection against mosquito bites in order to maximize their own benefits, effectively balancing the cost of protection and the risk of contracting the dengue fever. We find that even when the usage of protection is strictly voluntary, as soon as the cost of protection is about 10,000 times less than the cost of contracting dengue fever, the optimal level of protection will be within 5 % of the level needed for herd immunity.
Similar content being viewed by others

1 Introduction
Dengue virus is a leading cause of illness and death in the tropics and subtropics. The virus causes dengue fever, a vector-borne disease transmitted by mosquitoes. Each year, an estimated 50–500 million cases of dengue fever occur worldwide (Whitehorn and Farrar 2010; Bhatt et al. 2013). The principal vector, Aedes aegypti, is a domestic species highly susceptible to the dengue virus. It feeds on human blood usually during the daytime and takes multiple blood meals during one feeding cycle. It breeds in stagnant clean water in artificial containers and is, therefore, well adapted to urban life. A second vector, Aedes Stegomyia albopictus, is generally considered less effective as an epidemic vector because, unlike A. aegypti, it feeds on many animals other than humans and is less strongly associated with the domestic environment (Whitehorn and Farrar 2010). There are no approved vaccines for the dengue virus, and prevention depends on mosquito control and protection from mosquito bites (WHO 2009) although other methods have recently been proposed (Bian et al. 2010; Frentiu et al. 2010).
Game-theoretical models (see, for example, Bauch and Earn 2004) consider a situation where individuals face a decision either to do a potentially costly preventive action (such as to use repellents) or to risk that they contract the disease. Game theory has been applied to vaccination against major public health threats, including smallpox (Bauch et al. 2003), measles (Shim et al. 2012b), rubella (Shim et al. 2009), childhood diseases (Bauch 2005), influenza (Galvani et al. 2007) and many others. From the game-theoretical perspective, an individual takes an action (such as to use a repellent or to vaccinate the child) that will maximize their personal payoff, taking into account the disease incidence and risk of infection, which is determined by actions taken by the rest of the population (Shim et al. 2012a). The theory assumes that individuals are driven by self-interest rather than by the interest of the group (Shim et al. 2011). As the frequency of preventive actions increases, the incentive to make such an action decreases due to indirect protection by other protected individuals (Bauch and Earn 2004). This makes the eradication of disease very difficult, even when the cost of preventive action is very low (Geoffard and Philipson 1997).
In this paper, we follow the general approach of previous models such as Bauch and Earn (2004) and apply the approach to a situation where individuals choose to use repellent to reduce the mosquito biting rate and consequently to reduce the risk of contracting dengue fever.
2 Methods
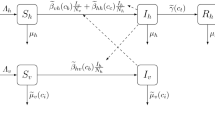
Our model relies heavily on the results and analysis of Amaku et al. (2014). In Amaku et al. (2014), the authors present a model involving all variables related to vector control of dengue fever. The model also considers the vertical transmission of dengue in the mosquitoes (Adams and Boots 2010). In Amaku et al. (2014), the authors deduce thresholds for the elimination of the disease. In particular, they derive a basic reproduction number for dengue fever
The notation is explained, and the values of the parameters are given in Table 1. We note that this expression holds for biologically reasonable values and may break down for values far away from those presented in Table 1. For example, if \(g\approx 1\), \(R_0\) would certainly not increase to \(\infty \) as (1) suggests.
For our purposes, the most important parameter is the average biting rate, a, as this is what every single individual can influence by their actions (such as using repellents and mosquito shields) and also because \(R_0\) is very sensitive to a.
Individuals can influence the rate mosquitoes bite them by the use of repellents. For the sake of simplicity, we assume that when \(r\in [0,1]\) denotes the frequency of repellent usage, the biting rate is expressed by
where \(a_{\mathrm{max}}\) is the maximum average biting rate when no protection is in place.
Based on Amaku et al. (2014), the probability an individual gets infected is given by \(\frac{\lambda }{\lambda +\mu _H}\), where \(\lambda = \lambda (r,r_{\mathrm{pop}})\) is the force of infection that depends on the individual’s repellent usage, r, as well as on the repellent usage in the population, \(r_{\mathrm{pop}}\), and can be expressed by
whenever \(R_0>1\). Note that \(\lambda = 0\) when \(R_0\le 1\). Also, note that \(\lambda (r,r_{\mathrm{pop}})\) seems to decrease in g but in fact does not as its component \(R_0\) increases in g.
Following Bauch and Earn (2004), the individual’s payoff of using r in the population using \(r_{\mathrm{pop}}\) is given by
where C is the cost of complete protection (\(r=1\)) relative to the cost of dengue fever to the individual.
For every given \(r_{\mathrm{pop}}\), we can find \(r_{\mathrm{max}}= r_{\mathrm{max}}(r_{\mathrm{pop}})\in [0,1]\) that maximizes \(E(\cdot , r_{\mathrm{pop}})\). We are looking for the Nash equilibrium value, \(r_{\mathrm{NE}}\), such that when all individuals use \(r_{\mathrm{NE}}\), nobody has an incentive to deviate from it.
3 Results
Let \(R_{0,\mathrm{max}}\), given by (1) when \(a=a_{\mathrm{max}}\), be the maximal basic reproduction number for dengue fever when nobody uses any protection (i.e., when \(r_{\mathrm{pop}}=0\)). It follows from (2) to (1) that \(R_0<1\) if and only if \(r_{\mathrm{pop}}>r_{\mathrm{HI}}\) where
corresponds to the level of protection that yields herd immunity against dengue fever. Consequently, if \(r_{\mathrm{pop}}>r_{\mathrm{HI}}\), there is little risk of contracting dengue fever, and thus, \(r_{\mathrm{max}}(r_{\mathrm{pop}})=0\). For the rest of this section, we will thus assume \(r_{\mathrm{pop}}<r_{\mathrm{HI}}\) and thus \(R_0>1\). Note that
and thus
Consequently, the maximum of \(E(\cdot , r_{\mathrm{pop}})\) is attained either at 0 or 1, and the equilibrium value \(r_{\mathrm{NE}}\) will be given by the solution of
which is equivalent to
Figure 1 shows the equilibrium values \(r_{\mathrm{NE}}\) as they depend on the relative cost of protection, C for various relative mosquito densities m. Figure 2 further shows that \(r_{\mathrm{HI}}-r_{\mathrm{NE}}\) depends on C in an almost linear way and in particular, when \(C\approx 10^{-4}\), the optimal value of protection is within 5 % of the protection level needed for herd immunity (and decreasing the cost further does not significantly improve the outcomes). Figure 3 further shows that the disease is very close to being eliminated for such low C.

Dependence of optimal protection level \(r_{\mathrm{NE}}\) on the relative cost of the protection from various mosquito prevalences. Note that \(r_{\mathrm{NE}}\) increases with m

Dependence of the difference between \(r_{\mathrm{HI}}\), the protection yielding the herd immunity for dengue fever, and the optimal protection level \(r_{\mathrm{NE}}\) on the relative cost of the protection from various mosquito prevalences. Note that the difference decreases with m

Dependence of the basic reproduction number \(R_0\) when everybody uses \(r_{\mathrm{NE}}\) on the relative cost of the protection when \(m=1\)
4 Discussion
Following Bauch and Earn (2004), we have provided a game-theoretical model to study the optimal usage of repellents in order to prevent the contraction of dengue fever. Our analysis is built on the model of dengue infection dynamics by Amaku et al. (2014) that, similarly to Coutinhoa et al. (2006), Burattini et al. (2008) and Massad et al. (2011) studied the efficiency of control strategies. Not surprisingly, as in Geoffard and Philipson (1997), Bauch and Earn (2004) and Bauch (2005), we found that eradication of dengue fever by voluntary usage of repellents to minimize mosquito biting rate is not possible. Nevertheless, we found that the optimal repellent usage is still very close to the levels needed for herd immunity, especially if the cost of repellents is low.
In our model, we assumed that individuals can achieve perfect protection. This is not always possible, and as shown, for example, in Wu et al. (2011), imperfect protection aggravates the dilemma of voluntary protective actions. We have also assumed that individuals have perfect information (about the dengue epidemics, the protection coverage in the population, etc.), base their decision solely on the expected payoff, and all (but a focal individual) use the same level \(r_{\mathrm{pop}}\). However, individuals can have different perceptions of the risk (Poletti et al. 2011) and can also base their decision based also on different social aspects (Xia and Liu 2013). Consequently, there is an inherent variability in the population protection level that is not captured in our model.
Nevertheless, game-theoretical models can still capture the interplay between disease prevalence, protective action coverage and individual behavior, and may help in managing the population dynamics of vaccinating behavior for the benefit of public health (Bauch 2005).
References
Adams B, Boots M (2010) How important is vertical transmission in mosquitoes for the persistence of dengue? Insights from a mathematical model. Epidemics 2(1):1–10
Amaku M, Coutinho FAB, Raimundo SM, Lopez LF, Burattini MN, Massad E (2014) A comparative analysis of the relative efficacy of vector-control strategies against dengue fever. Bull Math Biol 76(3):697–717
Bauch CT (2005) Imitation dynamics predict vaccinating behaviour. Proc R Soc B Biol Sci 272(1573):1669–1675
Bauch CT, Earn DJ (2004) Vaccination and the theory of games. Proc Natl Acad Sci USA 101(36):13391–13394
Bauch CT, Galvani AP, Earn DJ (2003) Group interest versus self-interest in smallpox vaccination policy. Proc Nat Acad Sci 100(18):10564–10567
Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, Drake JM, Brownstein JS, Hoen AG, Sankoh O et al (2013) The global distribution and burden of dengue. Nature 496(7446):504–507
Bian G, Xu Y, Lu P, Xie Y, Xi Z (2010) The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. PLoS Pathog 6(4):e1000833
Brownstein J, Hett E, O’Neill S (2003) The potential of virulent wolbachia to modulate disease transmission by insects. J Invertebr Pathol 84(1):24–29
Burattini M, Chen M, Chow A, Coutinho F, Goh K, Lopez L, Ma S, Massad E (2008) Modelling the control strategies against dengue in Singapore. Epidemiol Infect 136(03):309–319
Coutinhoa F, Burattinia M, Lopeza L, Massada E (2006) Threshold conditions for a non-autonomous epidemic system describing the population dynamics of dengue. Bull Math Biol 68(8):2263–2282
Frentiu FD, Robinson J, Young PR, McGraw EA, O’Neill OL (2010) Wolbachia-mediated resistance to dengue virus infection and death at the cellular level. PloS One 5(10):e13398
Galvani AP, Reluga TC, Chapman GB (2007) Long-standing influenza vaccination policy is in accord with individual self-interest but not with the utilitarian optimum. Proc Nat Acad Sci 104(13):5692–5697
Geoffard PY, Philipson T (1997) Disease eradication: private versus public vaccination. Am Econ Rev 87(1):222–230
Halstead SB (1990) Dengue and dengue hemorrhagic fever. Curr Opin Infect Dis 3(3):434–438
Index Mundi (2012) Life expectancy at birth. http://www.indexmundi.com/map/?v=30
Massad E, Coutinho FAB, Lopez LF, Da Silva DR (2011) Modeling the impact of global warming on vector-borne infections. Phys Life Rev 8(2):169–199
Ocampo CB, Wesson DM (2004) Population dynamics of Aedes aegypti from a dengue hyperendemic urban setting in Colombia. Am J Trop Med Hyg 71(4):506–513
Poletti P, Ajelli M, Merler S (2011) The effect of risk perception on the 2009 H1N1 pandemic influenza dynamics. PloS One 6(2):e16460
Scott TW, Morrison AC, Lorenz LH, Clark GG, Strickman D, Kittayapong P, Zhou H, Edman JD (2000) Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: population dynamics. J Med Entomol 37(1):77–88
Shim E, Kochin B, Galvani A (2009) Insights from epidemiological game theory into gender-specific vaccination against rubella. Math Biosci Eng 6(4):839–854
Shim E, Meyers LA, Galvani AP (2011) Optimal H1N1 vaccination strategies based on self-interest versus group interest. BMC Public Health 11(Suppl 1):S4
Shim E, Chapman GB, Townsend JP, Galvani AP (2012a) The influence of altruism on influenza vaccination decisions. J R Soc Interf 9:2234–2243
Shim E, Grefenstette JJ, Albert SM, Cakouros BE, Burke DS (2012b) A game dynamic model for vaccine skeptics and vaccine believers: measles as an example. J Theor Biol 295:194–203
Whitehorn J, Farrar J (2010) Dengue. Br Med Bull 95(1):161–173
WHO (2009) World Health Organization and Special Programme for Research and Training in Tropical Diseases and World Health Organization. Department of Control of Neglected Tropical Diseases and World Health Organization. Epidemic and Pandemic Alert. Dengue: guidelines for diagnosis, treatment, prevention and control. World Health Organization
Wu B, Fu F, Wang L (2011) Imperfect vaccine aggravates the long-standing dilemma of voluntary vaccination. PloS One 6(6):e20,577
Xia S, Liu J (2013) A computational approach to characterizing the impact of social influence on individuals’ vaccination decision making. PloS One 8(4):e60,373
Acknowledgments
This research was supported by the MAA’s National Research Experience for Undergraduates program funded by NSF Grants DMS-1156582 and DMS-1359016. The authors also would like to thank the anonymous reviewers for their comments that helped to improve the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Dorsett, C., Oh, H., Paulemond, M.L. et al. Optimal Repellent Usage to Combat Dengue Fever. Bull Math Biol 78, 916–922 (2016). https://doi.org/10.1007/s11538-016-0167-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11538-016-0167-z