
Abstract
Background
Investigations of Culicoides fauna, including inventories, were carried out in Morocco at different periods after the country had faced major bluetongue and African horse sickness outbreaks. However, no comprehensive reference publication has provided a clear overview of the Culicoides species diversity. This study reviewed available data on Culicoides biting midge species in Morocco from 1968 to 2015 (published and grey literature in French and English) in order to revise the current checklist, in light of state of the art taxonomic and systematic knowledge, and confirmed the checklist with morphological and molecular identifications of specimens collected from the region of Rabat.
Methods
Literature related to Culicoides collections in Morocco was collated. Authors were contacted to obtain raw data and additional information for the collections. Fresh Culicoides material was collected and examined from two sites around Rabat, the capital of Morocco. Each collected individual was examined and morphologically identified, if possible, to the species level. In addition, molecular identification was performed to separate closely related species, to confirm difficult morphological identifications and to confirm new species records.
Results
A total of 6121 individuals of Culicoides spp. were collected and analyzed and at least 17 species were identified: C. cataneii/C. gejgelensis, C. circumscriptus, C. fagineus, C. festivipennis, C. imicola, C. jumineri, C. kingi, C. longipennis, C. montanus, C. newsteadi, C. obsoletus, C. paolae, C. parotti, C. puncticollis, C. sahariensis, C. scoticus and C. subfagineus. Seven species were confirmed using phylogenetic analyses. Two new species records for Morocco are reported: C. paolae and C. subfagineus.
Conclusions
The Moroccan fauna of Culicoides now includes 54 valid species. Further work would certainly increase this total, as one of the clades we identified was not affiliated to any described and valid species.
Similar content being viewed by others

Background
Biting midges of the genus Culicoides Latreille, 1809 (Diptera: Ceratopogonidae) are responsible for the transmission of economically important viruses in animal production, most notably African horse sickness virus (AHSV), bluetongue virus (BTV), epizootic hemorrhagic disease virus (EHDV) and the Schmallenberg virus which emerged in 2011 in Europe [1,2,3,4]. Over the past two decades, multiple incursions of BTV serotypes [5] into the Mediterranean basin and the rest of Europe have dramatically changed the epidemiology of Culicoides-borne diseases in this region and increased the need for up to date taxonomic knowledge. Indeed, although Culicoides imicola Kieffer, 1913 is known as a proven BTV vector species for a very long time [6, 7], recent BTV outbreaks, especially in regions where C. imicola is absent, have highlighted that other species are able to transmit BTV. Within species present in the Mediterranean region, some other species of the subgenus Avaritia, such as Culicoides obsoletus (Meigen, 1818) and Culicoides scoticus Downes & Kettle, 1952, are considered probable BTV vectors based on their ecological habits, on virus isolation or viral genome detections from field-collected individuals and on experimental infections [8,9,10]. Moreover, several species of the subgenus Culicoides, such as Culicoides pulicaris (Linnaeus, 1758), Culicoides lupicaris Downes & Kettle, 1952 Culicoides punctatus (Meigen, 1804) or Culicoides newsteadi Austen, 1921, are considered possible BTV vectors based on their ecological habits and on virus isolation or viral genome detections from field-collected individuals [11,12,13,14]. Finally, BTV genome has been found in the abdomen and thorax of Culicoides paolae Boorman, 1996 [15], questioning its role as possible vector.
Previous investigations of the Culicoides fauna of Morocco have been prompted by major epizootics of Culicoides-borne diseases. The first intensive faunistic inventory started after the large AHSV outbreaks affecting northern Africa and Spain in 1965–66 [16]. Various surveys conducted in 1968–69 and 1970–71 throughout the country reported the existence of many Culicoides species and described two new species: Culicoides calloti Kremer, Delécolle, Bailly-Choumara & Chaker, 1979 and Culicoides landauae Kremer, Rebholtz-Hirtzel & Bailly-Choumara, 1975 [17,18,19,20,21]. Two additional species, Culicoides clastrieri Callot, Kremer & Deduit, 1962 and Culicoides duddingstoni Kettle & Lawson, 1955, were collected in Fès-Meknès Region in 1972, but the information was not published (Delécolle, pers. comm.). Taking into account the current classification of the genus Culicoides, the checklist in the 1970s consisted of at least 54 species (Table 1).
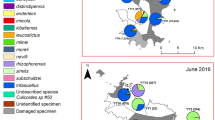
A second AHS epizootic occurred in Spain in 1987, from where it spread to Morocco in 1989 and persisted until 1991 [22]. At that time, the Afrotropical species C. imicola was the only proven AHSV vector and was considered the only species involved in AHSV transmission in Morocco [23, 24]. As a result, surveys conducted in Morocco in 1994–95 were focused on characterizing the distribution and abundance of C. imicola [23, 25]. Culicoides imicola is very widely dispersed, found at altitudes of up to 1275 meters and under climatic conditions ranging from Mediterranean to Saharan, with the highest catches in the lower areas of the northwest (between Tangier and Rabat) and in Marrakech (Fig. 1). Other species reported were Culicoides circumscriptus Kieffer, 1918, C. newsteadi, Culicoides puncticollis (Becker, 1903) and specimens close to Culicoides festivipennis Kieffer, 1914 (mentioned under its synonym name, Culicoides odibilis Austen, 1921), but also C. obsoletus and C. pulicaris. The two latter referred more likely to ‘group’ than to single species. Indeed, authors confirmed that morphologically close species might have been grouped with C. obsoletus or C. pulicaris, as identification focused mainly on C. imicola [23].

Map of the regions in the Kingdom of Morocco
The late 1990s were marked by the occurrence of massive and regular BTV outbreaks in the Mediterranean [5]. The scale of these successive epizootic diseases has led to the establishment of entomological surveillance networks dedicated to Culicoides in most Mediterranean countries. In Morocco, a cross-sectional study was therefore carried out in 2000–2003 by the veterinary services in 50 sites. This third study confirmed the wide distribution of C. imicola, which was the most frequent species in the collections [26], but did not provide additional inventory information, except for C. imicola; the samples collected by these authors were identified only at the level of species groups.
The first BTV outbreak in Morocco was reported in September 2004 [27]. Since then, BTV outbreaks have been reported every year, except for 2008, in the entire Moroccan territory, whereas the country faced EHDV outbreaks in 2006. Following snapshot entomological surveys carried out in BTV transmission sites [28], a new entomological surveillance network was set up in 2009–2010 as part of the epidemiological surveillance of animal diseases organized by the national veterinary services [29]. This fourth study resulted in the publication of an updated Culicoides checklist for Morocco, with the addition of a new species based on a single morphologically identified specimen: Culicoides helveticus Callot, Kremer & Deduit, 1962. The lack of molecular confirmation for this individual alone suggests caution in including this new species in the national checklist.
In the present study, we aimed to revise the current inventory of Culicoides species present in Morocco in light of the most up to date taxonomic and systematic knowledge. The studies in the 1970s used species names that are now considered as synonymous. Morphological identification of Culicoides is also difficult; variations in diagnostic characteristics are common and could lead to false affiliation of specimens [30, 31]. Therefore, we aimed to confirm the checklist for Morocco based on the examination of fresh material collected in the Rabat region, the Moroccan capital, by an integrative taxonomy approach using morphological identification supplemented by molecular identification to confirm species records.
Methods
Literature related to Culicoides collections in Morocco, including articles published in French, English and unpublished ‘grey’ literature (mainly reports or theses), was collated. Authors were contacted to obtain raw data on the collections and details on the level of identification (species or ‘group’).
In these previous studies, only morphological identification was used. We thus decided to collect and examine fresh Culicoides material from two sites around Rabat, a horse-riding center (33°56′N, 6°49′W) (site M1) and a goat farm (33°51′N, 6°51′W) (site M2) to make samples available for molecular identification. The objective was to confirm the record of some species for which morphological identification is difficult. Collection was made using an Onderstepoort Veterinary Institute black light suction trap operated from dusk until dawn, on two consecutive days every two weeks (from May to October 2016). When possible, collected individuals were morphologically identified to the species level using relevant morphological identification keys [32, 33].
Additionally, molecular identification was performed (i) to identify closely related species (such as C. obsoletus, C. scoticus or Culicoides montanus Shakirzjanova, 1962); (ii) to confirm difficult morphological identifications (damaged specimen or uncertain diagnostic features); and (iii) to confirm new species records. Genomic DNA was individually extracted using the NucleoSpin® Tissue DNA Kit (Macherey-Nagel, Duren, Germany) according to the manufacturer’s instructions. For C. obsoletus/C. scoticus, the allele-specific PCR from Nolan et al. [34] was used to separate the two morphologically similar species. For other species, cox1 amplification and sequencing using primers were produced following procedure described by Bakhoum et al. [35], before final species assignment was done by phylogenetic reconstructions and BLAST search of GenBank entries and an Afrotropical BOLD database (AFCUL001-18 to AFCUL1131-18).
The newly generated cox1 sequences were edited in GENEIOUS R6 (Biomatters, http://www.geneious.com/) then aligned using the MUSCLE algorithm [36]. Maximum Likelihood (ML) phylogenetic trees were computed under a substitution model estimated using the JMODELTEST tool within MEGA X [37]. The Bayesian information criterion implemented within JMODELTEST was used to determine the most suitable evolutionary model; this was the general time reversible (GTR) model with gamma distributed (+G) rate variation among sites (5 categories; parameter = 0.6015) and proportion of invariable sites (+I) of 22.82%. Missing data, including indels, were excluded from the analysis. The tree was subjected to 1000 bootstrap replications to assess topological reliability. Initial tree(s) for the heuristic search were obtained automatically by applying Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the Maximum Composite Likelihood (MCL) approach, and then selecting the topology with superior log likelihood value.
Specimens were stored in alcohol with one specimen per species slide-mounted and are available upon request to Maria Bourquia and Khalid Khallaayoune at Agronomic and Veterinary Institute Hassan II, Rabat, Morocco. Sequences were deposited in the GenBank database under the accession numbers MK732283–MK732313. Sequences from the GenBank and BOLD databases were used in the tree reconstruction. Sequences for Culicoides halophilus Kieffer, 1924 (GenBank: KX853271 and KX853272) were used although not considered as a valid species by the world catalogue of Culicoides [38].
Results
A total of 6121 Culicoides were examined, representing at least 17 species collected during a total of 12 collection sessions. Morphological identification confirmed the presence of 15 previously reported species and established two new records: C. paolae with 6 individuals and Culicoides subfagineus Delécolle & Ortega, 1998 with 3 individuals (Table 1). We dissected and mounted the wings of the different species including C. paolae and C. subfagineus (Fig. 2).

Wing patterns of Culicoides spp. collected during the present survey
Molecular identification was performed on 105 individuals. A total of 71 females morphologically identified as C. obsoletus/C. scoticus were identified with the species-specific PCR [34] allowing the identification of 59 C. obsoletus and 7 C. scoticus. Five individuals could not be identified to the species level and were kept as C. obsoletus/C. scoticus. cox1 sequences were generated from 34 individuals of which 3 individuals did not produce exploitable sequences. Alignment was 390 bp long and included 31 sequences plus 135 references sequences retrieved from GenBank, BOLD or expert molecular library.
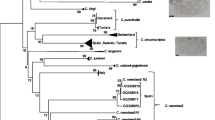
The resulting tree was used as a species affiliation tool and not to infer systematic relationships. Indeed, the node robustness was high (above 90%) only at the terminal nodes which allows affiliation at the species level. From the tree we infer the presence of Culicoides cataneii Clastrier, 1957 (2 sequences), C. circumscriptus (1 sequence), C. imicola (1 sequence), Culicoides jumineri Callot & Kremer, 1969 (5 sequences), C. montanus (1 sequence), C. obsoletus (1 sequence), C. paolae (6 sequences), Culicoides sahariensis Kieffer, 1923 (2 sequences), Culicoides kingi Austen, 1912 (3 sequences), C. subfagineus (3 sequences) and 6 sequences clustering in the same clade without voucher sequence (Fig. 3). Molecular affiliation of two sequences (MK732287 and MK732288) should be considered with caution. They clustered in proximity to C. jumineri sequences. However, these two individuals could not be considered with certainty as C. jumineri, due to the bootstrap values being too low, highlighting genetic differences between these two sequences and the C. jumineri reference sequences.

Phylogenetic tree based on cox1 polymorphism. In bold, sequences produced in this study with GenBank reference numbers in brackets. The evolutionary history was inferred using the Maximum Likelihood method and GTR+G+I model. This analysis involved 166 nucleotide sequences. All positions containing gaps and missing data were eliminated (complete deletion option); there were a total of 390 positions in the final dataset. The tree is drawn to scale, with branch lengths corresponding to the number of substitutions per site
Discussion
Species inventories allow completing and updating taxonomic and systematic revision to develop identification keys, establish national or regional species checklists and field guides, and make all the relevant information readily available through scientific publications and other dissemination tools. Regular updates of faunistic inventories of economically important biological groups are of great importance to maintain expertise and overcome taxonomic obstacles, train young scientists and ensure the stability of species community structure in an environment subject to the introduction and colonization of exotic species [39].
The present paper revised, updated and confirmed the checklist of Culicoides for Morocco using combined morphological and molecular identification and, to the authors’ knowledge, reports for the first time C. paolae and C. subfagineus in the country. The revised checklist includes 54 species of Culicoides recorded from the territory of Morocco. However, it is noted that the southernmost Saharan region of Eddakhla-Oued Eddahab has, to our knowledge, never been prospected (Fig. 1). The neighboring region of Laayoune-Sakia El Hamra has only been prospected around Laayoune in 2009–2010 [29]. The few specimens collected belonged to C. imicola, and to Newsteadi and Circumscriptus groups of species. Therefore, we have not included these two regions in Table 2. The small number of Culicoides specimens collected in Laayoune may be related to the arid climate prevailing in this region.
Our review of the literature highlighted the use of synonym names for 11 species (C. sahariensis, C. imicola, Culicoides univittatus Vimmer, 1932, C. newsteadi, Culicoides odiatus Austen, 1921, C. festivipennis, Culicoides pallidus Khalaf, 1957, Culicoides ravus de Meillon, 1936, Culicoides kibunensis Tokunaga, 1937, Culicoides sergenti Kieffer, 1921 and Culicoides truncorum Edwards, 1939). Some authors also re-evaluated their own collections. Culicoides riethi Kieffer, 1914 erroneously reported [17], was modified to C. puncticollis in 1971 [18]. Caution should therefore be exercised when using former publications. It is mandatory to refer to the world catalogue for Culicoides [38] to validate species names and update them if needed. Moreover, our literature review highlighted the lack of identification tools for Culicoides in Morocco, and more broadly in North Africa, as already published for the Western Palaearctic region [33].
The updated checklist for the country was established after a critical analysis of the literature and from our novel collections. Culicoides begueti Clastrier, 1957 is mentioned by Kremer et al. [18] from unpublished collections in the Marrakech region. As materials and references to these catches were not available, the species has been removed from the list pending further reports. The presence of individuals belonging to the ‘Kingi-schultzei group’ was reported by Bailly-Choumara & Kremer in Souss-Massa Region [17, 21]. According to our molecular and morphological analysis, this species could be C. kingi. Culicoides helveticus was collected as a unique individual by Lhor et al. [29]. This species can be confused with Culicoides stigma (Meigen, 1818) and Culicoides parroti Kieffer, 1922, which have the same wing pattern, but can be distinguished by the shape of the spermatheca [40]. In our collections, C. parroti was observed. Considering the known distribution of C. helveticus is the mountainous regions of the Palaearctic region such as in Russia [41,42,43], France [40], Sweden [44] and Ukraine [45], we considered it was appropriate to remove this species from Moroccan list of Culicoides.
Our molecular analysis revealed that the specimens morphologically identified as C. obsoletus/C. scoticus belong to C. obsoletus, C. scoticus and one specimen of C. montanus. Indeed, C. montanus is a rare species that is phylogenetically very close to C. obsoletus [46, 47] for which the taxonomic status is a little controversial [48, 49]. Recently, several authors have reported the existence of cryptic diversity within the C. obsoletus taxon [50, 51], namely C. obsoletus clade O2, C. obsoletus dark and one unnamed entity. Albeit from a limited number of individuals, we did not observe such cryptic diversity within the C. obsoletus taxon in Morocco.
A clade of 6 sequences (i.e. MK732297, MK732292, MK732298, MK732283, MK732284 and MK732286) received high bootstrap support. This clade is part of the same phylogroup as C. santonicus Callot, Kremer, Rault & Bach, 1966 which belongs to the Vexans group, grouping three species with Culicoides brunnicans Edwards, 1939 and Culicoides vexans (Staeger, 1839). We hypothesized that it is either a novel species or simply one without voucher sequences in molecular libraries. This could be investigated by increasing the number of voucher sequences to other Afrotropical or Palaearctic species. Moreover, the C. jumineri clade showed genetic variability that may indicate that cryptic species diversity is present within this clade.
To our knowledge, our study records C. paolae for the first time in Morocco, whereas it was previously known from Italy [52], Greece [53], Algeria [54] and Tunisia [55,56,57]. This species, largely distributed in the Mediterranean region, was not considered, until recently, to be associated with BTV transmission. Indeed, BTV genome was recovered from the thorax of field-collected individuals in Sardinia during BTV outbreaks [15], raising a possible role of this species in BTV transmission. Updating the distribution of this species is thus of primary interest. To our knowledge, we also describe C. subfagineus for the first time in the country. Our specimens were first morphologically identified as Culicoides grisescens Edwards, 1939 but tree reconstruction clearly assigned all three specimens to C. subfagineus. Culicoides subfagineus was recently reported for the first time in Portugal [58] and Germany [59]. Some authors confuse this species with Culicoides fagineus Edwards, 1939 and group it into C. subfagineus/C. fagineus as pairs of species [60].
Finally, Moroccan fauna is composed of species with a different geographical distribution and can be classified into three groups: (i) species with a wide distribution covering the Palaearctic region (from Scandinavia to the Mediterranean) such as C. circumscriptus, C. clastrieri, C. duddingstoni, Culicoides dzhafarovi Remm, 1967, C. fagineus, C. festivipennis, C. jumineri, C. kibunensis, Culicoides kurensis Dzhafarov, 1960, Culicoides maritimus Kieffer, 1924, C. montanus, C. newsteadi, C. obsoletus, C. odiatus, C. parroti, Culicoides pictipennis (Staeger, 1839), Culicoides picturatus Kremer & Deduit, 1961, C. pulicaris, C. punctatus, C. puncticollis, Culicoides saevus Kieffer, 1922, C. scoticus, Culicoides shaklawensis Khalaf, Ingram & Macfie, 1957, Culicoides simulator Edwards, 1939, Culicoides subfasciipennis Kieffer, 1919 and Culicoides vidourlensis Callot, Kremer, Molet & Bach, 1968; (ii) species distributed around the Mediterranean basin such as Culicoides azerbajdzhanicus Dzhafarov, 1962, C. cataneii, Culicoides derisor Callot & Kremer, 1965, Culicoides faghihi Navai, 1971, Culicoides heteroclitus Kremer & Callot, 1965, C. jumineri, Culicoides langeroni Kieffer, 1921, Culicoides longipennis Khalaf, 1957, Culicoides marcleti Callot, Kremer & Basset, 1968, C. paolae, Culicoides pseudopallidus Khalaf, 1961, C. sahariensis, C. santonicus, Culicoides sejfadinei Dzhafarov, 1958, Culicoides semimaculatus Clastrier, 1958, Culicoides sergenti Kieffer, 1921, C. subfagineus, C. univittatus; and (iii) Afrotropical species also present in the southern part of the Palaearctic zone, especially in the Mediterranean such as C. imicola, C. kingi, C. ravus and Culicoides similis Carter, 1920.
Some species have limited numbers of records in the literature making their known geographical distribution too sporadic to be assigned to the above groups: Culicoides badooshensis Khalaf, 1961 (reported in Irak, Iran, Morocco and Turkey), C. pallidus (Morocco), C. landauae (Morocco), C. calloti (Morocco) and Culicoides pumilus (Winnertz, 1852) (Ukraine, France and Morocco).
The two other countries of the Maghreb, Algeria and Tunisia, have many bio-climatic similarities such as a Mediterranean coastline in the north and Sahara desert in the south. The Culicoides fauna of these countries has been studied in two steps, as in Morocco. First, inventories were carried out in the 1950s and the 1980s in Algeria [61, 62], leading to the record of 31 species, and in the 1980s in Tunisia [63], leading to the record of 22 species. Then, inventories were completed during studies following the BTV emergence in the 2000s in the Maghreb and in the whole Mediterranean region. Today, the number of species recorded in Morocco (54 species) was slightly higher than in Algeria with 47 species [54] and in Tunisia with 35 species [55, 57, 64, 65]. The same increasing species richness gradient from Tunisia to Morocco is observed for mosquitoes [66], and may be related to the greater diversity of climates and landscapes found in Morocco, including the influence of the ocean climate prevailing on the northwest coast and not existing in the two neighboring countries.
Conclusions
The Culicoides fauna of Morocco includes 54 species, at least 8 of which are recognized vectors or potential vectors of Culicoides-borne viruses of economic importance in animal production. Among these, C. imicola is a proven BTV and AHSV vector [6, 7, 67] and was considered for a long time the only important vector. Most epidemiological studies have therefore focused on this species [23]. In this study, we confirmed by molecular assay the presence of both C. obsoletus and C. scoticus in Morocco, which are considered as probable BTV vectors in the western Palaearctic region [68, 69]. Moreover, we reported the presence of C. kingi, a potential EHDV vector [70, 71] and of C. paolae, which, together with C. newsteadi, C. pulicaris and C. punctatus [14, 15, 72], is regarded as a potential BTV vector. Establishing a comprehensive checklist of Culicoides for Morocco is a prerequisite for developing a barcode library and atlas of diagnostic characters, but also to plan further ecological studies, including large scale Culicoides collections focusing species of veterinary interest, to be able to carry out risk mapping for related Culicoides-borne diseases.
Availability of data and materials
Specimens are stored in ethanol with one specimen per species slide-mounted and are available upon request to Maria Bourquia and Khalid Khallaayoune at Agronomic and Veterinary Institute Hassan II, Rabat, Morocco.
Abbreviations
- AHSV:
-
African horse sickness virus
- BTV:
-
bluetongue virus
- EHDV:
-
epizootic hemorrhagic disease virus
References
Du Toit RM. The transmission of bluetongue and horse sickness by Culicoides. Onderstepoort J Vet. 1944;19:7–16.
Mellor PS, Boorman J, Baylis M. Culicoides biting midges. Their role as arbovirus vectors. Annu Rev Entomol. 2000;45:307–40.
Coetzer JAW, Guthrie AJ. African horse sickness. In: Coetzer JAW, Tustin RC, editors. Infectious diseases of livestock. 2nd ed. Cape Town: Oxford University Press; 2004. p. 1231–46.
Balenghien T, Pagès N, Goffredo M, Carpenter S, Augot D, Jacquier E, et al. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev Vet Med. 2014;116:360–9.
Wilson AJ, Mellor PS. Bluetongue in Europe: past, present and future. Philos Trans R Soc Lond B Biol Sci. 2009;364:2669–81.
Venter GJ, Graham SD, Hamblin C. African horse sickness epidemiology: vector competence of South African Culicoides species for virus serotypes 3, 5 and 8. Med Vet Entomol. 2000;14:245–50.
Mellor PS, Hamblin C. African horse sickness. Vet Res. 2004;35:445–66.
Mellor PS, Pitzolis G. Observations on breeding sites and light-trap collections of Culicoides during an outbreak of bluetongue in Cyprus. Bull Ent Res. 1979;69:229–34.
Savini G, Goffredo M, Monaco F, Di Gennaro A, Cafiero MA, Baldi L, et al. Bluetongue virus isolations from midges belonging to the Obsoletus complex (Culicoides, Diptera: Ceratopogonidae) in Italy. Vet Rec. 2005;157:133–9.
Carpenter S, McArthur C, Selby R, Ward R, Nolan DV, Luntz AJM, et al. Experimental infection studies of UK Culicoides species midges with bluetongue virus serotypes 8 and 9. Vet Rec. 2008;163:589–92.
Caracappa S, Torina A, Guercio A, Vitale F, Calabrò A, Purpari G, et al. Identification of a novel bluetongue virus vector species of Culicoides in Sicily. Vet Rec. 2003;153:71–4.
De Liberato C, Scavia G, Lorenzetti R, Scaramozzino P, Amaddeo D, Cardeti G, et al. Identification of Culicoides obsoletus (Diptera: Ceratopogonidae) as a vector of bluetongue virus in central Italy. Vet Rec. 2005;156:301–4.
Romon P, Higuera M, Delécolle JC, Baldet T, Aduriz G, Goldarazena A. Phenology and attraction of potential Culicoides vectors of bluetongue virus in Basque Country (northern Spain). Vet Parasitol. 2012;186:415–24.
Goffredo M, Catalani M, Federici V, Portanti O, Marini V, Mancini G, et al. Vector species of Culicoides midges implicated in the 2012–2014 bluetongue epidemics in Italy. Vet Ital. 2015;51:131–8.
Foxi C, Meloni G, Puggioni G, Manunta D, Rocchigiani A, Vento L, et al. Bluetongue virus detection in new Culicoides species in Sardinia, Italy. Vet Rec. 2019;184:621.
Sellers RF, Pedgley DE, Tucker MR. Possible spread of African horse sickness on the wind. J Hyg (Lond). 1977;79:279–98.
Bailly-Choumara H, Kremer M. Deuxième contribution à l’étude des Culicoides du Maroc (Diptera: Ceratopogonidae). Entomol Méd Parasitol. 1970;4:383–91.
Kremer M, Hommel M, Bailly-Choumara H. Troisième contribution à l’étude faunistique des Culicoides du Maroc. Ann Parasitol Hum Comp. 1971;46:661–70.
Kremer M, Rebholtz-Hirtzel C, Bailly-Choumara H. Quatrième contribution à l’étude faunistique des Culicoides (Diptera, Ceratopogonidae) du Maroc: description de C. landauae n. sp. (Diptera, Ceratopogonidae), redescription de C faghihi Navai et d’une forme de C. subfascipennis Kieffer. Entomol Méd Parasitol. 1975;4:205–14.
Kremer M, Delécolle JC, Bailly-Choumara H, Chaker E. Cinquième contribution à l’étude faunistique des Culicoides (Diptera, Ceratopogonidae). Entomo Méd Parasitol. 1979;3:195–9.
Chaker E, Bailly-Choumara H, Kremer M. Sixième contribution à l’étude faunistique des Culicoides du Maroc (Diptera, Cefratopogonidae). Bull Inst Sci Rabat. 1979;4:81–6.
Mellor PS, Boned J, Hamblin C, Graham S. Isolations of African horse sickness virus from vector insects made during the 1988 epizootic in Spain. Epidemiol Infect. 1990;105:447–54.
Baylis M, Bouayoune H, Touti J, El Hasnaoui H. Use of climatic data and satellite imagery to model the abundance of Culicoides imicola, the vector of African horse sickness virus in Morocco. Med Vet Entomol. 1997;12:255–66.
Rawlings P, Pro MJ, Pena I, Ortega MD, Capela R. Spatial and seasonal distribution of Culicoides imicola in Iberia in relation to the transmission of African horse sickness virus. Med Vet Entomol. 1997;11:49–57.
Bouayoune H, Touti J, El Hasnaoui H, Baylis M, Mellor PS. The Culicoides vectors of African horse sickness virus in Morocco: distribution and epidemiological implications. Arch Virol. 1998;14:113–25.
Fassi-Fihri O, Lhor Y. Fièvre catarrhale ovine (FCO). Historique et situation épidémiologique au Maroc. FAO report. Rome: FAO; 2008. p. 24.
World Organization for Animal Health. Bluetongue in Morocco. OIE disease information, Animal Health Data. 2004. https://web.oie.int/hs2/zi_pays.asp?c_pays=117&annee=2004. Accessed 2 Sep 2019.
Cêtre-sossah C, Baldet T. Mission d’expertise entomologique et virologique, fièvre catarrhale ovine - Maroc. Rapport de mission à l’IAV Hassan II. Montpellier: Cirad; 2004. p. 12.
Lhor Y, Khayli M, Bouslikhane M, El Harrak M, Fassi-Fihri O. Spatial and seasonal distribution of Culicoides species in Morocco in relation to the transmission of bluetongue viruses. Br J Virol. 2015;2:88–95.
Meiswinkel R, Gomulski LM, Delécolle JC, Goffredo M, Gasperi G. The taxonomy of Culicoides vector complexes-unfinished business. Vet Ital. 2004;40:151–9.
Harrup LE, Bellis GA, Balenghien T, Garros C. Culicoides Latreille (Diptera: Ceratopogonidae) taxonomy: current challenges and future directions. Infect Genet Evol. 2015;30:249–66.
Delécolle JC. Nouvelle contribution à lʼétude systématique et iconographique des espèces du genre Culicoides, (Diptéra: Cératopogonidae) du Nord-Est de la France. PhD thesis. Université Louis Pasteur de Strasbourg I, France; 1985.
Mathieu B, Cêtre-Sossah C, Garros C, Chavernac D, Balenghien T, Carpenter S, et al. Development and validation of IIKC: an interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasit Vectors. 2012;5:137.
Nolan DV, Carpenter S, Barber J, Mellor PS, Dallas JF, Mordue Luntz AJ, Piertney SB. Rapid diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet Microbiol. 2007;124:82–94.
Bakhoum MT, Labuschagne K, Huber K, Fall M, Mathieu B, Venter G, et al. Phylogenetic relationships and molecular delimitation of Culicoides Latreille (Diptera: Ceratopogonidae) species in the Afrotropical region: interest for the subgenus Avaritia. Syst Entomol. 2018;43:355–71.
Edgar RC. Muscle: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7.
Darriba D, Taboada GL, Doallo R, Posada D. jModelTest2: More models, new heuristics and parallel computing. Nature Med. 2012;9:772.
Borkent A. The subgeneric classification of species of Culicoides 2016 - thoughts and a warning. 2016. http://wwx.inhs.illinois.edu/files/5014/6532/8290/CulicoidesSubgenera.pdf. Accessed 2 Sep 2019.
Silveira LF, De Mello Beisiegel B, Curcio FF, Valdujo PH, Dixo M, Verdade VK, et al. What use do fauna inventories serve? Estudos Avançados. 2010;24:173–207.
Kremer M. Contribution à l’étude du genre Culicoides Latreille particulièrement en France. In: Encyclopédie entomologique, série A. Paris: Editions Paul Lechevalier; 1965.
Mezenev NP. Blood-sucking biting midges (Ceratopogonidae) in north central Siberia. Parazitologiya. 1990;24:28–36.
Aibulatov SV. Bloodsucking dipterans (Diptera: Ceratopogonidae, Culicidae, Simuliidae, Tabanidae) of the Kurgala Peninsula, Leningrad Province. Entomol Rev. 2009;89:645–58.
Sprygin AV, Fiodorova OA, Babin YY, Elatkin NP, Mathieu B, England ME, et al. Culicoides biting midges (Diptera, Ceratopogonidae) in various climatic zones of Russia and adjacent lands. J Vector Ecol. 2014;39:306–15.
Nielsen SA, Nielsen B, Chirico J. Monitoring of biting midges (Diptera: Ceratopogonidae: Culicoides Latreille) on farms in Sweden during the emergence of the 2008 epidemic of bluetongue. Parasitol Res. 2010;6:1197–203.
Filatov S, Szadziewski R. Annotated checklist and distribution of Culicoides biting midges of Ukraine (Diptera: Ceratopogonidae). J Nat Hist. 2017;51:487–511.
Gomulski LM, Meiswinkel R, Delécolle JC, Goffredo M, Gasperi G. Phylogenetic relationships of the subgenus Avaritia Fox, 1955 including Culicoides obsoletus (Diptera, Ceratopogonidae) in Italy based on internal transcribed spacer 2 ribosomal DNA sequences. Med Vet Entomol. 2006;20:229–38.
Mathieu B, Delécolle JC, Garros C, Balenghien T, Setier-Rioc ML, Candolfia E, et al. Simultaneous quantification of the relative abundance of species complex members: application to Culicoides obsoletus and Culicoides scoticus (Diptera: Ceratopogonidae), potential vectors of bluetongue virus. Vet Parasitol. 2011;182:297–306.
Kiehl E, Walldorf V, Klimpel S, Al-Quraishy S, Mehlhorn H. The European vectors of bluetongue virus: are there species complexes, single species or races in Culicoides obsoletus and C pulicaris detectable by sequencing ITS-1, ITS-2 and 18S RDNA? Parasitol Res. 2009;105:331–6.
Garros C, Mathieu B, Balenghien T, Cêtre-Sossah C, Delécolle JC, et al. Comments on Kiehl et al. (2009) “The European vectors of Bluetongue virus: are there species complexes, single species or races in Culicoides obsoletus and C. pulicaris detectable by sequencing ITS-1, ITS-2 and 18S-rDNA? Parasitol Res. 2010;107:731–4.
Wenk CE, Kaufmann C, Schaffner F, Mathis A. Molecular characterization of Swiss Ceratopogonidae (Diptera) and evaluation of real-time PCR assays for the identification of Culicoides biting midges. Vet Parasitol. 2012;184:258–66.
Meiswinkel R, De Bree F, Bossers-De Vries R, Elbers AR. An unrecognized species of the Culicoides obsoletus complex feeding on livestock in the Netherlands. Vet Parasitol. 2015;207:324–8.
Boorman J, Mellor PS, Scaramozzino P. A new species of Culicoides (Diptera, Ceratopogonidae) from southern Italy. Parassitologia. 1996;38:501–3.
Patakakis MJ, Papazahariadou M, Wilson A, Mellor PS, Frydas S, Papadopoulos O. Distribution of Culicoides in Greece. J Vector Ecol. 2009;34:243–51.
Djerbal M, Delécolle JC. Surveillance entomologique de la fièvre catarrhale ovine en Algérie. Rev Elev Méd Vét Pays Trop. 2009;62:81–180.
Chaker E, Sfari M, Rouis M, Babba H, Azaiez R. Faunistic note of Culicoides (Diptera, Ceratopogonidae) from Monastir (Tunisia). Parasite. 2005;12:359–61.
Augot D, Mathieu B, Hadj Henni L, Barriel V, Mena SZ, Smolis S, et al. Molecular phylogeny of 42 species of Culicoides (Diptera, Ceratopogonidae) from three continents. Parasite. 2017;24:23.
Sghaier S, Hammami S, Goffredo M, Hammami M, Portanti O, Lorusso A, et al. New species of the genus Culicoides (Diptera Ceratopogonidae) for Tunisia, with detection of bluetongue viruses in vectors. Vet Ital. 2017;53:357–66.
Ramilo DW, Diaz S, Fonseca I, Delécolle JC, Wilson A, Meireles J, et al. First report of 13 species of Culicoides (Diptera: Ceratopogonidae) in mainland Portugal and Azores by morphological and molecular characterization. PLoS One. 2012;7:e34896.
Ayllón T, Nijhof AM, Weiher W, Bauer B, Allène X, Clausen PH. Feeding behaviour of Culicoides spp. (Diptera: Ceratopogonidae) on cattle and sheep in northeast Germany. Parasit Vectors. 2014;7:34.
Pudar D, Petrić D, Allène X, Alten B, Ayhan N, Cvetkovikj A, et al. An update of the Culicoides (Diptera: Ceratopogonidae) checklist for the Balkans. Parasit Vectors. 2018;11:462.
Clastrier J. Notes sur les Cératopogonidés. II. Quelques Culicoides d’Algérie à ailes tachetées. Arch Inst Pasteur Algérie. 1957;35:404–44.
Szadziewski R. Ceratopogonidae (Diptera) from Algeria. VI. Culicoides Latr. Bull Entomo Pol. 1984;54:163–82.
Chaker E, Kremer M. Culicoides of Tunisia: morphological characteristics Chorology and ecology of species found. Arch Inst Pasteur Tunis. 1982;59:511–40.
Hammami S, Bouzid M, Hammou F, Fakhfakh E, Delécolle JC. Occurrence of Culicoides spp. (Diptera: Ceratopogonidae) in Tunisia, with emphasis on the Bluetongue vector C. imicola. Parasite. 2008;15:179–81.
Slama D, Babba H, Chaker E. Culicoides spp. (Diptera: Ceratopogonidae) in Tunisia. In: Shields VDC, editor. Biological control of pest and vector insects. London: InTechOpen; 2016. p. 213–29.
Gunay F, Picard M, Robert V. MosKeyTool, an interactive identification key for mosquitoes of Euro-Mediterranean. Version 2.1. http://www.medilabsecure.com/moskeytool.
Paweska JT, Venter GJ, Hamblin C. A comparison of the susceptibility of Culicoides imicola and C. bolitinos to oral infection with eight serotypes of epizootic haemorrhagic disease virus. Med Vet Entomol. 2005;19:200–7.
Goffredo M, Meiswinkel R, Federici V, Di Nicola F, Mancini G, Ippoliti C, et al. The ‘Culicoides obsoletus group’ in Italy: relative abundance, geographic range, and role as vector for Bluetongue virus. Vet Ital. 2016;52:235–41.
Paslaru AL, Mathis A, Torgerson P, Veronesi E. Vector competence of pre-alpine Culicoides (Diptera: Ceratopogonidae) for bluetongue virus serotypes 1, 4 and 8. Parasit Vectors. 2018;11:466.
Mellor PS, Osborne R, Jennings DM. Isolation of bluetongue and related viruses from Culicoides spp. in the Sudan. J Hyg (Lond). 1984;93:621–8.
Lee VH. Isolation of viruses from field populations of Culicoides (Diptera : Ceratopogonidae) in Nigeria. J Med Entomol. 1979;16:76–9.
Foxi C, Delrio G, Falchi G, Marche MG, Satta G, Ruiu L. Role of different Culicoides vectors (Diptera: Ceratopogonidae) in Bluetongue virus transmission and overwintering in Sardinia (Italy). Parasit Vectors. 2016;9:440.
Acknowledgments
The authors thank Matthew Baylis (University of Liverpool, UK) for sharing the raw data from 1994–1995 collections, François Thiaucourt for the help with the illustrations and finally the owner of the farms where 2016 collections were carried out.
Funding
This study was supported by Vectornet, funded by European Foof Safety Agency and European Center for Disease Control. MB was supported by a Grant from PHC Toubkal (Toubkal/17/38 campus France 36867VH) and Centre National de Recherche Scientifique et Technique (CNRST).
Author information
Authors and Affiliations
Contributions
MB, TB, CG and KK designed the study. MB and IB contributed to the collection of Culicoides. MB, IR, LG, KH, JCD and AM identified all Culicoides specimens. MB analysed the data. IR, LG, KH, IB, JCD, TB, AM and YL contributed to the manuscript after its first draft by MB, TB, CG and KK. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bourquia, M., Garros, C., Rakotoarivony, I. et al. Update of the species checklist of Culicoides Latreille, 1809 biting midges (Diptera: Ceratopogonidae) of Morocco. Parasites Vectors 12, 459 (2019). https://doi.org/10.1186/s13071-019-3720-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-019-3720-4