
Abstract
Background
Entomopathogenic nematodes of the genera Heterorhabditis and Steinernema are widely used as biocontrol agents against diverse insect pests. Their association with symbiont bacteria is found to be the primary cause of insect mortality. The present study reported a comprehensive characterization of a local Philippine EPN isolate using congruent morpho-taxometrical and molecular data and the associated bacterium in the infective dauer juveniles.
Results
Using an insect-baiting technique, a nematode isolate labeled F2H was recovered from the sandy soils in the coastal area of southern Philippines. Based on morphology and morpho-taxometrics, the F2H isolate collected from Iligan City was identified as Heterorhabditis indica. Analysis of the ITS region of rDNA revealed the highest sequence match to H. indica. The sequences of the bacterial 16S rDNA gene showed that the symbiont of H. indica presented 100% similitude with the sequences of Ochrobactrum anthropi. Further, pairwise alignment and phylogenetic analysis demonstrated that O. anthropi, O. lupini and O. cytisi shared 100% similarity and form a monophyletic clade. These strains have not presented any differences in the phylogenetic and 16S rDNA data, and O. lupini and O. cytisi should be considered a later heterotypic synonym of O. anthropi.
Conclusions
The description of H. indica-O. anthropi association was the first report in the Philippines. This provides additional account and collection of naturally isolated EPN-bacteria, contributing to limited knowledge on its diversity in the Philippines, which can be utilized in the biocontrol of pests after further comprehensive assessments.
Similar content being viewed by others

Background
The increasing need to reduce chemical pesticide use has driven the interest in utilizing entomopathogenic nematodes (EPNs) as biological control agents (BCA) against economic agricultural pests (Georgis et al. 2006). Unlike chemicals, EPNs are target-specific with a wide range, ecofriendly, and most notably, user safe (Ehlers 2003). Their effectiveness and success as BCAs depend on their adaptability to the environment. Local species or strains are of particular interest due to better adaptation to abiotic factors, local climate and other regulators (Campos-Herrera et al. 2012). The most successful EPN strains belong to the family of Heterorhabditidae. The genus Heterorhabditis (Nematoda: Rhabditida) has a worldwide natural occurrence with currently 19 valid species throughout the globe (Machado et al. 2021).
The two Heterorhabditis species, H. bacteriophora and H. indica, are suggested to have a global distribution, especially in the tropics and subtropics (Bhat et al. 2020). Heterorhabditis indica was first isolated from southern India as a type strain (Poinar et al. 1992) and later on in many other parts of the world with the most recent record in Fiji Islands (Kour et al. 2020) and in eastern Australia (Aryal et al. 2022). In the Philippines, new populations of H. indica from banana, rice (Navarez et al. 2021), peanut (Dichusa et al. 2021) and sugarcane fields (Ramos et al. 2022) were recently documented from Davao del Sur and Cotabato provinces in Mindanao island.
Morphological discrimination between EPN species starts with differences in cadaver coloration to morphometrics. However, Heterorhabditis have been shown to exhibit slight morphological differences between species (Dolinski et al. 2008), making species identification extremely difficult without the aid of a molecular technique (Darissa and Iraki 2014). Molecular characterization was done by the analysis of internal transcribed spacer (ITS) rDNA sequences, among many others (Nasmith et al. 1996). This gene is considered as informative marker for species identification, barcoding and phylogeographical studies of Heterorhabditis species (Dhakal et al. 2020). Even so, false assignment of sequences to a taxon can occur, creating risks in the genetic database. Thus, for an increased resolution and reliability, combining morphological, morpho-taxometrical, molecular and phylogenetic studies are suggested (Spiridonov 2017).
The EPNs form a mutualistic complex with specific symbiotic bacteria, of which the latter is the primary cause of insect mortality (Ciche 2007). They depend on the EPNs for penetration into insect hosts through natural openings while bacterial cells are carried in the infective juveniles’ (IJs) gut. Once inside the host, they are released into the hemolymph, producing virulence factors like the toxin complex a (Tca) among many others produced by Photorhabdus (Proteobacteria: Morganellaceae) symbionts, killing the insect by septicemia within 48 h. (Waterfield et al. 2009). The bacteria then degrade host tissues, supplying nutrients for the growth and development of the nematode population (Hazir et al. 2003). It is known that symbiotic bacteria also produce compounds to antagonize competitors like other bacterial species, fungi, and scavengers that may disturb the nematode development (Waterfield et al. 2009). However, several non-symbiotic bacteria have also been isolated and reported in association with EPN species, for instance, Ochrobactrum anthropi, O. intermedium, and Pseudomonas protegens (Ogier et al. 2020). It was reported recently that the presence of non-symbiotic bacteria is likely to reduce the recycling ability of H. indica in other insect larvae (Upadhyay and Mohan 2021). Despite this, their functions, specific requirements, and relationships are by far still unclear.
Vast natural genetic diversity in EPN populations across the country is yet to be explored and exploited. The collection of local EPNs may bring higher infectivity to target insect pests as a result of better adaptation. In this study, characterizing a local EPN strain collected from Iligan City, as part of the leading agricultural producer in the second largest island in the Philippines, Northern Mindanao was aimed. By employing complementary morphological, morpho-taxometrical, and molecular approaches, a more comprehensive characterization of our local H. indica isolate was conducted, providing an additional account to existing knowledge of EPNs in the country. Furthermore, while symbionts have biocontrol potential, some non-symbiotic bacteria such as Ochrobactrum (Proteobacteria: Brucellaceae) were reported to be opportunistic and pose high risks to humans’ health (Kettaneh et al. 2003). Hence prior to virulence assays and mass production, the non-symbiotic bacteria associated with our EPN species was also identified using the 16S rDNA region (Kämpfer et al. 2003). The outcomes of this study will be pivotal for the future establishment of biocontrol programs in the country.
Methods
Nematode and bacterial isolates
Heterorhabditis indica F2H was isolated from a sandy soil sample collected from a coastal area in Iligan City, Mindanao, Philippines at 15 cm depth (N 8°12′ 5.10″) in January 2019. Soil samples were moistened and placed in plastic containers, with the subsequent addition of five super worm larvae, Zophobas morio (Coleoptera: Tenebrionidae) in each container (Bedding and Akhurst 1975). Containers with the soil and insect baits were incubated in the dark at 25 ± 2 °C. Regular checking of the samples to obtain infected cadavers was conducted every 2–3 days until required. A modified White trap method was used to isolate IJs from cadavers. Dextrose saline solutions (NSS, 0.9% NaCl) were poured into the Petri plate (5 cm diam.) and sealed with parafilm. The pure culture was obtained by re-inoculating EPNs thrice in sand assays and isolating fresh nematodes a new with modified white traps after every infection (Hoy et al. 2008). This monoculture was used for characterization. For the bacterial isolate, Ochrobactrum anthropi F2B was extracted directly from a total of five clean monocultured IJs of H. indica F2H, following the description of Aryal et al. (2022). Briefly, IJs were surface sterilized by soaking in 2% NaOCl for 5 min to avoid secondary contaminations.
Morphological characterization of the nematode
For morpho-taxometrical characterization, IJs were collected directly from the White trap, while adults were obtained through the dissection of cadavers 3–6 days after reinfection. For mounting and fixation of IJs and adults, a modified method originally described by Seinhorst (1959) was followed. A stepwise transfer of heat-killed IJs, hermaphrodite, male and female nematodes to anhydrous glycerol were done. Three following solutions were prepared: Solution 1 (containing 50:50 ratio of formalin (4%) and glycerin), Solution 2 (containing 50:50 ratio of 96% ethanol and 4% formalin) and lastly, Solution 3 (containing pure glycerin). Nematodes were then picked and mounted in clean glass slides with wax rings within a drop of pure glycerin. A maximum of five nematodes were placed on each permanent slide. A Carl Zeiss inverted routine microscope (ZEISS AxioVert.A1) equipped with a camera and AxioVision imaging software were used for studying morphology and measuring the key characters. The isolates were classified using taxonomic keys from Nguyen and Hunt (2007) and Machado et al. (2021).
Molecular characterization of the nematode and bacterium
DNA extraction was performed following the method described by Spiridonov et al. (2004) and using a tissue genomic DNA extraction kit (Dongsheng, Biotech). Briefly, the sterilized IJs were crushed by a sterile scalpel in a 1.5-ml microcentrifuge tube containing buffer, followed by adding 20 µl of Proteinase K. This was incubated at 55 °C until a homogenous solution was obtained. Different solutions provided within the kit were added following the manufacturer’s protocol after each brief vortexing and centrifugation process. After elution, the obtained purified DNA was then kept at − 20 °C in a deep freezer. A fragment of rDNA containing the ITS rDNA was amplified using the primers TW81 (5’-GTTTCCGTAGGTGAACCTGC-3’) as a forward primer and AB28 (5’-ATATGCTTAAGTTCAGCGGGT-3’) as a reverse primer (Joyce et al., 1994). The thermal cycler program for EPNs was as follows: 1 cycle of 94 °C for 7 min, followed by 35 cycles of amplification at 94 °C for 60 s, 54 °C for 60 s and 72 °C for 60 s and a final extension at 72 °C for 10 min.
For the bacterium, the universal primers used to amplify 16S rDNA genes were 27F (5'-AGAGTTTGATCCTGGCTCAG-3') as the forward primer and 1492R (5′-GGTTACCTTGTTACGACTT-3′) as the reverse primer (Frank et al. 2008). The PCR was performed with the following conditions: 1 cycle at 94 °C for 2 min, followed by 35 cycles at 94 °C for 45 s, 55 °C for 60 s, 72 °C for 60 s and a final extension at 72 °C for 10 min. The amplified products were Sanger sequenced by Macrogen, Inc. (Seoul, South Korea). The sequencing results of ITS and 16S rDNA of nematodes and bacteria were submitted in the GenBank under accession numbers, MT023075 and MT012075, respectively.
Phylogenetic analyses of nematode and bacterium
The generated sequences were first edited using Chromas 2.6.6. A nucleotide basic local alignment search tool (BLASTn) was performed in order to compare to all sequences deposited in the National Centre for Biotechnology (NCBI) (Altschul et al. 1997). A cut of ≥ 97% identity was considered for the same nematode species (Bhat et al. 2021) and ≥ 99% identity for bacterial sequence (Wang et al. 2015). ITS and 16S rDNA sequences were aligned with their closest BLAST search matches, using ClustalW multiple alignment. Poorly aligned regions were removed from the alignments using MEGA-7. The base substitution model was evaluated using jModeltest 0.1.1 (Posada 2008). Phylogenetic trees were elaborated using the Bayesian inference method as implemented in the program MrBayes 3.2.7 (Ronquist et al. 2012). The HKY + (gamma distribution of rate variation with a proportion of invariable sites) model was selected. The selected model was initiated with a random starting tree and ran with the Markov chain Monte Carlo (MCMC) for 1 × 107 generations. The Bayesian tree was ultimately visualized using the FigTree program 1.4.4 (Rambaut 2018). The out group taxons used for phylogenetic analysis were Heterorhabditis zealandica in case of Heterorhabtidis, and Xenorhabdus nematophila and Photorhabdus akhurstii in bacteria. In addition, genetic pairwise distances were estimated using MEGA-7 software (Kumar et al. 2016).
Results
Morphological characterization of the nematode
Key morphological characters were identified according to Nguyen and Hunt (Nguyen and Hunt 2007). Infective juveniles (n = 20) and only few males (n = 5) were obtained, fixed and measured in µm. F2H isolate closely matched the original description of H. indica. Infective juveniles were characterized by their body length of 488.5 (441–550) μm, tail length of 95.5 (79.9–105.6) μm, maximum body diameter of 24.2 (18–28) μm and ratio E% of 98.8 (85.9–112.5), and ratio F of 0.3 (0.2–0.4). Males are found to be slightly smaller (642, 585–697 μm) than the type strain (721, 573–788 μm) and the H. indica (872, 765–988 μm) documented from Cebu, Philippines; however, comparable size can be observed with H. indica H.TN48 populations in Vietnam with males size of 689 (596–818) μm. They are characterized by their spicule length (40.8, 36–46 μm), gubernaculum length (21.1, 20.9–24 μm), and ratio SW% (1.8, 1.6–1.9), closely matching other described strains. Females are observed to have protruding vulval lips with transverse slit, located near mid-body and generally smaller compared to other H. indica female populations (data not shown). Comparative morphometrics of IJs and males of the present isolates with previously described H. indica populations are presented in Tables 1 and 2, respectively.
Molecular characterization of the nematode and bacterium
Nucleotide sequence analysis of the ITS rDNA indicated the F2H isolate as H. indica. There were 699 bp for the strain F2H. Upon BLASTn analysis of the ITS rDNA sequences, F2H showed 99.86% similarity with the H. indica strains reported from different geographical regions of the globe. When aligned with ITS rDNA sequences of already known H. indica, it displayed two nucleotide differences with the sequences of H. indica from India (MF973067). The distance matrix analyses based on ITS rDNA sequences showed that the present isolate of H. indica (F2H) is separated from other reported Heterorhabditis nematodes by 10–189 bp (Additional file 1: Table S1).
The sequencing of amplified product of 16S rDNA resulted in the production of a partial sequence of 1220 bp which is the size expected for 16S rDNAs. Using the BLASTn algorithm, the 16S rDNA gene of the present bacterial strain was identified as the most closely related to Ochrobactrum sequences available in the GenBank database. The highest levels of identity were obtained with O. anthropi: the 16S rDNA sequence of isolate F2B had 100% identity with O. anthropi reported from different geographical regions of the world. When aligned with 16 rDNA sequences of already described O. anthropi, it showed zero nucleotide differences and 100 percentage similarity with the sequences of O. anthropi from France (AJ867290), India (EU826071), and other valid descriptions of O. anthropi. Further, the pairwise comparisons between the present O. anthropi from the Philippines and O. lupini from Spain showed a zero total character difference and 100% similarity, while with O. cytisi showed one total character difference and 100 percentage similarity. Additionally, O. anthropi, O. lupini and O. cytisi genomes presented a similar G + C (55.53, 56.52 and 55.56%, respectively), and A + C (44.47, 44.48 and 44.44, respectively) content demonstrating that each of them indeed represents single species. However, with other described species of Ochrobactrum, the present isolate F2B was separated by 18–68 bp (Additional file 1: Table S2).
Phylogenetic analysis of the nematode and bacterium
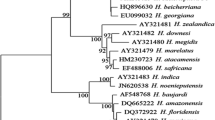
Phylogenetic analyses performed based on ITS1-5.8S-ITS2 rDNA showed that the present Heterorhabditis isolate (F2H) formed a monophyletic clade with Heterorhabditis indica reported from different countries of the world, thus confirming its identification. This monophyletic clade formed a group with closely related species, H. noenieputensis (Fig. 1) and together formed a group with other known members of Indica-clade. Members of Indica-clade formed a sister clade with described species of Bacteriophora- and Megidis-clade (Fig. 1). Thus, based on the phylogeny of ITS rDNA sequences, the F2H Heterorhabditis isolate from the Philippines was conspecific to H. indica.

Phylogenetic relationships of Heterorhabditis indica and other Heterorhabditis species as inferred from Bayesian analysis of sequences of the Internal Transcribed Spacer (ITS1-5.8S-ITS2) rDNA region. Bayesian posterior probabilities (%) equal to or more than 60% are given for appropriate clades. The scale bar shows the number of substitutions per site
Based on the 16S rDNA gene sequences, the bacterial isolate F2B was closely related to Ochrobactrum anthropi and shares 100% sequence similarity. Phylogenetic relationship reconstructions confirmed this observation and suggested that the bacterial isolate F2B belongs to the O. anthropi species (Fig. 2). In the phylogenetic tree of the 16S rDNA genes computed with sequences from all type strains of Ochrobactrum species that known, O. anthropi strains clustered with O. lupini and O. cytisi forming a monophyletic clade. Phylogenetic 16S marker gene sequence extracted from O. anthropi genome present similarity of 99% with O. lupini and 100% with O. cytisi.

Phylogenetic relationships of Ochrobactrum anthropi and other Ochrobactrum species as inferred from Bayesian analysis of sequences of the 16S rDNA region. Bayesian posterior probabilities (%) equal to or more than 60% are given for appropriate clades. The scale bar shows the number of substitutions per site
Discussion
Identification of these microscopic nematodes is often described to be rather difficult and requires meticulous efforts. At low taxonomic levels (i.e., order, family, and genus level), systematics was reported to be unstable. This is mainly due to their scantiness of observable and informative characters as well as overlapping morphometrics (Holterman 2008). H. indica IJs of this study were separated from other Heterorhabditis species by their smaller body length (< 600 μm). The rest of the characters are within the range of the original descriptions of Poinar et al. (1992). Males were slightly small at the type strain but comparable to the H. indica H.TN48 populations in Vietnam (Phan et al. 2003). However, Heterorhabditis exhibited slight morphological differences between species (Dolinski et al. 2008), making species identification extremely difficult (Darissa and Iraki 2014). With the advent of molecular diagnostic tools, morphological data can be complemented with modern techniques (Spiridonov 2017). In the present study, ITS rDNA region was used to identify and confirm the collected Heterorhabditis species.
Of particular highlight in this study was the bacterium detected in the IJs gut. These IJs initiate the parasitic-pathogenic life cycle of any EPNs as they carry bacteria in their guts or vesicle compartments while seeking their target insect hosts (Hazir et al., 2003). Thus, the natural bacteria present were determined in the IJs itself as it was rarely investigated in EPNs. Any phenotypic characterization, physiological, or biochemical profiling were not conducted to augment molecular analysis. Nonetheless, the conventionally used 16S rDNA gene analysis, which is the most used universal marker for identification of prokaryotes (Kämpfer et al. 2003) was used. For the first time, Ochrobactrum anthropi was reported as a non-symbiotic bacterium naturally associated with the IJs of H. indica F2H from the Philippines. Previous reports relating to existence of non-EPN bacterial symbionts are commonly recognized either as laboratory contaminants (Poinar 1966), symbiont’s inability to antagonize other species (Jackson et al. 1995), symbiont’s phase variations (Razia et al. 2011), or ephemeral associations (Gouge and Snyder 2006). Some findings on the natural occurrence of dixenic associations between the Ochrobactrum and Photorhabdus in H. indica were reported specifically inside the infected insect cadaver (Razia et al. 2011). For this reason, the previous idea of general contamination was dismissed as the identified Ochrobactrum bacteria were fresh harvests without laboratory transfer (Babić et al. 2000) and the method of bacterial isolation entailed surface disinfection on the nematodes (Fischer-Le Saux et al. 1998). Furthermore, the pairwise alignment and phylogenetic analysis of the 16S rDNA genes revealed that O. anthropi, O. lupini and O. cytisi shared 100% similarity and form a monophyletic clade. Thus, it may be concluded that O. cytisi status as a separate species needs to be validated with a whole-genome sequencing project for this species-type strain as was clarified for O. lupini by Volpiano et al. (2019)
Associated bacteria are usually isolated and identified from the insect host hemolymph and seldom from the IJ. With the traditional culture-dependent approach, several other bacterial associates from nematodes remain cryptic (Gouge and Snyder 2006). To investigate its community dynamics, a heterorhabditid-infected insect cadaver was profiled for bacterial communities using a culture-independent next-generation sequencing tool. Some studies have been also carried out in the model organism, C. elegans (Dirksen et al. 2016). This recent approach for EPN-microbe interactions revealed a gradual shift from Photorhabdus-dominant community toward one shared with non-symbiotic bacteria in the insect environment (Wollenberg et al. 2016). In order to elucidate findings, Ogier et al. (2020) employed a modern and systematic approach to directly examine for the first time the IJ microbiota of the Steinernema carpocapsae by metabarcoding. The authors found that instead of the standard monoxenic model of the EPN-bacteria complex, it proposes the pathobiome view. Ochrobactrum sp. was among many other bacteria described from Steinernema species. They further reported O. anthropi from H. bacteriophora microbiota, which validates our recent findings on the O. anthropi association with the IJs of H. indica. Although the traditional method was used, nevertheless it was complemented. As a future prospect, the IJ microbiota of the tested H. indica tropical isolates will be investigated.
Interestingly, only Ochrobactrum was identified in this study and neither symbiotic bacterium nor dixenic associations was detected inside the IJs gut. Directly, the bacterial DNA inside the IJ gut was extracted and then identified using the 16S rDNA. The lone detection could be attributed to the methods used or for some other reasons. For example, the possibility of the insensitivity and tolerance of this Ochrobactrum strain against antimicrobials produced by the symbiont (Jackson et al. 1995) or that the antimicrobial activity of the non-symbiotic species may have affected the primary symbiont and contributed to interspecies competition (Ogier et al. 2020). For this, it was assumed that this O. anthropi F2B non-symbiont may have dominated and decimated the symbiotic bacteria, or that they persisted well inside the IJ gut. This kind of sustainable association of bacteria other than the symbiont was demonstrated in the Steinernema-Xenorhabdus model, validating the persistence of non-symbionts like Ochrobactrum (Ogier et al. 2020). In a similar study, Ochrobactrum sp. was found persisting in the nematode gut of an EPN, Oscheius chongmingensis where it was also the most abundant bacterial genus at 59.82%, followed by Bacillus at 7.13% and others (Fu and Liu 2019). Ochrobactrum was further reported to be associated with the microbiome of other nematode species like the free-living cephalob nematode, Acrobeloides maximus (Baquiran et al. 2013), and in the model nematode C. elegans under stressful conditions (Dirksen et al. 2016), even indicated to coexist with several bacteria in nature. In general, Ochrobactrum is abundant in the soil, in the insect cadaver and in the IJ microbiota. As EPNs belong to the group of Rhabditida that feeds on microorganisms, they likely have retained this microbivorous behavior (Boemare et al. 1996), allowing non-symbionts to multiply in the nematode gut. Whether this bacterium is likewise maternally transmitted similar to a previously known elaborate pathway of Heterorhabditis symbionts (Ciche et al. 2008) is subject to further investigations.
The concern further is raised to health hazards. Although recently found not to be virulent on insects, the genus was earlier documented to be opportunistic pathogens to immune-compromised humans such as O. anthropi (Kettaneh et al. 2003). Nematodes then can be a reservoir of this opportunistic bacterium. As for the non-symbionts like Ochrobactrum, which can occur in laboratory culture, Boemare et al. (1996) emphasized that their association was not considered a risk because they cannot support long-term mass rearing of nematode populations. However, based on the documented persistence presented, risk cannot be ruled out. Thus, caution must always be practiced in the future outlook of applying EPN technology to the local agricultural industries for the farmers’ benefit. When applied in the field, several natural associations may occur, but in handling EPN cultures in the laboratory and mass producing EPN-based products, in vitro monoxenic culture in bioreactors is highly recommended. Symbiont bacterial cultures must be isolated and later introduced with axenic nematodes. This enables more control of undesired non-symbionts (Peters et al. 2017) described by two methods: first by surface sterilizing the IJs, and second by sterilizing the eggs to produce aposymbiotic nematodes. Proper axenization and sterile techniques should be employed. Thus, careful practice and preparation of EPNs are needed prior to virulence tests or mass production as a biocontrol agent.
Conclusions
The EPN Heterorhabditis indica F2H, isolated from coastal sandy soils in Iligan, was the first report of occurrence in northern Mindanao, Philippines. This isolate showed a high homology to H. indica isolates reported from different geographical areas of the world, extending the documentation and geographic distribution of this particular species, which may be used as a biocontrol agent against insect pests in the country. The discovery of the natural association to Ochrobactrum anthropi F2B, a non-symbiont bacterium has provided additional evidence that this is present and abundant not only in the soils, but also in nematode microbiota, persisting in the IJs gut. This is the first record in the Philippine archipelago. Since the species is reported to be opportunistic to humans, this study will provide insights into practicing caution during EPN development as biocontrol agents in the country. Handling EPN cultures in the laboratory and their mass production must be done through in vitro monoxenic cultures, an existing approach where eggs are surface sterilized before introduction to its particular symbiotic bacterial culture. Further, full genome sequences are required to confirm the status of some controversial bacterial species.
Availability of data and materials
All sequence data have been deposited with GenBank (NCBI). Other data and material of this manuscript are available upon reasonable request.
Abbreviations
- BCA:
-
Biological control agent
- EPN:
-
Entomopathogenic nematodes
- H. indica :
-
Heterorhabditis indica
- IJs:
-
Infective juveniles
- NCBI:
-
National Center of Biotechnology Information
- O. anthropi :
-
Ochrobactrum anthropic
References
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402. https://doi.org/10.1093/nar/25.17.3389
Aryal S, Nielsen UN, Sumaya NH, De Faveri S, Wilson C, Riegler M (2022) Isolation and molecular characterization of five entomopathogenic nematode species and their bacterial symbionts from eastern Australia. BioControl 67:63–74. https://doi.org/10.1007/s10526-021-10105-7
Babić I, Saux MF, Le Giraud E, Boemare N (2000) Occurrence of natural dixenic associations between the symbiont Photorhabdus luminescens and bacteria related to Ochrobactrum spp. in tropical entomopathogenic Heterophabditis spp. (Nematoda, Rhabditida). Microbiology 146:709–718
Baquiran JP, Thater B, Sedky S, De Ley P, Crowley D, Orwin PM (2013) Culture-independent investigation of the microbiome associated with the nematode Acrobeloides maximus. PLoS One. https://doi.org/10.1371/journal.pone.0067425
Bedding RA, Akhurst RJ (1975) A simple technique for the detection of insect paristic rhabditid nematodes in soil. Nematologica 21:109–110. https://doi.org/10.1163/187529275X00419
Bhat AH, Chaubey AK, Askary TH (2020) Global distribution of entomopathogenic nematodes, Steinernema and Heterorhabditis. Egypt J Biol Pest Control 30:31. https://doi.org/10.1186/s41938-020-0212-y
Bhat AH, Chaubey AK, Hartmann J, Nermut’ J, Půža V, (2021) Notes on the morphology, bionomics, distribution and efficacy of Steinernema siamkayai (Rhabditida: Steinernematidae) from western Uttar Pradesh, India. Nematology 23:1–20. https://doi.org/10.1163/15685411-bja10079
Boemare N, Laumond C, Mauleon H (1996) The entomopathogenic nematode-bacterium complex: biology, life cycle and vertebrate safety. Biocontrol Sci Technol 6:333–346. https://doi.org/10.1080/09583159631316
Campos-Herrera R, Barbercheck M, Hoy CW, Stock SP (2012) Entomopathogenic nematodes as a model system for advancing the frontiers of ecology. J Nematol Soc Nematol 44:162–176
Ciche T (2007) The biology and genome of Heterorhabditis bacteriophora. WormBook. https://doi.org/10.1895/wormbook.1.135.1
Ciche TA, Kim KS, Kaufmann-Daszczuk B, Nguyen KCQ, Hall DH (2008) Cell invasion and matricide during Photorhabdus luminescens transmission by Heterorhabditis bacteriophora nematodes. Appl Environ Microbiol 74:2275–2287. https://doi.org/10.1128/AEM.02646-07
Darissa OM, Iraki NM (2014) Molecular identification of six Steinernema isolates and characterization of their internal transcribed spacers regions. Jordan J Biol Sci 7:31–34. https://doi.org/10.12816/0008210
Dhakal M, Nguyen KB, Hunt DJ, Ehlers RU, Spiridonov SE, Subbotin SA (2020) Molecular identification, phylogeny and phylogeography of the entomopathogenic nematodes of the genus Heterorhabditis Poinar, 1976: a multigene approach. Nematology. https://doi.org/10.1163/15685411-bja10052
Dichusa CA, Ramos R, Aryal S, Sumaya NP, Sumaya NH (2021) Survey and identification of entomopathogenic nematodes in the province of Cotabato, Philippines, for biocontrol potential against the tobacco cutworm, Spodoptera litura (Fabricius)(Lepidoptera: Noctuidae). Egypt J Biol Pest Control 31:1
Dirksen P, Marsh SA, Braker I, Heitland N, Wagner S, Nakad R, Mader S, Petersen C, Kowallik V, Rosenstiel P, Félix MA, Schulenburg H (2016) The native microbiome of the nematode Caenorhabditis elegans: gateway to a new host-microbiome model. BMC Biol 14:38
Dolinski C, Kamitani FL, Machado IR, Winter CE (2008) Molecular and morphological characterization of heterorhabditid entomopathogenic nematodes from the tropical rainforest in Brazil. Mem Inst Oswaldo Cruz 103:150–159
Ehlers RU (2003) Biocontrol Nematodes. In: Hokkanen HMT, Hajek AE (eds) Environmental impacts of microbial insecticides. Progress in Biological Control, vol 1. Springer, Dordrecht. pp. 177–220. https://doi.org/10.1007/978-94-017-1441-9_10
Fischer-Le Saux M, Mauléon H, Constant P, Brunel B, Boemare N (1998) PCR-ribotyping of Xenorhabdus and Photorhabdus isolates from the caribbean region in relation to the taxonomy and geographic distribution of their nematode hosts. Appl Environ Microbiol 64:4246–4254. https://doi.org/10.1128/aem.64.11.4246-4254.1998
Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, Olsen GJ (2008) Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Appl Environ Microbiol 74:2461–2470. https://doi.org/10.1128/AEM.02272-07
Fu J, Liu Q (2019) Evaluation and entomopathogenicity of gut bacteria associated with dauer juveniles of Oscheius chongmingensis (Nematoda: Rhabditidae). Microbiologyopen. https://doi.org/10.1002/mbo3.823
Georgis R, Koppenhöfer AM, Lacey LA, Bélair G, Duncan LW, Grewal PS, Samish M, Tan L, Torr P, van Tol RWHM (2006) Successes and failures in the use of parasitic nematodes for pest control. Biol Control 38:103–123. https://doi.org/10.1016/j.biocontrol.2005.11.005
Gouge DH, Snyder JL (2006) Temporal association of entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) and bacteria. J Invertebr Pathol 91:147–157. https://doi.org/10.1016/j.jip.2005.12.003
Hazir S, Kaya HK, Stock S, Keskin N (2003) Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) for biological control of soil pests. Turkish J Biol 27:181–202
Holterman M (2008) Phylogenetic relationships within the phylum Nematoda as revealed by ribosomal DNA, and their biological implications. Dissertation, Wageningen University
Hoy CW, Grewal PS, Lawrence JL, Jagdale G, Acosta N (2008) Canonical correspondence analysis demonstrates unique soil conditions for entomopathogenic nematode species compared with other free-living nematode species. Biol Control 46:371–379. https://doi.org/10.1016/j.biocontrol.2008.06.001
Jackson TJ, Wang H, Nugent MJ, Griffin CT, Burnell AM, Dowds BCA (1995) Isolation of insect pathogenic bacteria, Providencia rettgeri, from Heterorhabditis spp. J Appl Bacteriol 78:237–244. https://doi.org/10.1111/j.1365-2672.1995.tb05022.x
Joyce SA, Burnell AM, Powers TO (1994) Characterization of Heterorhabditis isolates by PCR amplification of segments of mtDNA and rDNA genes. J Nematol 26:260–270
Kämpfer P, Buczolits S, Albrecht A, Busse HJ, Stackebrandt E (2003) Towards a standardized format for the description of a novel species (of an established genus): Ochrobactrum gallinifaecis sp. nov. Int J Syst Evol Microbiol. https://doi.org/10.1099/ijs.0.02710-0
Kettaneh A, Weill FX, Poilane I, Fain O, Thomas M, Herrmann JL, Hocqueloux L (2003) Septic shock caused by Ochrobactrum anthropi in an otherwise healthy host. J Clin Microbiol 41:1339–1341. https://doi.org/10.1128/JCM.41.3.1339-1341.2003
Kour S, Khurma U, Brodie G, Hazir S (2020) Natural occurrence and distribution of entomopathogenic nematodes (Steinernematidae, Heterorhabditidae) in Viti Levu, Fiji Islands. J Nematol. https://doi.org/10.21307/JOFNEM-2020-017
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874. https://doi.org/10.1093/molbev/msw054
Machado R, Bhat AH, Abolafia J, Muller A, Bruno P, Fallet P, Arce C, Turlings T, Bernal JS, Kajuga J, Waweru B, Toepfer S (2021) Multi-locus phylogenetic analyses uncover species boundaries and reveal the occurrence of two new entomopathogenic nematode species, Heterorhabditis ruandica n. sp. and Heterorhabditis zacatecana n. sp. J Nematol 53:e2021-89. https://doi.org/10.21307/jofnem-2021-089
Nasmith CG, Speranzini D, Jeng R, Hubbes M (1996) RFLP analysis of PCR amplified ITS and 26S ribosomal RNA genes of selected entomopathogenic nematodes (Steinernematidae, Heterorhabditidae). J Nematol 28:15–25
Navarez ML, Sangcopan R, Aryal S, Sumaya NPD, Bhat AH, Sumaya NH (2021) Native Philippine Heterorhabditis indica isolates from banana and rice fields and preliminary results of their virulence against the larvae of super worm (Zophobas morio Fabricius Coleoptera: Tenebrionidae). Egypt J Biol Pest Control 31:46. https://doi.org/10.1186/s41938-021-00388-4
Nguyen K, Hunt DJ (2007) Entomopathogenic nematodes: systematics, phylogeny and bacterial symbionts. Leiden, The Netherlands, Brill. https://doi.org/10.1163/ej.9789004152939.i-816
Ogier JC, Pagès S, Frayssinet M, Gaudriault S (2020) Entomopathogenic nematode-associated microbiota: from monoxenic paradigm to pathobiome. Microbiome 8:25. https://doi.org/10.1186/s40168-020-00800-5
Pascual PR, Pascual ML, Alburo HM (2017) First report of Entomopathogenic nematode Heterorhabditidae (Rhabditida) in organic vegetable farms in Cebu, Philippines. J Agric Manag 20:1–9
Peters A, Han R, Yan X, Leite LG (2017) Production of Entomopathogenic nematodes. In: Lacey L (ed) Microbial control of insect and mite pests: from theory to practice. Elsevier Inc., pp 157–170. https://doi.org/10.1016/B978-0-12-803527-6.00010-X
Phan KL, Subbotin SA, Nguyen NC, Moens M (2003) Heterorhabditis baujardi sp. n. (Rhabditida: Heterorhabditidae) from Vietnam and morphometric data for H. indica populations. Nematology 5:367–382. https://doi.org/10.1163/156854103769224368
Poinar GO (1966) The presence of Achromobacter nematophilus in the infective stage of a Neoaplectana sp. (Steinernematidae: Nematoda). Nematologica 12:105–108. https://doi.org/10.1163/187529266X00068
Poinar GO, Karunakar GK, David H (1992) Heterorhabditis indicus n. Sp. (Rhabditida : Nematoda) from India: separation of Heterorhabditis spp. by infective juveniles. Fundam Appl Nematol 15(5):467–472
Posada D (2008) jModelTest: phylogenetic model averaging. Mol Biol Evol 25:1253–1256. https://doi.org/10.1093/molbev/msn083
Rambaut A (2018) FigTree, a graphical viewer of phylogenetic trees (Version 1.4.4). Available at http://tree.bio.ed.ac.uk/software/figtree.
Ramos R Jr, Dichusa C, Aryal S, Crispolon E Jr, Sumaya NPD, Sumaya NH (2022) Isolation, identification and virulence of the local entomopathogenic nematode Heterorhabditis indica in the southern Philippines against the sugarcane white grub, Leucopholis irrorata (Chevrolat) (Coleoptera: Scarabaeidae). Int J Trop Insect Sci 42(3):2739–2750
Razia M, KarthikRaja R, Padmanaban K, Chellapandi P, Sivaramakrishnan S (2011) 16S rDNA-based phylogeny of non-symbiotic bacteria of entompathogenic nematodes from infected insect cadavers. Genomics Proteomics Bioinforma 9:104–112. https://doi.org/10.1016/S1672-0229(11)60013-2
Ronquist F, Teslenko M, Van Der Mark P, Ayres DL, Darling A, Ohna SH, Larget B, Liu L, Suchard MA, Huelsenbeck JP (2012) Software for systematics and evolution MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst Biol 61:539–542
Seinhorst JW (1959) A rapid method for the transfer of nematodes from fixative to anhydrous glycerin. Nematologica 4:67–69
Spiridonov SE, Reid AP, Podrucka K, Subbotin SA, Moens M (2004) Phylogenetic relationships within the genus Steinernema (Nematoda: Rhabditida) as inferred from analyses of sequences of the ITS1-5.8S-ITS2 region of rDNA and morphological features. Nematology 6:547–566
Spiridonov SE (2017) Entomopathogenic nematodes of the families steinernematidae and heterorhabditidae: morphology and taxonomy, In: Abd-Elgawad MMM, Askary TH, Coupland J (eds) Biocontrol agents: entomopathogenic and slug parasitic nematodes. CABI International, pp 45–62. https://doi.org/10.1079/9781786390004.0045
Upadhyay A, Mohan S (2021) Bacillus subtilis and B. licheniformis isolated from Heterorhabditis indica infected apple root borer (Dorysthenes huegelii) suppresses Nematode production in Galleria mellonella. Acta Parasitol 66(3):989–996
Volpiano CG, Sant’Anna FH, Ambrosini A, Lisboa BB, Vargas LK, Passaglia MPL (2019) Reclassification of Ochrobactrum lupini as a later heterotypic synonym of Ochrobactrum anthropi based on whole-genome sequence analysis. Int J Syst Evol Microbiol 69:2312–2314. https://doi.org/10.1099/ijsem.0.003465
Wang X, King Jordan I, Mayer L (2015) A phylogenetic perspective on molecular epidemiology. In: Tang YW, Sussman M, Liu D, Poxton I, Schwartzman J (eds) Molecular medical microbiology (second edition). Academic Press, Boston, pp 517–536
Waterfield NR, Ciche T, Clarke D (2009) Photorhabdus and a host of hosts. Annu Rev Microbiol 63:557–574
Wollenberg AC, Jagdish T, Slough G, Hoinville ME, Wollenberg MS (2016) Death becomes them: bacterial community dynamics and stilbene antibiotic production in cadavers of Galleria mellonella killed by Heterorhabditis and Photorhabdus spp. Appl Environ Microbiol 82:5824–5837
Acknowledgements
We would like to thank the Flora Biodiversity Laboratory-Nematology Research Team for the technical assistance on this research.
Funding
This research is funded by the Mindanao State University-Iligan Institute of Technology (SO No. 00254-IIT). The work of AHB is supported by Swiss Government Excellence Scholarship (Grant Nr. 2021.0463 to AHB).
Author information
Authors and Affiliations
Contributions
ALL, PN, SA, JGM, and NHS conceptualized and designed the experimental works. ALL carried out the sampling with the support of NHS. ALL, PN, and NHS performed the morphological and morphometric studies. AHB performed the data (sequence alignment and phylogenetic) analyses. NHS was responsible for research funding. ALL wrote the manuscript together with NHS and with the significant input of all other authors. All authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors of this manuscript do not have any kind of conflict of interest that needed disclosure to this journal.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Pairwise distances of the ITS region of the rDNA among species of the “Heterorhabditis”. Table S2. Pairwise distances of the 16S region of the rDNA among species of the “Ochrobactrum”.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Leonar, A.L., Nimkingrat, P., Aryal, S. et al. Natural association of the entomopathogenic nematode Heterorhabditis indica (Rhabditida: Heterorhabditidae) from the Philippines with the non-symbiotic bacterium Ochrobactrum anthropi (Proteobacteria: Brucellaceae). Egypt J Biol Pest Control 32, 83 (2022). https://doi.org/10.1186/s41938-022-00576-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00576-w