
Abstract
Mycobacterium bovis Bacillus Calmette–Guérin (BCG), an attenuated vaccine derived from M. bovis, is the only licensed vaccine against tuberculosis (TB). Despite its protection against TB in children, the protective efficacy in pulmonary TB is variable in adolescents and adults. In spite of the current knowledge of molecular biology, immunology and cell biology, infectious diseases such as TB and HIV/AIDS are still challenges for the scientific community. Genetic manipulation facilitates the construction of recombinant BCG (rBCG) vaccine that can be used as a highly immunogenic vaccine against TB with an improved safety profile, but, still, the manipulation of BCG vaccine to improve efficacy should be carefully considered, as it can bring in both favourable and unfavourable effects. The purpose of this review is not to comprehensively review the interaction between microorganisms and host cells in order to use rBCG expressing M. tuberculosis (Mtb) immunodominant antigens that are available in the public domain, but, rather, to also discuss the limitations of rBCG vaccine, expressing heterologous antigens, during manipulation that pave the way for a promising new vaccine approach.
Similar content being viewed by others


Introduction
Mycobacterium tuberculosis (Mtb) have attained global importance as a human pathogen [1]. In the early part of the 20th century, Bacillus Calmette–Guérin (BCG) vaccine was prepared at the Pasteur Institute, Lille, France in 1921 by Calmette and Guérin, consisting of a live attenuated strain of M. bovis, a closely related subspecies of Mtb. By repeating sub-culturing (231 passages) of the virulent strain on ox bile glycerine–potato media every 3 weeks over 13 years, they produced an attenuated strain, which, by the year 1919, was shown to be avirulent in guinea pigs, cows, horses, hamsters, mice and rabbits [2]. This attenuation promoted genetic deletions from M. bovis and resulted in 16 genomic regions of differentiation (RD1–RD16) as compared to the Mtb genome [3]. With reference to the regions of differentiation, RD1 is a DNA segment comprising a 10-Kb region, is deleted from all BCG strains but present in Mtb and M. bovis, encodes T-lymphocytes epitopes, viz. ESAT-6, CFP-10 and PPE proteins [4]; RD2 consists of a 10.7-Kb DNA segment and encodes proteins Mpt64 and CFP-21 [5]; RD4 corresponds to a 12.7-Kb region, deleted from M. bovis and all M. bovis BCG strains [3]. RD12 and RD13 are each about 2.5 Kb in size, and encode genes for a methyltransferase cytochrome P450 (RD12), a transcriptional regulator, a cytochrome P450 and a dehydrogenase (RD13). Both genomic regions are deleted in M. bovis and M. bovis BCG [3]. RD14 is a 9.1-Kb region of DNA encoding proteins of PE-PGRS and Rv1771 families [6]. The original BCG strain was maintained at the Pasteur Institute and it has also been distributed throughout laboratories in many countries before the original strain was lost. Most of the laboratories produced their own BCG strain and maintained it by sub-culturing [7], culminating in the evolution of more than 14 daughter strains, viz. BCG Russia, BCG Sweden, BCG Prague, BCG Moreau, BCG Phipps, BCG Pasteur 1173, BCG Tokyo, BCG Glaxo, BCG Tice, BCG Birkhaug, BCG Denmark, BCG China, BCG Frappier and BCG Connaught [8].
Following the establishment of Mtb infection within the lung, the acquired immune response is slow to identify the infection site [9] and by the time specific T cells arrive at the infection site, they play a critical role in determining the outcome of the immune response [10]. But, when the acquired immune response is absent, bacteria grow and the host dies rapidly [11]. The immunity to Mtb mainly recognises the participation of macrophages and cells of the adaptive immune system CD4+ and CD8+ T lymphocytes, as well as the cytokines TNF, IL-12 and IFN-γ, which are critical in the control of mycobacteria [12], but the improvement of current tuberculosis (TB) vaccines is limited by a lack of knowledge regarding the protective T cells capable of limiting the development of active TB [13].
Entry of Mtb into the macrophage is mediated by an array of receptors, including complement receptors, scavenger receptors and the mannose receptor [12]. While Mtb is established within lung, the bacilli are believed to be phagocytosed by the alveolar macrophages [14], neutrophils [15] and dendritic cells (DCs) [16]. There is evidence that Mtb modulates phagocytic function to prevent direct elimination from phagocytes by blocking maturation of the phagolysosome and by inhibiting apoptosis [17–19]. Mtb promotes necrotic death by inducing LXA4 (lipoxin A4), which inhibits the production of prostaglandin E2, resulting in mycobacterial spread [20–22]. MMP-1 (matrix metalloproteinases) is a collagenase that is up-regulated in TB patients and is associated with increased lung pathology in transgenic mice [23, 24] and MMP-9 has also been implicated in the pathogenesis of TB [25]. It has been demonstrated that, in humans, MMP-9 is responsible for worse outcomes in TB, suggesting a role in susceptibility to Mtb infection. Mice treated with anti-TNF antibodies or mice lacking the 55-kDa TNF receptor gene revealed that TNF is essential for the control of mycobacterial infection [19, 26]. However, lung neutrophils elicited the antigen-specific CD4+ T cells during mycobacterial infection and enhanced adaptive immune response by delivering the bacilli to DCs, which are more effective initiators of CD4+ T cell activation [27].
The pathogenicity of Mtb is related to its ability to export and secrete selected proteins that possess the capability to interact with the host cell. Mycobacterial export and secretion pathways play a central part in the survival of mycobacteria in divergent environments and hosts. Mycobacteria-specific ESX-1 and ESX-5 systems secrete/export Esx, Esp and PE/PPE proteins, respectively [28]. The key players of ESX-1 systems are ESAT-6 and CFP-10, which are encoded by genes esxA and esxB situated in the middle of the RD1 region. The PE and PPE genes are representatives of two large families of Mtb (around 7 % of the coding capacity) that encode proteins, which are derived from the motifs proline–glutamic acid (PE) and proline–proline–glutamic acid (PPE) N-terminal motifs. Within two families, several sub-families can be differentiated on the basis of middle and c-terminal sequences of its members, several of which carry highly repetitive motifs, polymorphic GC-rich repetitive sequences (PGRS) and major polymorphic tandem repeats (MPTR) [4, 29]. From a phylogenetics point of view, PE and PPE proteins seem to be associated with ESX systems [30], which are associated with protein secretion and export of the concerned domains. It has been shown that Mtb PE-LipY and M. marinum PPE-LipY were both exported to the bacterial surface associated with ESX-5-mediated secretion [31].
BCG vaccine can prevent miliary and meningeal TB in children, but its protective value in adults against pulmonary TB is questionable. A 15-year follow-up trial of BCG vaccination carried out in Chingleput and enrolling more than 360,000 individuals found that BCG offered no protection against pulmonary TB in adults [32]. The effectiveness of BCG in preventing TB in adults and infants is highly variable, with efficacies ranging between 0 to 80 % (average 50 %) having been reported from multiple clinical trials performed during the 20th century [33]. The reason for such variable protection may include BCG strain variation, the genetic variability amongst and different ages of the vaccinated individuals, routes of administration, geographic location, the dose of vaccine, interference by environmental, mycobacterial and helminthic infection, and patient nutritional status [6, 34, 35]. But, still, due to having unique properties, BCG vaccine could not be replaced by another vaccine because: (i) of the route of delivery of BCG vaccine (vaccine was delivered orally to humans between 1921 until the late 1940s and, since the late 1940s, administration followed the percutaneous or intradermal route); (ii) it is feasible to produce as compared to other vaccines; (iii) it is unaffected by maternal antibodies and, therefore, it can be given at any time after birth; (iv) it is stable and secure; (v) BCG is usually given as single dose, eliciting a long-lasting immunity [36].
Many factors have forced investigators to look for an alternative to BCG vaccine or to enhance the efficacy of BCG vaccine. In this context, it is required to have a better understanding of the interactions between microorganisms and host cells for a rational recommendation on the use of wild-type BCG and recombinant BCG (rBCG) should be explicated. The interest in BCG vaccine increased due to the development of different genetic systems for expressing foreign antigens in mycobacteria via different shuttle vector systems to express and secrete heterologous antigens and strategies for the transformation of mycobacteria.
Manipulation during rBCG vaccine can have both positive and negative aspects. In one aspect, rBCG expressing and secreting the immunodominant antigen Ag85B of Mtb was found to promote levels of protection greater than conventional BCG [37]. rBCG strains have been constructed which express cytokines such as IFN-γ or IL-2, IL-12 and granulocyte macrophage colony-stimulating factor (GM-CSF) to stimulate more potent immune responses against Mtb [38, 39]. In reverse, the protective efficacy of rBCG over-expressing LipY (PE_PGRS63 of Mtb) and the profile of host immune response generate an additional concern. During this manipulation, we found that over-expression of LipY in M. bovis BCG demolished the efficacy of BCG vaccine to protect against infection of Mtb, and the underlying mechanism was found to be down-regulation of the host immune system [40].
This review explains the knowledge available in the public domain with reference to rBCG strains that modulate immune response. In addition, some immunological deficits during the manipulation of rBCG over-expressing Mtb antigens are also discussed.
Approaches towards BCG vaccine
The way BCG vaccine has been managed in different countries and manufacturing units for several decades raises serious concern. After the first successful vaccination, BCG strains were distributed throughout the world and generated differently in various laboratories for several decades, resulting in both phenotypic and genotypic differences not only compared to the original BCG parent strain but also between the various BCG daughter strains [41]. Although the effect of these mutations is far from clear, strain variability has been suggested as an explanation for the variable protection found in clinical trials using different strains of BCG [6, 35]. It has been suggested that, over time, BCG vaccine may have lost a number of genes with potential relevance for protective immunity or, in other words, has gradually been attenuated to impotence [42]. Hence, it is important to provide BCG with selected Mtb-specific genes in order to enhance its immunogenicity and protective efficacy against TB [6].
Comparative genomics has yielded valuable information on the differences between BCG and virulent Mtb, revealing the absence of a number of genes and regions designated RD1–RD16 and encompassing 129 open reading frames in BCG vaccines [6, 43, 44]. Some of these genes which are present in virulent mycobacteria and absent in BCG are even likely associated with virulence and could play an important role for the failure of BCG. Reintroduction of selected genes from RD1–RD16 to BCG has, therefore, been suggested as a way towards enhancing the protective efficacy of the existing BCG vaccine [45]. Reintroduction of the RD1 region encodes Mtb ESAT-6 and CFP10 antigens into BCG, enhancing the protection against disseminated Mtb infection in mice and guinea pig [46].
There are some vaccination strategies currently in the development, all of them are primarily aimed at delaying disease outbreak and can potentially be optimised to achieve sterile eradication [47]. One of them takes advantage of prime vaccination with conventional BCG to strengthen the immune response by booster with a subunit vaccine. Several subunit vaccines have already entered phase I and phase II clinical trials [48, 49]. Agger and Andersen have shown that a subunit vaccine is not influenced by sensitisation with environmental mycobacteria and stimulates a protective T cell response, whereas BCG is dependent on the initial multiplication for its activity [50]. An alternative vaccination strategy is to replace BCG with a recombinant live vaccine, and two vaccine candidates of that type have now been entered into clinical trials. The first candidate is an rBCG expressing antigen 30-kDa major secretory protein [37]. rBCG expressing membrane-perforating listeriolysin (Hly) of Listeria monocytogenes showed better protection against Mtb aerosol infection than the parental BCG strain [51]. In a further study, comparison has been made to compare immune responses after vaccination with rBCG:ΔureC:Hly (which expresses Hly of L. monocytogenes and is devoid of urease C) and parental BCG with reference to identifying biomarkers that correlate with protection in a murine model of TB infection. The data revealed that rBCG induced type 1 and type 17 cytokine responses, whereas type 1 response was only induced by parental BCG. rBCG:ΔureC:Hly is more efficient than parental BCG against pulmonary TB in pre-clinical studies and has been successfully proven to be immunogenic in phase I clinical trials [52].
Immunological deficit of BCG vaccine
There are a number of issues in developing vaccines with enhanced protective immunity against TB. Several hypotheses have suggested that one reason for the attenuation of immunological characteristics of BCG is the lack of T cell antigens in BCG [45]. CD8+ T cells play a very important role in the host defence against TB infections [19], by using at least three different mechanisms: (a) direct extracellular killing of mycobacteria through antimicrobial activity,, (b) cytolysis of infected cells and (c) release of IFN-γ. Various human studies have demonstrated that CD8+ T cells specifically recognised Mtb-infected macrophages, as demonstrated by the production of IFN-γ, and lyse the infected macrophages, resulting in the simultaneous eradication of bacteria by the release of granules containing perforin and granulysin. So, CD8 cytotoxic T lymphocytes reduced the viability of the intracellular Mtb and can, hence, contribute to effective immunity against the pathogen [53, 54]. The crucial role of MHC class I-restricted CD8+ T cells was shown by the failure of β2-microglobulin (β2m)-deficient mice to control experimental Mtb infection [55]. Second, infection with mycobacteria is not able to induce sterilising immunity against reinfection with the same mycobacterium after clearance of the original infection with antibiotics. So, there is no vaccine against TB that has elicited sterilising immunity [56]. Third, the variability of the BCG vaccine has been attributed to genetic or nutritional differences between populations, as well as several ecological factors such as temperature, sunlight exposure and ultraviolet radiation that correlate with latitude, where the higher prevalence of environmental mycobacteria in tropical regions has been suggested to be the single most important factor for the observed low efficacy of BCG in these regions [35]. Therefore, BCG might attain its full potential only in developed countries where the population is not heavily exposed to environmental mycobacteria, because in the trials performed in the 1940s and 1950s in developed countries like Denmark, UK and North America, the BCG vaccine was found to be highly efficient (70–80 %), whereas more recent trials in developing countries demonstrated less or no protection against pulmonary TB [57].
Animal experiments showing protection provided by environmental mycobacteria partly conceal the effect of a subsequent BCG vaccination [58]. Rook et al. demonstrated that the environmental mycobacteria have a direct antagonistic influence and shift the immune response towards a T helper 2 (Th2) direction [59]. When Th2 cytokines are induced by exposure to high levels of the environmental mycobacteria or by vaccination, they can have a disease exacerbating role and suggested, on the basis of animal studies, that infection with environmental mycobacteria changed the immune reaction towards a detrimental humoral response that could not be abolished by following BCG vaccination [60]. In human TB patients, they correspond with poor clinical outcome [61]. Therefore, it is clear from the evidence that the effects of exposure to environmental mycobacteria on both the level of interfering with the efficacy of BCG and the degree of benefit of protection against Mtb are still not clear.
The cloud of doubt surrounding the efficacy of BCG vaccine has inspired investigators to improve BCG by making recombinants of various kinds, including genes of secretory proteins, cytokines, immunomodulators etc. Encouraging but mixed results have been obtained that correlate with both favourable and unfavourable consequences during the construction of rBCG.
rBCG vaccine
Human TB
Recombinant DNA technology enabled the construction of rBCG strains to be used as improved candidate TB vaccines with better immunogenicity. In this line of rBCG constructs with enhanced immunostimulatory properties, BCG was genetically engineered with different immunodominant antigens and cytokines. rBCG expressing and secreting the immunodominant antigen Ag85B of Mtb was found to promote levels of protection greater than conventional BCG [37]. Two rBCG strains (Connaught and Tice) over-expressing Ag85B in a guinea pig model of pulmonary TB were more efficient than BCG vaccine. rBCG strains have been constructed which express cytokines such as IFN-γ or IL-2, IL-12 and GM-CSF to stimulate more potent immune responses against Mtb [38, 39]. To achieve enhanced immunostimulatory properties, BCG was engineered to secrete r-human IFN-α (rhIFNα) under the control of mycobacterial heat shock protein (Hsp) 60 promoter and the α-antigen signal sequence. When compared with control BCG, rhIFNα-BCG was substantially more active in inducing the production of IFN-γ from human peripheral blood mononuclear cells [62].
It is mostly considered that important T cell antigens are missing in BCG, which is backed by extensive data published on ESAT-6, a low molecular mass protein of the RD1 region. It is considered a strong inducer of T cells [63–66] but deleted in BCG. Immunisation with ESAT-6 and DNA vaccines encoding ESAT-6 evoked protective responses [67–69]. Even a single epitope derived from ESAT-6 in the adjuvant DDA/MPL was found to confer efficient protection comparable to the protection afforded by BCG vaccine [70]. The immunomodulator effect of BCG vaccine has been recorded [39, 71], which indicates that cytokines plays a very significant role in improving this effect. This approach has allowed modulation of the immune system to respond with a specific and desired pattern of cytokines [72]. In another similar study, rBCG producing IL-2 enhanced a strong type 1 immune response in a murine model [73] and rBCG secreting IL-18 increased the type 1 immune response with the production of antigen-specific IFN-γ in vaccinated mice [74]. rBCG expressing IFN-γ resulted in an alteration in the pattern of inflammation and local tissue fibrosis.
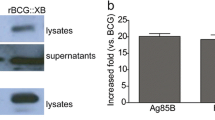
In addition, local expression of IFN-γ by rBCG resulted in more efficient bacterial clearance, which is accompanied by a reduction in tissue pathology [38]. Guinea pigs immunized with rBCG30, a BCG over-expressing the 85B antigen, and challenged with Mtb by aerosol, had less organ pathology, fewer bacteria in their lungs and spleen, and significantly greater survival than guinea pigs immunized with the parent strain of BCG [37]. This rBCG vaccine was the first vaccine reported to induce greater protective immunity against TB than the parent BCG vaccine in an animal model. Recently, the first double-blind phase I trial of rBCG30 in 35 adult humans showed that rBCG30 induced significantly increased Ag85b-specific T cell lymph proliferation, IFN-γ secretion and increased number of Ag85b-specific T cells capable of inhibiting intracellular mycobacteria [75]. An immense attempt has been dedicated to the assessment of BCG over-expressing members of the Ag85 complex (Ag85A, Ag85B and Ag85C), either individually [76, 77] or Ag85B associated to ESAT-6 [78] or Ag85B associated to other antigens [79] or IL-15 [80]. Several of these constructs afforded better protection than standard BCG. In another study, BCG was equipped with Hly of L. monocytogenes and showed significantly improved protection in a mouse model when compared to the parental BCG strain following aerosol challenge with Mtb [51]. Mice immunized with rBCG co-expressing Ag85B, CFP10 and interleukin-12 (rBCG::Ag85B–CFP10–IL-12) elicited strong immunogenicity and attenuation of mycobacterial growth as compared to BCG vaccine [81], and further extension of this study showed that rBCG::Ag85B–CFP10–IL-12 augmented the protection against Mtb by increasing the Th1 polarised response [82].
TB vaccines include viral vectored, mycobacterial whole cell or extract, protein or adjuvant, attenuated Mtb and recombinant live. Approximately 15 TB vaccine candidates are in various phases of clinical trials. According to the pipeline for new TB vaccines in August 2014, there is a phase I clinical trial including six vaccines, AdAg85A, TB/FLU-04 L and Crucell Ad35/MVA85A (viral vectored), MTBVAC (attenuated Mtb strain), ID93 + GLA-SE (protein/adjuvant) and DAR 901(mycobacterial whole cell or extract), and a phase II clinical trial including six vaccines, VPM 1002 (rBCG), H1/H56/H4 + IC3 (protein/adjuvant), RUTI (mycobacterial whole cell or extract) and Crucell Ad35/AERAS-402 (viral vectored). MVA85A (viral vectored) and M72 + AS01E (protein/adjuvant) are in a phase IIb clinical trial and one vaccine using M. vaccae (mycobacterial whole cell or extract) is in a phase III clinical trial [83].
Bovine TB
Mycobacterium bovis, causing bovine TB, is not only a serious animal or zoonotic disease that causes economic loss, but it is also a threat to public health [84]. The only current human vaccine, M. bovis BCG, provides protection against bovine TB, but with variable protective efficacy [85]. There have been several improvements in cattle vaccine development, like most of the promising approaches, including BCG-DNA [86], BCG–virus-vectored vaccine [87], BCG–protein vaccine [88, 89] and adjuvant vaccines that induce significantly superior protection compared to BCG alone [90]. Improved vaccines based on rBCG vaccines enhanced the protective efficacy of BCG vaccine, as shown in several studies with reference to human TB in a previous section of this paper, and several studies even demonstrated the enhanced protective immunity of rBCG vaccine against M. bovis challenge in cattle. It has been described that a live rBCG vaccine, rBCG30, provided more protection against Mtb in a guinea pig model of pulmonary TB [37]. The same rBCG vaccine produced greater protective immunity than BCG alone against M. bovis challenge, indicating a lower burden of M. bovis in the lung and spleen in rBCG30 immunised guinea pig [91].
When the protective immune response of M. bovis deleted mce2A and mce2B genes (double deletion mutant, M. bovis Δmce2) as an experimental vaccine, evaluation in cattle showed protection against M. bovis challenge, indicating that M. bovis Δmce2 is a promising vaccine candidate against M. bovis pathogenesis in cattle [92]. Khatri et al. tested the immunogenicity of two rBCG strains, namely, BCG Pasteur Δzmp1::aph and BCG Danish Δzmp1, in cattle and found that both strains induced superior T cell memory response compared to BCG alone [93]. Recently, the evaluation of M. bovis double knock-out mce2-phoP tested in mice as a vaccine candidate demonstrated that mice immunized with the double mutant protected against challenge with M. bovis [94]. A successive trial with a number of animal species specifies that the oral route of BCG vaccination attenuates the disease extremity after experimental challenge with M. bovis [85] and the administration of oral BCG vaccination was shown to prevent infection of wild possums against natural exposure to M. bovis [95]. The potential of oral vaccination for controlling TB has also been demonstrated in badgers [96].
Limitations of rBCG vaccine
In contrast to the above findings, one report has indicated that there is no effect on the immunogenicity of BCG vaccine during construction and a few others reported attenuation of immunogenicity of BCG vaccine during the construction of rBCG vaccine. Hereof, over-expression of the 19-kDa antigen (lipoprotein) did not change the capacity of BCG vaccine to protect against Mtb in mice [97]. It was noted that the 19-kDa antigen (Rv3763), a lipoprotein of Mtb, triggers high levels of IL-12 from macrophages in addition to suppressing the antigen presentation signalling cascade and its immunomodulatory properties. The polarisation of host immune responses towards Th2 subtypes confers the abolition of immunogenicity of rBCG19N (rBCG expressing Mtb 19-kDa lipoprotein) when used as a live vaccine against Mtb in guinea pigs indicates that over-expression of the 19-kDa antigen attenuates the BCG vaccine efficacy [98]. We assessed the immunogenicity of rBCG over-expressing LipY (PE_PGRS63 of Mtb) in mice against Mtb and found attenuation of the immunogenicity of rBCG vaccine to protect against Mtb infection in a murine model. rBCG over-expressing LipY vaccine offered no protection against challenge of Mtb as evident by the parameters, viz. viable counts of tubercle bacilli in the lungs and weight of infected mice, and pathology of the lungs and survival of challenged mice and immune response generated by this rBCG vaccine in murine model suggested down-regulation from Th1 to Th2 type [40]. These findings suggest that the detrimental effects mask the development of new promising rBCG vaccine approaches.
rBCG vaccine and other diseases
BCG vaccine usually prevents TB, but it is also an effective treatment for some non-muscle-invasive bladder cancers and has been used to treat it for more than 30 years. A putative model of the mechanism of action of BCG in bladder cancer has been shown elsewhere [99]. Raymond Pearl suggested that mycobacteria might be applicable to cancer therapy, observing in an autopsy study that cancer was less common in patients with active TB. Among various cancers including colon cancer, bladder cancer, lung cancer, leukaemia and melanoma, bladder cancer is the only cancer in which BCG is commonly used [71].
Several studies have reported that rBCG induced protection by using parasite, bacterial and viral antigens. Mice immunized subcutaneously with rBCG expressing the LCR1 antigen of Leishmania chagasi drove a type 1 immune response with IFN-γ production and, consequently, protection against challenge [100]. Interestingly, this rBCG was unable to cause significant levels of IFN-γ production when mice were immunized intraperitoneally and failed to ensure protection, advising that the route of administration is important for protection against L. chagasi. In another supporting study, BCG expressing the Sm14 antigen of Schistosoma mansoni induced IFN-γ production and predominantly type 1 cellular immune response in a murine model. Especially, the vaccinated animals were protected against cercarial challenge in this study [101]. Hamsters immunized with BCG expressing the LipL32 antigen of Leptospira interrogans were protected against challenge with L. interrogans. Autopsy examinations acknowledge, in this study, that rBCG-LipL32 was able to draw sterilising immunity against L. interrogans [102]. Rabbits inoculated with BCG expressing the cottontail rabbit papillomavirus L1 antigen developed neutralizing antibodies and showed smaller papilloma than the control group, demonstrating that rBCG could be used as a possible prophylactic against papillomavirus [103]. These studies manifest that rBCG has great potential as a vaccine vector, and rBCG vaccine offering protection against TB and several other diseases is credible.
Concluding remarks
Despite several controversies, Bacillus Calmette–Guérin (BCG) has several advantages, hence it is not easy to replace it with other vaccine candidates. The improvement of BCG remains the best alternative for the rational design of a vaccine against tuberculosis (TB). The post-genomic era could lead to the identification of novel Mycobacterium tuberculosis (Mtb) antigens that are absent from the BCG proteome and could, consequently, be applied to efficiently enhance the immunogenicity of BCG vaccine. The rationale to give a chance to foreign antigens in BCG is to enhance the efficacy and adjuvanticity of BCG as a recombinant vaccine. Success may play an important role when BCG continues to be applied to neonates, and boost the best subunit vaccine candidate, to stretch out the protection and efficacy of a vaccine. Improvements in recombinant BCG (rBCG) by expressing dominant antigens contained in the subunit vaccine are used for booster vaccinations and further improving the immune stimulatory capacity not merely as a vaccine against TB but also carrying a major role against many infectious diseases and, hence, making it a truly multi-valent vaccine. Manipulation of BCG vaccine should be carefully considered when discussing the potential of substituting BCG with new rBCG vaccines by using heterologous antigen expression in BCG, which also tends to attenuate the immune response in the murine model of TB and shares some disadvantages in rBCG vaccines.
References
Bloom BR, Murray CJ (1992) Tuberculosis: commentary on a reemergent killer. Science 57:1055–1064
Sakula A (1983) BCG: who were Calmette and Guérin? Thorax 38:806–812
Brosch R, Gordon SV, Pym A, Eiglmeier K, Garnier T, Cole ST (2000) Comparative genomics of the mycobacteria. Int J Med Microbiol 290:143–152. doi:10.1016/S1438-4221(00)80083-1
Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D et al (1998) Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393:537–544. doi:10.1038/31159
Joung SM, Ryoo S (2013) BCG vaccine in Korea. Clin Exp Vaccine Res 2:83–91. doi:10.7774/cevr.2013.2.2.83
Behr MA, Wilson MA, Gill WP, Salamon H, Schoolnik GK, Rane S et al (1999) Comparative genomics of BCG vaccines by whole-genome DNA microarray. Science 284:1520–1523
Tripathy SP (1983) The case for B.C.G. Ann Natl Acad Med Sci 19:11–21
da Costa AC, Nogueira SV, Kipnis A, Junqueira-Kipnis AP (2014) Recombinant BCG: innovations on an Old vaccine. Scope of BCG strains and strategies to improve long-lasting memory. Front Immunol 5:152. doi:10.3389/fimmu.2014.00152
Cooper AM (2009) Cell-mediated immune responses in tuberculosis. Annu Rev Immunol 27:393–422. doi:10.1146/annurev.immunol.021908.132703
Pearl JE, Torrado E, Tighe M, Fountain JJ, Solache A, Strutt T et al (2012) Nitric oxide inhibits the accumulation of CD4+CD44hiTbet+CD69lo T cells in mycobacterial infection. Eur J Immunol 42:3267–3279. doi:10.1002/eji.201142158
Geldmacher C, Zumla A, Hoelscher M (2012) Interaction between HIV and Mycobacterium tuberculosis: HIV-1-induced CD4 T-cell depletion and the development of active tuberculosis. Curr Opin HIV AIDS 7:268–275. doi:10.1097/COH.0b013e3283524e32
O’Garra A, Redford PS, McNab FW, Bloom CI, Wilkinson RJ, Berry MPR (2013) The immune response in tuberculosis. Annu Rev Immunol 31:475–527. doi:10.1146/annurev-immunol-032712-095939
Robinson RT, Orme IM, Cooper AM (2015) The onset of adaptive immunity in the mouse model of tuberculosis and the factors that compromise its expression. Immunol Rev 264:46–59. doi:10.1111/imr.12259
Schlesinger LS (1996) Entry of Mycobacterium tuberculosis into mononuclear phagocytes. Curr Top Microbiol Immunol 215:71–96
Eum S-Y, Kong J-H, Hong M-S, Lee Y-J, Kim J-H, Hwang S-H et al (2010) Neutrophils are the predominant infected phagocytic cells in the airways of patients with active pulmonary TB. Chest 137:122–128. doi:10.1378/chest.09-0903
Wolf AJ, Linas B, Trevejo-Nuñez GJ, Kincaid E, Tamura T, Takatsu K et al (2007) Mycobacterium tuberculosis infects dendritic cells with high frequency and impairs their function in vivo. J Immunol 179:2509–2519
Bhatt K, Salgame P (2007) Host innate immune response to Mycobacterium tuberculosis. J Clin Immunol 27:347–362. doi:10.1007/s10875-007-9084-0
Chen M, Gan H, Remold HG (2006) A mechanism of virulence: virulent Mycobacterium tuberculosis strain H37Rv, but not attenuated H37Ra, causes significant mitochondrial inner membrane disruption in macrophages leading to necrosis. J Immunol 176:3707–3716
Flynn JL, Chan J (2001) Immunology of tuberculosis. Annu Rev Immunol 19:93–129. doi:10.1146/annurev.immunol.19.1.93
Chen M, Divangahi M, Gan H, Shin DSJ, Hong S, Lee DM et al (2008) Lipid mediators in innate immunity against tuberculosis: opposing roles of PGE2 and LXA4 in the induction of macrophage death. J Exp Med 205:2791–2801. doi:10.1084/jem.20080767
Divangahi M, Chen M, Gan H, Desjardins D, Hickman TT, Lee DM et al (2009) Mycobacterium tuberculosis evades macrophage defenses by inhibiting plasma membrane repair. Nat Immunol 10:899–906. doi:10.1038/ni.1758
Behar SM, Divangahi M, Remold HG (2010) Evasion of innate immunity by Mycobacterium tuberculosis: is death an exit strategy? Nat Rev Microbiol 8:668–674. doi:10.1038/nrmicro2387
Elkington P, Shiomi T, Breen R, Nuttall RK, Ugarte-Gil CA, Walker NF et al (2011) MMP-1 drives immunopathology in human tuberculosis and transgenic mice. J Clin Invest 121:1827–1833. doi:10.1172/JCI45666
Elkington PTG, Nuttall RK, Boyle JJ, O’Kane CM, Horncastle DE, Edwards DR et al (2005) Mycobacterium tuberculosis, but not vaccine BCG, specifically upregulates matrix metalloproteinase-1. Am J Respir Crit Care Med 172:1596–1604. doi:10.1164/rccm.200505-753OC
Elkington PT, Ugarte-Gil CA, Friedland JS (2011) Matrix metalloproteinases in tuberculosis. Eur Respir J 38:456–464. doi:10.1183/09031936.00015411
Flynn JL, Goldstein MM, Chan J, Triebold KJ, Pfeffer K, Lowenstein CJ et al (1995) Tumor necrosis factor-alpha is required in the protective immune response against Mycobacterium tuberculosis in mice. Immunity 2:561–572
Blomgran R, Ernst JD (2011) Lung neutrophils facilitate activation of naive antigen-specific CD4+ T cells during Mycobacterium tuberculosis infection. J Immunol 186:7110–7119. doi:10.4049/jimmunol.1100001
Majlessi L, Prados-Rosales R, Casadevall A, Brosch R (2015) Release of mycobacterial antigens. Immunol Rev 264:25–45. doi:10.1111/imr.12251
Bottai D, Brosch R (2009) Mycobacterial PE, PPE and ESX clusters: novel insights into the secretion of these most unusual protein families. Mol Microbiol 73:325–328. doi:10.1111/j.1365-2958.2009.06784.x
Gey van Pittius NC, Sampson SL, Lee H, Kim Y, van Helden PD, Warren RM (2006) Evolution and expansion of the Mycobacterium tuberculosis PE and PPE multigene families and their association with the duplication of the ESAT-6 (esx) gene cluster regions. BMC Evol Biol 6:95. doi:10.1186/1471-2148-6-95
Daleke MH, Cascioferro A, de Punder K, Ummels R, Abdallah AM, van der Wel N et al (2011) Conserved Pro-Glu (PE) and Pro-Pro-Glu (PPE) protein domains target LipY lipases of pathogenic mycobacteria to the cell surface via the ESX-5 pathway. J Biol Chem 286:19024–19034. doi:10.1074/jbc.M110.204966
[No authors listed] (1999) Fifteen year follow up of trial of BCG vaccines in south India for tuberculosis prevention. Tuberculosis research centre (ICMR), Chennai. Indian J Med Res 110:56–69
Colditz GA, Brewer TF, Berkey CS, Wilson ME, Burdick E, Fineberg HV et al (1994) Efficacy of BCG vaccine in the prevention of tuberculosis. Meta-analysis of the published literature. JAMA 271:698–702
Black GF, Dockrell HM, Crampin AC, Floyd S, Weir RE, Bliss L et al (2001) Patterns and implications of naturally acquired immune responses to environmental and tuberculous mycobacterial antigens in northern Malawi. J Infect Dis 184:322–329. doi:10.1086/322042
Fine PE (1995) Variation in protection by BCG: implications of and for heterologous immunity. Lancet 346:1339–1345
Bastos RG, Borsuk S, Seixas FK, Dellagostin OA (2009) Recombinant Mycobacterium bovis BCG. Vaccine 27:6495–6503. doi:10.1016/j.vaccine.2009.08.044
Horwitz MA, Harth G, Dillon BJ, Maslesa-Galic’ S (2000) Recombinant bacillus calmette-guerin (BCG) vaccines expressing the Mycobacterium tuberculosis 30-kDa major secretory protein induce greater protective immunity against tuberculosis than conventional BCG vaccines in a highly susceptible animal model. Proc Natl Acad Sci U S A 97:13853–13858. doi:10.1073/pnas.250480397
Wangoo A, Brown IN, Marshall BG, Cook HT, Young DB, Shaw RJ (2000) Bacille Calmette-Guérin (BCG)-associated inflammation and fibrosis: modulation by recombinant BCG expressing interferon-gamma (IFN-gamma). Clin Exp Immunol 119:92–98
Murray PJ, Aldovini A, Young RA (1996) Manipulation and potentiation of antimycobacterial immunity using recombinant bacille Calmette-Guérin strains that secrete cytokines. Proc Natl Acad Sci U S A 93:934–939
Singh VK, Srivastava V, Singh V, Rastogi N, Roy R, Shaw AK et al (2011) Overexpression of Rv3097c in Mycobacterium bovis BCG abolished the efficacy of BCG vaccine to protect against Mycobacterium tuberculosis infection in mice. Vaccine 29:4754–4760. doi:10.1016/j.vaccine.2011.04.086
Behr MA (2002) BCG—different strains, different vaccines? Lancet Infect Dis 2:86–92
Behr MA, Small PM (1997) Has BCG attenuated to impotence? Nature 389:133–134. doi:10.1038/38151
Gordon SV, Brosch R, Billault A, Garnier T, Eiglmeier K, Cole ST (1999) Identification of variable regions in the genomes of tubercle bacilli using bacterial artificial chromosome arrays. Mol Microbiol 32:643–655
Jungblut PR, Schaible UE, Mollenkopf HJ, Zimny-Arndt U, Raupach B, Mattow J et al (1999) Comparative proteome analysis of Mycobacterium tuberculosis and Mycobacterium bovis BCG strains: towards functional genomics of microbial pathogens. Mol Microbiol 33:1103–1117
Mahairas GG, Sabo PJ, Hickey MJ, Singh DC, Stover CK (1996) Molecular analysis of genetic differences between Mycobacterium bovis BCG and virulent M. bovis. J Bacteriol 178:1274–1282
Pym AS, Brodin P, Majlessi L, Brosch R, Demangel C, Williams A et al (2003) Recombinant BCG exporting ESAT-6 confers enhanced protection against tuberculosis. Nat Med 9:533–539. doi:10.1038/nm859
Kaufmann SHE, Baumann S, Nasser Eddine A (2006) Exploiting immunology and molecular genetics for rational vaccine design against tuberculosis. Int J Tuberc Lung Dis 10:1068–1079
Weinrich Olsen A, van Pinxteren LA, Meng Okkels L, Birk Rasmussen P, Andersen P (2001) Protection of mice with a tuberculosis subunit vaccine based on a fusion protein of antigen 85b and esat-6. Infect Immun 69:2773–2778. doi:10.1128/IAI.69.5.2773-2778.2001
Goonetilleke NP, McShane H, Hannan CM, Anderson RJ, Brookes RH, Hill AVS (2003) Enhanced immunogenicity and protective efficacy against Mycobacterium tuberculosis of bacille Calmette-Guérin vaccine using mucosal administration and boosting with a recombinant modified vaccinia virus Ankara. J Immunol 171:1602–1609
Agger EM, Andersen P (2002) A novel TB vaccine; towards a strategy based on our understanding of BCG failure. Vaccine 21:7–14
Grode L, Seiler P, Baumann S, Hess J, Brinkmann V, Nasser Eddine A et al (2005) Increased vaccine efficacy against tuberculosis of recombinant Mycobacterium bovis bacille Calmette-Guérin mutants that secrete listeriolysin. J Clin Invest 115:2472–2479. doi:10.1172/JCI24617
Desel C, Dorhoi A, Bandermann S, Grode L, Eisele B, Kaufmann SHE (2011) Recombinant BCG ΔureC hly+ induces superior protection over parental BCG by stimulating a balanced combination of type 1 and type 17 cytokine responses. J Infect Dis 204:1573–1584. doi:10.1093/infdis/jir592
Stenger S, Mazzaccaro RJ, Uyemura K, Cho S, Barnes PF, Rosat JP et al (1997) Differential effects of cytolytic T cell subsets on intracellular infection. Science 276:1684–1687
Cho S, Mehra V, Thoma-Uszynski S, Stenger S, Serbina N, Mazzaccaro RJ et al (2000) Antimicrobial activity of MHC class I-restricted CD8+ T cells in human tuberculosis. Proc Natl Acad Sci U S A 97:12210–12215. doi:10.1073/pnas.210391497
Flynn JL, Goldstein MM, Triebold KJ, Koller B, Bloom BR (1992) Major histocompatibility complex class I-restricted T cells are required for resistance to Mycobacterium tuberculosis infection. Proc Natl Acad Sci U S A 89:12013–12017
Britton WJ, Palendira U (2003) Improving vaccines against tuberculosis. Immunol Cell Biol 81:34–45. doi:10.1046/j.0818-9641.2002.01143.x
Fine PE (1989) The BCG story: lessons from the past and implications for the future. Rev Infect Dis 11(Suppl 2):S353–S359
Palmer CE, Long MW (1966) Effects of infection with atypical mycobacteria on BCG vaccination and tuberculosis. Am Rev Respir Dis 94:553–568
Rook GA, Bahr GM, Stanford JL (1981) The effect of two distinct forms of cell-mediated response to mycobacteria on the protective efficacy of BCG. Tubercle 62:63–68
Hernandez-Pando R, Pavön L, Arriaga K, Orozco H, Madrid-Marina V, Rook G (1997) Pathogenesis of tuberculosis in mice exposed to low and high doses of an environmental mycobacterial saprophyte before infection. Infect Immun 65:3317–3327
Seah GT, Scott GM, Rook GA (2000) Type 2 cytokine gene activation and its relationship to extent of disease in patients with tuberculosis. J Infect Dis 181:385–389. doi:10.1086/315200
Luo Y, Chen X, Han R, O’Donnell MA (2001) Recombinant bacille Calmette-Guérin (BCG) expressing human interferon-alpha 2B demonstrates enhanced immunogenicity. Clin Exp Immunol 123:264–270
Brandt L, Oettinger T, Holm A, Andersen AB, Andersen P (1996) Key epitopes on the ESAT-6 antigen recognized in mice during the recall of protective immunity to Mycobacterium tuberculosis. J Immunol 157:3527–3533
Andersen P, Andersen AB, Sørensen AL, Nagai S (1995) Recall of long-lived immunity to Mycobacterium tuberculosis infection in mice. J Immunol 154:3359–3372
Lalvani A, Nagvenkar P, Udwadia Z, Pathan AA, Wilkinson KA, Shastri JS et al (2001) Enumeration of T cells specific for RD1-encoded antigens suggests a high prevalence of latent Mycobacterium tuberculosis infection in healthy urban Indians. J Infect Dis 183:469–477. doi:10.1086/318081
Ravn P, Demissie A, Eguale T, Wondwosson H, Lein D, Amoudy HA et al (1999) Human T cell responses to the ESAT-6 antigen from Mycobacterium tuberculosis. J Infect Dis 179:637–645. doi:10.1086/314640
Kamath AT, Feng CG, Macdonald M, Briscoe H, Britton WJ (1999) Differential protective efficacy of DNA vaccines expressing secreted proteins of Mycobacterium tuberculosis. Infect Immun 67:1702–1707
Li Z, Howard A, Kelley C, Delogu G, Collins F, Morris S (1999) Immunogenicity of DNA vaccines expressing tuberculosis proteins fused to tissue plasminogen activator signal sequences. Infect Immun 67:4780–4786
Brandt L, Elhay M, Rosenkrands I, Lindblad EB, Andersen P (2000) ESAT-6 subunit vaccination against Mycobacterium tuberculosis. Infect Immun 68:791–795
Olsen AW, Hansen PR, Holm A, Andersen P (2000) Efficient protection against Mycobacterium tuberculosis by vaccination with a single subdominant epitope from the ESAT-6 antigen. Eur J Immunol 30:1724–1732. doi:10.1002/1521-4141(200006)30:6<1724::AID-IMMU1724>3.0.CO;2-A
Alexandroff AB, Jackson AM, O’Donnell MA, James K (1999) BCG immunotherapy of bladder cancer: 20 years on. Lancet 353:1689–1694. doi:10.1016/S0140-6736(98)07422-4
O’Donnell MA, Aldovini A, Duda RB, Yang H, Szilvasi A, Young RA et al (1994) Recombinant Mycobacterium bovis BCG secreting functional interleukin-2 enhances gamma interferon production by splenocytes. Infect Immun 62:2508–2514
Young SL, O’Donnell MA, Buchan GS (2002) IL-2-secreting recombinant bacillus Calmette Guerin can overcome a Type 2 immune response and corticosteroid-induced immunosuppression to elicit a Type 1 immune response. Int Immunol 14:793–800
Biet F, Kremer L, Wolowczuk I, Delacre M, Locht C (2002) Mycobacterium bovis BCG producing interleukin-18 increases antigen-specific gamma interferon production in mice. Infect Immun 70:6549–6557
Hoft DF, Blazevic A, Abate G, Hanekom WA, Kaplan G, Soler JH et al (2008) A new recombinant bacille Calmette-Guérin vaccine safely induces significantly enhanced tuberculosis-specific immunity in human volunteers. J Infect Dis 198:1491–1501. doi:10.1086/592450
Dhar N, Rao V, Tyagi AK (2004) Immunogenicity of recombinant BCG vaccine strains overexpressing components of the antigen 85 complex of Mycobacterium tuberculosis. Med Microbiol Immunol 193:19–25. doi:10.1007/s00430-002-0170-x
Sugawara I, Li Z, Sun L, Udagawa T, Taniyama T (2007) Recombinant BCG Tokyo (Ag85A) protects cynomolgus monkeys (Macaca fascicularis) infected with H37Rv Mycobacterium tuberculosis. Tuberculosis (Edinb) 87:518–525. doi:10.1016/j.tube.2007.06.002
Badell E, Nicolle F, Clark S, Majlessi L, Boudou F, Martino A et al (2009) Protection against tuberculosis induced by oral prime with Mycobacterium bovis BCG and intranasal subunit boost based on the vaccine candidate Ag85B-ESAT-6 does not correlate with circulating IFN-gamma producing T-cells. Vaccine 27:28–37. doi:10.1016/j.vaccine.2008.10.034
Qie YQ, Wang JL, Zhu BD, Xu Y, Wang QZ, Chen JZ et al (2008) Evaluation of a new recombinant BCG which contains mycobacterial antigen ag85B-mpt64(190-198)-mtb8.4 in C57/BL6 mice. Scand J Immunol 67:133–139. doi:10.1111/j.1365-3083.2007.02048.x
Tang C, Yamada H, Shibata K, Maeda N, Yoshida S, Wajjwalku W et al (2008) Efficacy of recombinant bacille Calmette-Guérin vaccine secreting interleukin-15/antigen 85B fusion protein in providing protection against Mycobacterium tuberculosis. J Infect Dis 197:1263–1274. doi:10.1086/586902
Lin C-W, Su I-J, Chang J-R, Chen Y-Y, Lu J-J, Dou H-Y (2012) Recombinant BCG coexpressing Ag85B, CFP10, and interleukin-12 induces multifunctional Th1 and memory T cells in mice. APMIS 120:72–82. doi:10.1111/j.1600-0463.2011.02815.x
Chen Y-Y, Lin C-W, Huang W-F, Chang J-R, Su I-J, Hsu C-H et al (2014) Recombinant bacille Calmette-Guerin coexpressing Ag85b, CFP10, and interleukin-12 elicits effective protection against Mycobacterium tuberculosis. J Microbiol Immunol Infect. doi:10.1016/j.jmii.2014.11.019
Ahsan MJ (2015) Recent advances in the development of vaccines for tuberculosis. Ther Adv Vaccines 3:66–75. doi:10.1177/2051013615593891
Waters WR, Palmer MV, Buddle BM, Vordermeier HM (2012) Bovine tuberculosis vaccine research: historical perspectives and recent advances. Vaccine 30:2611–2622. doi:10.1016/j.vaccine.2012.02.018
Buddle BM, Wedlock DN, Denis M, Vordermeier HM, Hewinson RG (2011) Update on vaccination of cattle and wildlife populations against tuberculosis. Vet Microbiol 151:14–22. doi:10.1016/j.vetmic.2011.02.021
Hu X-D, Yu D-H, Chen S-T, Li S-X, Cai H (2009) A combined DNA vaccine provides protective immunity against Mycobacterium bovis and Brucella abortus in cattle. DNA Cell Biol 28:191–199. doi:10.1089/dna.2008.0790
Vordermeier HM, Villarreal-Ramos B, Cockle PJ, McAulay M, Rhodes SG, Thacker T et al (2009) Viral booster vaccines improve Mycobacterium bovis BCG-induced protection against bovine tuberculosis. Infect Immun 77:3364–3373. doi:10.1128/IAI.00287-09
Wedlock DN, Denis M, Painter GF, Ainge GD, Vordermeier HM, Hewinson RG et al (2008) Enhanced protection against bovine tuberculosis after coadministration of Mycobacterium bovis BCG with a Mycobacterial protein vaccine-adjuvant combination but not after coadministration of adjuvant alone. Clin Vaccine Immunol 15:765–772. doi:10.1128/CVI.00034-08
Wedlock DN, Denis M, Skinner MA, Koach J, de Lisle GW, Vordermeier HM et al (2005) Vaccination of cattle with a CpG oligodeoxynucleotide-formulated mycobacterial protein vaccine and Mycobacterium bovis BCG induces levels of protection against bovine tuberculosis superior to those induced by vaccination with BCG alone. Infect Immun 73:3540–3546. doi:10.1128/IAI.73.6.3540-3546.2005
Rizzi C, Bianco MV, Blanco FC, Soria M, Gravisaco MJ, Montenegro V et al (2012) Vaccination with a BCG strain overexpressing Ag85B protects cattle against Mycobacterium bovis challenge. PLoS One 7, e51396. doi:10.1371/journal.pone.0051396
Horwitz MA, Harth G, Dillon BJ, Maslesa-Galić S (2006) A novel live recombinant mycobacterial vaccine against bovine tuberculosis more potent than BCG. Vaccine 24:1593–1600. doi:10.1016/j.vaccine.2005.10.002
Blanco FC, Bianco MV, Garbaccio S, Meikle V, Gravisaco MJ, Montenegro V et al (2013) Mycobacterium bovis Δmce2 double deletion mutant protects cattle against challenge with virulent M. bovis. Tuberculosis (Edinb) 93:363–372. doi:10.1016/j.tube.2013.02.004
Khatri B, Whelan A, Clifford D, Petrera A, Sander P, Vordermeier HM (2014) BCG Δzmp1 vaccine induces enhanced antigen specific immune responses in cattle. Vaccine 32:779–784. doi:10.1016/j.vaccine.2013.12.055
García E, Bianco MV, Gravisaco MJ, Rocha RV, Blanco FC, Bigi F (2015) Evaluation of Mycobacterium bovis double knockout mce2-phoP as candidate vaccine against bovine tuberculosis. Tuberculosis (Edinb) 95:186–189. doi:10.1016/j.tube.2015.01.001
Tompkins DM, Ramsey DSL, Cross ML, Aldwell FE, de Lisle GW, Buddle BM (2009) Oral vaccination reduces the incidence of tuberculosis in free-living brushtail possums. Proc Biol Sci 276:2987–2995. doi:10.1098/rspb.2009.0414
Murphy D, Costello E, Aldwell FE, Lesellier S, Chambers MA, Fitzsimons T et al (2014) Oral vaccination of badgers (Meles meles) against tuberculosis: comparison of the protection generated by BCG vaccine strains Pasteur and Danish. Vet J 200:362–367. doi:10.1016/j.tvjl.2014.02.031
Yeremeev VV, Stewart GR, Neyrolles O, Skrabal K, Avdienko VG, Apt AS et al (2000) Deletion of the 19kDa antigen does not alter the protective efficacy of BCG. Tuber Lung Dis 80:243–247. doi:10.1054/tuld.2000.0252
Rao V, Dhar N, Shakila H, Singh R, Khera A, Jain R et al (2005) Increased expression of Mycobacterium tuberculosis 19 kDa lipoprotein obliterates the protective efficacy of BCG by polarizing host immune responses to the Th2 subtype. Scand J Immunol 61:410–417. doi:10.1111/j.1365-3083.2005.01569.x
Redelman-Sidi G, Glickman MS, Bochner BH (2014) The mechanism of action of BCG therapy for bladder cancer—a current perspective. Nat Rev Urol 11:153–162. doi:10.1038/nrurol.2014.15
Streit JA, Recker TJ, Donelson JE, Wilson ME (2000) BCG expressing LCR1 of Leishmania chagasi induces protective immunity in susceptible mice. Exp Parasitol 94:33–41. doi:10.1006/expr.1999.4459
Varaldo PB, Leite LCC, Dias WO, Miyaji EN, Torres FIG, Gebara VC et al (2004) Recombinant Mycobacterium bovis BCG expressing the Sm14 antigen of Schistosoma mansoni protects mice from cercarial challenge. Infect Immun 72:3336–3343. doi:10.1128/IAI.72.6.3336-3343.2004
Seixas FK, da Silva EF, Hartwig DD, Cerqueira GM, Amaral M, Fagundes MQ et al (2007) Recombinant Mycobacterium bovis BCG expressing the LipL32 antigen of Leptospira interrogans protects hamsters from challenge. Vaccine 26:88–95. doi:10.1016/j.vaccine.2007.10.052
Govan VA, Christensen ND, Berkower C, Jacobs WR Jr, Williamson A-L (2006) Immunisation with recombinant BCG expressing the cottontail rabbit papillomavirus (CRPV) L1 gene provides protection from CRPV challenge. Vaccine 24:2087–2093. doi:10.1016/j.vaccine.2005.11.029
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
None.
Competing interests
The authors have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Singh, V.K., Srivastava, R. & Srivastava, B.S. Manipulation of BCG vaccine: a double-edged sword. Eur J Clin Microbiol Infect Dis 35, 535–543 (2016). https://doi.org/10.1007/s10096-016-2579-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10096-016-2579-y