
Abstract
Schistosomiasis, caused mainly by S. mansoni, S. haematobium and S. japonicum, continues to be a serious tropical disease and public health problem resulting in an unacceptably high level of morbidity in countries where it is endemic. Praziquantel, the only drug currently available for treatment, is unable to kill developing schistosomes, it does not prevent re-infection and its continued extensive use may result in the future emergence of drug-resistant parasites. This scenario provides impetus for the development and deployment of anti-schistosome vaccines to be used as part of an integrated approach for the prevention, control and eventual elimination of schistosomiasis. This review considers the present status of candidate vaccines for schistosomiasis, and provides some insight on future vaccine discovery and design.
Similar content being viewed by others


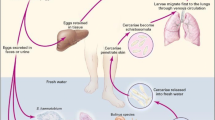
Background
The World Health Organization (WHO) considers schistosomiasis to be second only to malaria as the most devastating parasitic disease in terms of socioeconomic importance and public health impact [1]. Human infection is due to three main species, namely Schistosoma mansoni and S. japonicum which cause intestinal/hepatic schistosomiasis [2, 3] and S. haematobium, which results in urinogenital disease [4].
Human treatment with praziquantel (PZQ) is playing a central role in the control and prevention of schistosomiasis, being the only effective drug currently available [5]. However, the drug does not prevent re-infection and its exclusive use for the prevention and control of schistosomiasis is problematic; having been used for more than three decades, the emergence of PZQ-resistant schistosomes is a constant threat. Other drawbacks with PZQ are its poor activity against immature schistosomes, resulting in sub-optimal outcomes during mass drug administration campaigns and, as its mechanism of action remains unclear, the design of alternative drug formulations has proven difficult [5]. Furthermore, a substantial infrastructure is required to ensure the drug is supplied regularly in timely fashion to all parts of an endemic area.
Originally used as a major preventative measure [6], snail control, through the use of molluscicides (e.g. niclosamide), is now not the recommended method in isolation for the prevention of schistosomiasis [5]. In order to control and finally eliminate schistosomiasis, a vaccine will likely be a key component of an integrated approach (i.e. involving mass chemotherapy, targeted mollusciciding, environmental modification, health education, improved sanitation and vaccination). A transmission blocking vaccine for use in bovines could serve as a vital component in the control of S. japonicum [7], whereas clinical vaccines against S. mansoni and S. haematobium need to be developed. However, it is a sobering thought that no commercial vaccine is available currently against any of the human schistosomes, thereby emphasising the need for continued efforts towards achieving this goal. This review evaluates the current status of schistosome vaccine development.
Strategies for vaccine development
Although complex, the schistosome life-cycle, with its various stages each expressing distinct antigens, provides a vehicle for identifying many alternative molecules for vaccine development. The fact that the different stages reside in different host niches (larvae in the skin and lungs, adults in the liver and intestine or bladder capillaries) can help in the design of possible vaccines to prevent the migration of schistosome parasites and their maturation to adult worms. Importantly, the fact that schistosomes do not replicate in the definitive host makes partial reduction of the parasite burden sufficient to control schistosomiasis, strengthening the argument for developing an effective vaccine as a control intervention [8]. When identifying a suitable vaccine candidate, it is prudent to select key schistosome molecules in the live parasite that are (a) exposed to the host immune system; and (b) are essential for parasite survival. Such components may, for example, function in migration, immune evasion, nutrient uptake or attachment.
Adjuvant selection and mode of vaccine formulation and delivery are other important considerations in vaccine design and deployment as they can have a considerable impact on the protective effectiveness of the vaccine. It is well known that, in contrast to attenuated cercarial vaccines, other types, such as subunit vaccines, require an appropriate adjuvant to help stimulate the immune system. A number of different adjuvants are available such as gels, emulsions, particulates, cytokines, microbial products (e.g. CpG, cholera toxin) and proteases. Adjuvants can overcome immune senescence in older individuals, prolong the immunological memory of a vaccine, broaden the antibody repertoire and direct the immune system to a Th1 biased, Th2 biased or mixed Th1/Th2 biased response [9]. For example, a Th1 driving adjuvant such as IL-12, administered with irradiated cercariae, provided up to 90 % protection in murine schistosomiasis [10, 11]. Currently, in order to increase the protective efficacy of schistosome vaccines, a strategy of combining existing adjuvants with novel ones, developed based on emerging immunological targets, has been muted [11].
Some of the other challenges in schistosomiasis vaccine development are the risk of an atopic IgE response to a candidate vaccine [12], a lack in understanding of the nature of the immune response and the correlates of protective immunity in humans and other mammalian hosts, the transmission of other pathogens in schistosomiasis endemic areas resulting in co-infected individuals which can impact on vaccine efficacy, and antigenic polymorphism [13].
Schistosoma mansoni and S. haematobium vaccine candidates
Over 100 schistosome vaccine antigens have been identified, of which about a quarter have shown some level of protection in the murine model of schistosomiasis [14]. Disappointingly, however, only three molecules, S. mansoni fatty acid binding protein (Sm14), S. mansoni tetraspanin (Sm-TSP-2) and S. haematobium glutathione S-transferase (Sh28GST), have entered human clinical trials with Smp80 (calpain) undergoing testing in non-human primates [15].
A recent report has suggested that the murine model of schistosomiasis may be intrinsically flawed for pre-clinical testing of vaccine candidates as a result of the fragility of the pulmonary capillaries in mice which can prevent maturation of a large proportion of schistosome cercariae upon challenge; this may lead to the incorrect assumption that vaccine antigen-induced acquired protective immunity has been generated [16]. This article has stimulated vigorous debate and its conclusions require rigorous testing but, with this caveat, some of the S. mansoni and S. haematobium vaccine candidates that have been identified are now highlighted below and in Table 1.
Sm14
Schistosomes lack an oxygen-dependent pathway for the synthesis of sterols and fatty acids. Therefore, they are entirely dependent on the mammalian host to provide these essential lipids. Schistosomes use fatty acid binding proteins (FABPs) to absorb, transport and compartmentalize fatty acids from the host and, because of this critical biological function, Sm14 has long been considered a potential vaccine candidate [17]. Recombinant Sm14 (rSm14) provided up to 67 % protection in terms of reduced S. mansoni worm burden in outbred Swiss mice without the use of an adjuvant, and encouragingly, no autoimmune response was observed even though its structure is identical in basic form with mammalian host homologues [18]. It has been shown to be cross-species protective against both S. mansoni and Fasciola hepatica infection. Development of a dual vaccine effective against both fluke infections has great appeal in terms of human and animal health. Recombinant Sm14 with glucopyranosyl lipid adjuvant stable emulsion (GLA-SE) adjuvant entered and successfully completed a phase 1 clinical trial in healthy adult volunteers in Brazil, confirming its status as safe and immunogenic [19]. Further immunogenicity and safety phase 2 trials of rSm14 (adjuvanted with GLA-SE) are planned for schistosomiasis-endemic areas in Brazil and Africa [20].
Sh28GST
Schistosome 28 kDa glutathione S-transferase enzymes play a role in fatty acid metabolism and prostaglandin D2 synthesis and may help the parasite evade the host immune system. The enzyme present in S. mansoni (Sm28GST) has been tested extensively as a recombinant protein vaccine in various experimental models and has shown partial protection in terms of reduced worm burdens, inhibition of female worm fecundity and a reduction in egg viability [21]. Along with Sm14, Sm28GST was one of the six S. mansoni antigens originally independently tested under the auspices of TDR/WHO [22]. The S. haematobium homologue, Sh28GST (Bilhvax), formulated with alum adjuvant, has undergone human clinical trials [19, 23]. Cytokine production triggered by this vaccine candidate was shown to be influenced by factors such as host age, schistosome infection status and PZQ treatment history and this has provided an indication as to the features which should be considered for determining the efficacy of the GST-based vaccine during its testing in targeted endemic communities [23]. The Phase 1 and 2 clinical testing showed Bilhvax was safe for healthy and infected adults and children [13]. It was scheduled to complete a phase 3 self-contained, randomized, double blind clinical trial in 2012 (https://clinicaltrials.gov/ct2/show/NCT00870649) evaluating whether co-administration of the vaccine with PZQ could delay pathology relapse due to S. haematobium infection in children. The trial results, however, have yet to be released which has raised doubts about the effectiveness of the vaccine.
Sm-p80
Calpain is a calcium activated neutral cysteine protease [24]. Prime-boost vaccination (priming with DNA and boosting with recombinant protein) with Sm-p80, the large subunit of S. mansoni calpain, in combination with resiquimod adjuvant, resulted in 49 % worm burden reduction, while 50 % protection was achieved using the recombinant protein as primary and boost vaccine (immunized and then boosted by recombinant protein) in mice [25]. With the same approach, but using a different adjuvant, oligodeoxynucleotide (ODN) 10104, 70 % worm burden reduction and 75 % egg reduction was achieved with Sm-p80 [26]. Moreover, a 58 % worm burden reduction in baboons (Papio anubis) was reported recently with the Sm-p80-based vaccine adjuvanted with resiquimod and CpG ODN [27].
Using a different approach, a Sm-p80 DNA vaccine conferred 59 % worm burden reduction and 84 % decrease in egg production in mice [28]. In baboons the vaccine provided levels of protection against S. mansoni infection comparable to those achieved by the irradiated cercarial vaccine; moreover, antibodies and IFN-γ were shown to play an important role in the protective immunity generated in this non-human primate model [29, 30].
Importantly, recombinant Sm-p80 has also been shown to exhibit cross-species protection against S. haematobium in hamsters and baboons [31]. Promisingly, enduring antibody titers were detected in mice at sixty weeks post-vaccination with recombinant Sm-p80, and IgG specific for Sm-p80 was detected in baboons 5–8 years after initial vaccination with the Sm-80 DNA vaccine [32]. It is anticipated that the recombinant Sm-p80/GLA-SE vaccine, “SchistoShield”, will move forward to phase 1 and 2 human clinical trials in 2017 [31, 32]. Furthermore, it has also been shown that Sm-p80 has a therapeutic effect in vaccinated baboons through decreasing the numbers of established worms, reducing the retention of eggs in tissues, and decreasing the number of eggs excreted in faeces [33].
Sm-TSP-2
The tetraspanins are a group of proteins that are highly abundant in the schistosome tegument where they are found at the outer-most membrane of the intra-mammalian stage of the parasite, and hence are highly exposed to the host immune system [34]. The major tetraspanin proteins in S. mansoni are Sm-TSP-1 and Sm-TSP-2 with the latter conferring protection in S. mansoni challenge animal models and also correlating with protective immunity in naturally resistant people [35]. Sm-TSP-2 is currently being developed by the Sabin Vaccine Institute Product Development Partnership as a 9 kDa recombinant Sm-TSP-2/Alhydrogel® vaccine in combination with the GLA-AF adjuvant; it has undergone toxicology studies [36], has shown good preclinical results, expression of the antigen can be readily scaled up [37] (http://www.sabin.org/updates/pressreleases/phase-1-clinical-trial-sm-tsp-2-schistosomiasis-vaccine), and the vaccine has completed a phase 1 clinical trial although the outcome is not yet known. Disappointedly, whereas high levels of IgG1 and IgG2 antibodies were generated in mice against the recombinant TSP-2 protein homologue from S. japonicum, no consistent protective efficacy was achieved [38].
Sm29
Sm29 is present in the tegument of adult worms and schistosomula and in its recombinant form it induces high level production of IgG1 and IgG3 isotypes among individuals resistant to infection and re-infection [39]. There are reports that recombinant Sm29 can prevent infection in animals previously exposed to S. mansoni. For example, 26–48 % protection was observed in BALB/c mice that were previously infected with a Brazilian strain of S. mansoni and treated with PZQ [40]. Recently an increased level of protection was obtained through combining Sm29 with Sm14 in the presence of poly I:C adjuvant; 40.3, 68.2 and 57.9 % reductions in adult worm burden, liver egg burden and intestinal eggs, respectively, was achieved, along with a reduction in granuloma size and number in the livers of immunized mice [41]. Another report showed that by fusing Sm29 and Sm14, a 48.4 % reduction in adult worm burden in mice was achieved [42]. Similarly, fusing Sm29 with Sm-TSP-2 resulted in an increased reduction (from 22 to 35 %) in the number of worms, a higher titre of IgG1 and IgG2 antibodies and increased levels of IFNγ and TNFα than Sm29 alone in challenged mice [43, 44]. In a further advance, recombinant Sm29 was subjected to high hydrostatic pressure which dissociated the aggregated protein resulting in a successfully folded, soluble, stable and structured molecule produced in high yield, which was protective against S. mansoni, thereby paving the way for its industrial production down track [45].
Antioxidants
The antioxidants Cu–Zn superoxide dismutase (SOD) and glutathione S peroxidase (GPX) induced greater than 40 % reduction in worm burdens when administered as DNA-based vaccines in S. mansoni-challenged mice [46]. Similarly, a recent study showed a protective effect against schistosomiasis in baboons [47]. These non-human primates, vaccinated with two different formulations of SOD (SmCT-SOD and SmSP-SOD) and one of GPX, with a protocol of priming with naked DNA and boosting with the respective recombinant antioxidant proteins encapsulated in polylactic acid (PLA) microspheres, exhibited a robust immune response, which resulted in a reduction in worm numbers, and a pronounced anti-pathology effect compared with control animals [47].
Digestive tract proteins
Schistosoma mansoni worms ingest host blood which passes through the oesophagus before arriving at an area in the gut where many peptidases catalyse its proteolysis. The processing of the blood and the resulting uptake of nutrients are functions essential for the survival of the parasite. Blocking these critical processes represents an important strategy for vaccine development and a number of digestive tract proteins, that are not recognized by host immune responses during normal infection, but are essential for parasite survival, have been tested as cryptic vaccine candidates [48]. One example was the trialling of a soluble form of schistosome lysosome-associated membrane glycoprotein (Sm-LAMP), located in the gastrodermis, which resulted in a reduction in worm burden (16–25 %) and in faecal eggs (52–60 %); moreover, its insoluble form produced up to 38 % reduction in liver egg burden [49]. Another recent study identified a number of esophageal secreted proteins, encoded by microexon genes (MEGs), that are involved in the initial processing of ingested blood and these, along with lysosomal hydrolase, also localised to the oesophagus, may prove to be novel immune targets [50]. Other examples of digestive proteins as potential vaccine targets are described in Table 1 and elsewhere [51].
DLC/LC8
Proteins with the dynein light chain family (DLC/LC8) domain, which is evolutionarily conserved in different organisms, have been considered recently as vaccine targets against schistosomiasis as some homologues, such as the TAL family (Tegumental Allergen Like), are present in the distal cytoplasm of cells, in the tegument and on the membranous surface of schistosomes [52]. Recombinant S. mansoni DLC 12 and DLC 13, adjuvanted with alhydrogel, demonstrated a 43 and 51 % reduction in worm burdens respectively, in vaccinated and challenged mice [52]. Furthermore, both DLCs reduced the size of granulomas in hepatic tissues by up to 70 %. Their encouraging immunogenicity and protective efficacy, coupled with the absence of any allergic reactivity, warrants their further study as individual vaccine antigens or as part of a multi-component vaccine [52]. As DLC proteins are associated with motor myofibrillar proteins with intracellular function, the mechanism by which DLC immunization conferred protection remains to be determined.
Attenuated S. mansoni vaccines
Schistosoma mansoni cercariae, attenuated by heat, chemical or ultraviolet treatment, or by ionizing radiation (gamma or X-ray), have been shown to provide protection against S. mansoni challenge in several mammalian species [24, 53]. Recent studies undertaken with UV-radiation-attenuated cercariae given once or, more effectively, multiple times to C57BL/6 mice have confirmed earlier findings with significant reductions in worm and hepatic and intestinal egg numbers in vaccinated animals [54, 55]. Moreover, tegumental changes in the adult worms (swelling, fusion of tegumental folds, vesicle formation and loss or shortening of spines on the tubercles) were evident [54, 55].
A recent systematic meta-analysis and review of the available publications in the area on mice indicated the irradiated cercarial vaccine has the potential to achieve protection as high as 78 % with one vaccination dose [53]. Although the models showed the generated protection waned, it remained elevated for at least eight months after vaccination, reinforcing the view that the level of protective immunity obtained, although partial, would reduce both schistosome transmission and parasite-associated morbidity [53]. This re-emphasises the potential of developing an attenuated cercarial vaccine, a concept proposed some years back that might well be worth revisiting [56], with the caveat that it would in all likelihood be problematical for human use (perhaps less so for application in reservoir hosts of S. japonicum), since such an attenuated vaccine would, for example, likely carry too high a risk of side effects or of partially or unattenuated parasites reaching the mesenteric veins and becoming patent. Long-term storage of such a vaccine before deployment is another challenge that would need to be overcome.
Vaccine candidates for S. japonicum
Whereas clinical vaccines will need to be developed against S. haematobium and S. mansoni, the zoonotic nature of schistosomiasis japonica allows for a complementary approach for S. japonicum involving a transmission blocking vaccine for livestock animals, particularly bovines [24]. The vaccine would be used in reservoir hosts of S. japonicum to reduce transmission to humans [24].
Attenuated S. japonicum vaccines
Vaccination of bovines with gamma irradiated schistosomula resulted in a significant reduction in adult worms and liver eggs compared with control animals [57]. Moreover, vaccination of pigs using UV-attenuated cercariae, produced variable, although highly promising, levels of protection in terms of reduced adult worms and hepatic egg burden in animals receiving a single immunization or those vaccinated three times [58]. The protective response was shown to be associated with IFNγ and IgG2 antibody production [59]. A pilot study in miniature pigs, using similar methodology (but incorporating a lower number of challenging cercariae), resulted in more than 80 % worm reduction [60]. However, it has been reported that unstable and relatively low protection is induced in C57BL/6 mice by attenuated S. japonicum cercariae. This is likely due to the poor Th1 response generated and the less robust antibody response produced in comparison with BALB/c mice, and suggests this mouse strain might be a sub-optimal model for studying the mechanisms of immune protection against this schistosome species [61].
Sj97
Paramyosin is a myofibrillar 97 kDa protein present in the muscle layers and the tegument of schistosomes that has long been regarded as a vaccine candidate against S. japonicum and S. mansoni infection [24]. An early study showed that mice vaccinated intraperitoneally with purified paramyosin (without the use of an adjuvant) stimulated 62–86 % resistance against S. japonicum cercarial challenge [62]. In addition to preventing infection, a longitudinal treatment-re-infection design study in Leyte, the Philippines showed that a Th2 bias in response to Sj97 predicted a longer time to human re-infection and a lower re-infection intensity after treatment with PZQ [63, 64]. Moreover, it was reported in the human Leyte cohort that individuals who produce IgE but not IgG4 in response to rSj97 had 77 % lower re-infection intensity after 12 months >of treatment with PZQ [64, 65]. Sj97 is now in early preclinical testing with process development and further proof of concept studies taking place in mice and water buffalo [13, 64].
Sj26GST
A S. japonicum 26 kDa GST plasmid DNA vaccine (Sj26GST) resulted in a significant reduction in worm numbers and in hepatic and faecal eggs in vaccinated mice; when the DNA vaccine was given in combination with interleukin 18 (IL-18), a potent IFN-γ inducing factor, the protective efficacy was improved [66]. DNA vaccines have some advantages over other types of immunization but they have some limitations concerning the gene delivery system. A recent study reported on a novel nanoparticle formulation of the Sj26GST DNA vaccine; although there was no significant reduction in worm burden, a highly significant decline in tissue egg burden and the fecundity of female adult worms resulted [67]. Toll-like receptor (TLR) 7/8 ligands (e.g. R848) and TLR 9 ligands (e.g. CpG oligodeoxynucleotides, or CpG) as adjuvants can increase vaccine effectiveness through activating the innate immune system and ultimately activating and directing the adaptive immune system. Such adjuvants have been shown to potentiate the activity of the Sj26GST DNA vaccine in mice by increasing splenocyte proliferation, elevating IgG, IgG2a, IFNγ and TNFα levels, and preventing Treg-mediated immunosuppression [68]. In another development, Sj26GST alone, or in combination with fatty acid binding protein (SjFABP), expressed in recombinant pseudorabies virus (PRV) Bartha-K61, induced significant levels of specific immunity and protection in mice and, importantly, sheep, emphasising the potential effectiveness of this live vector for vaccination against schistosomiasis japonica in animal reservoirs [69].
SjTPI
The glycolytic pathway enzyme triose-phosphate isomerase (TPI), found in all stages of the schistosome life-cycle, is another targeted vaccine candidate for schistosomiasis japonica. An early study showed that a S. japonicum (Chinese strain) TPI (SjCTPI) plasmid DNA vaccine (with or without an IL-12 DNA plasmid) protected pigs against challenge infection [70]. Synergistic enhancement of immunogenicity and protection in mice against S. japonicum challenge was achieved with codon optimization and electroporation delivery of the SjTPI DNA vaccine [71], showing a similar level of protection as a replication-defective recombinant optimized SjTPI (rAdV-SjTPI) adenoviral vaccine [72] which was enhanced using a heterologous prime-boost strategy [73]. A study, conducted in Chinese water buffalo with a DNA vaccine encoding SjCTPI alone or fused with bovine HSP-70 with booster immunizations co-administered using a plasmid encoding IL-12, resulted in a significant reduction in worm numbers, liver and faecal eggs and in faecal miracidial hatching [74]. SjCTPI, delivered by a heterologous “prime-boost” regimen, has been used to vaccinate bovines in China as part of a multi-component integrated control package [75].
SjIRs
Schistosoma japonicum possess two types of insulin receptors (SjIRs) which, on binding to mammalian host insulin, can activate the parasite’s insulin pathway, which is pivotal for glucose uptake, growth, and maturation [76]. Recombinant ligands of both S. japonicum insulin receptor 1 and 2 (SjLD1, SjLD2), tested in vaccine/challenge trials in mice resulted in significant reductions in faecal egg output, in reduced mature intestinal eggs and stunting of adult worms [77, 78]. The retardation in growth of the worms likely resulted from reduced glucose uptake [77]. Furthermore, knockdown of the SjIRs using RNA interference (RNAi) resulted in their reduced expression coupled with a reduction in the transcription level of downstream genes within the insulin pathway that are associated with glucose metabolism and schistosome fecundity [79], thereby reinforcing their vaccine potential.
Sj14
In early studies, Sj14 (fatty acid binding protein; SjFABP), the S. japonicum homologue of Sm14, generated no or only a limited level of protection [24], but when given to mice as a DNA vaccine with a plasmid coding for IL-18 as adjuvant, the level of protection was increased substantially [80]. The latter study also showed that SjFABP + IL-18 increased the Th1 immune response by producing a higher level of IFNγ and a lower level of IL-4 compared with mice vaccinated only with SjFABP [80]. Somewhat disappointedly, the Sj14 DNA vaccine, coupled with Sj26GST to form a bivalent DNA-based vaccine, resulted in a reduced level of protective efficacy [81].
Sj23
Sj23, like Sm-TSP-2, a member of the tetraspanin family, is a 23-kDa surface-exposed integral membrane protein expressed in all infective parasite stages. It was shown in BALB/c mice to elicit a rapid humoral immune response dominated by IgG2a antibodies, but not IgG1, and did not provide protection against cercarial challenge after priming with recombinant Semliki forest virus (SFV) particles followed by a boost with recombinant protein [82]. A subsequent report of mice vaccinated with purified recombinant protein LHD-Sj23-GST (large hydrophilic domain of Sj23 fused with Sj26GST) in combination with one of three adjuvants (Freund’s adjuvant (FA), Montanide ISA 206 or Montanide ISA 70 M), and parasite challenged, resulted in high-level production of LHD-Sj23-GST-specific IgG1, IgG2a and IgG3 antibodies and significant reductions in worm burden [83]. In order to further improve on the level of protection, a multivalent DNA vaccine comprising Sj23, glyceraldedyde-3 phosphate dehydrogenase (SjGAPDH), SjFABP and Sj26 was tested in mice which resulted in very high levels of protective efficacy in terms of reduced worms (70.8 % reduction) and liver eggs (60.7 % reduction) [84]. Another study in mice, using three cocktail DNA vaccines encoding Sj23, SjCTPI and NP30, boosted by electroporation in vivo and a protein vaccine boost to this regimen, resulted in a 60 % reduction in adult worm numbers and more than 60 % reduction in the liver egg burden [85].
Details of these and a number of other vaccine candidates tested against S. japonicum challenge infection are presented in Table 2 and elsewhere [78].
New antigen discovery: a way forward
New antigen discovery has been aided by major recent advances in schistosome genomics, transcriptomics and post-genomic technologies [86]. Proteomics is another important and now widely used tool that can identify potential vaccine targets with a focus on the protein constituents of different schistosome sources such as the host-parasite interface comprising tegument or gut [87]. Studies on the tegument have used a number of procedures including biotin-labelling of live parasites and subsequent isolation and characterisation of the biotinylated proteins using tandem mass spectrometry (MS/MS) to identify surface-located proteins, and therefore those accessible to host antibodies. Proteomics of the schistosome gut and its contents has shed new light on the functionality of this important region of the parasite [87]. Coupled with other approaches such as metabolomics, interrogation of the schistosome proteome, particularly the surface, provides a mechanism to identify important clinically-relevant proteins and those having potential as new vaccine targets [87].
Vaccinomics is another powerful innovation which provides a foundation for searching critical determinants of immunity and can promote antigen discovery and the design of novel vaccines for complex pathogens such as the schistosomes [88]. A recent vaccinomics approach for discovering novel schistosome antigens that may not be revealed by conventional proteomics involved the design and manufacture of an immunomics protein microarray, the first to be generated for a multi-cellular pathogen. Mostly surface-derived proteins (215 in total) from S. japonicum and S. mansoni were selected and they were produced using a rapid in vitro translation system, and then printed as a vaccine discovery tool [87, 89, 90]. The reactivity of microarray proteins can be measured with antisera from human patients or schistosomiasis-resistant/exposed animals using a labelled secondary antibody and a laser microarray scanner; highly reactive proteins are then assessed as putative vaccines.
One application of the array used antibodies from acutely- or chronically-infected Chinese individuals with early/advanced schistosomiasis japonica, and subjects exposed, but stool negative for S. japonicum eggs, for screening. This resulted in the identification of 25 immunodominant antigens, including a number of vaccine candidates, transporters, tetraspanin-related proteins, and unannotated proteins [91]. The array has also been screened for IgG subclass and IgE responses, using sera from a human Brazilian cohort of putatively resistant (PR) and chronically S. mansoni-infected (CI) individuals stratified by worm intensity levels (high, medium, low), determined by faecal egg counts, so as to identify antibody signatures reflective of protective vs non-protective immune responses [92]. Probing for IgE responses allowed the identification of antigens that might induce potentially deleterious hypersensitivity reactions if used as subunit vaccines in endemic populations so it was encouraging that the PR individuals did not mount an intense IgE response to these antigens compared with CI subjects [92].
This immunomics-based approach to schistosomiasis vaccine antigen discovery was further validated by the identification of targets of the protective IgG1 immune response in PZQ-induced resistant subjects exposed to S. haematobium; uncharacterized proteins and a number of recognised vaccine antigens (e.g. glucose transporters, tetraspanins, glutathione-S-transferases, calpain) were identified [93]. The same report described the use of sera from rhesus macaques experimentally rendered resistant to S. japonicum infection to screen for antigen targets, and the discovery of new and known vaccine candidates, including many recognized by the human subjects.
Another important application has been the immune screening of the schistosome microarray with antibody secreting cell (ASC)-probes [89, 90, 94], generated from lymph nodes draining the sites of larval S. japonicum migration [95]. This technique is especially advantageous for recognizing antigens with low immunogenicity (selective pressure may have an influence on important protective epitopes which evolve over time with low immunogenicity) or those only temporarily exposed to the immune response [90]. In one study, ASC probes (from skin and lung) and sera from semi-permissive rats and sera from susceptible mice were used to screen the array after infection and re-infection with S. japonicum [94]. A total of 29 antigens, including a number of recognised vaccine candidates and several S. japonicum homologues of human schistosomiasis resistance markers - the tegument allergen-like proteins - were differentially recognized by infected hosts from which eight proteins were prioritized as putative novel schistosome vaccine and diagnostic antigens [94]. In a related study, the protein microarray, screened with ASC probes generated from S. japonicum-infected rats, resulted in the identification of a novel antigen, termed S. japonicum Ly-6-like protein 1 (Sj-L6L-1) which shares structural and sequence features with the Ly-6 protein family and has several other features suggesting it is a promising vaccine candidate against the developing larvae [96].
Conclusion
Schistosomiasis remains a substantial public health problem due to the very high levels of morbidity it causes in many parts of the world. Currently, treatment is entirely dependent on PZQ chemotherapy. As exclusive use of the drug may lead to the emergence of drug resistant strains, development and deployment of a vaccine as part of an integrated approach for prevention and control of schistosomiasisis is to be encouraged. Much of our current understanding of immunity and immune mechanisms against schistosomiasis rely on studies conducted on mice, but vaccines based on studies performed only in the mouse model could have undesirable effects if taken prematurely to human clinical trials. The recent concern raised about using mice for determining the efficacy of vaccine candidates further reinforces the argument that additional critical examination of any identified candidate vaccine antigen, whether or not it has foundation in acquired immunity is essential, and that moving to studies using larger models such as rabbits, pigs or bovines in the case of S. japonicum, or non-human primates for S. mansoni and S. haematobium, is clearly necessary. Furthermore, much of our knowledge regarding immune protection has resulted from studies focused on S. mansoni, so further studies on S. japonicum and, particularly, S. haematobium are needed. Similarly, protection levels of many candidate vaccines show improvement after modification of antigen formulation and improved delivery systems. Combining different genes or antigens can also result in higher levels of vaccine-induced protection. Targeting key biological functions of schistosomes such as tegumental integrity, fecundity, and nutrient uptake using RNAi represent key potential sites to target the parasites for elimination through vaccination [97]. Although in its infancy, CRISPR technology may provide a novel approach identifying specific protein-encoding schistosome genes for vaccine candidate discovery [98].
Schistosomiasis vaccine development has proven highly challenging and costly and new funding is required to promote the generation of a schistosome vaccine antigen pipeline, similar to that in place for many other infectious diseases, and to progress existing promising candidates into clinical trials. It is becoming apparent that mass drug administration alone will not eliminate schistosomiasis and that a vaccine will be an essential component of any future schistosomiasis control intervention toolbox.
Abbreviations
- ASC:
-
Antibody secreting cell
- CI:
-
Chronically-infected
- CPG:
-
5′-C-phosphate-G-3′
- CRISPR:
-
Clustered regularly interspaced short palindromic repeats
- DLC:
-
Dynein light chain
- FABPs:
-
Fatty acid binding proteins
- GLA-AF:
-
Glucopyranosyl lipid A
- GLA-SE:
-
Glucopyranosyl lipid adjuvant stable emulsion
- LHD-Sj23-GST:
-
(large hydrophilic domain of Sj23 fused with Sj26GST)
- MEGs:
-
Microexon genes
- MS:
-
Mass spectrometry
- NP30:
-
Murine monoclonal anti-idiotypic antibody NP30
- ODN:
-
Oligodeoxynucleotide
- PLA:
-
Polylactic acid
- PR:
-
Putatively resistant
- PRV:
-
Pseudorabies virus
- PZQ:
-
Praziquantel
- R848:
-
Resiquimod
- rAdV-SjTPI:
-
Replication-defective recombinant optimized SjTPI adenoviral vaccine
- RNAi:
-
RNA interference
- rSj97:
-
Recombinant Schistosoma japonicum paramyosin
- rSm14:
-
Recombinant Sm14
- SFV:
-
Semliki forest virus
- Sh28GST:
-
Schistosoma haematobium 28 kDa (kilodalton) glutathione S-transferase
- Sj14:
-
SjFABP, Schistosoma japonicum fatty acid binding protein
- Sj23:
-
Schistosoma japonicum 23-kDa surface-exposed integral membrane protein
- Sj26GST:
-
Schistosoma japonicum 26 kDa glutathione S-transferase
- Sj97:
-
Schistosoma japonicum paramyosin
- SjCTPI:
-
S. japonicum (Chinese strain) TPI
- SjFABP:
-
Schistosoma japonicum fatty acid binding protein
- SjGAPDH:
-
Schistosoma japonicum glyceraldedyde-3 phosphate dehydrogenase
- SjIRs:
-
Schistosoma japonicum insulin receptors
- Sj-L6L-1:
-
Schistosoma japonicum Ly-6-like protein 1
- SjLD1, SjLD2:
-
ligands of Schistosoma japonicum insulin receptor 1 and 2
- Sm14:
-
Schistosoma mansoni fatty acid binding protein
- Sm28GST:
-
Schistosoma mansoni 28 kDa glutathione S-transferase
- Sm29:
-
Schistosoma mansoni 29 kDa antigen
- SmCT-SOD:
-
Cu/Zn cytosolic superoxide dismutase signal peptide-containing SOD
- Sm-LAMP:
-
Schistosoma mansoni lysosome-associated membrane glycoprotein
- Smp80:
-
Schistosoma mansoni calpain
- SmSP-SOD:
-
Signal peptide-containing SOD
- Sm-TSP-1:
-
Schistosoma mansoni tetraspanin-1
- Sm-TSP-2:
-
Schistosoma mansoni tetraspanin 2
- SOD:
-
Superoxide dismutase
- TAL:
-
Tegumental Allergen Like
- TDR:
-
The Special Programme for Research and Training in Tropical Diseases
- TLR:
-
Toll-like receptor
- TPI:
-
Triose-phosphate isomerase
- WHO:
-
World Health Organisation
References
Sarvel AK, Oliveira AA, Silva AR, Lima AC, Katz N. Evaluation of a 25-year-program for the control of schistosomiasis mansoni in an endemic area in Brazil. PLoS Negl Trop Dis. 2011;5(3):e990.
World Health Organization. Schistosomiasis. In: Schistosomiasis Fact Sheet No 115. World Health Organization. 2015. http://www.who.int/mediacentre/factsheets/fs115/en/. Accessed Aug 2015.
Nour NM. Schistosomiasis: health effects on women. Rev Obst Gynecol. 2010;3(1):28.
Christinet V, Lazdins-Helds JK, Stothard JR, Reinhard-Rupp J. Female genital schistosomiasis (FGS): from case reports to a call for concerted action against this neglected gynaecological disease. Int J Parasitol. 2016;46(7):395–404.
Cioli D, Pica-Mattoccia L, Basso A, Guidi A. Schistosomiasis control: praziquantel forever? Mol Biochem Parasitol. 2014;195(1):23–9.
Sturrock RF. Schistosomiasis epidemiology and control: how did we get here and where should we go? Mem Inst Oswaldo Cruz. 2001;96(Suppl):17–27.
Ross AG, Olveda RM, Acosta L, Harn DA, Chy D, Li Y, et al. Road to the elimination of schistosomiasis from Asia: the journey is far from over. Microbes Infect. 2013;15(13):858–65.
Hewitson JP, Maizels RM. Vaccination against helminth parasite infections. Expert Rev Vaccines. 2014;13(4):473–87. doi:10.1586/14760584.2014.893195.
Knudsen NP, Olsen A, Buonsanti C, Follmann F, Zhang Y, Coler RN, et al. Different human vaccine adjuvants promote distinct antigen-independent immunological signatures tailored to different pathogens. Sci Rep. 2016;6:19570. doi:10.1038/srep19570.
Lebens M, Sun JB, Czerkinsky C, Holmgren J. Current status and future prospects for a vaccine against schistosomiasis. Expert Rev Vaccines. 2004;3(3):315–28.
Stephenson R, You H, McManus DP, Toth I. Schistosome vaccine adjuvants in preclinical and clinical research. Vaccines. 2014;2(3):654–85.
Diemert DJ, Pinto AG, Freire J, Jariwala A, Santiago H, Hamilton RG, et al. Generalized urticaria induced by the Na-ASP-2 hookworm vaccine: implications for the development of vaccines against helminths. J Allergy Clin Immunol. 2012;130(1):169–76.
Mo AX, Agosti JM, Walson JL, Hall BF, Gordon L. Schistosomiasis elimination strategies and potential role of a vaccine in achieving global health goals. Am J Trop Med Hyg. 2014;90(1):54–60. doi:10.4269/ajtmh.13-0467.
Siddiqui AA, Siddiqui BA, Ganley-Leal L. Schistosomiasis vaccines. Hum Vaccin. 2011;7(11):1192–7.
Merrifield M, Hotez PJ, Beaumier CM, Gillespie P, Strych U, Hayward T, et al. Advancing a vaccine to prevent human schistosomiasis. Vaccine. 2016;34(26):2988–91.
Wilson RA, Li XH, Castro-Borges W. Do schistosome vaccine trials in mice have an intrinsic flaw that generates spurious protection data? Parasit Vectors. 2016;9(1):89.
Tendler M, Simpson AJ. The biotechnology-value chain: development of Sm14 as a schistosomiasis vaccine. Acta Trop. 2008;108(2–3):263–6.
Tendler M, Brito CA, Vilar MM, Serra-Freire N, Diogo CM, Almeida MS, et al. A Schistosoma mansoni fatty acid-binding protein, Sm14, is the potential basis of a dual-purpose anti-helminth vaccine. Proc Natl Acad Sci. 1996;93(1):269–73.
Santini-Oliveira M, Coler RN, Parra J, Veloso V, Jayashankar L, Pinto PM, et al. Schistosomiasis vaccine candidate Sm14/GLA-SE: Phase 1 safety and immunogenicity clinical trial in healthy, male adults. Vaccine. 2016;34(4):586–94.
Tendler M, Almeida M, Simpson A. Development of the Brazilian anti schistosomiasis vaccine based on the recombinant fatty acid binding protein Sm14 plus GLA-SE adjuvant. Front Immunol. 2015;6:218.
Capron A, Capron M, Dombrowicz D, Riveau G. Vaccine strategies against schistosomiasis: from concepts to clinical trials. Int Arch Allergy Immunol. 2001;124(1–3):9–15.
Bergquist NR, Colley DG. Schistosomiasis vaccine:research to development. Parasitol Today. 1998;14(3):99–104.
Bourke CD, Nausch N, Rujeni N, Appleby LJ, Trottein F, Midzi N, et al. Cytokine responses to the anti-schistosome vaccine candidate antigen glutathione-S-transferase vary with host age and are boosted by praziquantel treatment. PLoS Negl Trop Dis. 2014;8(5):e2846.
McManus DP, Loukas A. Current status of vaccines for schistosomiasis. Clin Microbiol Rev. 2008;21(1):225–42.
Ahmad G, Zhang W, Torben W, Noor Z, Siddiqui AA. Protective effects of Sm-p80 in the presence of resiquimod as an adjuvant against challenge infection with Schistosoma mansoni in mice. Int J Infect Dis. 2010;14(9):e781–7.
Ahmad G, Zhang W, Torben W, Haskins C, Diggs S, Noor Z, et al. Prime-boost and recombinant protein vaccination strategies using Sm-p80 protects against Schistosoma mansoni infection in the mouse model to levels previously attainable only by the irradiated cercarial vaccine. Parasitol Res. 2009;105(6):1767–77.
Ahmad G, Zhang W, Torben W, Ahrorov A, Damian RT, Wolf RF, et al. Preclinical prophylactic efficacy testing of Sm-p80-based vaccine in a nonhuman primate model of Schistosoma mansoni infection and immunoglobulin G and E responses to Sm-p80 in human serum samples from an area where schistosomiasis is endemic. J Infect Dis. 2011;204(9):1437–49.
Ahmad G, Torben W, Zhang W, Wyatt M, Siddiqui AA. Sm-p80-based DNA vaccine formulation induces potent protective immunity against Schistosoma mansoni. Parasite Immunol. 2009;31(3):156–61. 29.
Zhang W, Ahmad G, Torben W, Noor Z, Le L, Damian RT, et al. Sm-p80-based DNA vaccine provides baboons with levels of protection against Schistosoma mansoni infection comparable to those achieved by the irradiated cercarial vaccine. J Infect Dis. 2010;201(7):1105–12.
Ahmad G, Zhang W, Torben W, Damian RT, Wolf RF, White GL, et al. Protective and antifecundity effects of Sm-p80-based DNA vaccine formulation against Schistosoma mansoni in a nonhuman primate model. Vaccine. 2009;27(21):2830–7.
Karmakar S, Zhang W, Ahmad G, Torben W, Alam MU, Le L, et al. Cross-species protection: Schistosoma mansoni Sm-p80 vaccine confers protection against Schistosoma haematobium in hamsters and baboons. Vaccine. 2014;32(11):1296–303.
Zhang W, Ahmad G, Le L, Rojo JU, Karmakar S, Tillery KA, et al. Longevity of Sm-p80-specific antibody responses following vaccination with Sm-p80 vaccine in mice and baboons and transplacental transfer of Sm-p80-specific antibodies in a baboon. Parasitol Res. 2014;113(6):2239–50.
Karmakar S, Zhang W, Ahmad G, Torben W, Alam MU, Le L, et al. Use of an Sm-p80-based therapeutic vaccine to kill established adult schistosome parasites in chronically infected baboons. J Infect Dis. 2014;209(12):1929–40.
Braschi S, Borges WC, Wilson RA. Proteomic analysis of the schistosome tegument and its surface membranes. Mem Inst Oswaldo Cruz. 2006;101 Suppl 1:205–12.
Loukas A, Tran M, Pearson MS. Schistosome membrane proteins as vaccines. Int J Parasitol. 2007;37(3–4):257–63.
Beaumier CM, Gillespie PM, Hotez PJ, Bottazzi ME. New vaccines for neglected parasitic diseases and dengue. Transl Res. 2013;162(3):144–55.
Curti E, Kwityn C, Zhan B, Gillespie P, Brelsford J, Deumic V, et al. Expression at a 20 L scale and purification of the extracellular domain of the Schistosoma mansoni TSP-2 recombinant protein: a vaccine candidate for human intestinal schistosomiasis. Hum Vaccin Immunother. 2013;9(11):2342–50.
Zhang W, Li J, Duke M, Jones MK, Kuang L, Zhang J, et al. Inconsistent protective efficacy and marked polymorphism limits the value of Schistosoma japonicum tetraspanin-2 as a vaccine target. PLoS Negl Trop Dis. 2011;5(5):e1166.
Cardoso FC, Pacifico RN, Mortara RA, Oliveira SC. Human antibody responses of patients living in endemic areas for schistosomiasis to the tegumental protein Sm29 identified through genomic studies. Clin Exp Immunol. 2006;144(3):382–91.
Alves CC, Araujo N, dos Santos VC, Couto FB, Assis NR, Morais SB, et al. Sm29, but not Sm22.6 retains its ability to induce a protective immune response in mice previously exposed to a Schistosoma mansoni infection. PLoS Negl Trop Dis. 2015;9(2):e0003537.
Ewaisha RE, Bahey-El-Din M, Mossallam SF, Amer EI, Aboushleib HM, Khalil AM. Combination of the two schistosomal antigens Sm14 and Sm29 elicits significant protection against experimental Schistosoma mansoni infection. Exp Parasitol. 2014;145:51–60.
Mossallam SF, Amer EI, Ewaisha RE, Khalil AM, Aboushleib HM, Bahey-El-Din M. Fusion protein comprised of the two schistosomal antigens, Sm14 and Sm29, provides significant protection against Schistosoma mansoni in murine infection model. BMC Infect Dis. 2015;15:147.
Pinheiro CS, Ribeiro AP, Cardoso FC, Martins VP, Figueiredo BC, Assis NR, et al. A multivalent chimeric vaccine composed of Schistosoma mansoni SmTSP-2 and Sm29 was able to induce protection against infection in mice. Parasite Immunol. 2014;36(7):303–12.
Goncalves De Assis NR, Batistoni de Morais S, Figueiredo BC, Ricci ND, de Almeida LA, da Silva Pinheiro C, et al. DNA Vaccine encoding the chimeric form of Schistosoma mansoni Sm-TSP2 and Sm29 confers partial protection against challenge infection. PLoS One. 2015;10(5):e0125075.
Chura-Chambi RM, Nakajima E, de Carvalho RR, Miyasato PA, Oliveira SC, Morganti L, et al. Refolding of the recombinant protein Sm29, a step toward the production of the vaccine candidate against schistosomiasis. J Biotechnol. 2013;168(4):511–9.
Shalaby KA, Yin L, Thakur A, Christen L, Niles EG, LoVerde PT. Protection against Schistosoma mansoni utilizing DNA vaccination with genes encoding Cu/Zn cytosolic superoxide dismutase, signal peptide-containing superoxide dismutase and glutathione peroxidase enzymes. Vaccine. 2003;22(1):130–6.
Carvalho-Queiroz C, Nyakundi R, Ogongo P, Rikoi H, Egilmez NK, Farah IO, et al. Protective potential of antioxidant enzymes as vaccines for schistosomiasis in a non-human primate model. Front Immunol. 2015;6:273.
Figueiredo BC, Ricci ND, de Assis NR, de Morais SB, Fonseca CT, Oliveira SC. Kicking in the guts: Schistosoma mansoni digestive tract proteins are potential candidates for vaccine development. Front Immunol. 2015;6:22.
Nawaratna SS, Gobert GN, Willis C, Mulvenna J, Hofmann A, McManus DP, et al. Lysosome-associated membrane glycoprotein (LAMP) - preliminary study on a hidden antigen target for vaccination against schistosomiasis. Sci Rep. 2015;5:15069.
Wilson RA, Li XH, MacDonald S, Neves LX, Vitoriano-Souza J, Leite LC, et al. The schistosome esophagus is a ‘hotspot’ for microexon and lysosomal hydrolase gene expression: implications for blood processing. PLoS Negl Trop Dis. 2015;9(12):e0004272.
Pearson MS, Ranjit N, Loukas A. Blunting the knife: development of vaccines targeting digestive proteases of blood-feeding helminth parasites. Biol Chem. 2010;391(8):901–11.
Diniz PP, Nakajima E, Miyasato PA, Nakano E, de Oliveira Rocha M, Martins EA. Two SmDLC antigens as potential vaccines against schistosomiasis. Acta Trop. 2014;140:193–201.
Fukushige M, Mitchell KM, Bourke CD, Woolhouse ME, Mutapi F. A meta-analysis of experimental studies of attenuated Schistosoma mansoni vaccines in the mouse model. Front Immunol. 2015;6:85.
El-Shabasy EA, Reda ES, Abdeen SH, Said AE, Ouhtit A. Transmission electron microscopic observations on ultrastructural alterations in Schistosoma mansoni adult worms recovered from C57BL/6 mice treated with radiation-attenuated vaccine and/or praziquantel in addition to passive immunization with normal and vaccinated rabbit sera against infection. Parasitol Res. 2015;114(4):1563–80.
Reda ES, Ouhtit A, Abdeen SH, El-Shabasy EA. Structural changes of Schistosoma mansoni adult worms recovered from C57BL/6 mice treated with radiation-attenuated vaccine and/or praziquantel against infection. Parasitol Res. 2012;110(2):979–92.
McManus D. Back to the future for schistosome vaccines? Parasite Immunol. 2006;28(6):246–7.
Hsu SY, Xu ST, He YX, Shi FH, Shen W, Hsu HF, et al. Vaccination of bovines against schistosomiasis japonica with highly irradiated schistosomula in China. Am J Trop Med Hyg. 1984;33(5):891–8.
Lin D, Tian F, Wu H, Gao Y, Wu J, Zhang D, et al. Multiple vaccinations with UV- attenuated cercariae in pig enhance protective immunity against Schistosoma japonicum infection as compared to single vaccination. Parasit Vectors. 2011;4:103.
Tian F, Lin D, Wu J, Gao Y, Zhang D, Ji M, et al. Immune events associated with high level protection against Schistosoma japonicum infection in pigs immunized with UV-attenuated cercariae. PLoS One. 2010;5(10):e13408.
Abdel-Hafeez EH, Watanabe K, Kamei K, Kikuchi M, Chen H, Daniel B, et al. Pilot study on interferon-gamma-producing T cell subsets after the protective vaccination with radiation-attenuated cercaria of Schistosoma japonicum in the miniature pig model. Trop Med Health. 2014;42(4):155–62.
Zhang M, Tian F, Gao Y, Ji M, Wu G. Ultraviolet- attenuated cercariae of Schistosoma japonicum fail to effectively induce a Th1 response in spite of up-regulating expression of cytotoxicity-related genes in C57BL/6 mice. J Biomed Res. 2010;24(4):277–84.
Ramirez BL, Kurtis JD, Wiest PM, Arias P, Aligui F, Acosta L, et al. Paramyosin: a candidate vaccine antigen against Schistosoma japonicum. Parasite Immunol. 1996;18(1):49–52.
Leenstra T, Acosta LP, Wu HW, Langdon GC, Solomon JS, Manalo DL, et al. T-helper-2 cytokine responses to Sj97 predict resistance to reinfection with Schistosoma japonicum. Infect Immun. 2006;74(1):370–81.
Jiz MA, Wu H, Olveda R, Jarilla B, Kurtis JD. Development of paramyosin as a vaccine candidate for Schistosomiasis. Front Immunol. 2015;6:347.
Jiz M, Friedman JF, Leenstra T, Jarilla B, Pablo A, Langdon G, et al. Immunoglobulin E (IgE) responses to paramyosin predict resistance to reinfection with Schistosoma japonicum and are attenuated by IgG4. Infect Immun. 2009;77(5):2051–8.
Wei F, Liu Q, Gao S, Shang L, Zhai Y, Men J, et al. Enhancement by IL-18 of the protective effect of a Schistosoma japonicum 26 kDa GST plasmid DNA vaccine in mice. Vaccine. 2008;26(33):4145–9.
Mbanefo EC, Kumagai T, Kodama Y, Kurosaki T, Furushima-Shimogawara R, Cherif MS, et al. Immunogenicity and anti-fecundity effect of nanoparticle coated glutathione S-transferase (SjGST) DNA vaccine against murine Schistosoma japonicum infection. Parasitol Int. 2015;64(4):24–31.
Wang X, Dong L, Ni H, Zhou S, Xu Z, Hoellwarth JS, et al. Combined TLR7/8 and TLR9 ligands potentiate the activity of a Schistosoma japonicum DNA vaccine. PLoS Negl Trop Dis. 2013;7(4):e2164.
Wei F, Zhai Y, Jin H, Shang L, Men J, Lin J, et al. Development and immunogenicity of a recombinant pseudorabies virus expressing Sj26GST and SjFABP from Schistosoma japonicum. Vaccine. 2010;28(32):5161–6.
Zhu Y, Si J, Harn DA, Xu M, Ren J, Yu C, et al. Schistosoma japonicum triose-phosphate isomerase plasmid DNA vaccine protects pigs against challenge infection. Parasitology. 2006;132(Pt 1):67–71.
Zhu Y, Lu F, Dai Y, Wang X, Tang J, Zhao S, et al. Synergistic enhancement of immunogenicity and protection in mice against Schistosoma japonicum with codon optimization and electroporation delivery of SjTPI DNA vaccines. Vaccine. 2010;28(32):5347–55.
Dai Y, Wang X, Zhao S, Tang J, Zhang L, Dai J, et al. Construction and evaluation of replication-defective recombinant optimized triosephosphate isomerase adenoviral vaccination in Schistosoma japonicum challenged mice. Vaccine. 2014;32(7):771–8.
Dai Y, Wang X, Tang J, Zhao S, Xing Y, Dai J, et al. Enhancement of protective efficacy through adenoviral vectored vaccine priming and protein boosting strategy encoding triosephosphate isomerase (SjTPI) against Schistosoma japonicum in mice. PLoS One. 2015;10(3):e0120792.
Da’dara AA, Li YS, Xiong T, Zhou J, Williams GM, McManus DP, et al. DNA-based vaccines protect against zoonotic schistosomiasis in water buffalo. Vaccine. 2008;26(29–30):3617–25.
Gray DJ, Li YS, Williams GM, Zhao ZY, Harn DA, Li SM, et al. A multi-component integrated approach for the elimination of schistosomiasis in the People’s Republic of China: design and baseline results of a 4-year cluster-randomised intervention trial. Int J Parasitol. 2014;44(9):659–68.
You H, Zhang W, Jones MK, Gobert GN, Mulvenna J, Rees G, et al. Cloning and characterisation of Schistosoma japonicum insulin receptors. PLoS One. 2010;5(3):e9868.
You H, Gobert GN, Duke MG, Zhang W, Li Y, Jones MK, et al. The insulin receptor is a transmission blocking veterinary vaccine target for zoonotic Schistosoma japonicum. Int J Parasitol. 2012;42(9):801–7.
You H, McManus DP. Vaccines and diagnostics for zoonotic schistosomiasis japonica. Parasitology. 2015;142(2):271–89.
You H, Gobert GN, Cai P, Mou R, Nawaratna S, Fang G, et al. Suppression of the insulin receptors in adult Schistosoma japonicum impacts on parasite growth and development: further evidence of vaccine potential. PLoS Negl Trop Dis. 2015;9(5):e0003730.
Wei F, Liu Q, Zhai Y, Fu Z, Liu W, Shang L, et al. IL-18 enhances protective effect in mice immunized with a Schistosoma japonicum FABP DNA vaccine. Acta Trop. 2009;111(3):284–8.
Tu Y, Hu Y, Fan G, Chen Z, Liu L, Man D, et al. Protective effects of membrane-anchored and secreted DNA vaccines encoding fatty acid-binding protein and glutathione S-transferase against Schistosoma japonicum. PLoS One. 2014;9(1):e86575.
Jiang N, Cai P, Yin J, Hao L, Lu H, Wang X, et al. Characterization of antibody responses to the Sj23 antigen of Schistosoma japonicum after infection and immunization. Acta Trop. 2010;116(1):9–14.
Zhu Z, Fu Z, Zhang M, Han Y, Hong Y, Li D, et al. Protective efficacy evaluation induced by recombinant protein LHD-Sj23-GST of Schistosoma japonicum emulsified with three different adjuvants. Parasite Immunol. 2012;34(6):341–4.
Zhu L, Liu HF, Lu MB, Long QK, Shi YE, Yu LJ. Construction, purification, and evaluation of multivalent DNA vaccine against Schistosoma japonicum. Parasitol Res. 2011;108(1):115–21.
Dai Y, Zhu Y, Harn DA, Wang X, Tang J, Zhao S, et al. DNA vaccination by electroporation and boosting with recombinant proteins enhances the efficacy of DNA vaccines for schistosomiasis japonica. Clin Vaccine Immunol. 2009;16(12):1796–803.
Ricciardi A, Ndao M. Still hope for schistosomiasis vaccine. Hum Vaccin Immunother. 2015;11(10):2504–8.
Driguez P, McManus DP, Gobert GN. Clinical implications of recent findings in schistosome proteomics. Expert Rev Proteomics. 2016;13(1):19–33.
De Sousa KP, Doolan DL. Immunomics: a 21st century approach to vaccine development for complex pathogens. Parasitology. 2016;143(2):236–44.
Driguez P, Doolan DL, Loukas A, Felgner PL, McManus DP. Schistosomiasis vaccine discovery using immunomics. Parasit Vectors. 2010;3:4.
McWilliam HE, Driguez P, Piedrafita D, McManus DP, Meeusen EN. Novel immunomic technologies for schistosome vaccine development. Parasite Immunol. 2012;34(5):276–84.
Driguez P, Li Y, Gaze S, Pearson MS, Nakajima R, Trieu A, et al. Antibody signatures reflect different disease pathologies in patients with schistosomiasis due to Schistosoma japonicum. J Infect Dis. 2016;213(1):122–30.
Gaze S, Driguez P, Pearson MS, Mendes T, Doolan DL, Trieu A, et al. An immunomics approach to schistosome antigen discovery: antibody signatures of naturally resistant and chronically infected individuals from endemic areas. PLoS Pathog. 2014;10(3):e1004033.
Pearson MS, Becker L, Driguez P, Young ND, Gaze S, Mendes T, et al. Of monkeys and men: immunomic profiling of sera from humans and non-human primates resistant to schistosomiasis reveals novel potential vaccine candidates. Front Immunol. 2015;6:213.
Driguez P, McWilliam HE, Gaze S, Piedrafita D, Pearson MS, Nakajima R, et al. Specific humoral response of hosts with variable schistosomiasis susceptibility. Immunol Cell Biol. 2016;94(1):52–65.
McWilliam HE, Driguez P, Piedrafita D, Maupin KA, Haab BB, McManus DP, et al. The developing schistosome worms elicit distinct immune responses in different tissue regions. Immunol Cell Biol. 2013;91(7):477–85.
McWilliam HE, Driguez P, Piedrafita D, McManus DP, Meeusen EN. Discovery of novel Schistosoma japonicum antigens using a targeted protein microarray approach. Parasit Vectors. 2014;7:290.
Gobert GN, You H, McManus DP. Gaining biological perspectives from schistosome genomes. Mol Biochem Parasitol. 2014;196(1):21–8.
Cai P, Gobert GN, You H, McManus DP. The Tao survivorship of schistosomes: implications for schistosomiasis control. Int J Parasitol. 2016;46(7):453–63.
Tran MH, Pearson MS, Bethony JM, Smyth DJ, Jones MK, Duke M, et al. Tetraspanins on the surface of Schistosoma mansoni are protective antigens against schistosomiasis. Nat Med. 2006;12(7):835–40.
Pearson MS, Pickering DA, McSorley HJ, Bethony JM, Tribolet L, Dougall AM, et al. Enhanced protective efficacy of a chimeric form of the schistosomiasis vaccine antigen Sm-TSP-2. PLoS Negl Trop Dis. 2012;6(3):e1564.
Boulanger D, Reid GD, Sturrock RF, Wolowczuk I, Balloul JM, Grezel D, et al. Immunization of mice and baboons with the recombinant Sm28GST affects both worm viability and fecundity after experimental infection with Schistosoma mansoni. Parasite Immunol. 1991;13(5):473–90.
Cardoso FC, Macedo GC, Gava E, Kitten GT, Mati VL, de Melo AL, et al. Schistosoma mansoni tegument protein Sm29 is able to induce a Th1-type of immune response and protection against parasite infection. PLoS Negl Trop Dis. 2008;2(10):e308.
Martins VP, Morais SB, Pinheiro CS, Assis NR, Figueiredo BC, Ricci ND, et al. Sm10.3, a member of the micro-exon gene 4 (MEG-4) family, induces erythrocyte agglutination in vitro and partially protects vaccinated mice against Schistosoma mansoni infection. PLoS Negl Trop Dis. 2014;8(3):e2750.
El Ridi R, Tallima H, Selim S, Donnelly S, Cotton S, Gonzales Santana B, et al. Cysteine peptidases as schistosomiasis vaccines with inbuilt adjuvanticity. PLoS One. 2014;9(1):e85401.
Ricciardi A, Dalton JP, Ndao M. Evaluation of the immune response and protective efficacy of Schistosoma mansoni Cathepsin B in mice using CpG dinucleotides as adjuvant. Vaccine. 2015;33(2):346–53.
Chlichlia K, Bahgat M, Ruppel A, Schirrmacher V. DNA vaccination with asparaginyl endopeptidase (Sm32) from the parasite Schistosoma mansoni: anti-fecundity effect induced in mice. Vaccine. 2001;20(3–4):439–47.
Figueiredo BC, Assis NR, Morais SB, Ricci ND, Pinheiro CS, Martins VP, et al. Schistosome syntenin partially protects vaccinated mice against Schistosoma mansoni infection. PLoS Negl Trop Dis. 2014;8(8):e3107.
Chen H, Nara T, Zeng X, Satoh M, Wu G, Jiang W, et al. Vaccination of domestic pig with recombinant paramyosin. against Schistosoma japonicum in China. Vaccine. 2000;18(20):2142–6.
Xu X, Zhang D, Sun W, Zhang Q, Zhang J, Xue X, et al. A Schistosoma japonicum chimeric protein with a novel adjuvant induced a polarized Th1 immune response and protection against liver egg burdens. BMC Infect Dis. 2009;9:54. doi:10.1186/1471-2334-9-54.
Qiu C, Liu S, Hong Y, Fu Z, Wei M, Ai D, et al. Molecular characterization of thyroid hormone receptor beta from Schistosoma japonicum and assessment of its potential as a vaccine candidate antigen against schistosomiasis in BALB/c mice. Parasit Vectors. 2012;5:172.
Cao X, Hong Y, Zhang M, Han Y, Wu M, Wang X, et al. Cloning, expression and characterization of protein disulfide isomerase of Schistosoma japonicum. Exp Parasitol. 2014;146:43–51.
Hu C, Zhu L, Luo R, Dao J, Zhao J, Shi Y, et al. Evaluation of protective immune response in mice by vaccination the recombinant adenovirus for expressing Schistosoma japonicum inhibitor apoptosis protein. Parasitol Res. 2014;113(11):4261–9.
Liu J, Wang J, Wang S, Xu B, Liu X, Wang X, et al. Molecular cloning and characterization of Schistosoma japonicum aldose reductase. Parasitol Res. 2013;112(2):549–58.
Wang L, Liu W, Yang M, Peng D, Chen L. Development of a Streptococcus gordonii vaccine strain expressing Schistosoma japonicum Sj-F1 and evaluation of using this strain for intranasal immunization in mice. Parasitol Res. 2013;112(4):1701–8.
Acknowledgement
The authors gratefully acknowledge the National Health and Medical Research Council of Australia for support of their research on schistosomiasis.
Funding
Not applicable.
Availability of data and material
Not applicable.
Authors’ contributions
BMT drafted the article which was modified by all other authors. All authors read and approved the final version of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Tebeje, B.M., Harvie, M., You, H. et al. Schistosomiasis vaccines: where do we stand?. Parasites Vectors 9, 528 (2016). https://doi.org/10.1186/s13071-016-1799-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-016-1799-4