
Abstract
Background
The percentage of the multidrug resistant Klebsiella pneumoniae clinical isolates is increasing worldwide. The excessive exposure of K. pneumoniae isolates to sublethal concentrations of biocides like benzalkonium chloride (BAC) in health care settings and communities could be one of the causes contributing in the global spread of antibiotic resistance.
Results
We collected 50 K. pneumoniae isolates and these isolates were daily exposed to gradually increasing sublethal concentrations of BAC. The consequence of adaptation to BAC on the cell surface hydrophobicity (CSH) and biofilm formation of K. pneumoniae isolates was explored. Remarkably, 16% of the tested isolates showed an increase in the cell surface hydrophobicity and 26% displayed an enhanced biofilm formation. To evaluate whether the influence of BAC adaptation on the biofilm formation was demonstrated at the transcriptional level, the RT-PCR was employed. Noteworthy, we found that 60% of the tested isolates exhibited an overexpression of the biofilm gene (bssS). After sequencing of this gene in K. pneumoniae isolates before and after BAC adaptation and performing pairwise alignment, 100% identity was detected; a finding that means the absence of mutation after adaptation to BAC.
Conclusion
This study suggests that the widespread and increased use of biocides like BAC at sublethal concentrations has led to an increase biofilm formation by K. pneumoniae isolates. Enhanced biofilm formation could result in treatment failure of the infections generated by this pathogen.
Similar content being viewed by others


Background
The bacterial resistance to antibiotics was first revealed in 1940s. Nevertheless, this problem has continued to progress and evolve as one decade passed into the other [1]. In recent times, the multidrug-resistant bacterial pathogens have become much more common, and they are causing many problems in the treatment of bacterial infections [2]. It is established that the increased spread of resistance among bacterial pathogens has been largely attributed to the improper usage of antimicrobials in both hospitals and communities. Moreover, failure among the healthcare providers to comply with the infection control precautions is an important cause for the dissemination of resistance [3]. Currently, Klebsiella pneumoniae is an important pathogen, being the causative agent of about 14–20% of the infections in respiratory tract, lower biliary duct, surgical wounds, and urinary tract [4]. This opportunistic pathogen can form biofilm as one of its main virulence factors, giving it the ability to attach to both biotic and abiotic surfaces, which contributes to antibiotic resistance [5]. The contribution of the biofilm in bacterial resistance is due to the exopolymeric matrix of biofilm which acts as a diffusion barrier for the penetration of various antibiotics and the physiological differences between planktonic and sessile bacterial cells [6]. Biocides including antiseptics, disinfectants and preservatives have been used in both clinical and domestic applications for more than half a century. At the present time, the use of different biocides is questioned, as the chronic exposure to these agents could have several harmful effects on the human health and could select for less susceptible isolates towards antibiotics and biocides [7, 8]. Quaternary ammonium compounds like benzalkonium chloride (BAC) are commonly used as preservatives in different cosmetics and pharmaceuticals and disinfectants in medical and food-processing environments [9]. In the current work, we investigated the impact of the repeated exposure of K. pneumoniae clinical isolates to sublethal concentrations of BAC on the cell surface hydrophobicity (CSH) and biofilm formation.
Methods
Bacterial isolates
A total of 50 K. pneumoniae isolates were collected from the different departments of [Tanta University] Hospital. The clinical isolates were subjected to both microscopical examination and standard biochemical tests [10]. Klebsiella pneumoniae ATCC 13883 was used as a reference strain.
Chemicals
All the utilized chemicals in this study were of pure analytical grade and they were purchased from Sigma-Aldrich, USA.
Determination of MIC and MBC of BAC
The minimum inhibitory concentration (MIC) of BAC was identified by broth microdilution method in Muller-Hinton broth (MHB) (Oxoid, UK) in 96-well microtitration plates [11]. MIC is defined as the lowest antimicrobial concentration that inhibits bacterial growth indicated by the absence of turbidity in the wells compared to both the growth control and the uninoculated wells. The wells that showed inhibition of growth were subcultured on MH agar (MHA) (Oxoid, UK) for identification of the MBC (minimum bactericidal concentration) which is the lowest concentration of BAC that kills bacteria after overnight incubation at 37 °C. All these determinations were conducted in independent triplicates. As described by Moen et al. [12], bacterial isolates that show MIC > 12 μg/ml were estimated to be tolerant to BAC.
Adaptation to BAC
It was accomplished by daily exposing the tested isolates to sublethal concentrations of BAC with a gradual increase [13] beginning with a concentration of 0.5 × MBC of BAC. When the growth was detected, a 10-fold diluted culture was transferred to fresh MHB supplemented with a slightly higher concentration of BAC. This step was further continued until no growth was noticed after overnight incubation at 37 °C. Afterward, the bacterial suspension from the last tube that showed growth was inoculated to MHA and incubated overnight at 37 °C for growth conformation. All isolates were tested in triplicate.
Determination of the cell surface hydrophobicity (CSH)
It was identified before and after BAC adaptation as described by El-Banna et al. [14]. Briefly, after centrifugation of the bacterial suspensions and collecting the pellets, they were resuspended in saline and transferred to MHB, then they were incubated for only 1 h at 37 °C. The bacterial suspensions then were centrifuged and the pellets were resuspended in the phosphate urea magnesium sulfate buffer (PUM buffer, pH 6.9). Different volumes of n-hexane, ranging from 0.3 to 1.8 ml, were added to 4.8 ml of bacterial suspension in PUM buffer, thoroughly mixed for 2 min, and after complete phase partition, the aqueous phase was taken and its absorbance was measured at 540 nm. The hydrophobicity index (HI) was calculated using the following formula:
Impact of adaptation on biofilm formation using crystal violet assay
Biofilm formation of the tested isolates before and after BAC adaptation was assessed as described by Yang et al. [15]. After overnight incubation of the tested bacteria, its OD600 was adjusted at 1.0, and they were diluted using sterile brain heart infusion (BHI) broth (Oxoid, UK) to a ratio of 1:100. Then, 100 μL of each bacterial suspension was seeded in 96-well microtitration plates, utilizing 3 wells for each isolate, and the wells that contained BHI broth alone were the negative control. After 24 and 48 h of incubation at 37 °C, the wells were washed 3 times with water. The formed biofilms were then fixed with 100 μL of methanol and left for 20 min and air-dried for 1 h. The fixed bacteria were stained with crystal violet solution and left for 10 min, then washed 5 times with water and air-dried. Finally, the dye was solubilized using 100 μl acetic acid solution and OD490 was measured by ELISA AutoReader (Sunrise Tecan, Austria).
The tested isolates were classified into four groups based on their ability to form a biofilm (their ODs) as follows:
-
a-
NBP: not biofilm producer (ODc < OD < 2 ODc),
-
b-
WBP: weak biofilm producer (2 ODc < OD < 4 ODc),
-
c-
MBP: moderate biofilm producer (4 ODc < OD < 6 ODc) and
-
d-
SBP: strong biofilm producer (6 ODc < OD).
The cut-off OD (ODc) of the microtitration plate is three standard deviations above the mean OD of the negative control.
Determination of the viability of microbial cells inhabiting the biofilm population
The MTT (3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide) method that is based on the reduction of a tetrazolium salt was accomplished using the method previously described [16]. In brief, the bacterial suspension was incubated in 96-well microtitration plates and the free-floating cells were removed. About 150 μl of phosphate-buffered saline (PBS) containing 50 μl of MTT solution, at a concentration of 0.3%, were added to each well and incubated for 2 h at 37°C. The MTT solution was then withdrawn from the wells and 150 μl of dimethyl sulphoxide (DMSO) was added and incubated at room temperature for 15 min. The absorbance of the resulting solution was measured using ELISA AutoReader at a wavelength of 550 nm.
RT-PCR
Detection of the relative gene expression of the biofilm specific gene using Rotor-Gene Q 5 plex apparatus (Qiagen, Germany) was accomplished before and after adaptation of 10 K. pneumoniae isolates to BAC utilizing the Rotor-Gene Q software. The gene gapA was utilized as the housekeeping gene [17]. Total RNA was extracted according to the protocol of the manufacturer of Purelink® RNA Mini Kit (Thermo Scientific, USA). RNA was then converted into cDNA using the power cDNA synthesis kit (first-strand cDNA synthesis) (iNtRON Biotechnology, Korea). The amplification of the biofilm gene (bssS) was carried out using Power SYBR® Green master mix (Thermo SCIENTIFIC, USA). Primers designed as previously described by Hassan et al. [18] with the following sequences: the forward primer 5′-GATTCAATTTTGGCGATTCCTGC-3′ and the reverse primer 5′-TAATGAAGTCATTCAGACTCATCC-3′. Relative gene expression was calculated by the 2−ΔΔCt method [19], and the isolates before BAC adaptation were utilized as control samples. When there was an increase of 2-fold or more in comparison with that of the control samples, overexpression was designated (Fernández-Cuenca et al. 2015). All the experiments were performed in triplicate and the result values were expressed as mean ± SD.
PCR and nucleotide sequencing
The genomic DNA was extracted using the GeneJET Genomic DNA Extraction Kit following the manufacturer’s protocol (Thermo Scientific, USA). Amplification using the forward and reverse primers of the biofilm gene bssS (mentioned before) was performed in a total volume of 25 μl containing 1 μl DNA extract, 1 μl of forward primer (10 μM), 1 μl of reverse primer (10 μM), 12.5 μl of Dream green PCR master mix (Thermo Scientific, USA) and 9.5 μl of nuclease-free water. The thermocycler (Thermo Scientific, USA) was programmed with the following conditions including initial denaturation at 94 °C for 2 min, followed by 35 cycles of denaturation at 94 °C for 40 s, annealing at 48 °C for 1 min, extension at 72 °C for 1 min, and final cycle of amplification at 72 °C for 5 min. For the conformation of production of PCR product, the generated amplicon (225 bp) was visualized on 1.5% agarose gel electrophoresis stained with ethidium bromide and illuminated under UV transilluminator (Kowell, Spain). The PCR amplicons were purified using the PCR Purification Kit (PP-201XS; Jena Bioscience, Germany) according to the manufacturer’s instructions. The purified PCR products were then sent to LGC Company (Germany) to be sequenced in the forward direction using ABI 3730XL DNA sequencer (Thermo SCIENTIFIC, USA).
Bioinformatic analysis
The sequences were analyzed using the Chromas 2.6.5 program (https://technelysium.com.au/wp/chromas/) and the identity of the sequenced PCR products was examined using Blast search against GenBank database (http://blast.ncbi.nlm.nih.gov/Blast.cgi). The alignments and assembly of the sequences were performed using Jalview 2.10.2 software.
Statistical analysis
Independent repeating of experiments (at least three times) was carried out in order to achieve reproducibility. The expression of the data was in the form of mean and standard deviation. One-way ANOVA was utilized to detect the significant differences among the tested groups (p < 0.05) using IBM SPSS (17.0, IBM, USA).
Results
In the current study, 50 K. pneumoniae isolates were collected from various clinical samples including wound (13; 26%), urine (11 isolates; 22%), sputum (19 isolates; 38%), and blood (7 isolates; 14%).
Cell surface hydrophobicity
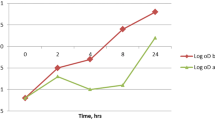
After partitioning of the bacterial cells between the aqueous and organic phases, HI was calculated after measuring the absorbance of the aqueous phase at 540 nm. It was found that 16% of K. pneumoniae isolates exhibited a significant increase in HI after BAC adaptation as shown in Fig. 1.

Chart showing the significant increase in HI in adapted 8 K. pneumonia isolates (16% of isolates)
Influence of BAC adaptation on biofilm formation
The percentage of biofilm-forming K. pneumoniae isolates (MBP plus SBP) increased from 30 to 56% after adaptation to BAC as shown in Table 1. Besides, the viability of K. pneumoniae isolates inhabiting the biofilm population was tested using MTT tetrazolium assay before and after BAC adaptation. It was found that there was non-significant change (p > 0.05) in the viability of the bacterial cells after BAC adaptation.
RT-PCR
This test was performed for more comprehension of the influence of BAC adaptation on the ability of K. pneumoniae isolates form a biofilm. The biofilm gene expression (bssS) was measured in the adapted cells relative to the corresponding non-adapted cells using RT-PCR in 10 K. pneumoniae isolates. As shown in Table 2, 60% of the tested isolates (6 out of 10 isolates) exhibited overexpression of the biofilm gene after adaptation to BAC (i.e., > 2-fold increase in gene expression).
DNA sequencing and alignments of the biofilm gene (bssS)
The sequences of the biofilm gene (bssS) of 6 K. pneumoniae isolates (that showed an increase in their gene expression by RT-PCR) were submitted to GenBank database and assigned the following accession numbers: MH727630, MH727631, MH727632, MH727633, MH727634, and MH727635. Data analysis of these sequences using BLAST showed 98% identity of these nucleotide sequences with K. pneumoniae biofilm gene (bssS). After performing pairwise alignment of each one of these sequences with the sequence of its corresponding non-adapted isolates using Jalview software, they all showed 100% identity.
Discussion
The high percentage of multidrug-resistant K. pneumoniae isolates is a worldwide problem because these pathogens are considered as a major threat to cause both hospital-acquired and community-acquired infections [19, 20]. Hence, the factors which could contribute to the dissemination of resistance among K. pneumoniae clinical isolates should be extensively elucidated and studied in order to be prevented as much as possible The frequent exposure of K. pneumoniae isolates to different biocides at sublethal concentrations in community and health care settings could be one of these factors. Therefore, the objectives of this study were to comprehend and evaluate the impact of adaptation of K. pneumoniae isolates to BAC on its ability to form biofilm as this can have a significant role in the development and proliferation of antibiotic resistance among these isolates.
Biofilm is a matrix formed from extracellular polymeric substances such as exopolysaccharides, nucleic acids, and proteins. It can slow down the growth of bacterial cells and it can form a strong mechanical barrier against the penetration of antimicrobials leading to the development of resistance [21]. In the current research, both CSH and biofilm formation of K. pneumoniae isolates were tested before and after BAC adaptation. As a response to adaptation, the cellular surfaces of bacteria may undergo certain chemical modifications in order to repel the water-soluble compounds resulting in a change in its CSH [22,23,24]. It is well recognized that CSH plays an important role in bacterial adherence to different surfaces [22, 23]. Adhesion of bacterial cells is the first step in biofilm formation [20]. Hydrophobicity index is used as a measure for CSH and it was found that 16% of K. pneumoniae isolates showed a significant increase in HI after BAC adaptation. Nevertheless, CSH is not the sole factor affecting biofilm formation and there are a large number of other factors that have been suggested to affect biofilm formation, such as the hydrodynamic forces, nutrient supply, surface charges, and the existence of other adhesion proteins [24].
The semi-quantitative measurement of the mature biofilms which are produced by K. pneumoniae isolates using crystal violet assay revealed that 26% of the tested isolates became moderately and strongly biofilm producers after BAC adaptation. This phenomenon could lead to many detrimental impacts as biofilm formation is strongly believed to have a major role in the survival and pathogenicity of K. pneumoniae isolates [25]. This is similar to the finding of the research paper conducted by Houari and Di Martino [26] who observed that the adaptation of K. pneumoniae and Escherichia coli isolates to BAC has increased in the bacterial ability to form a biofilm. Moreover, other researchers found that adaptation to biocides has increased the biofilm formation in Escherichia coli, Proteus mirabilis and Staphylococcus aureus isolates [27,28,29]. The MTT assay was utilized in the present study in order to test the viability of the cells embedded in the biofilm before and after BAC adaptation. Our results showed that there was non-significant change in the viability of the bacterial cells after adaptation to BAC. We decided to choose the MTT test in this work as a method to test the bacterial viability instead of their enumeration via serial dilution agar plate technique as the MTT method is faster and more convenient than the traditional methods [30]. The influence of adaptation to BAC on biofilm formation was further investigated by RT-PCR. We noticed that the repeated exposure to sublethal concentrations of BAC has resulted in an increase in the expression of the biofilm gene (bssS) in 60% of the tested isolates. Additionally, sequence analysis of the biofilm amplicon from the tested isolate before and after adaptation to BAC showed no variation in the sequence after pairwise alignment (100% identity) which indicates the absence of mutation in the biofilm gene (bssS) after adaptation; however, this needs further investigations.
Conclusions
We have presented that the repeated exposure of K. pneumonia isolates to sublethal doses of BAC has resulted in a relative increase in the cell surface hydrophobicity and biofilm formation. A factor that could have a role in the emergence and dissemination of resistance to numerous antimicrobials. The findings of our study highly recommend that the underlying hazards connected with the prolonged exposure to sublethal concentrations of biocides like BAC should be of a great focus in hospital and community sanitation. As the observed phenomenon may have vast clinical implications concerning bacterial resistance to antimicrobial chemotherapy and subsequently worse prognosis of different infectious diseases. Accordingly, much more efforts are required to diminish and reduce the increasing usage of biocides, and to develop novel biological systems able to efficiently degrade such agents from the environment.
Availability of data and materials
Please contact corresponding author for data requests.
Abbreviations
- BAC:
-
Benzalkonium chloride
- BHI:
-
Brain heart infusion
- CSH:
-
Cell surface hydrophobicity
- DMSO:
-
Dimethyl sulphoxide
- HI:
-
Hydrophobicity index
- MBP:
-
Moderate biofilm producer
- MBC:
-
Minimum bactericidal concentration
- MIC:
-
Minimum inhibitory concentration
- MHA:
-
Muller-Hinton agar
- MHB:
-
Muller-Hinton broth
- NBP:
-
Not biofilm producer
- OD:
-
Optical density
- PBS:
-
Phosphate buffered saline
- PUM:
-
Phosphate urea magnesium sulphate buffer
- SBP:
-
Strong biofilm producer
- WBP:
-
Weak biofilm producer
References
Abdelaziz A, El Banna T, Sonbol F, Gamaledin N, Maghraby G (2014) Optimization of niosomes for enhanced antibacterial activity and reduced bacterial resistance: in vitro and in vivo evaluation. Exp Opin Drug Deliv 12:163–180
Sonbol F, Elbanna T, Abdelaziz A, Elekhnawy E (2018) Biological characters of vancomycin resistant Staphylococcus aureus isolates from a university hospital in Egypt. Arch Clin Microb 9:1–6
El Banna T, Sonbol F, Abd El Aziz A, Elekhnawy E (2018) Prevalence and antimicrobial susceptibility of vancomycin resistant staphylococci in an Egyptian university hospital. J Med Microb Diag 7:1–8
De Rosa FG, Corcione S, Cavallo R, Di Perri G, Bassetti M (2015) Critical issues for Klebsiella pneumoniae KPC-carbapenemase producing K. pneumoniae infections: a critical agenda. Futur Microbiol 10(2):283–294. https://doi.org/10.2217/fmb.14.121
Seifi K, Kazemian H, Heidari H, Rezagholizadeh F, Saee Y, Shirvani F (2016) Evaluation of biofilm formation among Klebsiella pneumoniae isolates and molecular characterization by ERIC-PCR. Jundishapur J Microbiol 9:e30682
Ebrahimi A, Hemati M, Shabanpour Z, Dehkordi S, Bahadoran S, Lotfalian S (2015) Effects of benzalkonium chloride on planktonic growth and biofilm formation by animal bacterial pathogens. Jundishapur J Microbiol 8:e16058
Sonbol F, Elbanna T, Abdelaziz A, Elekhnawy E (2018) Impact of triclosan adaptation on membrane properties, efflux and antimicrobial resistance of Escherichia coli clinical isolates. J Appl Microbiol 126:730–739
Houari A, Di Martino P (2007) Effect of chlorhexidine and benzalkonium chloride on bacterial biofilm formation. Lett App Microbiol 45(6):652–656. https://doi.org/10.1111/j.1472-765X.2007.02249.x
Gerba CP (2015) Quaternary ammonium biocides: efficacy in application. App Environ Microbiol 81:464–469
MacFaddin JF (1976) Biochemical tests for identification of medical bacteria. Williams & Wilkins Co., Baltimore
Moen B, Rudi K, Bore E, Langsrud S (2012) Subminimal inhibitory concentrations of the disinfectant benzalkonium chloride select for a tolerant subpopulation of Escherichia coli with inheritable characteristics. Int J Mol Sci 13(4):4101–4123. https://doi.org/10.3390/ijms13044101
Soumet C, Meheust D, Pissavin M, Le Grandois P, Fremaux B, Feurer M (2016) Reduced susceptibilities to biocides and resistance to antibiotics in food-associated bacteria following exposure to quaternary ammonium compounds. J Appl Microbiol 121(5):1275–1281. https://doi.org/10.1111/jam.13247
El-Banna T, Abdelaziz A, Sonbol F, El-Ekhnawy E (2019) Adaptation of Pseudomonas aeruginosa clinical isolates to benzalkonium chloride retards its growth and enhances biofilm production. Mol Biol Rep 46(3):3437–3443. https://doi.org/10.1007/s11033-019-04806-7
Yang B, Lei Z, Zhao Y, Ahmed S, Wang C, Zhang S (2017) Combination susceptibility testing of common antimicrobials in vitro and the effects of sub-MIC of antimicrobials on Staphylococcus aureus biofilm formation. Front Microbiol 8:2125. https://doi.org/10.3389/fmicb.2017.02125
Trafny EA, Lewandowski R, Zawistowska-Marciniak I, Stepińska M (2013) Use of MTT assay for determination of the biofilm formation capacity of microorganisms in metal working fluids. World J Microbiol Biotechnol 29(9):1635–1643. https://doi.org/10.1007/s11274-013-1326-0
Huguet A, Pensec J, Soumet C (2013) Resistance in Escherichia coli: variable contribution of efflux pumps with respect to different fluoroquinolones. J App Microbiol 114(5):1294–1299. https://doi.org/10.1111/jam.12156
Hassan R, El-Naggar W, El-Sawy E, El-Mahdy A (2011) Characterization of some virulence factors associated with Enterbacteriaceae isolated from urinary tract infections in Mansoura hospitals. Egypt J Med Microbiol 20:9–17
Fernández-Cuenca F, Tomás M, Caballero-Moyano F, Bou G, Martínez L, Vila J (2015) Reduced susceptibility to biocides in Acinetobacter baumannii: association with resistance to antimicrobials, epidemiological behaviour, biological cost and effect on the expression of genes encoding porins and efflux pumps. J Antimicrob Chemother 70(12):3222–3229. https://doi.org/10.1093/jac/dkv262
Abdelaziz A, Sonbol F, Elbanna T, El-Ekhnawy E (2019) Exposure to sublethal concentrations of benzalkonium chloride induces antimicrobial resistance and cellular changes in Klebsiellae pneumoniae clinical isolates. Microb Drug Resist 25(5):631–638. https://doi.org/10.1089/mdr.2018.0235
Gallardo-Moreno AM, González-Martín ML, Pérez-Giraldo C, Garduno E, Bruque JM, Gómez-García AC (2002) Thermodynamic analysis of growth temperature dependence in the adhesion of Candida parapsilosis to polystyrene. Appl Environ Microbiol 68(5):2610–2613. https://doi.org/10.1128/aem.68.5.2610-2613.2002
Rodrigues DF, Elimelech M (2009) Role of type 1 fimbriae and mannose in the development of Escherichia coli K12 biofilm: from initial cell adhesion to biofilm formation. Biofouling 25(5):401–411. https://doi.org/10.1080/08927010902833443
Heilmann C (2011) Adhesion mechanisms of staphylococci. Adv Exp Med Biol 715:105–123. https://doi.org/10.1007/978-94-007-0940-9_7
Andersson S, Kuttuva G, Carl R, Land J, Dalhammar G (2008) Biofilm formation and interactions of bacterial strains found in wastewater treatment systems. FEMS Microbiol Lett 283(1):83–90. https://doi.org/10.1111/j.1574-6968.2008.01149.x
Krasowska A, Sigler K (2014) How microorganisms use hydrophobicity and what does this mean for human needs? Front Cell Infect Microbiol 4:112
Beceiro A, Tomás M, Bou G (2013) Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clin Microbiol Rev 26(2):185–230. https://doi.org/10.1128/CMR.00059-12
Moretro T, Hermansen L, Holck AL, Sidhu MS, Rudi K, Langsrud S (2003) Biofilm formation and the presence of the intercellular adhesion locus ica among staphylococci from food and food processing environments. Appl Environ Microbiol 69(9):5648–5655. https://doi.org/10.1128/AEM.69.9.5648-5655.2003
Pagedar A, Singh J, Batish VK (2012) Adaptation to benzalkonium chloride and ciprofloxacin affects biofilm formation potential, efflux pump and haemolysin activity of Escherichia coli of dairy origin. J Dairy Re 79(4):383–389. https://doi.org/10.1017/S0022029912000295
Wang H, Cheng H, Wang F, Wei D, Wang X (2010) An improved 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay for evaluating the viability of Escherichia coli cells. J Microbiol Methods 82(3):330–333. https://doi.org/10.1016/j.mimet.2010.06.014
Elekhnawy E, Sonbol F, Abdelaziz A, Elbanna T (2021) An investigation of the impact of triclosan adaptation on Proteus mirabilis clinical isolates from an Egyptian university hospital. Braz J Microbiol. Under press. https://doi.org/10.1007/s42770-021-00485-4
Grela E, Ząbek A, Grabowiecka A (2015) Interferences in the optimization of the MTT assay for viability estimation of Proteus mirabilis. Avicenna J Med Biotech 7:159–167
Acknowledgements
The authors would like to thank the central laboratory of Tanta University for the technical support.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
E.E. carried out the experiment. E.E and F.S wrote the manuscript. T.E and A.A conceived the original idea. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
We declare that this manuscript is only submitted to Egyptian Journal of Medical Human Genetics and it is not currently under consideration elsewhere.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Elekhnawy, E.A., Sonbol, F.I., Elbanna, T.E. et al. Evaluation of the impact of adaptation of Klebsiella pneumoniae clinical isolates to benzalkonium chloride on biofilm formation. Egypt J Med Hum Genet 22, 51 (2021). https://doi.org/10.1186/s43042-021-00170-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43042-021-00170-z