
Abstract
Background
The production of hydrogen from biomass by fermentation is one of the routes that can contribute to a future sustainable hydrogen economy. Lignocellulosic biomass is an attractive feedstock because of its abundance, low production costs and high polysaccharide content.
Results
Batch cultures of Caldicellulosiruptor saccharolyticus and Thermotoga neapolitana produced hydrogen, carbon dioxide and acetic acid as the main products from soluble saccharides in Miscanthus hydrolysate. The presence of fermentation inhibitors, such as furfural and 5-hydroxylmethyl furfural, in this lignocellulosic hydrolysate was avoided by the mild alkaline-pretreatment conditions at a low temperature of 75°C. Both microorganisms simultaneously and completely utilized all pentoses, hexoses and oligomeric saccharides up to a total concentration of 17 g l-1 in pH-controlled batch cultures. T. neapolitana showed a preference for glucose over xylose, which are the main sugars in the hydrolysate. Hydrogen yields of 2.9 to 3.4 mol H2 per mol of hexose, corresponding to 74 to 85% of the theoretical yield, were obtained in these batch fermentations. The yields were higher with cultures of C. saccharolyticus compared to T. neapolitana. In contrast, the rate of substrate consumption and hydrogen production was higher with T. neapolitana. At substrate concentrations exceeding 30 g l-1, sugar consumption was incomplete, and lower hydrogen yields of 2.0 to 2.4 mol per mol of consumed hexose were obtained.
Conclusion
Efficient hydrogen production in combination with simultaneous and complete utilization of all saccharides has been obtained during the growth of thermophilic bacteria on hydrolysate of the lignocellulosic feedstock Miscanthus. The use of thermophilic bacteria will therefore significantly contribute to the energy efficiency of a bioprocess for hydrogen production from biomass.
Similar content being viewed by others


Background
In view of the transition to hydrogen as a major energy carrier in the future, new routes for hydrogen production need to be explored. The production of hydrogen from biomass is one of the options for contributing to the supply of exploitable renewable resources. Hydrogen can be produced from a vast range of biomass, using thermochemical, as well as fermentative, processes. Carbohydrates, such as sugars, starch or (hemi)cellulose, are the prime substrates for fermentative processes. For future sustainability of the energy supply, the utilization of (hemi)cellulose is of prime interest, as this component is most abundant in crops that can be grown for the purpose of energy supply.
To date, many studies have been done on fermentative hydrogen production from pure sugars and from feedstocks, such as by-products from the agricultural and food industry, municipal waste, or wastewaters [1]. However, only a few studies describe the production of hydrogen from lignocellulosic biomass (reviewed in [2]). Some of these feedstocks were offered as solid materials, such as office paper waste [3], wheat straw waste [4], delignified wood fibres [5], and a variety of cellulosic waste materials [6]. Other types of biomass, such as Miscanthus [7], paper sludge [8], corn stover [9], and corn stalks [10] were first pretreated and/or hydrolyzed to obtain a soluble substrate of mixed sugars and oligosaccharides. Bacterial consortia from anaerobic digester sludge or cow dung compost, as well as single cultures of mesophilic (Clostridium acetobutylicum) and thermophilic bacteria (Clostridium thermocellum, Caldicellulosiruptor saccharolyticus and Thermotoga elfii), have been used as hydrogen producers. The reported hydrogen yields on these lignocellulosic substrates varied greatly from approximately 10% to more than 80% of the theoretical value, which is 4 mol of hydrogen per mol of hexose. The diversity of the applied feedstocks and pretreatment methods hardly allow a comparison of hydrogen production efficiency.
It is proposed that research should focus on the conversion of biomass to fermentable substrates to obtain a successful introduction of cellulosic biomass for the production of biofuels [11]. The fermentability of lignocellulose is improved by pretreatment of the biomass. This is required to overcome the recalcitrance of the lignocellulosic complex by altering its structure, which makes the cellulose and hemicelluloses accessible to the enzymes [12]. One of the methods of interest is pretreatment with an alkaline agent at relatively low temperatures (<100°C). One of the main effects of the alkali is the disruption of the intermolecular bonding between xylan and other biomass components, such as lignin or other hemicellulosic components, resulting in increased porosity of the lignocellulosic biomass. In addition, applying alkali in a water solution generally leads to swelling of cell wall material, thereby increasing the internal surface of the lignocellulosic matrix. These effects (that is, increase of porosity; swelling of fibrous material) lead to an increase in enzymatic degradability, as the lignocellulosic matrix is more accessible for enzymes. Another effect, less well understood, is the modification of lignin during alkaline pretreatment. Depending on the alkali used and the biomass type, alkaline pretreatment may lead to lignin depolymerisation, and (partial) dissolution of lignin components.
This study describes the efficiency of hydrogen production from Miscanthus, a lignocellulosic energy crop. Miscanthus is a rapidly growing perennial C4 grass with relatively high yields of 8 to 15 ton dry weight per ha in Western European regions, which requires only low inputs of nutrients for cultivation. It has been studied for years because of its potential for future energy supply. Previously, we have compared mechanical methods of pretreatment, milling and extrusion, in combination with chemical treatment. The applied methods were aimed at the high conversion of polysaccharides and high yields of monomeric sugars [7]. In the present study, the effect of various chemicals used for alkali pretreatment on the fermentability of the feedstock is investigated. For this, two extreme thermophilic bacteria were used, the cellulolytic bacterium Caldicellulosiruptor saccharolyticus of the order Clostridiales [13], and the moderately halophilic bacterium Thermotoga neapolitana of the order Thermotogales [14], which grow at optimum temperatures of 70 and 80°C, respectively. Thermophilic bacteria are superior with respect to hydrogen yield [15–17], due to favourable thermodynamical conditions at high temperatures, and reduced variety in by-product formation. Furthermore, many thermophilic bacteria, including T. neapolitana and C. saccharolyticus, are able to utilize a wide range of substrates for growth from simple sugars to complex carbohydrates [18, 19]. Unlike T. neapolitana, C. saccharolyticus is capable of growth on crystalline cellulose [13], and on lignocellulosic feedstocks, although degradation of these substrates was limited [20]. Therefore, the pretreatment was followed by enzymatic hydrolysis of Miscanthus to prepare fermentable substrates. The soluble fraction, that is, the hydrolysate, will be a complex mixture of monomeric C6 and C5 sugars, and di- and oligosaccharides. The production of hydrogen from all of these sugars is required to develop an efficient process. The results showed that both thermophiles were able to consume most, if not all, soluble carbohydrates present in the Miscanthus hydrolysate. Hydrogen was produced at high yields of more than 75% of the theoretical value.
Results and discussion
Comparison of alkaline pretreatment methods
Two types of alkali, NaOH and Ca(OH)2, were tested for the pretreatment of Miscanthus. The dosage of these types of alkali was based on earlier reported work [7, 21]. Indicators of the efficiency of lignocellulosic biomass pretreatment are the pH after the addition of the alkali at the start of the pretreatment at 85°C, the amount of delignification after pretreatment, and the enzymatic degradability of the pretreated biomass. The results showed that NaOH was most effective because of the higher initial pH of the biomass, and the lower lignin content of the insoluble fraction of the material that was pretreated with NaOH (Table 1). Still, a reasonable amount of monomeric sugars can be released from Ca(OH)2-pretreated Miscanthus, despite the insignificant delignification of the biomass during pretreatment (Table 2).
The selection of an optimal pretreatment method will not only be determined by the efficiencies of the biomass pretreatment, and the enzymatic hydrolysis of the polysaccharides, but also by the fermentability of the hydrolysates. Hydrolysates were prepared from batches of approximately 200 g of milled Miscanthus, which were pretreated using NaOH or Ca(OH)2, titrated with different acids prior to enzymatic hydrolysis, and hydrolyzed with commercial enzyme preparations, after which the sugar composition of the hydrolysates was determined (Table 2). In addition, the acid usage in the preparation of hydrolysate IV was reduced by circa 50% through washing with water prior to pH adjustment. Glucose and xylose were the main monosaccharides in the hydrolysates. The highest amount of monomeric sugars was found in hydrolysate III, which has been prepared by pretreatment with the highest amount of alkali. This resulted in a polysaccharide conversion efficiency of circa 55%.
Fermentability of hydrolysates on small scale and inhibitory compounds
The fermentability of the four hydrolysates was tested in small-scale experiments using closed flasks. For this, the production of hydrogen and the main organic acids (acetate and lactate) by C. saccharolyticus and T. neapolitana on a concentration range of diluted hydrolysate was monitored and compared to production in control fermentations using pure sugars. The final sugar concentration in all samples was adjusted to 10 g l-1 by adding pure sugars (glucose and xylose, 7:3, w w-1). Both bacteria were able to grow and produce hydrogen with all hydrolysates (Table 3). C. saccharolyticus performed well on hydrolysates II and IV. The fermentability of hydrolysate I, and to a lesser extent hydrolysate III, was poor as production of organic acids was already inhibited at low sugar concentrations. T. neapolitana also showed good growth and organic acid production during growth on hydrolysate IV, and fermentation was slightly inhibited with hydrolysate I. In contrast to C. saccharolyticus, organic acid production was inhibited on hydrolysate II.
The inhibition of fermentation might be caused by the chemicals used for pretreatment and pH adjustment. Growth of C. saccharolyticus is inhibited by high ionic strength of the culture medium [22]. Indeed, 50% inhibition of growth and the production of hydrogen and organic acids were observed at approximately 130 mM acetate (Table 4). This effect was not observed with T. neapolitana, which is in line with its moderately halophilic nature. The high concentration of acetate in hydrolysate III probably caused the reduced organic acid production with C. saccharolyticus. The washing of the alkaline-pretreated material prior to pH adjustment reduced the use of acetic acid (Table 2) and prevented the inhibition of fermentation (Table 3).
Salts, such as calcium chloride, potassium phosphate, and sodium sulfate, also inhibit C. saccharolyticus (Table 4). The presence of calcium and sulfate ions in hydrolysate I (Table 2) probably contributed to the strong inhibition with this hydrolysate. The inhibition of C. saccharolyticus was not observed with hydrolysate II, because the concentration of calcium ions was reduced through formation of an insoluble salt when phosphoric acid, instead of sulfuric acid, was used for pH adjustment. The growth of T. neapolitana appeared to be stimulated by the presence of calcium ions up to 50 mmol l-1, and the organism is less sensitive for phosphate and sulfate (Table 4). The inhibition of fermentation by hydrolysate I and II is, therefore, likely to be caused by other components in these hydrolysates.
Inhibitory compounds may be generated during the pretreatment and hydrolysis of lignocellulosic biomass. These include degradation products of sugars, such as the aldehydes furfural and 5-hydroxymethyl furfural (HMF). The effects of 0 to 4 g l-1 of furfural and HMF on the fermentability of glucose were tested in small flasks. Both C. saccharolyticus and T. neapolitana were inhibited by these compounds. C. saccharolyticus appeared to be more sensitive (Table 4). Actively growing cells of C. saccharolyticus fully metabolized furfural to mainly furfuryl alcohol (>80%). However, at 4 g l-1, hardly any growth was observed and the furfural was no longer converted. Similar results were obtained with HMF, although the conversion products were not identified. T. neapolitana also metabolized furfural and HMF. Furfural was partly converted to furfuryl alcohol, while other conversion products were not identified. Previous reports also showed inhibition of growth of other bacteria by similar concentrations of furfural and HMF (IC50 at 1 to 4 g l-1; [23, 24]), but a stimulatory effect (up to 3 g l-1) on the growth of Clostridium beijerinckii has also been reported [25]. The reduction of the furaldehydes to alcohols has been mentioned earlier as a detoxification step under both aerobic and anaerobic conditions [23]. In yeast, alcohol dehydrogenases and xylose reductase were responsible for the reduction of furfural and HMF [26, 27]. Bacterial enzymes involved in furaldehyde reduction have not been identified yet, but the reduction appeared to be constitutive with reduced nicotinamide adenine dinucleotide (phosphate) (NAD(P)H) acting as the electron donor [28]. The concentration of furaldehydes in hydrolysate I and II was less than 0.1 g l-1 and, therefore, the inhibition of T. neapolitana was not caused by these compounds.
Hydrolysate preparation on bench scale and fermentability
The best fermentability was observed with hydrolysate IV. This batch was prepared from NaOH-pretreated Miscanthus that was washed with water to remove a large part of the alkali (and solubilised lignin), thus reducing the amount of acetic acid needed for pH adjustment. In order to validate this method and the process conditions at the bench scale, 1.35 kg of Miscanthus was cut and sieved to pieces of circa 1 cm, and subsequently treated with 9% NaOH (w w-1) for 6 h at 75°C in a 10 l stirred tank reactor. After dewatering, the pH of the slurry was adjusted with acetic acid, and the pulp was enzymatically hydrolyzed by using a fed-batch procedure. The viscosity of the slurry with 27.5% dry matter was rapidly reduced during the hydrolysis process (Figure 1). The clarified hydrolysate contained 38.3 g l-1 of monosaccharides (Table 5). The conversion efficiency was comparable to the small-scale experiment (hydrolysate IV), despite the shorter period of alkali treatment (6 h instead of 16 h), and lower pretreatment temperature (75°C instead of 85°C). The total amount of monosaccharides in the hydrolysate increased by 19% after sulfuric acid treatment, indicating that other soluble sugars were present as di- and/or oligosaccharides (Table 5). More than half of the extra monomers were xylose. The incomplete degradation of the polysaccharides to monosaccharides is partly due to product inhibition of the hydrolytic enzyme activity. Xylan degradation to xylose appeared to be extra inefficient, most likely because the enzyme preparation was not optimally balanced for the complete degradation of xylans to xylose [29]. The contribution of carbohydrates to the total chemical oxygen demand (COD) of the hydrolysate was 73% (Table 5).

Enzymatic hydrolysis of alkaline-pretreated Miscanthus on bench scale. (a) The slurry immediately after addition of the enzyme preparation (GC 220 at a final loading rate of 15 IFPU per g dry matter). (b) After 24 h of hydrolysis. Miscanthus was pretreated with 9% NaOH (w w-1) for 6 h at 75°C.
During enzymatic hydrolysis, the pH of the slurry decreased from the initial pH of 5.2 to 4.3. Besides the release of organic acids from the lignocellulosic material during hydrolysis, this decline can be ascribed to lactic acid formation by contaminating bacteria. The relative amount of other, unknown, organic compounds amounted to 21% of the total COD of the hydrolysate. Because of the mild process conditions (moderate temperature, ambient pressure, and no extreme acidic pH), sugar degradation products are not expected to be formed. Indeed, furfural and HMF were not detected in the hydrolysate (detection limit 10 mg l-1).
The fermentability of this hydrolysate was tested in small flasks with C. saccharolyticus and T. neapolitana. The production of hydrogen and organic acids (acetate and lactate) was observed with hydrolysate concentrations up to 28 g l-1 of monosaccharides (Figure 2). At 10 g l-1, hydrogen production from the hydrolysate by both thermophiles was less when compared to production from the corresponding pure sugars (glucose and xylose at 7 and 3 g l-1, respectively), and decreased further at higher hydrolysate concentrations. The production of organic acids by both bacteria was very similar for all hydrolysate concentrations, and comparable to that from pure sugars. Only the rate of organic acid production by C. saccharolyticus appeared to be lower at the highest hydrolysate concentration. The difference between hydrogen and organic acid production was due to increased lactate production at higher hydrolysate concentrations, which is not accompanied by hydrogen production. The production of lactate might be prevented in favour of acetate and hydrogen formation during fermentations under pH and hydrogen pressure (p H2) controlled conditions.

Hydrogen and organic acid production by cultures of C. saccharolyticus and T. neapolitana grown on Miscanthus hydrolysate. (a) and (c) C. saccharolyticus. (b) and (d) T. neapolitana. (a) and (b) Hydrogen production. (c) and (d) Organic acid (acetic and lactic acid) production. Measurements were done after 16 and 40 h after the start of the fermentation. Hydrolysate concentration is given in g monosaccharides l-1. In the control without hydrolysate, 10 g l-1 of a glucose/xylose (7:3, w w-1) mixture was used as the carbon source. The error bars show the range about the mean of the values from two flasks.
Anaerobic batch fermentations under controlled conditions in a bioreactor
Fermentations under controlled conditions were done with Miscanthus hydrolysate and the glucose-xylose mixture using a continuously-stirred bioreactor (CSTR). The p H2 was kept below the critical concentration of circa 10 kPa [22] by flushing the culture medium with N2. Details of the fermentations are shown in Table 6. At concentrations of 10 and 14 g l-1, the consumption of pure sugars was completed within 22 to 34 h. At the end of the T. neapolitana fermentation some fructose was observed (<0.3 g l-1). Fructose might be the product of the activity of a xylose isomerase identified in this bacterium, which is able to catalyze the isomerisation of glucose to fructose [30]. Although T. neapolitana is able to grow on fructose, consumption is slow and usually incomplete (unpublished results). At a higher concentration of 28 g l-1 of pure sugars, consumption by C. saccharolyticus was still incomplete after 71 h. The fermentation by T. neapolitana was stopped after 45 h because of brown-colouring of the culture medium due to the occurrence of Maillard reactions at the high fermentation temperature of 80°C. At that moment, hydrogen production was low (approximately 0.3% in the off gas).
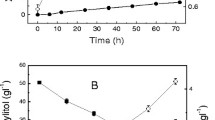
With low hydrolysate concentrations, the duration of the fermentation was prolonged by a few hours as compared to fermentations on pure sugars, except for the 14 g l-1 fermentation by C. saccharolyticus, where the sugar consumption was significantly retarded. At the highest hydrolysate concentration, substrate consumption by C. saccharolyticus was incomplete and T. neapolitana cultures were severely coloured brown. Both thermophiles utilized other organic compounds in the hydrolysate besides glucose and xylose. Based on COD measurements, the extra consumption was circa 20%. Among the organic compounds were arabinose (approx. 3%), and di- and/or oligosaccharides. The latter were consumed within the first 25 h of the fermentations at a hydrolysate concentration of 14 g l-1 of monosaccharides (Figure 3). The consumption of di- and/or oligosaccharides also occurred at the highest substrate concentration, but was initially less rapid.

Consumption of carbohydrates by cultures of C. saccharolyticus and T. neapolitana grown on Miscanthus hydrolysate. Monosaccharide consumption (filled symbols) and consumption of di- and/or oligosaccharides (open symbols) by C. saccharolyticus (a) and T. neapolitana (b) in a medium containing Miscanthus hydrolysate. The hydrolysate concentration was at 14 (triangles) and 28 (diamonds) g monosaccharides l-1.
Both thermophiles simultaneously consumed glucose and xylose, but T. neapolitana preferred glucose, while C. saccharolyticus consumed both sugars at a very similar rate (Figure 4). It has been shown previously that C. saccharolyticus has a preference for xylose if both sugars are present at equal concentration [31]. At higher glucose/xylose ratios, such as in the Miscanthus hydrolysate, a preference for xylose was no longer observed.

Simultaneous consumption of monosaccharides by cultures of C. saccharolyticus and T. neapolitana grown on Miscanthus hydrolysate. Consumption of glucose (circle) and xylose (triangle) by C. saccharolyticus (filled symbols) and T. neapolitana (open symbols) in a medium containing Miscanthus hydrolysate. The monosaccharide concentration was 14 g l-1.
The main products of the fermentations were hydrogen, acetate and CO2 (Table 6). The highest amount of hydrogen, 319 mmol l-1 or 7.1 l l-1, was produced by C. saccharolyticus in a fermentation of 14 g l-1 monosaccharides from the Miscanthus hydrolysate. In general, hydrogen and acetate production by C. saccharolyticus were higher than by T. neapolitana. For the fermentations at lower sugar concentrations, this was partly due to the slightly higher lactate production by T. neapolitana as compared to C. saccharolyticus. Lactate production by C. saccharolyticus was very low in fermentations on pure sugars, as well as on the hydrolysate. T. neapolitana produced a little more lactate on the hydrolysate than on pure sugars. Both bacteria grew to higher densities on the hydrolysate than on pure sugars (Table 6 and Figure 5), which might be due to the supplementation of the medium with some nutrients originating from the hydrolysate.

Hydrogen production by cultures of C. saccharolyticus and T. neapolitana grown on pure sugars and Miscanthus hydrolysate. Growth (squares) and hydrogen production (circles) in cultures of C. saccharolyticus (a) and T. neapolitana (b) grown on a glucose/xylose (7:3, w w-1) mixture (open symbols) or Miscanthus hydrolysate (filled symbols). The monosaccharide concentration was 14 g l-1.
On pure sugars at 10 g l-1, carbon balances were more than 90%. At increased substrate concentrations the carbon balances often became less, indicating that more non-identified products were formed. Part of these products were volatile components, since COD balances, including the gaseous hydrogen, were still incomplete and volatile components in the gas stream were not collected. The large amount of non-defined products in the T. neapolitana fermentation at high hydrolysate concentration was due to the Maillard reactions.
Hydrogen yield and productivity
The molar yields of products on the consumed substrates are shown in Table 7. The hydrogen yields (YH2) at low-sugar concentrations were generally more than 3 mol H2 per mol hexose, with a maximum of 3.4 mol per mol hexose obtained in fermentations with C. saccharolyticus. For C. saccharolyticus, the hydrogen yield after growth on hydrolysate was equally high as that from pure sugars. The yields with cultures of T. neapolitana on hydrolysate varied between 89% to almost 100% of the yield on pure sugars. The yield dropped substantially at the highest substrate concentrations. The biomass yield (Yxs) of both thermophiles also decreased with increasing substrate concentrations.
The increased production of compounds other than biomass and acetate apparently was not accompanied by hydrogen production. In all fermentations, except those with Maillard reactions, the hydrogen to acetate molar ratio varied between 2.0 to 2.2, which is close to the theoretical value of 2. The CO2 to acetate molar ratio was circa 1.1 at the low substrate concentrations. This ratio increased to 1.3 at the highest substrate concentration, which suggests that a decarboxylation step is involved in the metabolic pathways for the other products. One of these products could be ethanol. Both thermophiles are able to form ethanol, but it is often lost in the gas stream at the high fermentation temperatures because of its volatility.
Most of the hydrogen was produced during the exponential growth phase, but even when the cell density decreased, hydrogen was still produced, although often at a lower rate (Figure 5). The hydrogen production rate by T. neapolitana was very similar on pure sugars and on Miscanthus hydrolysate. The productivity by C. saccharolyticus on hydrolysate was lower than the productivity on pure sugars (Figure 5), which is in line with the lower sugar consumption rate. Apparently, C. saccharolyticus is inhibited by the hydrolysate.
Initially, hydrogen productivity by both thermophiles on low concentrations of Miscanthus hydrolysate was comparable, but after circa 20 h the productivity by C. saccharolyticus declined as compared to that by T. neapolitana (Figure 6). At this point, the cell density of C. saccharolyticus started to decrease (Figure 5). The cell density of T. neapolitana decreased after circa 22 h, but T. neapolitana appeared to be able to maintain reasonable hydrogen productivity as long as the substrate was not completely consumed (Figure 6(b)). The reason for the early decrease in cell density of the thermophiles while substrate is still available is unknown. Some nutrients may have become limiting or the bacteria sense a high cell density and react by arresting growth. When the growth of C. saccharolyticus stopped, the concentration of organic acids (acetate and lactate) was approximately 90 mM. This is far below the concentration of acetate (150 to 175 mM), where cell lysis starts to dominate [22]. At the highest hydrolysate concentration of 28 g l-1 monosaccharides, hydrogen productivity by both thermophiles was low. After circa 20 h, hydrogen and organic acid production by T. neapolitana virtually stopped.

Effect of Miscanthus hydrolysate concentration on the production of hydrogen, acetate and lactate by cultures of C. saccharolyticus and T. neapolitana. Production of hydrogen (circle), acetate (triangle) and lactate (diamond) in cultures of C. saccharolyticus (filled symbols) and T. neapolitana (open symbols) grown on Miscanthus hydrolysate at concentrations of 10 (a), 14 (b) and 28 (c) g monosaccharide l-1.
The maximum volumetric hydrogen productivity (QH2, max) occurred at the late exponential growth phase of the thermophiles. The highest QH2, max of 14.5 mmol l-1 h-1 was observed with T. neapolitana that was fermenting 10 g l-1 of pure sugars (Table 7). The maximum p H2 measured in the off gas was 4.6 kPa. In the C. saccharolyticus fermentations, hydrogen pressures of circa 4 kPa were found, which is significantly lower than the critical value of 10 to 20 kPa. For T. neapolitana the critical value has not been established yet.
Conclusion
Cultures of C. saccharolyticus and T. neapolitana produced hydrogen and acetic acid as the main organic acid during the fermentation of sugars in hydrolysates prepared from the lignocellulosic energy crop Miscanthus. The mild conditions of alkaline treatment enabled the optimized pretreatment protocol to be compatible with the thermophilic fermentations. The formation of potential inhibitors, for example, sugar degradation products, was negligible. For future applicability on an industrial scale this one-stage process needs to be further optimized with respect to alkali usage, for instance by regeneration and recycling of the alkali agent.
Both thermophiles, C. saccharolyticus and T. neapolitana, appeared to be able to simultaneously and completely utilize all soluble monomeric C5 and C6 sugars, di- and oligosaccharides up to a total sugar concentration of 17 g l-1. The capacity of co-fermenting glucose and xylose by C. saccharolyticus was recently confirmed by whole-genome transcriptome analysis [32]. Simultaneous and complete substrate utilization from complex feedstocks, such as the hydrolysates of lignocellulosic biomass, will add to an energy-efficient process and is a major advantage in industrial scale production facilities.
The observed hydrogen yields resulting from the thermophilic fermentations were 74 to 85% of the theoretical value of 4 mol per mol hexose in fermentations with circa 17 g l-1 total sugars. These are amongst the highest hydrogen yields obtained in the fermentation of sugars in lignocellulosic hydrolysates reported to date. C. saccharolyticus offers the advantage of a nearly 10% higher hydrogen yield during growth on Miscanthus hydrolysates as compared to T. neapolitana. However, the rate of substrate consumption and hydrogen production by T. neapolitana was higher. C. saccharolyticus seemed to be hampered by the increase in ionic strength of the culture medium during fermentation and showed lower hydrogen productivity in the stationary phase. Because T. neapolitana is a moderately halophilic organism, it tolerates conditions with higher ionic strength, and possibly with higher osmolalities.
Cultures growing on Miscanthus hydrolysates had low volumetric hydrogen productivity with a maximum of 13 mmol l-1 h-1 and a mean productivity of circa 7 mmol l-1 h-1. Higher volumetric hydrogen productivities have been reported for systems with higher cell densities, such as the carrier-induced granular sludge bed (CIGSB) bioreactor. Productivities of more than 300 mmol H2 l-1 h-1 have been observed, but the hydrogen yield of circa 50% on pure sucrose was low [33].
The COD reduction of the culture medium containing Miscanthus hydrolysate was limited to 30% because the carbohydrates were only partially oxidized to organic acids. More hydrogen can potentially be produced from the organic acids in the effluent by photobacteria, which use light as an extra energy source. Theoretically, another 4 moles of H2 and 2 moles of CO2 can be produced from acetic acid. The first experiment was done with supernatant that was obtained from a culture of T. neapolitana after growth on Miscanthus hydrolysate. Rhodobacter capsulatus, a purple non-sulfur bacterium, was able to grow and produce hydrogen on the supernatant, which was diluted twice, and supplemented with Fe(III)-citrate [34]. The yield of the two fermentations was circa 4.5 moles of hydrogen per mole hexose, that is, 37% of the theoretical value of 12 moles of hydrogen per mole of glucose. Further research is aimed at improving the hydrogen production efficiency of this combined thermophilic and photoheterotrophic fermentation [35].
Methods
Biomass pretreatment and enzymatic hydrolysis
Miscanthus giganteus was collected in the spring of 2004 from a location in Groningen, The Netherlands and consisted primarily of stems. The dry matter content of the harvested stems was circa 80 to 85% on a wet weight basis. The total carbohydrate and lignin content on a dry weight basis was 63% (including 42% glucose, 19% xylose, and 1% arabinose) and 23%, respectively.
Alkaline pretreatment experiments at the lab scale (2 l pulp mixer, Quantum Mark V reactor) were done with milled Miscanthus (Retsch SM 200 mill equipped with a 2 mm screen). An amount of 225 g of biomass (circa 200 g dry matter) was pretreated with 9 to 12% Ca(OH)2 or NaOH (w w-1 dry matter, for details see Tables 1 and 2) at a solid:liquid ratio of 0.125 (w w-1). Pretreatment was done at 85°C for 16 h. One batch of pretreated Miscanthus was washed three times with demineralised water. Prior to enzymatic hydrolysis, the pH of the pretreated material was adjusted to 4.8 to 5.1 with 17% phosphoric, 20% sulfuric or 25% acetic acid (v v-1). Enzymatic hydrolysis of the pretreated biomass was done using commercial enzyme preparations (Cellubrix and Novozymes 188 from Novozymes, Bagsvaerd, Denmark and GC 220 from Genencor, Rochester, NY, USA). The amount of enzyme added per 100 g dry matter was 28, 9, and 13 ml of Cellubrix, Novozymes 188 and GC 220, respectively (for details see Table 2). The enzyme concentration was selected to warrant a similar cellulase activity of 15 IFPU per g dry matter, on the basis of cellulase activity measurements [29]. Incubation was done at 50°C for 24 h.
A pretreatment and hydrolysis experiment was also done at the bench scale in a 10 l stirred vessel. An amount of 1.35 kg (circa 1 kg dry matter) of milled Miscanthus (Pallmann type PS 3–5 knife mill equipped with a screen of 10 mm × 10 mm square opening) was added to the vessel, together with NaOH, under continuous mixing (90 rpm). The amount of NaOH was 9% (w w-1) at a solid:liquid ratio of 0.125 (w w-1). The pretreatment was done for 6 h at 75°C. The material was then dewatered using a manual piston press to make a slurry of 275 g dry matter l-1. The pH of the remaining viscous pulp was adjusted to 5 using a 20% (v v-1) acetic acid solution. Enzymatic hydrolysis was done by a fed-batch procedure of GC 220 addition. The incubation was at 50°C for 24 h.
Hydrolysates were collected after neutralization of the pH and removal of the solids of the enzymatically hydrolyzed material by centrifugation. The hydrolysates were stored at -15°C until use.
Microorganisms and medium
C. saccharolyticus DSM 8903 and T. neapolitana DSM 4359 were obtained from the Deutsche Sammlung von Mikroorganismen und Zellkulturen (DSMZ). The culture medium consisted of (per l) KH2PO4 0.3 g, K2HPO4 0.3 g, MgCl2.6H2O 0.4 g, NH4Cl 0.9 g, yeast extract 1.0 g, cysteine-HCl 0.75 g, FeCl3.6H2O 2.5 mg, SL-10 trace elements 1 ml, and resazurin 0.5 mg. The pH was adjusted to 7.0 at room temperature. NaCl (5 g per l) was added to the culture medium of T. neapolitana. The culture medium for flask experiments was supplemented with 50 mM 4-morpholine propanesulfonic acid (MOPS) to increase the buffering capacity of the medium. A mixture of glucose and xylose (7:3, w w-1) or Miscanthus hydrolysate sugars were used as the carbon source. The medium was made anoxic by flushing with N2. The experiments were carried out under non-sterile conditions. C. saccharolyticus and T. neapolitana were grown at 72 and 80°C, respectively.
The fermentability of hydrolysates was tested using flasks of 118 ml with 20 ml culture medium under a nitrogen atmosphere. The total monosaccharide concentration was 10 g l-1 coming from the pure sugar mixture, the hydrolysate, or a combination of both. The flasks were inoculated with 5% (v v-1) of a preculture that was grown overnight on the same pure sugar mixture. After 16 and 40 h, samples were withdrawn from the headspace (duplicate gas sample of 0.2 ml) and the culture medium (single sample of 1 ml) for analyses of the hydrogen production, cell density, pH, and organic acid production. The experiments were carried out in duplicate (two flasks per condition). The inhibition of compounds was tested using the same method, except that 10 g l-1 of glucose was used as the carbon source.
Batch fermentations under controlled conditions were carried out in a jacketed 2 l bioreactor (Applikon, Delft, The Netherlands) with a working volume of 1 l. The pH was controlled at circa 6.8 (measured at room temperature) by automatic addition of 2 N NaOH. The cultures were continuously stirred at 350 rpm and sparged with N2 at 7 l h-1. Inoculation was done by adding 10% (v v-1) of a preculture that was grown overnight on the glucose/xylose mixture. A fermentation was considered to have ended when the hydrogen concentration was less than 0.2% in the off gas. Samples of 7 ml were regularly taken from the culture medium for measurement of the cell density and substrate and product analyses. Hydrogen and CO2 were measured in the off gas each hour. Data are from one representative fermentation per condition.
Analytical methods
Determination of the acid-soluble and acid-insoluble lignins was performed according to the Tappi method [36]. Organic acids were analyzed by high performance liquid chromatography (HPLC) using a Shodex ionpak KC811 column (Waters, The Netherlands), as described earlier [37]. Monosaccharides, di- and oligosaccharides, furfural, HMF and furfuryl alcohol were analyzed by HPLC using an Altech IOA-1000 column at 90°C, with 3 mM sulfuric acid as the mobile phase (0.4 ml per min), followed by detection by differential refractometry. Fructose was determined enzymatically (Megazyme International Ireland Ltd, Bray, Ireland). Hydrogen in the headspace of the serum bottles and hydrogen and CO2 production in the bioreactors were measured as previously described [37]. COD measurements of the culture medium and hydrolysates were done using the LCK test kit 014 of Hach Lange (Düsseldorf, Germany). The hydrolysis of di- and oligosaccharides in hydrolysates to monomeric sugars was done by the addition of concentrated sulfuric acid (95 to 97%) to a final concentration of 1 M and incubation at 95°C for 1 h. The optical density of the cultures was measured against a water blank at 580 nm after dilution of the culture broth with deionised water. The cell dry weight was determined from the highest value of the optical densities using the relation CDW (g l-1) = (0.377 × OD580) + 0.011 for C. saccharolyticus [37] and CDW (g l-1) = (0.528 × OD580) for T. neapolitana. The molecular weight of T. neapolitana was assumed to be the same as the measured value for C. saccharolyticus, that is, 24.6 g (mol C)-1 [37].
Yield and productivity
The amount of consumed substrate, including non-defined organic compounds, was used for calculating product yields. Yields were expressed as mol product per mol C6 sugar. Because the theoretical hydrogen and acetate yields per C-mol are equal for glucose and xylose [8], the molar amount of xylose was converted to a molar amount of hexose. The consumed unknown organic compounds were determined from COD measurements. They were considered to be carbohydrates with the same product yield as for glucose and xylose. COD in mmol O2 l-1 was converted to mmol hexose l-1, according to the equation C6H12O6 + 6O2 → 6CO2 + 6H2O. The maximum volumetric hydrogen productivity was calculated from the time interval with the highest percentage of hydrogen in the off gas.
References
Li C, Fang HH: Fermentative hydrogen production from wastewater and solid wastes by mixed cultures. Crit Rev Environ Sci Technol 2007, 37: 1-39.
Saratale GD, Chen S-D, Lo Y-C, Saratale RG, Chang J-S: Outlook of biohydrogen production from lignocellulosic feedstock using dark fermentation – a review. J Sci Ind Res 2008, 67: 962-979.
Valdez-Vazquez I, Sparling R, Risbey D, Rinderknecht-Seijas N, Poggi-Varaldo M: Hydrogen generation via anaerobic fermentation of paper mill wastes. Bioresour Technol. 2005,96(17):1907-1913.
Fan Y-T, Zhang Y-H, Zhang S-F, Hou H-W, Ren B-Z: Efficient conversion of wheat straw wastes into biohydrogen gas by cow dung compost. Bioresour Technol. 2006,97(3):500-505.
Levin DB, Islam R, Cicek N, Sparling R: Hydrogen production by Clostridium thermocellum 27405 from cellulosic biomass substrates. Int J Hydrogen Energy 2006, 31: 1496-1503.
Magnusson L, Islam R, Sparling R, Levin D, Cicek N: Direct hydrogen production from cellulosic waste materials with a single-step dark fermentation process. Int J Hydrogen Energy 2008, 33: 5398-5403.
de Vrije T, de Haas GG, Tan GB, Keijsers ER, Claassen PA: Pretreatment of Miscanthus for hydrogen production by Thermotoga elfii . Int J Hydrogen Energy 2002, 27: 1381-1390.
Kádár Z, de Vrije T, van Noorden GE, Budde MA, Szengyel Z, Réczey K, Claassen PA: Yields from glucose, xylose, and paper sludge hydrolysate during hydrogen production by the extreme thermophile Caldicellulosiruptor saccharolyticus . Appl Biochem Biotechnol. 2004, 113–116: 497-508.
Datar R, Huang J, Maness P-C, Mohagheghi A, Czernik S, Chornet E: Hydrogen production from the fermentation of corn stover biomass pretreated with steam-explosion process. Int J Hydrogen Energy 2007, 32: 932-939.
Ren N, Wang A, Gao L, Xin L, Lee D-J, Su A: Bioaugmented hydrogen production from carboxymethyl cellulose and partially delignified corn stalks using isolated cultures. Int j hydrogen Energy 2008, 33: 5250-5255.
Lynd LR, Laser MS, Bransby D, Dale BE, Davison B, Hamilton R, Himmel M, Keller M, McMillan JD, Sheehan J, Wyman CE: How biotech can transform biofuels. Nat Biotechnol. 2008,26(2):169-172.
Mosier N, Wyman CE, Dale B, Elander R, Lee YY, Holtzapple M, Ladisch M: Features of promising technologies for pretreatment of lignocellulosic biomass. Bioresource Technology 2005, 96: 673-686.
Rainey FA, Donnison AM, Janssen PH, Saul D, Rodrigo A, Bergquist PL, Daniel RM, Stackebrandt E, Morgan HW: Description of Caldicellulosiruptor saccharolyticus gen. nov., sp. nov: an obligately anaerobic, extremely thermophilic, cellulolytic bacterium. FEMS Microbiol Lett 1994, 120: 263-266.
Jannasch HW, Huber R, Belkin S, Stetter KO: Thermotoga neapolitana sp. nov. of the extremely thermophilic, eubacterial genus Thermotoga . Arch Microbiol 1988, 150: 103-104.
de Vrije T, Claassen PAM: Dark hydrogen fermentations. In Bio-methane & Bio-hydrogen. Edited by: Reith JH, Wijffels RH, Barten H. The Hague: Smiet Offset; 2003:103-123.
Hallenbeck PC: Fundamentals of the fermentative production of hydrogen. Water Sci Technol 2005, 52: 21-29.
Jones PR: Improving fermentative biomass-derived H 2 -production by engineering microbial metabolism. Int J Hydrogen Energy 2008, 33: 5122-5130.
Blumer-Schuette SE, Kataeva I, Westpheling J, Adams MW, Kelly RM: Extremely thermophilic microorganisms for biomass conversion: status and prospects. Curr Opin Biotechnol. 2008,19(3):210-217.
VanFossen AL, Lewis DL, Nichols JD, Kelly RM: Polysaccharide degradation and synthesis by extremely thermophilic anaerobes. Ann NY Acad Sci 2008, 1125: 322-337.
Donnison AM, Brockelsby CM, Morgan HW, Daniel RM: The degradation of lignocellulosics by extremely thermophilic microorganisms. Biotechnol Bioeng 1989, 33: 1495-1499.
Kim S, Holtzapple MT: Lime pretreatment and enzymatic hydrolysis of corn stover. Bioresource Technology 2005, 96: 1994-2006.
van Niel EW, Claassen PA, Stams AJ: Substrate and product inhibition of hydrogen production by the extreme thermophile, Caldicellulsiruptor saccharolyticus . Biotechnol Bioeng 2003, 81: 255-262.
Boopathy R, Bokang H, Daniels L: Biotransformation of furfural and 5-hydroxymethyl furfural by enteric bacteria. J Industrial Microbiol 1993, 1: 147-150.
Zaldivar J, Martinez A, Ingram LO: Effect of selected aldehydes on the growth and fermentation of ethanologenic Escherichia coli . Biotechnol Bioeng 1999, 65: 24-33.
Ezeji T, Qureshi N, Blaschek HP: Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and butanol fermentation. Biotechnol Bioeng 2007, 97: 1460-1469.
Almeida JR, Roder A, Modig T, Laadan B, Liden G, Gorwa-Grauslund MF: NADH- vs NADPH-coupled reduction of 5-hydroxymethyl furfural (HMF) and its implications on product distribution in Saccharomyces cerevisiae . Appl Microbiol Biotechnol 2008, 78: 939-945.
Almeida JR, Modig T, Roder A, Liden G, Gorwa-Grauslund MF: Pichia stipitis xylose reductase helps detoxifying lignocellulosic hydrolysate by reducing 5-hydroxymethyl-furfural (HMF). Biotechnol Biofuels 2008, 1: 12.
Gutiérrez T, Buszko ML, Ingram LO, Preston JF: Reduction of furfural to furfuryl alcohol by ethanologenic strains of bacteria and its effect on ethanol production from xylose. Appl Biochem Biotechnol. 2002, 98–100: 327-340.
Kabel MA, van der Maarel MJ, Klip G, Voragen AG, Schols HA: Standard assays do not predict the efficiency of commercial cellulase preparations towards plant materials. Biotechnol Bioeng 2005, 93: 56-63.
Vieille C, Hess JM, Kelly RM, Zeikus JG: xyl A cloning and sequencing and biochemical characterization of xylose isomerase form Thermotoga neapolitana . Appl Environ Microbiol 1995, 61: 1867-1875.
Kengen SWM, Goorissen HP, Verhaart M, van Niel EWJ, Claassen PAM, Stams AJM: Biological hydrogen production by anaerobic microorganisms. In Biofuels. Edited by: Soetaert W, Vandamme EJ. Chichester: John Wiley & Sons; 2009:197-221.
Werken HJG, Verhaart MRA, VanFossen AL, Willquist K, Lewis DL, Nichols JD, Goorissen HP, Mongodin EF, Nelson KE, van Niel EWJ, Stams AJM, Ward DE, de Vos WM, Oost J, Kelly RM, Kengen SWM: Hydrogenomics of the extreme thermophilic bacterium Caldicellulosiruptor saccharolyticus . Appl Environ Microbiol 2008, 74: 6720-6729.
Lee K-S, Lo Y-C, Lin P-J, Chang J-S: Improved biohydrogen production in a carrier-induced granular sludge bed by altering physical configuration and agitation pattern of the bioreactor. Int J Hydrogen Energy 2006, 31: 1648-1657.
Uyar B, Schumacher M, Gebicki J, Modigell M: Photoproduction of hydrogen by Rhodobacter capsulatus from thermophilic fermentation effluent. Bioprocess Biosystems Eng 2008.
Claassen PA, de Vrije T: Non-thermal production of pure hydrogen from biomass: HYVOLUTION. Int J Hydrogen Energy 2006, 31: 1416-1423.
TAPPI: T249 cm-85 Carbohydrate composition of extractive-free wood and wood pulp by gas-liquid chromatography. In Test Methods 1998–1999. Atlanta: TAPPI Press; 1999.
de Vrije T, Mars AE, Budde MA, Lai MH, Dijkema C, de Waard P, Claassen PA: Glycolytic pathway and hydrogen yield studies of the extreme thermophile Caldicellulosiruptor saccharolyticus . Appl Microbiol Biotechnol 2007, 74: 1358-1367.
Acknowledgements
This study was financially supported by the Commission of the European Communities, Sixth Framework Programme, Priority 6, Sustainable Energy Systems (019825 HYVOLUTION), the Dutch Programme EET (Economy, Ecology, Technology), a joint initiative of the Ministries of Economic Affairs, of Education, Culture and Sciences and of Housing, Spatial Planning and the Environment (EETK03028 BWPII), and the Dutch Ministry of Agriculture, Nature and Food Quality. Mr. A. Drenth (Agromiscanthus B.V.) is acknowledged for the supply of Miscanthus.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
TdV carried out most of the chemical inhibitor studies and analyzed the chemical composition of the hydrolysate, and drafted the manuscript. RRB performed the alkaline pretreatment, enzymatic hydrolysis experiments and the analysis of hydrolysates, and helped to draft the manuscript. MAWB carried out the 1 litre-scale fermentations. MHL carried out the hydrolysate fermentability tests and some of the chemical inhibitor studies. AEM helped to draft the manuscript. PAMC participated in the coordination of the study and helped to draft the manuscript. All the authors read and accepted the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under license to BioMed Central Ltd. This is an open access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
de Vrije, T., Bakker, R.R., Budde, M.A. et al. Efficient hydrogen production from the lignocellulosic energy crop Miscanthus by the extreme thermophilic bacteria Caldicellulosiruptor saccharolyticus and Thermotoga neapolitana. Biotechnol Biofuels 2, 12 (2009). https://doi.org/10.1186/1754-6834-2-12
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1754-6834-2-12