
Abstract
Background
Thermotoga maritima and T. neapolitana are hyperthermophile bacteria chosen by many research teams to produce bio-hydrogen because of their potential to ferment a wide variety of sugars with the highest theoretical H2/glucose yields. However, to develop economically sustainable bio-processes, the culture medium formulation remained to be optimized. The main aim of this study was to quantify accurately and specifically the effect of thiosulfate, used as sulfured nutriment model, on T. maritima growth, yields and productivities of hydrogen. The results were obtained from batch cultures, performed into a bioreactor, carefully controlled, and specifically designed to prevent the back-inhibition by hydrogen.
Results
Among sulfured nutriments tested, thiosulfate, cysteine, and sulfide were found to be the most efficient to stimulate T. maritima growth and hydrogen production. In particular, under our experimental conditions (glucose 60 mmol L−1 and yeast extract 1 g L−1), the cellular growth was limited by thiosulfate concentrations lower than 0.06 mmol L−1. Under these conditions, the cellular yield on thiosulfate (Y X/Thio) could be determined at 3617 mg mmol−1. In addition, it has been shown that the limitations of T. maritima growth by thiosulfate lead to metabolic stress marked by a significant metabolic shift of glucose towards the production of extracellular polysaccharides (EPS). Finally, it has been estimated that the presence of thiosulfate in the T. maritima culture medium significantly increased the cellular and hydrogen productivities by a factor 6 without detectable sulfide production.
Conclusions
The stimulant effects of thiosulfate at very low concentrations on T. maritima growth have forced us to reconsider its role in this species and more probably also in all thiosulfato-reducer hyperthermophiles. Henceforth, thiosulfate should be considered in T. maritima as (1) an essential sulfur source for cellular materials when it is present at low concentrations (about 0.3 mmol g−1 of cells), and (2) as both sulfur source and detoxifying agent for H2 when thiosulfate is present at higher concentrations and, when, simultaneously, the pH2 is high. Finally, to improve the hydrogen production in bio-processes using Thermotoga species, it should be recommended to incorporate thiosulfate in the culture medium.
Similar content being viewed by others

Background
Today, the amount of energy derived from fossils fuels (petroleum, natural gas, and coal) represents about 80% of the world energy consumption. It is henceforth recognized that their use has induced very serious environmental pollutions. The accumulation of the greenhouse-gas carbon dioxide in the atmosphere and the depletion of fossil fuels, altogether with the high prices and the ever-increasing demand have forced most of the countries to start looking for cleaner and renewable energy sources. Therefore, major research efforts focusing on solar and wind energy, geothermal resources, and energy derived from biomass were undertaken to develop new technologies suitable for industrial use.
In this context, although dihydrogen (H2) is not a primary energy source, it is currently seen as a very promising carbonless “energy carrier” which may be used to store energy and provide an efficient alternative to fossil fuels. Up to now, most of the H2 is currently industrially produced by steam reforming of natural gas or by alternative processes based on electrolysis or thermolysis of water [1]. However, in the last decade, biological processes employing bacteria for H2 production have received a significant and increasing attention [2–11]. Indeed, like with other biofuels such as ethanol, butanol, fatty acids, and methane, bio-hydrogen can be produced by processes using living organisms such as green algae (photolysis of water), phototrophic and anaerobic microorganisms (photofermentation of organic acids), and anaerobic fermentative microorganisms (dark fermentation of organic substrates) [2, 7, 8, 11–19]. Among these, due to the large spectrum of catabolic activities of H2-producing microorganisms, dark fermentation is considered as one of the most promising route. The fermentation processes of these microorganisms potentially allow producing hydrogen from renewable energy sources derived from biomass or various carbohydrate-rich waste streams [2, 20–26].
Depending on the microorganism species used, dark fermentations can be performed at either moderate or elevated temperature. In the former case, hydrogen productivities (QH2/time) were generally higher whereas higher yields (YH2/substrate) were reached in the latter [4]. Moreover, the advantages of high versus low temperatures for the bio-hydrogen production include better pathogen destruction, reduced risks of methanogen or acetogen growth (hydrogen consuming), and less sensitivity of hyperthermophiles to the H2 partial pressure [27–30]. However, in order to develop economically sustainable hydrogen-producing bio-processes, both productivities and yields should be significantly increased. Another economic aspect for these bio-processes is the high fresh water requirement, a resource which needs to be preserved. The use of marine microorganisms, which produce hydrogen in seawater from a wide variety of sugars, appears, in this case, to be a promising approach.
Thermotoga spp. are hyperthermophile or thermophile fermentative anaerobic bacteria belonging to a deep-branching lineage within the domain Bacteria [31–34]. They inhabit various hot ecosystems, including hot springs, hydrothermal vents, and oil reservoirs [35–37]. Among Thermotoga species, T. maritima, originally isolated from a geothermal-heated marine sediment at Vulcano, Italy [32], has received considerable interest as potential hydrogen producer [38]. Indeed, T. maritima is able to produce hydrogen with high productivity and yield [29, 39] from a wide variety of sugars, ranging from hexose and pentose monomers to starch and xylan polymers [40]. In addition, compared to the other hydrogen-producing microorganisms, Thermotogales, including T. maritima, exhibit the highest H2 yields, close to the Thauer limit (4 mol H2 per mol glucose). For instance, a yield of 4 mol of H2 per mol of glucose has been reported by Schöder et al. [39] when T. maritima growth was limited by glucose, the energy source, and under very low hydrogen partial pressure (down to 1.3% as hydrogen partial pressure). Actually, T. maritima harvests energy by glycolysis via the Embden–Meyerhof pathway (EMP) as the main route and via the Entner–Doudoroff (ED) pathway to a lesser extent (about 85 and 15% of consumed glucose, respectively) [39, 41]. The ultimate pyruvate-reduction steps resulted in acetate, H2 and CO2 (1:2:1 as molar proportions, respectively) as major end-products of glucose fermentation, and in lactate, alanine, and extracellular polysaccharides (EPS) as minor end-products. The production of these fermentation products was shown to depend on culture conditions (culture medium, nutritional or oxidative stress, operating conditions such as pH, pH2, stirring, Eh,…) [35, 39, 42–46]. In agreement with this classical fermentation model, the H2 yield is optimal (4 H2:2 CO2:2 acetate as molar proportions per mol of glucose or other hexose) only when all the glucose is converted to acetate because the lactate, alanine, and EPS productions are bypassing pathways impairing the H2 production. Furthermore, it has been reported in the literature that Thermotogales reduced sulfur-containing compounds such as elemental sulfur, polysulfide, and thiosulfate to hydrogen sulfide [47–49]. When T. maritima and T. neapolitana, two very closely related species [50], were cultivated in the presence of elemental sulfur, and with glucose as energy source, final cell yields were significantly enhanced but their growth rate remained unaffected [39, 51, 52]. In addition, it has been shown that, in the presence of sulfur, glucose was not more efficiently used. Indeed fermentation–carbon–product pattern remained similar (e.g., acetate and CO2 as major fermentation products and low amounts of lactate), and hydrogen sulfide was produced at the expense of H2 [39]. These findings argue that sulfur reduction was not coupled with energy conservation [39]. In addition, it has been found that (a) hydrogen at high partial pressure inhibited the growth of T. maritima [39], T. neapolitana [16, 29, 53, 54], and other strains of Thermotoga [42, 55], and (b) the presence of sulfur stimulated the growth of these bacteria on glucose at high H2 pressures rather than at low H2 pressures [39]. Finally, these results were consistent with the Huber proposal suggesting that growth stimulation by sulfur reduction in T. maritima, and probably other Thermotogales, was explained as an electron-sink reaction preventing the accumulation of inhibitory concentrations of the fermentation product H2 [32, 35]. In the same way, other studies have shown that, in T. neapolitana and T. maritima, addition of other sulfured compounds, such as cystine, dimethyl sulfide, and thiosulfate, could also relieve the inhibition power of H2 and/or enhanced the cellular growth [48, 49, 51].
With the aim to optimize the growth of T. maritima and to better control its nutritional requirements of sulfur compounds, their effects on growth and glucose catabolism were addressed in this report study. After a comparative study performed in serum bottles, thiosulfate was finally chosen as model among the tested sulfured compounds because of its efficiency for T. maritima growth and its stability at high temperature. In a concentration range between 0 and 0.24 mmol L−1, the effects of thiosulfate on the growth and glucose catabolism were accurately analyzed from batch cultures, performed into a bioreactor, carefully controlled for pH, temperature, and agitation. In addition, in order to prevent the inhibition of T. maritima cultures by hydrogen, the bioreactor was equipped with a specific device allowing controlling and maintaining the hydrogen partial pressure in the bioreactor headspace below the critical limit which was determined under our experimental conditions.
Methods
Strain and medium for routine cultures
Thermotoga maritima MSB8 was obtained from the DSMZ (DSM 3109T). The basal culture medium (BM) used for growth was prepared using anaerobic techniques as developed by Hungate and Macy [56, 57]. It contained (per liter): NaCl 20 g, yeast extract (Fluka Biochemical, Spain) 1 g, NH4Cl 0.5 g, KH2PO4 0.3 g, K2HPO4 0.3 g, MgCl2 0.2 g, KCl 0.1 g, CaCl2 0.1 g, and Balch trace mineral element solution [58] 10 mL. The medium was adjusted to pH 7.0 with 1 mol L−1 KOH and then boiled and cooled down to room temperature under a stream of O2-free N2. It was then distributed into 100 mL serum bottles (35 mL of medium) as previously described [59]. After sealing the serum bottles, the gaseous phase was flushed with a stream of O2-free N2:CO2 (80:20%) for 30 min. The medium was then autoclaved at 120 °C for 20 min and stored at room temperature. Before inoculation, the culture medium was supplemented with 0.75 mL of NaHCO3 (100 g L−1) and 0.75 mL of glucose (1 mol L−1). After inoculation with 1 mL of overnight T. maritima culture, final concentrations of NaHCO3 and glucose in the culture medium were 2 g L−1 and 20 mmol L−1, respectively. All T. maritima cultures performed in serum bottles were incubated at 80 °C. The stock solutions of NaHCO3 and glucose were prepared under anoxic conditions as described by Miller and Wolin [59], and stored under N2:CO2 (80:20%). The glucose solution was sterilized by filtration and the NaHCO3 solution by autoclaving (120 °C for 20 min).
Culture media for the experiments concerning the study of the sulfur compounds
Stock solutions of DMSO, Na2S, methionine, thiosulfate, elemental sulfur, and cysteine were sterilized by filtration and distributed into 100 mL serum bottles. Anoxia was obtained by flushing the bottle headspaces with an O2-free N2 gas stream for 30 min. DMSO, Na2S, methionine, cysteine, and elemental sulfur stock-solution concentrations were 14 mmol L−1 and thiosulfate stock-solution concentration was 7 mmol L−1. To study sulfur compounds, 100 mL serum bottles, containing 19 mL of basal culture medium (BM) prepared under O2-free N2:CO2 (80:20%), were used. Before inoculation, the culture medium was supplemented with 0.45 mL of NaHCO3 (100 g L−1), 0.55 mL of glucose (1 mol L−1), and 0.5 mL of sulfured compound stock solution. After inoculation with 1 mL of overnight T. maritima culture, final concentrations of NaHCO3, glucose, and sulfured compound in the culture medium were 2 g L−1, 25, and 0.3 mmol L−1 sulfur equivalent, respectively. In this study, two successive cultures, incubated at 80 °C, were done in triplicate for each sulfured compound.
Culture media for the bioreactor experiments
The basal medium BM for the bioreactor contained (per liter): NaCl 20 g, Yeast extract (Fluka Biochemical, Spain) 1 g, NH4Cl 0.5 g, KH2PO4 0.3 g, K2HPO4 0.3 g, MgCl2 0.2 g, KCl 0.1 g, CaCl2 0.1 g, and Balch trace mineral element solution 10 mL [58]. The BM medium was supplemented with glucose (25 or 60 mmol L−1) and thiosulfate at various concentrations ranging between 0 and 0.24 mmol L−1. The medium was adjusted to pH 6.5 with 1 mol L−1 KOH. Fifteen liters of medium were prepared routinely in a tank (20 L), autoclaved at 120 °C for 45 min, and then cooled down to room temperature under an O2-free-N2 gas stream. The medium tank was then connected, under sterile conditions, to the feed pump of the bioreactor, and maintained continuously under a stream of O2-free N2.
Experimental material and bioreactor
Thermotoga maritima was batch cultured in a 2.3 L double-jacket glass bioreactor (FairMenTec, France) with a 1.5 L working volume. The fermentor was run with stirring driven by two axial impellers, and was equipped with sensors to monitor temperature (Prosensor pt 100, France), pH (Mettler Toledo InPro 3253, Switzerland), and redox potential (Mettler Toledo InPro 3253, Switzerland). The incoming gas stream (O2-free N2 or O2-free N2 and H2), prepared via one or two mass-flow meters (Bronkhorst, range 0–100 or 0–100 and 0–10 mL min−1, Netherland, respectively), was injected through a nozzle immersed in the bioreactor. The steam in the outgoing gas stream was condensed in a water-cooler glass exhaust [temperature controlled at 4 °C with a cooling bath equipped with a pump (Julabo SE 6, France)] to prevent liquid loss in the bioreactor (water–vapor condensates were returned to the culture vessel). On the outgoing gas streamline, downstream from the water-cooler glass exhaust, a micro-GC, equipped with a catharometric detector (MS5A, SRA Instrument, France), a GC-FPD, equipped with a flame photometric detector (PR 2100, Perichrom, France), and a CARBOCAP CO2 probe (Vaisala GMT 221, Finland), allowed online measurement of H2, N2 (micro-GC), H2S (GC-FPD), and CO2 (Probe) contents (see below for the analytic conditions). To prevent air from entering the bioreactor, the outgoing gas streamline was closed off by a hydraulic seal (2 cm deep immersion in oil). The bioreactor was heated by hot-water circulation in the double jacket using a heated bath equipped with a pump (Julabo F25, France). Bioreactor liquid volume and NaOH consumption, which was used to regulate culture pH, were followed via two scales [Sartorius Combics 1 and BP 4100 (France), respectively]. Temperature, pH, gas stream flow rates, and stirrer speed were regulated through control units (local loops). The bioreactor was connected to two pumps dedicated to the supply of fresh culture medium and to empty the reactor. All this equipment was connected to a Wago PLC (France) via a serial link (RS232/RS485), a 4–20 mA analog loop or a digital signal. The PLC was connected to a computer for process monitoring and data acquisition. BatchPro software (Decobecq Automatismes, France) was used to monitor and manage the process with good flexibility and total traceability.
Operating conditions for the bioreactor
Before each series of fermentation cycles, the reactor was dismantled, washed, and sterilized by autoclaving at 120 °C for 30 min. One series comprised about 15 successive fermentation cycles. One fermentation cycle included three steps: the reactor feeding, the fermentation phase, and the reactor emptying.
For each experimental condition tested, three or four successive fermentation cycles were carried out. In general, the two or three last ones were reproducible for growth and fermentation patterns.
Description of the three steps of a fermentation cycle:
Step 1: Reactor feeding. The reactor was filled with 1.4 L of fresh basal medium BM supplemented with glucose and thiosulfate depending on the experiments. During the filling step, an incoming gas stream (O2-free N2) adjusted at 500 mL min−1 was used to maintain the anoxia in the bioreactor.
Step 2: Fermentation phase. For the first fermentation cycle of a series, the bioreactor inoculation was performed with 100 mL of a recent T. maritima culture coming from serum bottles. For the next fermentation cycles, 100 mL of the previous fermentation (n − 1) was kept in the bioreactor to inoculate the current fermentation cycle (n).
For the fermentation phase, temperature and stirrer speed were regulated at 80 ± 0.5 °C and 350 ± 5 rpm, respectively. pH was regulated at 7.0 ± 0.1 by adding 1 mol L−1 NaOH. pH and redox probes were calibrated separately at 80 °C with pH −4.22 and −7.04 buffers (Mettler Toledo, Switzerland) and a 124 mV redox buffer at pH 7.0 (Mettler Toledo, Switzerland), respectively. The probe calibrations were verified after each series of fermentation cycles (about every 15 fermentation cycles).
At the beginning of the fermentation phase (step 2), the incoming gas stream (O2-free N2) was adjusted initially at 10 mL min−1 and maintained until the hydrogen percentage in the bioreactor outgoing gas reached the set point of 5%. When the set point was reached, the fermentation process controlled the debit of the incoming gas stream (O2-free N2) to maintain the hydrogen into the outgoing gas at 5% until the end of the fermentation phase.
Regarding the preliminary experiments, focusing on the effects of partial pressure of hydrogen on growth and glucose catabolism in T. maritima, different incoming gaseous mixtures (H2/N2) (v/v), in a range of (85/15) to (1/99), were used to perfuse the culture. For these experiments, the debit of the incoming gaseous mixtures was constant and adjusted at 50 mL min−1 during the fermentation phase.
Step 3: Reactor emptying. The end of the fermentation phase (step 2) was characterized by a decrease of the regulated debit of the incoming gas stream (O2-free N2) due to the hydrogen production weakening by glucose starvation. Thus, when this debit fell below 15 mL min−1, the process triggered the emptying phase of the bioreactor, consisting in removing 1.4 L of culture from the bioreactor, leaving 100 mL to be used as inoculum for the next fermentation cycle. The process was therefore ready for a new cycle.
Analytical methods
All growths of T. maritima were followed by measuring optical density (OD). OD was determined in triplicate at 600 nm with a S2100 Diode array UV–Visible spectrophotometer (WPA Biowave, France). Cell dry weight was determined as one unit OD corresponding to 330 mg L−1.
As described earlier, the gas produced during fermentation runs were analyzed continuously with a micro-GC, a GC-FPD, and a CO2 probe. Regarding the micro-GC, dedicated to H2 and N2 measurements, the temperatures of the injector, the column, and the detector were adjusted to 90, 100, and 100 °C, respectively. The pressure of argon, used as carrier gas, was 200 kPa. The gas analysis was repeated every 2 min. The chromatogram treatments were performed by SOPRANE software (SRA Instrument, France).
Regarding the GC-FPD, dedicated to the measure of the H2S present in the out-coming gas, the column was a capillary RESEK RTX-1 and the detector was a flame photometer. Operating conditions were as follows: the gradient for oven-temperature increase was adjusted from 50 to 200 °C with a rate of 15 °C per minute, the temperatures of injector and detector were adjusted to 180 and 230 °C, respectively. The pressure of helium, used as carrier gas, was 60 kPa. The frequency of sample-injection gas was set every 20 min and the chromatogram treatments were performed via the WINILAB III software (Perichrom, France).
Glucose, acetate, lactate, and fructose concentrations were determined by HPLC as follows: 1 mL of culture sample was centrifuged for 5 min at 14500 rpm, and 20 L was then loaded onto an Animex HPX-87H column (Biorad) set at 35 °C, and eluted at 0.5 mL min−1 with a H2SO4 solution (0.75 mmol L−1). The product concentrations were determined with a differential refractometer detector (Shimadzu RID 6 A, Japan) connected to a computer running WINILAB III software (Perichrom, France). All analyses were performed in triplicate. l-alanine concentrations in centrifuged culture samples were determined by HPLC as described by Moore et al. [60]. Microbial extracellular polysaccharides (EPS) were quantified in centrifuged culture sample by the colorimetric method described by Dubois et al. [61]. Throughout this paper, EPS values were converted in glucose equivalent and expressed in mmol L−1.
Determination of H2 and CO2 production rates
During the experiments, the data of N2 debits and the gas analyses (N2, H2, and CO2) were recorded and used to calculate the fluxes of CO2 and H2, which then led to the cumulative amounts of CO2 and H2 produced in the bioreactor.
To determine the production of H2 and CO2, we used a mathematical model based on the material balances of the 3 gaseous compounds (N2, H2, and CO2):
Here \(p_{{{\text{N}}_{2} }}^{\text{out}}\), \(p_{{{\text{CO}}_{2} }}^{\text{out}},\) and \(p_{{{\text{H}}_{2} }}^{\text{out}}\) are the partial pressures, in the outlet-gas stream, of N2, CO2, and H2, respectively, \(p_{{{\text{N}}_{2} }} \; = \;p_{{{\text{CO}}_{2} }} \; = \;p_{{{\text{H}}_{2} }} \; = \;\;100\%\) are the partial pressures of N2 (carrier gas), CO2, and H2 (biological gas produced during fermentation), respectively, \(V_{\text{HR}}\) (960 mL) is the bioreactor headspace volume, and \(V_{\text{Steam}}\)(320 mL) is water–vapor volume. Vapor volume was calculated according to the Antoine equation at 69 °C (median headspace temperature during the fermentation run). \(Q_{{{\text{N}}_{2} }}\), \(Q_{{{\text{CO}}_{2} }}\), \(Q{}_{{{\text{H}}_{ 2} }},\) and \(Q_{T}^{\text{out}}\) are the N2, CO2, and H2 flows and the sum of these three gases, respectively. At 69 °C (headspace temperature), carrier gas flow rate \(Q_{{{\text{N}}_{2} }}\) was calculated as follows:
.
To determine the total production of CO2, ([CO2]aq + \([{\text{HCO}}_{3}^{ - } ]\) + \([{\text{CO}}_{3}^{2 - } ]\)) in the liquid phase of the bioreactor was estimated by the following relations:
Here pCO 2 is the partial pressure of CO2 in the headspace of the bioreactor.
Results and discussion
Before carrying out our study on the sulfur-compound effects on T. maritima growth, a specific formulation for the growth medium was determined. In order to emphasize the impact of these compounds, the glucose and yeast extract concentrations were determined in such a way that the bacterial growth was limited only by the nutrients present in the yeast extract. This point was essential because of the presence, in the yeast extract, of sulfur compounds such as cystine and methionine. Based on the experiments performed in the bioreactor (see Figs. 1 and 2 presented in the part II) [62], it was established that, in the presence of a glucose excess (concentrations greater than 20 mmol L−1), T. maritima growth was only limited by the yeast extract for concentrations ranging from 0 to 1 g L−1. Consequently, we have chosen, for all the following experiments, to use a culture medium containing 1 g L−1 of yeast extract with 25 and 60 mmol L−1 glucose for fermentation runs performed in serum bottles and bioreactor, respectively.

Comparisons of different sulfured nutriments on Thermotoga maritima cultures. All sulfured compounds were added at the rate of 0.3 mmol L−1 sulfur equivalent in a medium containing glucose (25 mmol L−1), yeast extract (1 g L−1), and salts (see “Methods” section). DMSO, S°, and Na2S meant dimethyl sulfoxide, elementary sulfur, and sodium sulfide, respectively. Productivities of cells (Qcells), and of hydrogen (QH2), and consumption of glucose (Qglu) were calculated during the growth phase (after 14.5 h of incubation). All batch cultures were performed in triplicate in serum bottles. The control corresponded to T. maritima grown without adding the sulfured compounds

Cellular and hydrogen productivities versus hydrogen partial pressure. Productivities of T. maritima cells (Qcells) were expressed in mg per liter of medium and per hour. Glucose consumption rates (Qglu) were expressed in mmol per liter of medium and per hour. These fermentation parameters were calculated during the growth phase from fermentation runs performed in bioreactor (in triplicate). For all fermentations, culture medium contained glucose (60 mmol L−1) and yeast extract (1 g L−1)
Effects of several sulfur compounds on T. maritima growth performed in serum bottles
Dimethyl sulfoxide (DMSO), methionine, cysteine, thiosulfate, elementary sulfur, and sodium sulfide (Na2S), at a concentration of 0.3 mmol L−1 equivalent sulfur, were tested as sulfured sources on T. maritima cultures. These cultures were performed in serum bottles under anoxic conditions in the presence of glucose as energy source.
For all the cultures, results of C-recovery ranged from 88.1 to 96.2%, showing that almost all of the carbon of the fermented glucose was recovered as lactate, acetate, and CO2, the latter being estimated by considering that one mole of CO2 was produced per mole of acetate (Table 1). In addition, the levels of the molar ratios “H2/acetate,” found to be close to 2.0 (between 1.7 and 2.2) (Table 1), indicated that the hydrogen and acetate produced were correctly measured.
To evaluate the effect of the various sulfur compounds on growth and fermentation of glucose, three parameters, cellular production rates (Qcells), glucose consumption rates (Qglu), and hydrogen production rates (QH2), calculated during the first 14.5 h of fermentation (growth phase), were used (Fig. 1).
The results in Fig. 1 showed that, by comparison with the control culture (culture grown without adding the sulfured compounds) the presence of DMSO had no significant effect on T. maritima growth and glucose fermentation. In contrast, the presence of the five other sulfur compounds—elementary sulfur, methionine, thiosulfate, cysteine, and sodium sulfide—accelerated T. maritima growth and fermentation, as indicated by the increasing of Qcells, Qglu, and QH2. Finally, among these compounds, thiosulfate, cysteine, and sodium sulfide were significantly the most efficient (Fig. 1).
Childers et al. [51] already reported beneficial effects of sulfur compounds such as cystine, dimethylsulfide, or elementary sulfur on T. neapolitana growth, a species close to T. maritima [51]. Similarly, it was reported that most Thermotoga species, including T. maritima, were able to reduce thiosulfate and/or elementary sulfur into H2S and that their growths were enhanced in the presence of thiosulfate and/or elementary sulfur [32, 35, 42, 47, 48, 50]. So far, it is generally admitted that the improving of Thermotoga species growth, due to the reductive process of sulfur compounds (elementary sulfur, thiosulfate, cystine, dimethyl sulfide, or polysulfur) into sulfide, was the result of a detoxifying process preventing H2 accumulation, a powerful inhibitor for growth, rather than an energy-yielding electron-sink reaction [32, 35, 39, 47]. However, in our experimental conditions, the idea of a “detoxifying process preventing H2 accumulation” cannot be retained to explain the observed enhancing of T. maritima growth since the concentration of thiosulfate (0.15 mmol L−1) was too low. Indeed, if the added thiosulfate was only used for oxidizing H2, 0.15 mmol L−1 of thiosulfate could only allow oxidizing 0.6 mmol L−1 of H2 (4 mol of H2 are necessary to reduce 1 mol of thiosulfate into sulfide). This amount of oxidized hydrogen (0.6 mmol L−1) appears insignificant in comparison to the 47.3 mmol L−1 of hydrogen produced during the fermentation of glucose (Table 1). In consequence, given the small amount of hydrogen potentially oxidized by the thiosulfate, the enhancing of T. maritima growth cannot be attributed to a detoxification concept. The only remaining valid assumption would be that thiosulfate should be considered, under our experimental conditions, as a sulfur source dedicated to the anabolism and thus to the synthesis of cellular components such as proteins.
Experiments performed in bioreactor
To deepen and specify the stimulating effect of sulfur-compound low concentrations on the fermentation of glucose by T. maritima, thiosulfate was selected as the sulfur source for all subsequent experiments in the bioreactor. In contrast to Na2S and cysteine, thiosulfate is both non-volatile and thermally stable under our experimental conditions (temperature and pH were controlled at 80 °C and 7.0, respectively). In addition, to minimize as much as possible, the inhibitory effects of hydrogen on growth and metabolism of T. maritima, the reactor was equipped with a specific device to control the hydrogen partial pressure in the headspace (see “Methods” section).
Determination of the critical limit of hydrogen partial pressures (pH2) to prevent the inhibition of T. maritima cultures
In order to determine the technical specifications of the pH2 control, enabling, at the same time, on one hand, not to inhibit the T. maritima cultures, and, on the other hand, to estimate correctly the hydrogen and CO2 productions, several fermentation runs were performed varying the pH2, in the bioreactor headspace, within a range from 7 to 607 mbar at 80 °C. To perform these experiments, the bioreactor was fed with different incoming gaseous mixtures (H2/N2) used at a constant total debit (50 mL min−1).
The results presented in Fig. 2 showed that the cellular production rates (Qcells) and the glucose consumptions rates (Qglu) were unaffected when pH2 was maintained in a range of 7.1–178.5 mbar. In contrast, the providing of H2 at a partial pressure of 607 mbar within the bioreactor headspace caused an approximate twofold decrease in Qglu and Qcells levels (Fig. 2).
These results were different from those reported by Schroder et al. [39], which have shown that T. maritima growth was reduced even for hydrogen partial pressures as low as 28 mbar. On the contrary, our results were totally consistent with numerous authors who concluded that (1) a pH2 lower than 200 mbar is required for a fully functional hydrogen-producing reactor in which the growth was optimal [44, 63–65], and (2) a pH2 estimated at 2900 mbar at 85 °C was completely inhibiting T. maritima growth [32].
Furthermore, as already mentioned in literature [39, 42], our results showed that the hydrogen partial pressure increase, within the bioreactor, led to a shift of the glucose catabolism from acetate towards lactate (molar ratios lactate/glucose increased from 0.5 to 0.8 and molar ratios acetate/glucose changed inversely from 1.3 to 1.0 at 7 and 607 mbar of pH2, respectively) (Table 2).
Moreover, it is noteworthy that, whatever the pH2 tested, the fermentation of glucose by T. maritima led to a significant reduction of the culture medium as indicated by the decrease of Eh measurements during the growth phases (between t 1 and t 2) (Table 2). In addition, the results showed that the initial Eh measurements (at t 0 before starting the fermentation phase) were conversely correlated to the level of pH2 (Eh measurements at t0 decreased from −130 to −239 mV when pH2 increased from 7 to 607 mbar, respectively) (Table 2). Taken together, these results indicated that the phenomenon of reduction of the culture medium was due to both biological and chemical activities. As discussed in a previous study [43], this capacity of T. maritima to reduce the culture medium by itself suggested that the reducing compounds such as cysteine and/or Na2S usually added in anaerobic medium cultures for Thermotogales growth were unnecessary.
In addition, concerning the estimates of H2 and CO2 productions, the monitoring of the fermentation runs, carried out with an incoming gaseous mixture (N2/H2) containing 85% of hydrogen (pH2 at 607 mbar), has revealed that, under these conditions, the hydrogen production accuracy was insufficient. In contrast, for the other fermentation runs, fed with gaseous mixtures containing less than 25% of H2 (batches with pH2 lower than 179 mbar), both measures and method to calculate H2 and CO2 productions (“Determination of H2 and CO2 production rates” in the “Methods” section) were correct as confirmed by molar ratios H2/CO2 and CO2/acetate close to 2 and 1, respectively (corresponding to T. maritima glucose catabolism [39]) (Table 2).
In accordance with our results, we have therefore chosen, for all the following fermentation runs, to control pH2 at a maximal value of 35 mbar, corresponding to a maximum of 5% (v/v) H2 in the gaseous outflow of the bioreactor (for more details on the pH2 control see paragraph ≪Operating conditions for the bioreactor≫ in the “Methods” section). Under these experimental conditions, our results showed that hydrogen and CO2 productions were correctly estimated, and growth and glucose catabolism were unaffected by H2.
Thermotoga maritima growth in culture medium limited by thiosulfate
The effects of the limitation of thiosulfate, as the main sulfur growth nutriment, were studied on the growth and on the pattern of glucose fermentation products in T. maritima. Fermentation runs were performed varying the thiosulfate concentration within a range from 0 to 1.0 mmol L−1, in the bioreactor, with a culture medium containing yeast extract and high concentrations of glucose (1 g L−1 and 60 mmol L−1, respectively).
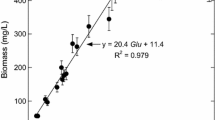
Under our experimental conditions, the thiosulfate concentration had a significant effect on the maximal cellular concentration obtained during fermentation runs (“cells” values reported at t 2 in Table 3). Figure 3 shows that the cellular growth was limited by thiosulfate only when its concentration was lower than 0.06 mmol L−1. For higher concentrations up to 1.0 mmol L−1, cellular concentration approached an upper limit of 400 mg L−1, probably revealing other nutritional limitations, or back-inhibition by fermentation products such as acetate and lactate. In thiosulfate-limited batch cultures (concentrations ranging from 0 to 0.06 mmol L−1), the cellular yield on thiosulfate (Y X/Thio) could be determined at 3617 ± 176 mg (cell dry weight) of cells per mmol of thiosulfate initially present in the culture medium (Fig. 3). Otherwise, the culture, performed in mineral medium (absence of yeast extract) with glucose as sole carbon and energy source, showed that thiosulfate was essential for T. maritima growth (Fig. 3). Under these culture conditions, the cellular concentrations reached 27 mg L−1 with 0.03 mmol L−1 of thiosulfate, up to a limit of approximately 40 mg L−1 with 1 mmol L−1 (Fig. 3). The cellular productions, obtained with 1 g L−1 of yeast extract (about 128 mg L−1 of cells as maximum concentration) (Table 3) without adding thiosulfate, could be explained by the sulfured compounds, such as cystine and methionine, present in the yeast extract. In addition, it could be concluded that among all the nutrients provided by the yeast extract, the sulfur compounds were likely those limiting T. maritima growth. Finally, the extrapolation of the linear regression presented in Fig. 3 suggested that the sulfur nutrients present in 1 g L−1 of yeast extract were equivalent at 0.03 mmol L− 1 of thiosulfate in terms of effect on the growth of T. maritima. On the other hand, as shown in Fig. 3, the significant offset between the cellular concentrations obtained in the presence and absence of yeast extract, with thiosulfate in excess (concentrations greater than 0.2 mmol L−1), unambiguously indicated that the yeast extract also provided, in addition to sulfured compounds, some specific nutritional factors strongly stimulating T. maritima growth. These compounds could be some vitamins as reported by Childers et al. [51] rather than amino acids, demonstrated to be poorly used by T. maritima [46].

Maximum cellular concentrations versus thiosulfate concentrations. In the absence of yeast extract, maximal biomass is obtained with glucose (25 mmol L−1) as energy source. The cultures were performed in triplicate in serum bottles after nine subcultures under the same conditions. In the presence of yeast extract at 1 g L−1, maximal biomass is obtained with glucose (60 mmol L−1). These cultures were performed in triplicate in bioreactor
In order to evaluate the sulfur quantity from thiosulfate incorporated in the cellular material, the quantities of sulfur present both in the cells and originally in the culture medium were calculated for two specific culture conditions (Table 4). The first condition corresponds to organic sulfur-limited growth provided by 1 g L−1 of yeast extract (medium without adding thiosulfate). The second condition corresponds to a growth limited by the sulfur provided by both 0.06 mmol L−1 of thiosulfate and 1 g L−1 of yeast extract as shown in Fig. 3. The amount of organic sulfur (found in the form of cystine, a cysteine dimer, and methionine) present in 1 g L−1 yeast extract was calculated from the yeast extract composition provided by Sigma (0.008 mmol of cystine and 0.054 mmol of methionine or about 0.07 mmol of organic sulfur (S-YE) per gram of yeast extract). Both these two amino acids can be assimilated by T. maritima as previously shown (Fig. 1). The amount of sulfur present in 0.06 mmol of thiosulfate represented 0.12 mmol (S-thiosulfate). Finally, cellular sulfur (S-cells) was calculated from the elemental composition of T. maritima cells given by Kelly et al. (C1 H1.6 O0.6 N0.2 S0.005) [46] and from the maximum cell concentrations obtained under both culture conditions (Tables 3 and 4).
From these results, in the absence of thiosulfate, the cells incorporated 35% (Table 4) of the organic sulfur initially present in the yeast extract. Considering that organic sulfur is the growth-limiting element, it can be estimated that the remaining 65% of the S-YE are not accessible to the cells.
In the presence of thiosulfate, the calculations show that 36% of the initial sulfur (S-YE and S-thiosulfate) have been incorporated into the cellular material (Table 4). Two options concerning the origin of the S-cell can be considered. Either both the yeast extract and the thiosulfate contributed to the S-cell, or the thiosulfate was the only sulfur source. In the first case, if the assimilable S-YE was primarily incorporated into the biomass, the remaining S-cell would correspond to 36% of the initial S-thiosulfate. In the second case, 56% of the thiosulfate was incorporated into the cellular material. Therefore, in both cases, the fraction of the S-thiosulfate incorporated in the cellular material is lower than 56% of the initially present S-thiosulfate. Given the dissymmetry of the oxidation level of the two sulfurs in thiosulfate (S–SO3 −), these observations would suggest that only one of the two sulfurs would actually be incorporated into the cellular material.
In Table 3, the analysis of carbon balances, obtained for various thiosulfate concentrations, showed that carbon recovery decreased drastically when thiosulfate concentrations limited T. maritima growth (e.g., inferior to 0.06 mmol L−1). In the absence of thiosulfate, the production of cells, lactate, acetate, and CO2 represented about only 54.9% of the carbon from the consumed glucose versus 78–87% for fermentation runs performed with more than 0.06 mmol L−1 of thiosulfate. The additional analyses of culture fermentation products, performed in the absence of thiosulfate and in the presence of 0.12 and 0.24 mmol L−1 of thiosulfate, revealed the presence of l-alanine (1.39, 3.8, and 3.8 mmol L−1, respectively) and extracellular polysaccharides (EPS) (4.7, 3.6, 2.9 mmol L−1 in equivalent glucose). Taking into account the carbon, represented by l-alanine and EPS, for these three culture conditions (0.0, 0.12, and 0.24 mmol L−1 of thiosulfate), the levels of C-recoveries (C2-recoveries) (87.2, 92.2, and 95.8%) concluded that almost all of the carbon from the fermented glucose was recovered. Under these three culture conditions, the production of l-alanine expressed as a percentage (c-alanine/fermented c-glucose) evolved almost constantly (between 4.2 and 4.6%). This result was entirely consistent with those published in the literature. Indeed, it should be noted that, in T. maritima, alanine production never represented more than 4–5% of the carbon from the consumed carbohydrate, whatever the evaluated growth conditions [43, 46, 49]. In contrast, EPS production, expressed as a percentage (C-EPS/C-glucose fermented), increased considerably in the absence of thiosulfate (28.6, 8.5, 7.0% versus 0.0, 0.12, and 0.24 mmol L−1 of thiosulfate, respectively). It should be noted that EPS are probably underestimated. Indeed, when preparing the samples for the EPS assay, the cultures were centrifuged to remove the cells which also eliminated the fraction of the EPS that is trapped with the cells. This EPS overproduction, revealed in the absence of thiosulfate, is not surprising since EPS could account for up to more than 20% of the carbon from the consumed carbohydrate depending on culture conditions [43, 46].
It must be emphasized that, in T. maritima, EPS production was associated with stress conditions such as oxidative stress or deficiency in ammonium [43, 45, 46]. Indeed, they should play a significant role in the defense strategy employed by this species and other anaerobes to cope with unfavorable environmental constraints [45, 46, 66, 67]. From this perspective, our results demonstrated, for the first time, that, in T. maritima, the deficiency of sulfur nutriment should be also considered as a stress condition marked by a stimulation of EPS production as an end-product of glucose fermentation.
In T. maritima cultures, performed under oxidative stress by oxygen [43] or under nutritional stress by sulfur nutriment deficiency (here the thiosulfate), both acetate and lactate yields were found to decline concomitantly in favor of the EPS yields. Interestingly, the major difference between patterns for the end-products of glucose fermentation in these two under-stress cultures was marked by the change in proportions between lactate and acetate molar yields. Indeed, T. maritima grown under oxidative stress by oxygen [43] showed an additional shift of glucose catabolism towards lactate, where 0.8 mol of lactate was produced per mol of acetate [43]. Although the limitation of T. maritima growth by thiosulfate was found to decrease drastically both acetate and lactate yields down to a low limit (0.7 and 0.3 mol mol−1, respectively in absence of thiosulfate), the proportion between these two molar yields remained however constant for all evaluated conditions (about 1 mol of lactate produced per 2 mol of acetate produced) (Table 3).
The studies addressing T. maritima metabolism have showed that this species harvested energy by glycolysis via the Embden–Meyerhof pathway (EMP) as the main route [39, 41]. For the ultimate steps of pyruvate reductions, acetate, H2, and CO2 (1:2:1 as molar proportions) represented the major end-products of glucose fermentation, whereas lactate, alanine, and EPS were minor. In agreement with this classical fermentation model, the H2 yield is only optimized when all glucose is converted to acetate. The highest H2 molar yield, that can be therefore achieved by this fermentation model, was 4 mol of H2 per mole of glucose (or other hexose) referred to as the Thauer limit [38, 39, 68, 69]. Under our experimental conditions, the results showed that all molar yields, including H2/glu, acetate/glu, and lactate/glu, declined sharply in thiosulfate-limited T. maritima growth (range 0–0.06 mmol L−1 of thiosulfate) (Table 3). In contrast, when thiosulfate was in excess (concentrations higher than 0.12 mmol L−1), the molar yields on glucose for acetate, lactate, and H2 reached 1–1.1, 0.4–0.5, and 2–2.2 mol mol−1 as upper limit, respectively (Table 3). Similarly, the kinetic patterns for the hydrogen specific productivity (q H2) and glucose specific consumption (q glucose) increased when the initial thiosulfate concentration increased (Table 3). The changes in the values of these two parameters indicated that the presence of thiosulfate accelerated the glucose consumption and, consequently, the hydrogen production to 18.8 and 40.4 mmol g−1 h−1 as upper limits, respectively (Table 3). In addition, consequently to the increases, all together of cellular concentration, cellular yield (cells/glu), and specific rates (q H2 and q glucose) (Table 3), the effect of thiosulfate was even greater on the volumetric hydrogen and cellular production, and volumetric glucose consumption (Qcells, QH2, and Qglu, respectively) (Fig. 4). As shown in the Fig. 4, Qcells and QH2 were increased sixfold and Qglu fourfold when thiosulfate concentrations were not limiting T. maritima growth (higher than 0.12 mmol L−1) in comparison to culture performed in absence of thiosulfate (Fig. 4). The upper limits for Qcells, QH2, and Qglu reached, in these cases, 25 mg L−1 h−1, 5.6, and 2.8 mmol L−1 h−1, respectively (Fig. 4).

Glucose consumption and hydrogen and cells productivities versus thiosulfate concentrations. Hydrogen productivities (QH2) and glucose consumptions (Qglu) were expressed in mmol per liter of medium and per hour. T. maritima cells productivities (Qcells) were expressed in mg per liter of medium and per hour. These fermentation parameters were calculated during the growth phase from fermentation runs performed in bioreactor (in triplicate). For all fermentations, culture medium contained glucose (60 mmol L−1) and yeast extract (1 g L−1)
Compared to the theoretical Thauer limit (4 mol mol−1), the relative weakness of the best H2 molar yields (H2/glu) (2–2.2 mol mol−1) (Table 3) obtained in this work is naturally explained by the nutrient constraints (excess of glucose with regard to the yeast extract), which had to be imposed in order to achieve the objectives of our study (effects of the concentration of thiosulfate on T. maritima growth). Nevertheless, these yields (2–2.2 mol mol−1) were coherent with the range of H2 molar yields (1.7–4.0 mol mol−1) reported in literature for T. maritima and T. neapolitana [16, 29, 39, 44, 53, 55, 68, 70–72]. It is the same for the highest QH2 (5.6 mmol L−1 h−1) (Fig. 3) which was also ranked in the middle of the range of QH2 values (1 to 14 mmol L−1 h−1) reported in literature [16, 29, 68, 70–72]. Thus, the comparison between the data in this study, for H2/glu, QH2, as well as cellular yields on glucose (cells/glu), with those found in the literature (8.5–9.6 g mol−1 of glucose, as in Table 3, versus 45 g mol−1 as reported by Schroder et al. [39]) suggested that, except for thiosulfate and glucose, the other compounds of our culture medium have to be optimized to reach the theoretical Thauer limit (4 mol of H2 per mole of glucose) and the maximum H2 productivity.
Conclusions
This study has highlighted the necessary requirement, in a culture medium, of sulfur sources, including sulfide, cysteine, or thiosulfate, to grow T. maritima. The focus on thiosulfate, used as model, demonstrated that, in extremely controlled experimental conditions and with glucose as energy source, T. maritima growth was drastically limited by thiosulfate in the range of 0–0.06 mmol L−1. Under such experimental conditions, solely limited by thiosulfate, the cellular yield (Y X/Thio) was accurately determined (3617 ± 176 mg of cells per mmol of thiosulfate consumed). This evaluation was necessary to build a mathematical Monod-based model using glucose, yeast extract, and thiosulfate concentrations and the partial pressure of hydrogen as variables. This model, which simulates T. maritima growth and hydrogen production from glucose fermentation, will be published in the Part II of this publication [62].
Moreover, the results of this study showed that, under yeast extract-limited culture conditions, among all the nutrients present in the yeast extract, sulfur compounds, including both cystine (cysteine dimer) and methionine, were the ones limiting T. maritima growth.
So far, thiosulfate, as well as sulfur, was considered only as a detoxifying agent preventing the accumulation of H2 by oxidation into sulfide. This oxidation therefore relieves the inhibition by H2 of T. maritima and most Thermotoga species growth. This study underlined the increase of T. maritima growth with very low thiosulfate concentrations, for which the detoxifying effect is negligible. Instead, when its concentration was low, thiosulfate was found to be a sulfured nutriment required for the growth, forcing to reconsider its role in this species and most probably also in all thiosulfato-reducer hyperthermophiles. From now on, thiosulfate should be considered in T. maritima (1) as a sulfur source used for the synthesis of cellular materials (anabolism including proteins and Fe–S clusters dedicated to hydrogenase and ferredoxins, for instance) when thiosulfate is present at low concentrations (about 0.06 mmol L−1 under our experimental conditions), and (2) as both sulfur source and detoxifying agent at higher concentrations. Concerning this latter case, the intensity of the hydrogen detoxification function will depend on the thiosulfate availability and the level of the hydrogen partial pressure within culture medium as discussed in the part II of this manuscript at the end of “Results and discussion” section [62].
Otherwise, based on the comparison of the patterns for the end-products of glucose fermentation, obtained from T. maritima grown under different stress conditions such as nutritional deficiencies by nitrogen or thiosulfate, or oxidative stress by the presence of oxygen, amazing analogies were highlighted and discussed.
Finally, as demonstrated in this study, the thiosulfate addition, in the culture medium formulation, in a range of 0.12–0.24 mmol L−1 was able to increase significantly the cellular and hydrogen productivities in T. maritima by a factor of 6 compared to a thiosulfate-free medium. Based on these results, it can be recommended, for all works dealing with the optimization of hydrogen production from hexoses by dark fermentation using T. maritima or T. neapolitana, to add thiosulfate in the medium in the proportion of about 0.3 mmol of thiosulfate per g of cells (calculated from Y X/thio = 3617 mg of cells (cell dry weight) per mmol of thiosulfate). Thus, it will stimulate the cellular growth and hydrogen production while limiting the sulfide production within biogas, which is a damaging product for biotechnological applications in the energy field.
Abbreviations
- EPS:
-
extracellular polysaccharides
- EMP:
-
Embden–Meyerhof pathway
- ED:
-
Entner–Doudoroff pathway
- pH2:
-
partial pressure of dihydrogen expressed in millibars
- Eh:
-
measurement, expressed in millivolts, of the reduction potential relative to a standard hydrogen electrode
- DSMZ:
-
Deutsche Sammlung von Mikroorganismen und Zellkulturen
- DMSO:
-
dimethyl sulfoxide
- Na2S:
-
sodium sulfide
- micro-GC:
-
micro gas chromatograph
- GC-FPD:
-
gas chromatograph flame photometric detector
- OD:
-
optical density
- CDW:
-
cell dry weight
- HPLC:
-
high Pressure liquid chromatograph
- Qcells:
-
cellular production rate [mg (cdw) L−1 h−1)]
- Qglu:
-
glucose consumption rate (mmol L−1 h−1)
- QH2 :
-
hydrogen production rate (mmol L−1 h−1)
References
Rao KK, Cammack R. Producing hydrogen as a fuel. In: Cammack R, Frey M, Robson R, editors. Hydrogen as a fuel: learning from nature. New York: CRC Press; 2001. p. 201–29.
Singh A, Sevda S, Abu Reesh I, Vanbroekhoven K, Rathore D, Pant D. Biohydrogen production from lignocellulosic biomass: Technology and Sustainability. Energies. 2015;8(11):12357.
Ren N, Guo W, Liu B, Cao G, Ding J. Biological hydrogen production by dark fermentation: challenges and prospects towards scaled-up production. Curr Opin Biotechnol. 2011;22(3):365–70.
Hallenbeck PC, Ghosh D. Advances in fermentative biohydrogen production: the way forward? Trends Biotechnol. 2009;27(5):287–97.
Pant D, Van Bogaert G, Diels L, Vanbroekhoven K. A review of the substrates used in microbial fuel cells (MFCs) for sustainable energy production. Bioresour Technol. 2010;101(6):1533–43.
Turner J, Sverdrup G, Mann MK, Maness PC, Kroposki B, Ghirardi M, et al. Renewable hydrogen production. Int J Energy Res. 2008;32(5):379–407.
Redwood MD, Paterson-Beedle M, Macaskie LE. Integrating dark and light bio-hydrogen production strategies: towards the hydrogen economy. Rev Environ Sci Bio/Technol. 2009;8(2):149–85.
Rai PK, Singh SP, Asthana RK, Singh S. Biohydrogen production from sugarcane bagasse by integrating dark-and photo-fermentation. Bioresour Technol. 2014;152:140–6.
Guwy AJ, Dinsdale RM, Kim JR, Massanet-Nicolau J, Premier G. Fermentative biohydrogen production systems integration. Bioresour Technol. 2011;102(18):8534–42.
Chookaew T, Prasertsan P, Ren ZJ. Two-stage conversion of crude glycerol to energy using dark fermentation linked with microbial fuel cell or microbial electrolysis cell. New Biotechnol. 2014;31(2):179–84.
Basak N, Das D. The prospect of purple non-sulfur (PNS) photosynthetic bacteria for hydrogen production: the present state of the art. World J Microbiol Biotechnol. 2007;23(1):31–42.
Urbaniec K, Bakker RR. Biomass residues as raw material for dark hydrogen fermentation—a review. Int J Hydrog Energy. 2015;40(9):3648–58.
Sarma SJ, Pachapur V, Brar SK, Le Bihan Y, Buelna G. Hydrogen biorefinery: potential utilization of the liquid waste from fermentative hydrogen production. Renew Sustain Energy Rev. 2015;50:942–51.
Ghimire A, Frunzo L, Pirozzi F, Trably E, Escudie R, Lens PNL, et al. A review on dark fermentative biohydrogen production from organic biomass: process parameters and use of by-products. Appl Energy. 2015;144:73–95.
Das D, Veziroglu TN. Advances in biological hydrogen production processes. Int J Hydrog Energy. 2008;33(21):6046–57.
D’Ippolito G, Dipasquale L, Vella FM, Romano I, Gambacorta A, Cutignano A, et al. Hydrogen metabolism in the extreme thermophile Thermotoga neapolitana. Int J Hydrog Energy. 2010;35(6):2290–5.
Kumar N, Das D. Enhancement of hydrogen production by Enterobacter cloacae IIT-BT 08. Process Biochem. 2000;35(6):589–93.
Chen CC, Lin CY, Chang JS. Kinetics of hydrogen production with continuous anaerobic cultures utilizing sucrose as the limiting substrate. Appl Microbiol Biotechnol. 2001;57(1–2):56–64.
Ren N-Q, Cao GL, Guo WQ, Wang AJ, Zhu YH, B-f Liu, et al. Biological hydrogen production from corn stover by moderately thermophile Thermoanaerobacterium thermosaccharolyticum W16. Int J Hydrog Energy. 2010;35(7):2708–12.
Ren N, Wang A, Cao G, Xu J, Gao L. Bioconversion of lignocellulosic biomass to hydrogen: potential and challenges. Biotechnol Adv. 2009;27(6):1051–60.
Kapdan IK, Kargi F. Bio-hydrogen production from waste materials. Enzyme Microb Technol. 2006;38(5):569–82.
Show KY, Lee DJ, Tay JH, Lin CY, Chang JS. Biohydrogen production: current perspectives and the way forward. Int J Hydrog Energy. 2012;37(20):15616–31.
Chen CC, Chuang YS, Lin CY, Lay CH, Sen B. Thermophilic dark fermentation of untreated rice straw using mixed cultures for hydrogen production. Int J Hydrog Energy. 2012;37(20):15540–6.
Valdez-Vazquez I, Rios-Leal E, Esparza-Garcia F, Cecchi F. Poggi-Varaldo HcM. Semi-continuous solid substrate anaerobic reactors for H2 production from organic waste: mesophilic versus thermophilic regime. Int J Hydrog Energy. 2005;30(13):1383–91.
Tawfik A, El-Qelish M. Continuous hydrogen production from co-digestion of municipal food waste and kitchen wastewater in mesophilic anaerobic baffled reactor. Bioresour Technol. 2012;114:270–4.
Varrone C, Rosa S, Fiocchetti F, Giussani B, Izzo G, Massini G, et al. Enrichment of activated sludge for enhanced hydrogen production from crude glycerol. Int J Hydrog Energy. 2013;38(3):1319–31.
Sahlström L. A review of survival of pathogenic bacteria in organic waste used in biogas plants. Bioresour Technol. 2003;87(2):161–6.
Van Groenestijn JW, Hazewinkel JHO, Nienoord M, Bussmann PJT. Energy aspects of biological hydrogen production in high rate bioreactors operated in the thermophilic temperature range. Int J Hydrog Energy. 2002;27(11):1141–7.
Nguyen TAD, Kim JP, Kim MS, Oh YK, Sim SJ. Optimization of hydrogen production by hyperthermophilic eubacteria, Thermotoga maritima and Thermotoga neapolitana in batch fermentation. Int J Hydrog Energy. 2008;33(5):1483–8.
Kadar Z, de Vrije T, van Noorden GE, Budde MAW, Szengyel Z, Réczey K, et al. Yields from glucose, xylose, and paper sludge hydrolysate during hydrogen production by the extreme thermophile Caldicellulosiruptor saccharolyticus. Appl Biochem Biotechnol. 2004;114(1–3):497–508.
Darimont B, Sterner R. Sequence, assembly and evolution of a primordial ferredoxin from Thermotoga maritima. EMBO J. 1994;13(8):1772–81.
Huber R, Langworthy TA, König H, Thomm M, Woese CR, Sleytr UB, et al. Thermotoga maritima sp. nov. represents a new genus of unique extremely thermophilic eubacteria growing up to 90 °C. Arch Microbiol. 1986;144(4):324–33.
Woese CR. Bacterial evolution. Microb Mol Biol R. 1987;51(2):221–71.
Nelson KE, Clayton RA, Gill SR, Gwinn ML, Dodson RJ, Haft DH, et al. Evidence for lateral gene transfer between Archaea and Bacteria from genome sequence of Thermotoga maritima. Nature. 1999;399(6734):323–9.
Huber R, Stetter KO. The order Thermotogales. In: Balows A, Tripper HG, Dworkin M, Harder W, Schleifer KH, editors. The Prokaryotes. New York: Springer; 1992. p. 3809–15.
Jeanthon C, Reysenbach A-L, L’Haridon S, Gambacorta A, Pace N, Glénat P, et al. Thermotoga subterranea sp. nov., a new thermophilic bacterium isolated from a continental oil reservoir. Arch Microbiol. 1995;164(2):91–7.
Fardeau ML, Ollivier B, Patel BKC, Magot M, Thomas P, Rimbault A, et al. Thermotoga hypogea sp. nov., a xylanolytic, thermophilic bacterium from an oil-producing well. Int J Syst Bacteriol. 1997;47(4):1013–9.
Chou C-J, Jenney FE, Adams MWW, Kelly RM. Hydrogenesis in hyperthermophilic microorganisms: implications for biofuels. Metab Eng. 2008;10(6):394–404.
Schröder C, Selig M, Schönheit P. Glucose fermentation of acetate, CO2 and H2 in the anaerobic hyperthermophilic eubacterium Thermotoga maritima: involvement of the Embden-Meyerhof pathway. Arch Microbiol. 1994;161(6):460–70.
Chhabra SR, Shockley KR, Conners SB, Scott KL, Wolfinger RD, Kelly RM. Carbohydrate-induced differential gene expression patterns in the Hyperthermophilic Bacterium Thermotoga maritima. J Biol Chem. 2003;278(9):7540–52.
Selig M, Xavier KB, Santos H, Schönheit P. Comparative analysis of Embden-Meyerhof and Entner–Doudoroff glycolytic pathways in hyperthermophilic archaea and the bacterium Thermotoga. Arch Microbiol. 1997;167(4):217–32.
Janssen PH, Morgan HW. Heterotrophic sulfur reduction by Thermotoga sp. strain FjSS3.B1. FEMS Microbiol Lett. 1992;96(2–3):213–7.
Lakhal R, Auria R, Davidson S, Ollivier B, Dolla A, Hamdi M, et al. Effect of oxygen and redox potential on glucose fermentation in Thermotoga maritima under controlled physicochemical conditions. Int J Microbiol. 2010:1–10.
De Vrije T, Budde MAW, Lips SJ, Bakker RR, Mars AE. Claassen PAM. Hydrogen production from carrot pulp by the extreme thermophiles Caldicellulosiruptor saccharolyticus and Thermotoga neapolitana. Int J Hydrog Energy. 2010;35(24):13206–13.
Le Fourn C, Fardeau M-L, Ollivier B, Lojou E, Dolla A. The hyperthermophilic anaerobe Thermotoga maritima is able to cope with limited amount of oxygen: insights into its defence strategies. Environ Microbiol. 2008;10(7):1877–87.
Rinker KD, Kelly RM. Effect of carbon and nitrogen sources on growth dynamics and exopolysaccharide production for the hyperthermophilic archaeon Thermococcus litoralis and bacterium Thermotoga maritima. Biotechnol Bioeng. 2000;69(5):537–47.
Huber R, Hannig M. Thermotogales. In: Balows A, Tripper HG, Dworkin M, Harder W, Schleifer KH, editors. The Prokaryotes. New York: Springer; 2006. p. 899–922.
Ravot G, Ollivier B, Magot M, Patel BKC, Crolet J, Fardeau M, et al. Thiosulfate reduction, an important physiological feature shared by members of the order Thermotogales. Appl Environ Microbiol. 1995;61(5):2053–5.
Ravot G, Ollivier B, Fardeau ML, Patel BK, Andrews KT, Magot M, et al. l-alanine production from glucose fermentation by hyperthermophilic members of the domains Bacteria and Archaea: a remnant of an ancestral metabolism? Appl Environ Microbiol. 1996;62(7):2657–9.
Jannasch HW, Huber R, Belkin S, Stetter KO. Thermotoga neapolitana sp. nov. of the extremely thermophilic, eubacterial genus Thermotoga. Arch Microbiol. 1988;150(1):103–4.
Childers SE, Vargas M, Noll KM. Improved methods for cultivation of the extremely thermophilic bacterium Thermotoga neapolitana. Appl Environ Microbiol. 1992;58(12):3949–53.
Belkin S, Wirsen CO, Jannasch HW. A new sulfur-reducing, extremely thermophilic eubacterium from a submarine thermal vent. Appl Environ Microbiol. 1986;51(6):1180–5.
Nguyen TAD, Han SJ, Kim JP, Kim MS, Sim SJ. Hydrogen production of the hyperthermophilic eubacterium, Thermotoga neapolitana under N2 sparging condition. Bioresour Technol. 2010;101(1):S38–41.
Levin DB, Pitt L, Love M. Biohydrogen production: prospects and limitations to practical application. Int J Hydrog Energy. 2004;29(2):173–85.
Eriksen NT, Riis ML, Holm NK, Iversen N. H2 synthesis from pentoses and biomass in Thermotoga spp. Biotechnol Lett. 2011;33(2):293–300.
Hungate RE. A roll-tube method for the cultivation of strict anaerobes. In: Norris JR, Ribbons DW, editors. Methods in microbiology. London: Academic Press; 1969. p. 117–32.
Macy JM, Snellen JE, Hungate RE. Use of syringe methods for anaerobiosis. Am J Clin Nutr. 1972;25(12):1318–23.
Balch WE, Fox GE, Magrum LJ, Woese CR, Wolfe RS. Methanogens: reevaluation of a unique biological group. Microb Mol Biol R. 1979;43(2):260.
Miller TL, Wolin MJ. A serum bottle modification of the Hungate technique for cultivating obligate anaerobes. Appl Microbiol. 1974;27(5):985–7.
Moore S, Spackman DH, Stein WH. Chromatography of amino acids on sulfonated polystyrene resins: an improved system. Anal Chem. 1958;30(7):1185–90.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350–6.
Auria R, Boileau C, Davidson S, Casalot L, Christen P, Liebgott PP, Combet-Blanc Y. Hydrogen production by the hyperthermophilic bacterium Thermotoga maritima Part II: modeling and experimental approaches for hydrogen production. Biotechnol Biofuels. 2016. doi:10.1186/s13068-016-0681-0.
Angenent LT, Karim K, Al-Dahhan MH, Wrenn BA, Domiguez-Espinosa R. Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol. 2004;22(9):477–85.
van Niel EWJ, Claassen PAM, Stams AJM. Substrate and product inhibition of hydrogen production by the extreme thermophile Caldicellulosiruptor saccharolyticus. Biotechnol Bioeng. 2003;81(3):255–62.
Hawkes FR, Hussy I, Kyazze G, Dinsdale R, Hawkes DL. Continuous dark fermentative hydrogen production by mesophilic microflora: principles and progress. Int J Hydrog Energy. 2007;32(2):172–84.
Lapaglia C, Hartzell PL. Stress-induced production of biofilm in the hyperthermophile Archaeoglobus fulgidus. Appl Environ Microbiol. 1997;63(8):3158–63.
Pysz MA, Conners SB, Montero CI, Shockley KR, Johnson MR, Ward DE, et al. Transcriptional analysis of biofilm formation processes in the anaerobic, hyperthermophilic bacterium Thermotoga maritima. Appl Environ Microbiol. 2004;70(10):6098–112.
Eriksen NT, Nielsen TM, Iversen N. Hydrogen production in anaerobic and microaerobic Thermotoga neapolitana. Biotechnol Lett. 2008;30(1):103–9.
Nandi R, Sengupta S. Microbial production of hydrogen: an overview. Crit Rev Microbiol. 1998;24(1):61–84.
Munro SA, Zinder SH, Walker LP. The fermentation stoichiometry of Thermotoga neapolitana and influence of temperature, oxygen, and pH on hydrogen production. Biotechnol Prog. 2009;25(4):1035–42.
Ngo TA, Kim M-S, Sim SJ. Thermophilic hydrogen fermentation using Thermotoga neapolitana DSM 4359 by fed-batch culture. Int J Hydrog Energy. 2011;36(21):14014–23.
De Vrije T, Bakker RR, Budde MAW, Lai MH, Mars AE, Claassen PAM. Efficient hydrogen production from the lignocellulosic energy crop Miscanthus by the extreme thermophilic bacteria Caldicellulosiruptor saccharolyticus and Thermotoga neapolitana. Biotechnol Biofuels. 2009;2(1):1.
Authors’ contributions
CB performed most of the experiments, analyzed the data, and revised the manuscript. RA analyzed the data and revised the manuscript. SD supervised experimental part of the project, and participated in results analysis. LC participated in results analysis, drafted, and revised the manuscript. PPL was involved in the design and carrying out part of the experimental study, and revised the manuscript. PC participated in the design of experiments and helped the interpretation of results. YCB acted as coordinator of the project, designed this study, analyzed experimental results, drafted, and revised the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The authors thank the Decobecq team (Decobecq Automatismes, France) for technical support on the BatchPro software used for bioreactor monitoring and management. This work was supported by the French Agence Nationale pour la Recherche (National Research Agency) under the HYCOFOL_BV project. The laboratories, where all the experiments were performed, are in compliance with ISO 9001-2015.
Competing interests
The authors declare that they have no competing interests.
Availability of supporting data
All data generated or analyzed during this study are included in this article.
Consent for publication
All authors approved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Boileau, C., Auria, R., Davidson, S. et al. Hydrogen production by the hyperthermophilic bacterium Thermotoga maritima part I: effects of sulfured nutriments, with thiosulfate as model, on hydrogen production and growth. Biotechnol Biofuels 9, 269 (2016). https://doi.org/10.1186/s13068-016-0678-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-016-0678-8