
Abstract
We propose a novel ultrasound approach with the primary aim of establishing the temporal relationship of structure and function in athletes of varying sporting demographics. 92 male athletes were studied [Group IA, (low static–low dynamic) (n = 20); Group IC, (low static–high dynamic) (n = 25); Group IIIA, (high static–low dynamic) (n = 21); Group IIIC, (high static–high dynamic) (n = 26)]. Conventional echocardiography of both the left ventricles (LV) and right ventricles (RV) was undertaken. An assessment of simultaneous longitudinal strain and LV volume/RV area was provided. Data was presented as derived strain for % end diastolic volume/area. Athletes in group IC and IIIC had larger LV end diastolic volumes compared to athletes in groups IA and IIIA (50 ± 6 and 54 ± 8 ml/(m2)1.5 versus 42 ± 7 and 43 ± 2 ml/(m2)1.5 respectively). Group IIIC also had significantly larger mean wall thickness (MWT) compared to all groups. Athletes from group IIIC required greater longitudinal strain for any given % volume which correlated to MWT (r = 0.4, p < 0.0001). Findings were similar in the RV with the exception that group IIIC athletes required lower strain for any given % area. There are physiological differences between athletes with the largest LV and RV in athletes from group IIIC. These athletes also have greater resting longitudinal contribution to volume change in the LV which, in part, is related to an increased wall thickness. A lower longitudinal contribution to area change in the RV is also apparent in these athletes.
Similar content being viewed by others


Introduction
The assessment of the ‘athletes heart’ (AH) has received significant attention with literature highlighting structural adaptation of both the left ventricles (LV) [1] and right ventricles (RV) [2]. Structural adaptation may mimic that of inherited cardiac disease such as hypertrophic cardiomyopathy and arrhythmogenic right ventricular cardiomyopathy and therefore a grey zone of differential diagnosis is often apparent [3, 4]. In order to aid differential diagnosis some studies have assessed cardiac function using conventional echocardiography [5]. More recently others [6–8] have utilised novel techniques such as strain (ε) imaging. Absolute longitudinal ε values presented in these and other studies have been variable further complicating the differential diagnosis of physiological from pathological adaptation [9–11]. These studies rarely take into account the different chamber size or temporal changes in functional data from athletes of varying workloads/sporting disciplines which may, in part, explain any inconsistencies that have been observed.
We have been developing a combination of 2D and ε imaging methods to provide simultaneous temporal relationships of LV and RV structure and longitudinal function. This technique elucidates, non-invasively, the structure-function relationship throughout systole and diastole and provides estimates of the relative contribution of longitudinal mechanics to volume/area change. It is known that RV structure, anatomy, mechanics and function are very different to the LV. In fact, unlike the LV, RV filling in healthy subjects is predominantly determined by kinetic energy generated through gravity and respiration [12]. It is, therefore, likely that ε-area/volume relationships may differ in the RV and LV in a range of highly trained athletes which may well highlight the primary physiological differences during the filling/diastolic phase of the cardiac cycle.
In view of this, the aim of our study was to characterise LV and RV longitudinal ε-volume/area relationships throughout the cardiac cycle, utilising a novel post-processing technique, in athletes with high and low volumes of static and dynamic exercise training. We hypothesise that:
-
1.
When calculated to end diastolic chamber size RV and LV ε will not differ across athlete groups.
-
2.
The longitudinal contribution to volume/area change will be similar in systole compared to diastole within the LV but different in the RV.
Materials and methods
Study design and population
Ninety-two Caucasian, male athletes were recruited into this cross-sectional study and sub-grouped into ‘Mitchells Classification’ [13] based upon their sporting discipline. This allowed for four sub-groups: Group IA, low static–low dynamic (<20 % maximal voluntary contraction (MVC) [defined as the greatest amount of tension the relevant muscle (groups) can generate and hold] and <40 % maximal oxygen uptake (maxO2)). Group IC, high static–low dynamic (>50 % MVC and <40 % maxO2). Group IIIA, low static–high dynamic (<20 % MVC and >70 % maxO2) and group IIIC, high static–high dynamic (>40 % MVC and >70 % maxO2). Group IA consisted of cricketers (n = 20, mean age ± SD, 28 ± 4 years), group IIIA were weightlifters, aikido athletes and gymnasts (n = 21, mean age ± SD, 27 ± 10 years), group IC consisted of footballers (n = 25, mean age ± SD, 25 ± 4 years) and group IIIC comprised of cyclists and boxers (n = 26, mean age ± SD, 26 ± 6 years). All athletes were classified as elite and performed at national or international level. Training status was high and involved a combination of static, dynamic and sporting practice/competition relative to their sporting discipline. Mean weekly training hours were 27, 18, 25 and 29 h for groups IA, IIIA, IC and IIIC respectively. All participants were healthy and free from cardiovascular disease and avoided alcohol and caffeine 24 h prior to data collection and refrained from training for at least 6 h prior to the examinations. Ethics approval was granted by the ethics committee of Liverpool John Moores University.
Procedures
Body mass (Seca 217, Hannover, Germany) and height (Seca Supra 719, Hannover, Germany) were recorded and body surface area (BSA) was calculated as previously described [14]. All athletes completed a health questionnaire to exclude cardiovascular symptoms, family history of sudden cardiac death and any other cardiovascular history and/or abnormalities. A screening 12-lead electrocardiogram (CardioExpress SL6, Spacelab Healthcare, Washington, US) confirmed the absence of non-training related abnormalities [15]. A full standard echocardiogram was undertaken by a single experienced sonographer using a commercially available ultrasound system (Vivid Q, GE Healthcare, Horten, Norway) and a 1.5–4 MHz phased array transducer. All images were acquired in accordance with British Society of Echocardiography guidelines [16]. Images were stored in a raw DICOM format and exported to an offline workstation (EchoPac version 6.0, GE Healthcare, Horten, Norway) for subsequent analysis. All data was analysed by a single experienced sonographer overseeing three student sonographers.
Conventional 2D echocardiography
Standard measurements of the LV were made in accordance with American Society of Echocardiography (ASE) guidelines [17]. LV linear dimensions (LVDd) were measured from a parasternal long axis orientation and LV mass was calculated using the ASE corrected equation. In order to provide a comprehensive assessment of LV wall thickness, eight measurements were made from a parasternal short axis orientation at basal and mid-levels from the antero-septum, infero-septum, posterior wall and lateral wall [18]. Mean wall thickness (MWT) was calculated as an average of all eight segments and the maximum value of wall thickness (MaxWT) was also reported. LV end diastolic volume (LVEDV), LV end systolic volume (LVESV) and ejection fraction (EF) were calculated from a Simpson’s biplane method utilising both apical four and two chamber orientations. An indication of LV geometry and relative wall thickness was determined by dividing LVEDV into LV mass (LVmass/LVEDV).
Standard 2-dimensional measurements of the RV were also made in accordance with ASE guidelines [19]. The RV outflow tract (RVOT) was measured at three locations, from the parasternal long axis (RVOTplax) and proximal (RVOT1) and distal (RVOT2) from a parasternal short axis orientation. The RV inflow was measured from a modified apical four chamber orientation and included the base (RVD1) the mid-level (RVD2) and the length (RVD3). RV diastolic area (RVDa) and RV systolic area (RVSa) were measured from the same acoustic window and RV fractional area change was calculated (RVFAC). RV wall thickness was measured from a sub-costal approach.
For direct comparison between groups, all structural indices were scaled allometrically to BSA based on the principle of geometrical similarity [20, 21]. Hence, linear dimensions were scaled to BSA0.5, areas directly to BSA and volumes to BSA1.5.
Myocardial speckle tracking
Images for offline assessment of myocardial ε and volume/area were acquired from the standard examination using a focused apical four-chamber view for the LV and a modified apical four-chamber view for the RV. In both views frame rates were adjusted to between 40 and 90 frames per second (FPS).
During the offline analysis (EchoPac, Version 6.0, GE Healthcare, Horten, Norway) a region of interest was placed around the LV from basal septum through to the basal lateral wall ensuring the whole of the myocardium was encompassed within. This provided six myocardial segments and an average of these provided a global index of LV longitudinal ε. For the RV the region of interest was constrained to the lateral wall only providing three segments from base to apex and the average was used to determine global longitudinal RV ε.
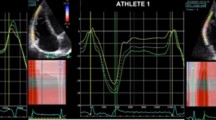
Novel assessment of strain-volume/area relationships were calculated for each participant (see Fig. 1). The raw ε data was exported to a spreadsheet (Excel, Microsoft Corp, Washington, US) and the global temporal values underwent cubic spline interpolation to provide 300 points in systole and 300 points in diastole in order to correct for variable heart rates [22]. The 600 ε values were subsequently split into 5 % increments of the cardiac cycle ensuring the raw peak value was included. The absolute time points for each of the ε values were noted and the same image and cardiac cycle were used to trace LV and RV monoplane volume and area respectively providing simultaneous measurements of volume or area and ε. A ε-volume/area loop was created for each participant and a polynomial regression of two orders was applied to both the systolic and diastolic components. These equations were then used to calculate ε at % increments of EDV and EDA. The difference between systolic and diastolic ε at each % increment of EDV and EDA was termed ‘systolic–diastolic coupling’ and reflects the longitudinal contribution to volume or area change between systole and diastole.

Methods for generation of strain-volume/area loops
Statistical analysis
All data is expressed as mean and standard deviation. A one-way sample ANOVA was applied to all variables across all four athlete groups. A p value of <0.05 was considered statistically significant. Where derived ε values indexed to volume or area and/or the systolic–diastolic coupling was significantly different between any groups a standard Pearson’s correlation was used to establish any relationship between absolute chamber size and these novel indices. Twenty randomly selected athletes were re-analysed by two separate operators to establish inter-observer variability of both right and left ventricular loops across each time-point and following calculation of strain and systolic–diastolic coupling for given EDV and EDA. The full data is available in a supplementary file.
Results
Baseline demographics for all athlete groups are presented in Table 1. All participants were matched for age. Training hours were similar between groups with exception of IIIA who trained for significantly fewer hours per week compared to all groups. Group IIIC had significantly lower body mass and BSA then all the other three groups. Heart rate was significantly lower in group IIIC compared to groups IA and IIIA and lower in group IC compared to group IIIA.
The left ventricle
Structural and functional indices of the LV are presented in Table 2. Indexed values of LV cavity size were elevated in those athletes with a high dynamic component. Other parameters of LV geometry were significantly higher in the IIIC group for LVmass index (compared to groups IA and IIIA), MWT index (compared to group IA) and MaxWT index (compared to groups IA and IC). There was no difference in EF or LVmass/LVEDV between any of the groups.
Peak LV ε was significantly lower in groups IC and IIIA compared to groups IA and IIIC. Graphical representation of LV ε and simultaneous volume is demonstrated in Fig. 2. Values for ε are different between groups but when indexed to the initial EDV the pattern of volume change is similar across groups (Fig. 3) and is also reflected in the ε-volume loops (Fig. 4). Polynomial regression revealed no difference in ε at any % EDV between groups IA, IC and IIIA, in either systole or diastole (see Table 3). The same polynomial regression from the ε-volume loops demonstrated a significantly higher longitudinal ε throughout both systole and diastole in group IIIC (see Fig. 3; Table 3). Linear systolic–diastolic coupling was evident in all groups consistent with similar changes in ε as volume altered throughout the cardiac cycle.
There was a weak to moderate but significant positive correlation between increased systolic ε in the physiological range [70–30 % of LVEDV (r = 0.246–0.406, p < 0.0001)] and increased MWT in group IIIC only. This finding was also evident with regards to diastolic ε at 40 and 30 % of LVEDV (r = 0.365 and 0.423, p < 0.0001).

Simultaneous left ventricular longitudinal ε and volume from all athlete groups

Predicted strains for given % LVEDV working in the physiological range of a 70 % ejection fraction based on polynomial regression of individual deformation-volume loops from all athlete groups

Left ventricular ε-volume loops for all athlete groups
The right ventricle
All structural and global functional indices from the RV are presented in Table 4. Group IIIC had significantly larger RV outflow dimensions compared to groups with low dynamic activity. There was no difference in outflow size between group IC and IIIC. Both RVD1 and RVD3 were also significantly larger in group IIIC compared to all the other groups. There was no difference in RV wall thickness between any of the groups. RVFAC was significantly higher in group IIIC compared to IC.
There was no significant difference in peak longitudinal ε between all groups. Figure 5 demonstrates simultaneous RV ε and area change across the cardiac cycle in all groups. Although ε appears homogenous across the cardiac cycle this is at different initial starting volumes. Polynomial regression provided similar ε throughout systole and diastole for any given % EDA between groups IA, IC and IIIA (see Table 5). A trend for lower RV ε in group IIIC was noted throughout the cardiac cycle which was significant between 50 and 70 % EDA (see Fig. 6). RV ε-area loops highlight variation between systole and diastole at any given volume (see Fig. 7). There were no significant correlations between absolute RV wall thickness and longitudinal ε in all groups.

Simultaneous right ventricular longitudinal ε and area from all athlete groups

Predicted strains for given % RVEDA working in the physiological range of a 50 % fractional area change based on polynomial regression of individual deformation-volume loops from all athlete groups

Right ventricular ε-area loops for all athlete groups
Discussion
Using traditional measures of cardiac function and structure, we confirm previous observation that physiological adaptation is primarily driven by high dynamic exercise. The novel deformation-area relationship, however, reveals potential differences between different elite athletes. The main novel findings from this study are: (1) different LV and RV peak longitudinal ε in groups IA, IC and IIIA were normalised for % EDV and EDA, (2) athletes in group IIIC require a greater longitudinal contribution to volume change in the LV which in part is related to an increased wall thickness, (3) athletes in group IIIC require a lower longitudinal contribution to area change in the RV when compared to other athlete groups and (4) longitudinal systolic–diastolic coupling is observed in the LV but reduced in the RV in all groups.
Left ventricular structural and functional adaptation
Those athletes involved in high static but low dynamic activity demonstrate a lack of cardiac adaptation with no evidence to highlight an increased wall thickness. Early work proposed a dichotomous type of adaptation of concentric hypertrophy in resistance athletes and eccentric hypertrophy in endurance athletes [23]. This hypothesis has received recent scrutiny with evidence to refute the resistance ‘limb’ of the dichotomy [6]. This phenomenon is further complicated by the multi-training nature of different sporting disciplines. The current study provides a unique assessment of cardiac adaptation in athletes from the “four corners” of Mitchell’s classification hence reflecting the variable training nature of elite sport. Our data demonstrate that those athletes involved only in dynamic activity present with LV chamber enlargement. In addition, athletes with high static and high dynamic components demonstrate a greater wall thickness, consistent with the partial development of an eccentric type of hypertrophy. Based on this it is apparent that a continuum of cardiac adaptation exists that is primarily driven by dynamic activity and further enhanced with co-existing static exercise. In view of this the presence of cardiac enlargement in low dynamic sports should be interpreted with caution.
Global LV function, as determined by EF, was not different between athlete groups however peak longitudinal ε was. Following ε-volume assessment all groups, with the exception of IIIC, presented with similar peak longitudinal ε. This confirms that differences in peak ε between these groups is not an inherent functional difference but merely a consequence of differences in cavity size. Based on a reported correlation between ε and wall thickness we can speculate that the greater wall thicknesses seen in IIIC athletes in combination with the increased cavity size causes structural refinement of the longitudinal matrix [24] and enhances its contribution to ejection. It may also be possible that ‘over-perfusion’ from enhanced resting coronary artery flow seen in endurance athletes [25] allows greater perfusion of the endocardium. It is difficult to articulate the physiological benefit for this shift in mechanics but may provide a greater reserve in circumferential/radial mechanics [11] to contribute more efficiently during exercise.
Right ventricular structure and function
We also observed larger RV dimensions in group IIIC only, suggesting the combination of high dynamic and high static training is again the primary driver for adaptation. This effects the RV inflow to a greater extent and is in fitting with previous work in the endurance population [2, 26]. This suggests that the grey area for differentiating from right-sided pathology is smaller in those athletes that do not integrate a combination of higher levels of dynamic and static exercise into their training/sport.
Absolute peak RV ε was not different between groups. Following generation of ε-area loops, group IIIC only (those with the greatest chamber adaptation) had a lower longitudinal ε for any given % change in EDA, which is at odds with the data reported for the LV. The physiological differences observed between ventricles at rest is likely reflective of different longitudinal and circumferential/radial reserve in the RV and LV respectively that underpin dichotomous mechanics in the RV and LV in order to generate higher stroke volumes during exercise. Previous work has highlighted lower RV regional ε in athletes with dilated ventricles [27] and our indexed data is in support of this. The lack of correlations of ε at specific % EDA with RV dimensions in the IIIC group may be a consequence of the complex shape of the RV with the recommended linear dimensions not fully representing its unique geometry [28]. It is also important to note that we chose to assess the RV lateral wall in isolation in order to conform with other studies that have assessed RV ε in the athletes heart. Due to the known homogenous distribution of ε in the RV myocardium in a physiological model, it is unlikely that the inclusion of the septum would provide different findings. That aside, further work aiming to better reflect RV geometry and wall thickness may elucidate potential structural and functional links.
Systolic–diastolic coupling
The ε-volume/area loops for the LV and RV provide insight into the relative contribution of longitudinal ε to structural change throughout the cardiac cycle. As predicted the shapes of the loops were similar between groups but different between the RV and LV. LV filling requires the rapid relaxation of the myocardium to generate the low LV pressure and the subsequent LA–LV pressure gradient [29]. This is maintained throughout early and late diastole by a combination of active relaxation and compliance. Ultimately this interaction throughout diastole allows for the generation of vortices which in turn provide the ‘suction’ of blood through the left side of the heart. The loops highlight the importance of longitudinal relaxation to this process with evidence of similar ε at any given volume during both systole and diastole. RV filling is very different to that of the LV. In healthy subjects, kinetic energy generated by gravity and respiration contribute significantly to filling [12]. This only occurs with normal right atrial pressures and excellent compliance of the RV and pulmonary vasculature. In our healthy athlete population we demonstrate the presence of systolic–diastolic “uncoupling” as determined by an elevation of longitudinal ε in diastole compared to the systolic counterpart at any given % EDA. It is difficult to fully explain this finding but it may well reflect the significant influence of variable loading conditions on RV physiology.
It would be valuable to establish LV and RV longitudinal systolic–diastolic coupling in different pathologies such as hypertrophic cardiomyopathy and arrhythmogenic right ventricular cardiomyopathy where differentiation from the AH is challenging. It is apparent that different training loads do not impact upon this specific functional index and therefore future work should aim to provide data from these pathological populations.
Limitations
The aim of the ε-volume/area loops was to provide simultaneous longitudinal ε and structure. This is only feasible using a single 2D image and hence we constrained its application to the assessment of longitudinal function only. The application to circumferential, radial and twist is feasible but this would not provide a direct ‘same cardiac cycle comparison’ and hence was not used in this analysis. Furthermore the short axis assessment suffers significantly from out-of-plane motion [30] which would further compound the problem. Three-dimensional echocardiography would overcome this and can provide simultaneous volume and strain in all planes. However a ‘real-time’ 3D acquisition provides very low frame rates (approximately 5–10 fps gives a temporal resolution of 100–200 ms) which would under-sample important components of the cardiac cycle. High frame rate 3D imaging is in development and when this comes to fruition it will be sensible to apply this technique.
The current methods for undertaking this type of analysis are time-consuming taking up to 30 min per subject to acquire both LV and RV loops. We are hopeful that with automated tracking, industrial partners will allow access to temporal volume data as well as temporal strain data which would make this type of analysis much more efficient and feasible in the ‘real-world’ clinical setting. The concept of area-strain assessment that is available on some ultrasound vendors may provide an alternative to the methodology used in the current study and equally overcome some of the issues related to analysis time. That aside, the modelling of the data to express ε at % EDV and EDA will require additional post processing. Likewise, the assessment of the RV would be problematic using this approach.
In order to move away from conventional classifications of endurance and resistance based athletes, we utilised Mitchells classification of sport which is based on relative dynamic and static workloads. This means that athletes from very different sports are often placed within the same classification. The allocation of each sport to a specific classification was based on consensus opinion by a Task Force working group. It is therefore pertinent to offer some critical perspective of this classification with the potential for bias and a potential lack of scientific rigor related to the true haemodynamic demands of individual sporting disciplines. Although it challenges preconceived views of sport we believe it is currently the ‘best’ workload classification available and hence its inclusion in this study. In addition, we based specific training loads on this classification system which provides general data on sporting disciplines rather than quantify overall training load on any individual/cohort. Although we feel this is an improvement on previous work it is apparent that future work would benefit from providing data pertaining to individual static and dynamic workloads.
Conclusion
The largest LV and RV morphology occurs in athletes that are engaged in a combination of high dynamic and high static exercise (group IIIC). This athlete group also have greater resting longitudinal contribution to volume change in the LV when compared to all other athlete groups which, in part, is related to an increased wall thickness. A lower longitudinal contribution to area change in the RV is also apparent for these athletes. The variable peak longitudinal ε seen in other athlete groups is merely a reflection of cardiac chamber size and does not indicate any intrinsic differences in function. Finally novel ε-volume/area loops highlight significant differences in longitudinal contribution to diastolic filling between the left and right ventricles in all athletes.
References
Pelliccia A, Culasso F, Di Paolo FM, Maron BJ (1999) Physiologic left ventricular cavity dilatation in elite athletes. Ann Intern Med 130:23–31
Oxborough D, Sharma S, Shave R, Whyte G, Birch K, Artis N, Batterham A, George K (2012) The right ventricle of the endurance athlete: the relationship between morphology and deformation. J Am Soc Echocardiogr 25:263–271
Caselli S, Maron MS, Urbano-Moral J, Pandian NG, Maron BJ, Pelliccia A (2014) Differentiating left ventricular hypertrophy in athletes from that in patients with hypertrophic cardiomyopathy. Am J Cardiol 114:1383–1389
Zaidi A, Sheikh N, Jongman JK, Gati S, Panoulas V, Carr-White G, Papadakis M, Sharma R, Behr E, Sharma S (2015) Clinical differentiation between physiological remodeling and arrhythmogenic right ventricular cardiomyopathy in athletes with marked electrocardiographic repolarization anomalies. J Am Coll Cardiol 65:2702–2711
Pluim BM, Zwinderman AH, van der Laarse A, van der Wall EE (2000) The athlete’s heart: a meta-analysis of cardiac structure and function. Circulation 101:336–344
D’Ascenzi F, Caselli S, Solari M, Pellicia A, Cameli M, Focardi M, Padeletti M, Corrado D, Bonifazi M, Mondillo S (2016) Novel echocardiographic techniques for the evaluation of athletes’ heart: a focus on speckle-tracking echocardiography. Eur J Prev Cardiol 23(4):437–446. doi:10.1177/204748731558609
Spence AL, Carter HH, Murray CP, Oxborough D, Naylor L, George K, Green D (2013) Magnetic resonance imaging-derived right ventricular adaptations to endurance versus resistance training. Med Sci Sports Exerc 45:534–541
Utomi V, Oxborough D, Ashley E, Lord R, Fletcher S, Stembridge M, Shave R, Hoffman M, Whyte G, Somauroo J, Sharma S, George K (2014) Predominance of normal left ventricular geometry in the male ‘athlete’s’ heart’. Heart 100:1264–1271
Baggish AL, Wang F, Weiner RB, Elinoff J, Tournoux F, Boland A, Picard M, Hutter A, Wood M (2008) Training-specific changes in cardiac structure and function: a prospective and longitudinal assessment of competitive athletes. J Appl Physiol 104:1121–1128
Nottin S, Doucende G, Schuster-Beck I, Dauzat M, Obert P (2008) Alteration in left ventricular normal and shear strains evaluated by 2D-strain echocardiography in the athlete’s heart. J Physiol 586:4721–4733
Richand V, Lafitte S, Reant P, Serri K, Lafitte M, Brette S, Kerouani A, Chalabi H, Dos Santos P, Douard H, Roudaut R (2007) An ultrasound speckle tracking (two-dimensional strain) analysis of myocardial deformation in professional soccer players compared with healthy subjects and hypertrophic cardiomyopathy. Am J Cardiol 100:128–132
Sengupta PP, Narula J (2013) RV form and function: a piston pump, vortex impeller, or hydraulic ram? J Am Coll Cardiol Cardiovasc Imaging 6:636–639
Mitchell JH, Haskell W, Snell P, Van Camp SP (2005) Task force 8: classification of sports. J Am Coll Cardiol 45:1364–1367
Mosteller R (1987) Simplified calculation of body surface area. N Engl J Med 317:1098
Corrado D, Pelliccia A, Heidbuchel H, Sharma S, Link M, Basso C, Biffi A, Buja G, Delise P, Gussac I, Anastasakis A, Borjesson M, Bjornstad H, Carre F, Deligiannis A, Dugmore D, Fagard R, Hoogsteen J, Mellwig J, Panhuyzen-Goedkoop N, Solberg E, Vanhees L, Drezner J, Estes M, Iliceto S, Maron B, Peidro R, Schwartz P, Stein R, Thiene G, Zepelli P, McKenna W (2010) Recommendations for interpretation of 12-lead electrocardiogram in the athlete. Eur Heart J 31:243–259
Wharton G, Steeds R, Allen J, Phillips H, Jones R, Kanagala P, Lloyd G, Masani N, Mathew T, Oxborough D, Rana B, Sandoval J, Wheeler R, O’Gallagher K, Sharma V (2015) A minimum dataset for a standard adult transthoracic echocardiogram: a guideline protocol from the British Society of Echocardiography. Echo Res Pract 2:1
Lang RM, Badano LP, Mor-Avi V, Afilalo J, Armstrong A, Ernande L, Flachskampf F, Foster E, Goldstein S, Kuznetsova T, Lancelotti P, Muraru D, Picard M, Rietzschel E, Rudski L, Spencer K, Tsang W, Voigt J (2015) Recommendations for cardiac chamber quantification by echocardiography in adults: an update from the American Society of Echocardiography and the European Association of Cardiovascular Imaging. Eur Heart J Cardiovasc Imaging 16:233–271
Wigle ED, Sasson Z, Henderson M, Ruddy T, Fulop J, Rakowski H, Williams W (1985) Hypertrophic cardiomyopathy. The importance of the site and the extent of hypertrophy. A review. Prog Cardiovasc Dis 28:1–83
Rudski LG, Lai WW, Afilalo J, Hua L, Handschumacher M, Chandrasekaran K, Solomon S, Louie E, Schiller N (2010) Guidelines for the echocardiographic assessment of the right heart in adults: a report from the American Society of Echocardiography endorsed by the European Association of Echocardiography, a registered branch of the European Society of Cardiology, and the Canadian Society of Echocardiography. J Am Soc Echocardiogr 23:685–713
Batterham A, George K (1998) Modeling the influence of body size and composition on M-mode ecocardiographic dimensions. Am J Physiol Heart Circ Physiol 274:701–708
Dewey FE, Rosenthal D, Murphy DJ, Froelicher VF, Ashley E (2008) Does size matter? Clinical applications of scaling cardiac size and function for body size. Circulation 117:2279–2287
Oxborough D, Shave R, Warburton D, Williams K, Oxborough A, Charlesworth S, Foulds H, Hoffman M, Birch K, George K (2011) Dilatation and dysfunction of the right ventricle immediately after ultraendurance exercise: exploratory insights from conventional two-dimensional and speckle tracking echocardiography. Circ Cardiovasc Imaging 4:253–263
Morganroth J, Maron BJ, Henry W, Epstein S (1975) Comparative left ventricular dimensions in trained athletes. Ann Intern Med 82:521–524
Ho SY (2009) Anatomy and myoarchitecture of the left ventricular wall in normal and in disease. Eur J Echocardiogr 10:3–7
Heinonen I, Nesterov SV, Liukko K, Kemppainen J, Nagren K, Luotolahti M, Virsu P, Oikonen V, Nuutila P, Kujala U, Kainulainen H, Boushel R, Knuuti J, Kalliokoski K (2008) Myocardial blood flow and adenosine A2A receptor density in endurance athletes and untrained men. J Physiol 586:5193–5202
D’Andrea A, Riegler L, Golia E, Cocchia R, Scarafile R, Salerno G, ezzullo E, Nunziata L, Citro R, Cuomo S, Caso P, Di Salvo G, Cittadini A, Russo M, Calabro R, Bossone E (2013) Range of right heart measurements in top-level athletes: the training impact. Int J Cardiol 164:48–57
Teske AJ, Cox MG, De Boeck BW, Doevendans P, Hauer RN, Cramer MJ (2009) Echocardiographic tissue deformation imaging quantifies abnormal regional right ventricular function in arrhythmogenic right ventricular dysplasia/cardiomyopathy. J Am Soc Echocardiogr 22:920–927
Ho SY, Nihoyannopoulos P (2006) Anatomy, echocardiography, and normal right ventricular dimensions. Heart 92:i2–i13
Esch BT, Warburton DER. (2009) Left ventricular torsion and recoil: implications for exercise performance and cardiovascular disease. J Appl Physiol 106:362–369
Nesser H, Mor-Avi V, Gorissen W, Weinert L, Steringer-Mascherbauer R, Niel J, Sugeng L, Lang R (2009) Quantification of left ventricular volumes using three-dimensional echocardiographic speckle tracking: comparison with MRI. Eur Heart J 30:1565–1573
Acknowledgments
We would like to thank Cardiac Risk in the Young for their support of this project. This project received no external funding.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Informed consent
Informed consent was obtained from all individual participants included in the study.
Research involving human rights
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Oxborough, D., Heemels, A., Somauroo, J. et al. Left and right ventricular longitudinal strain-volume/area relationships in elite athletes. Int J Cardiovasc Imaging 32, 1199–1211 (2016). https://doi.org/10.1007/s10554-016-0910-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10554-016-0910-4