
Abstract
Feruloyl esterases (FAEs) represent a diverse group of carboxyl esterases that specifically catalyze the hydrolysis of ester bonds between ferulic (hydroxycinnamic) acid and plant cell wall polysaccharides. Therefore, FAEs act as accessory enzymes to assist xylanolytic and pectinolytic enzymes in gaining access to their site of action during biomass conversion. Their ability to release ferulic acid and other hydroxycinnamic acids from plant biomass makes FAEs potential biocatalysts in a wide variety of applications such as in biofuel, food and feed, pulp and paper, cosmetics, and pharmaceutical industries. This review provides an updated overview of the knowledge on fungal FAEs, in particular describing their role in plant biomass degradation, diversity of their biochemical properties and substrate specificities, their regulation and conditions needed for their induction. Furthermore, the discovery of new FAEs using genome mining and phylogenetic analysis of current publicly accessible fungal genomes will also be presented. This has led to a new subfamily classification of fungal FAEs that takes into account both phylogeny and substrate specificity.
Similar content being viewed by others

Background
Plant biomass is a magnificent renewable source of biopolymers. It offers a wealth of possibilities for development and production of sustainable raw materials and energy which fit perfectly with the development of a bio-based economy [1]. Lignocellulosic biomasses from agricultural, agro-industrial, crop, and forestry wastes as well as herbaceous prairie grass, energy crops, and marine algae are regarded as the prospective feedstocks for modern bioethanol and biochemical production [2, 3]. The enzymatic hydrolysis of lignocellulosic biomass has many advantages when compared to chemical conversion in bioethanol production. There is no substrate loss due to chemical modifications; non-corrosive operational conditions may be used and the process is more environmentally friendly [4]. However, plant cell walls have evolved to defend against external factors, including mechanical, thermal, chemical, and biological stress [5, 6]. To efficiently and completely depolymerize different types of lignocellulosic materials, an arsenal of carbohydrate-active and lignin-acting enzymes is required [7, 8]. Feruloyl esterases (FAEs, also known as ferulic/cinnamic acid esterases, EC 3.1.1.73) are responsible for removing ferulic acid residues and cross-links from polysaccharides. They act as accessory (or auxiliary) enzymes that assist the other enzymes in gaining access to their site of action during biomass conversion [9, 10]. In addition to their potential role in bioethanol production, FAEs and their hydrolytic or transesterification products are of great interest for various biotechnological applications, in particular as modified natural antioxidants or food flavor precursors [11, 12]. Therefore, discovery of new FAEs with novel properties and applications is of considerable interest to industry [13]. In this review, we describe (1) the roles of FAEs in plant biomass degradation, (2) an overview of biochemical properties as well as the conditions that induce FAE production, (3) discovery of FAEs and insight into their phylogenetic relationships among fungal genomes, (4) an updated subfamily classification for fungal FAEs and (5) the recent applications of FAEs in biotechnological processes.
Ferulic and hydroxycinnamic acids in plant cell walls
Different types of lignocellulosic biomass can be used for second generation bioethanol production. In order to select the appropriate specificity of enzymes required for biomass degradation, we briefly summarize the occurrence of ferulic and hydroxycinnamic acids in the different types of plant biomass. A variety of hydroxycinnamic acids are present in the plant cell walls (up to 3% of cell wall dry weight), usually esterified or etherified to the polymers within the lignocellulosic matrix [14, 15]. Ferulic acid (ferulate, 4-hydroxy-3-methoxycinnamic acid, mainly trans- or E-form; Fig. 1) and to a lesser extent p-coumaric acid (p-coumarate, 4-hydroxycinnamic acid) are the most abundant hydroxycinnamic acids (hydroxycinnamates) in the plant cell wall polysaccharides. They are regarded as essential and unique structural components in the family Poales of commelinid monocots (e.g., wheat, rice, barley, oat, corn, sorghum, and sugarcane) [15, 16]. Ferulic acid is linked to cell wall polysaccharides mainly through ester-bonds between their carboxyl group and the O-5 hydroxyl group of α-l-arabinofuranosyl residues in glucuronoarabinoxylan (Fig. 1e) [17, 18]. In eudicotyledons, ferulic acid is mainly found in the order ‘core’ Caryophyllales (e.g., sugar beet; [19]). It is ester-linked to pectin at the O-2 and O-5 hydroxyl group of α-l-arabinofuranosyl residues in arabinan as well as at the O-6 hydroxyl group of β-d-galactopyranosyl residue in (arabino-)galactan, both of which are side chains of rhamnogalacturonan I (Fig. 1f) [16, 20–22]. Ferulic acid can oxidatively cross-link to form intermolecular ester-bonds to another arabinoxylan [mainly 5,5′-, 8-O-4′-, 8,5′-, 8,8′-diferulic acids (Fig. 1 g–l)], and ester-ether bonds between polysaccharide and lignin (arabinoxylan–ferulate–lignin) [15, 16, 19, 23–25]. Diferulic acids have been mainly detected in the high-arabinose substitution region of arabinoxylan, because dimerization requires the ferulic acid to be in close proximity [26]. In addition, cross-linking of cell wall polysaccharides and lignin by hydroxycinnamic acids leads to a dramatic increase in mechanical strength of the plant cell wall, decelerates wall extension, and acts as a barrier to block the ingress of microbial invaders as well as hydrolytic enzymes [16, 23, 27].

Model structures of hydroxycinnamic acids, feruloylated plant cell wall polysaccharides and the site of attack by the carbohydrate-active enzymes (modified from [8, 15]). a p-coumaric acid, b caffeic acid, c ferulic acid, d sinapic acid, e feruloylated glucuronoarabinoxylan, f feruloylated pectic rhamnogalacturonan I, g 8,5′-(benzofuran)-diferulic acid, h 8,5′-diferulic acid, i 5,5′-diferulic acid, j 8,4′-diferulic acid, k 8,8′-diferulic acid, l 8,8′-(aryl)-diferulic acid. ABF α-arabinofuranosidase; ABN endoarabinanase; ABX exoarabinanase; AXE acetyl xylan esterase; BXL β-1,4-xylosidase; FAE feruloyl esterase; GAL β-1,4-endogalactanase; GUS α-glucuronidase; LAC β-1,4-galactosidase; RGAE rhamnogalacturonan acetyl esterase; RGL rhamnogalacturonan lyase; RGX exorhamnogalacturonase; RHG endorhamnogalacturonase; XLN β-1,4-endoxylanase
Ferulic acid is also detected in all families of gymnosperms, ester-linked to the primary cell walls, with an amount ranging from 0.01 to 0.16% [28]. However, up to now there is still no identification of which polysaccharides ferulic acid is linked to in gymnosperms [14].
Role of FAEs in plant biomass degradation
Opening up the plant cell wall is a significant part of the process design for bioethanol and biochemical production. Due to the heterogeneity and complexity of the plant cell walls, a variety of carbohydrate- and lignin-active enzyme sets with complementary activities and specificities are required for complete enzymatic hydrolysis of plant biomass (for details see [8, 29]). As ferulic acid is linked to the lignin–carbohydrate complexes, disruption of the ester bond of the lignin–ferulate–arabinoxylan complex is important for complete cell wall deconstruction. FAEs play a key role in providing accessibility for glycoside hydrolases and polysaccharide lyases to the lignocellulose fibers by removal of the ester-bonds between plant polymers [9, 10]. FAEs not only act synergistically with xylanolytic enzymes to convert xylan into its monomers, but have also proved to enhance overall saccharification of lignocellulosic biomass, e.g., wheat straw [30] and sugarcane bagasse [31], when co-incubated with cellulase and xylanase. Moreover, overexpression of FAEs in planta reduces the levels of cell wall esterified phenolics and in most cases also enhances sugar release and improves cell wall digestibility [32–36]. This technique has also been applied to create self-processing transgenic plants that can alter their composition upon activation of the enzyme(s), e.g., to reduce recalcitrance of cell walls prior to saccharification (e.g., [37]; see application section below).
Overview of substrate specificity of characterized FAEs and their other properties
FAE discovery
The first FAEs were discovered in the late 80’s when a new type of esterase capable of releasing the covalently linked ferulic acid from xylan was reported [38–41]. During this period, most FAEs were identified by direct purification from culture supernatant, which required an appropriate induction condition [42–44]. The first fungal FAE encoding genes were identified from Aspergillus niger and Aspergillus tubingensis [45]. Peptide sequencing was used to identify short amino acid sequences of FAEs followed by degenerate or rapid amplification of cDNA ends PCR to obtain the whole gene sequence (Additional file 1: Table S1). Screening of cDNA libraries was also used for discovery of FAEs particularly for anaerobic rumen fungi (e.g., [46–48]). In recent years, publicly available fungal genome sequences have facilitated similarity-based discovery, and genome mining has become the most promising discovery technique (Additional file 1: Table S1). Databases such as carbohydrate-active Enzymes (CAZy) database (http://www.cazy.org; [49]) are very powerful tools for discovery of alternative enzymes of existing families. Discovery of novel enzyme classes or alternative enzymes from enzyme families not included in the database (such as several FAE families) requires other approaches. However, it should be noted that FAEs are a very diverse enzymes, so similarity-based discovery does not necessarily guarantee the same function.
Activity and properties of FAEs
Although FAEs have been identified in various plant cell wall degrading microbes, to date fungi are still the main source of FAEs used in industry [50]. Thorough data collection for the physicochemical properties of purified FAEs has been previously reported [9, 50–53], and therefore here we only present the properties of characterized fungal FAEs for which amino acid sequences have been reported (Additional file 1: Table S1; see also update classification section).
FAEs are active in a broad pH (from pH 3 to 10) and temperature (from 20 to 75 °C) range, but generally they are mainly active at pH 4–7 and temperatures below 50 °C (Additional file 1: Table S1; [50, 53]). A few reports also showed the effect of metal ions and inhibitors on FAEs [54–57]. It should be noted that the structures of only two fungal FAEs have been reported until now: A. niger (AnFaeA—[58–61]) and Aspergillus oryzae (AoFaeB—[62]), of which only AoFaeB contains a calcium binding site in its structure. It is located far from the active site but may have a role in stabilizing the protein structure. FAEs catalyze the hydrolysis of the substrate following the mechanism utilized by serine proteases [63] with a conserved Ser-His-Asp/Glu catalytic triad [64]. Glu as a part of catalytic triad, instead of Asp, was recently reported in several Basidiomycetes, which is uncommon among FAEs, but found in some members of the α/β-hydrolase-fold superfamily [57, 65, 66]. Differences in amino acid residues within loops and domains that situate in close proximity to the catalytic and substrate binding sites enable different FAEs to target different substrates [59, 62, 64]. The catalytic mechanism of FAEs involves two steps, the initial acylation of the nucleophilic serine residue forming acyl-enzyme intermediate followed by deacylation of the intermediate. In the deacylation step, nucleophilic water (hydrolysis) or other hydroxyl molecule from e.g., carbohydrate or alcohol (transesterification, see also industrial applications section) can attack the intermediate and cause the release of the product [58, 64].
Different substrates were used for characterization of FAEs: polysaccharides (e.g., wheat bran and sugar beet pulp [67]), feruloylated oligosaccharides (e.g., feruloylate-Ara-Xyl1–3, feruloylate-Ara1–3, p-coumaroylate-Ara-Xyl1–3 [22]), and monomeric hydroxycinnamate model substrates (e.g., methyl, ethyl, p-nitrophenyl, or α-naphthyl ferulate [46, 68–70]). Short chain fatty acid model substrates (e.g., α-naphthyl acetate, umbelliferyl acetate, and umbelliferyl butyrate) are also used for the activity assay. However, they only show whether the enzyme is active, but not whether they are specific to ferulic or hydroxycinnamic acid.
Reversed phase HPLC/UV is the most used technique for detection of ferulic and hydroxycinnamic acids, and their release from feruloylated poly- and oligosaccharides [22, 71, 72]. However, it is time consuming and usually requires prior isolation/extraction step before the analysis, which makes it less useful for rapid or high-throughput screening [70]. For the activity screening, a spectrophotometry-based method using monomeric hydroxycinnamate model substrates which detects the release of chromophore group (e.g., p-nitrophenyl, α-naphthyl ferulate) or the reduction of substrate (e.g., methyl, ethyl ferulate) is rapid and easy to perform. The spectrophotometric assay is widely accepted, even though there is a concern about the spectral overlapping between substrate and product, e.g., methyl substrates and their aromatic acids. Recently, other methods have been developed such as high-performance thin layer chromatography and electrochemical sensor for rapid detection of ferulic acid which may be useful for enzymatic screening [73, 74].
Several fungi produce more than one FAE isozyme and different substrates are required to determine their substrate specificity. The classical examples are two A. niger FAEs, AnFaeA and AnFaeB [45, 67, 75]. Regarding the monomeric substrates, AnFaeA is specific for ferulic and sinapic acid methyl esters, while AnFaeB is specific for ferulic, p-coumaric, and caffeic acid methyl esters (Fig. 1a–d). Of the oligomeric substrates (derived from wheat bran and sugar beet pulp), AnFaeA catalyzes the hydrolysis of the feruloylated (1,5) arabinosyl xylo-oligosaccharides from wheat arabinoxylan, but is less active towards feruloylated (1,2) arabino- and (1,6) galacto-oligosaccharides from sugar beet pulp. AnFaeB is active towards feruloylated oligosaccharides derived from both monocot and dicot cell walls [15, 68, 76]. Regarding the polymeric substrates, both FAEs also show opposite substrate preference. AnFaeA highly prefers to hydrolyze wheat arabinoxylan over sugar beet pectin and can also release the diferulic acid (5,5′, 8-O-4′), whereas AnFaeB is more active towards sugar beet pectin but cannot release diferulic acid [15, 75].
Inducing substrates, regulation, and production
Production of FAEs in nature depends highly on the available carbon sources or inducing compounds. Ferulic acid, and related hydroxycinnamic acids (e.g., caffeic, p-coumaric acids) and phenolic compounds (e.g., vanillic acid, vanillin, and veratric acid) can induce the production of FAEs [75]. Feruloylated plant biomass such as wheat bran, sugar beet pulp/pectin, and maize bran are frequently used as substrates for production of FAEs (Additional file 1: Table S1). Recent transcriptomic data from different fungal species suggested that the low- to non-feruloylated biofuel feedstocks such as the straw from wheat, barley, corn, rice, and soybean as well as the woody substrates from both softwood (pine) and hardwood (aspen) can substantially upregulate the expression of fae genes [77–80]. Although the presence of ferulic acid in the cultivation is not absolutely required, addition of ferulic acid can considerably improve the production of FAEs [76]. Xylose induces the production of AnFaeA, whereas most monosaccharides do not appear to support the production of other FAEs [75].
Detailed expression studies of FAE encoding genes are rare and have so far been mainly performed in species of the genus Aspergillus. Here expression of fae genes is presumed to be controlled by at least three independent regulatory systems [81]. The xylanolytic transcriptional activator XlnR, a zinc binuclear cluster motif (Zn(II)2Cys6), is a key factor in the regulation of hemicellulolytic and cellulolytic genes in Aspergilli [82–84]. In A. niger, faeA and other genes encoding xylan degrading enzymes (e.g., xlnB, xlnC, xlnD, axeA, axhA, and aguA) are under control of XlnR [81, 82]. Another major regulator that is responsible for carbon catabolite repression in many filamentous fungi is the conserved zinc-finger regulator CreA [85, 86]. Expression of faeA was influenced by the balance between induction by XlnR and repression by CreA, whereas faeB was not activated by XlnR, but still sensitive to CreA-mediated repression [75, 87]. creA deletion mutants showed improved production of secreted lignocellulose degrading enzymes including FAEs [75, 77]. In addition, both faeA and faeB are expressed in the presence of ferulic acid and other hydroxycinnamic acids [75], indicating the presence of a ferulic acid- or hydroxycinnamic acid-responsive transcriptional regulator. It should be noted that the ferulic acid induction is independent of XlnR and the combined ferulic acid induction and XlnR effect on expression of A. niger faeA is larger than the sum of the two effects alone [81]. However, it is unclear whether the ferulic acid- or hydroxycinnamic acid induction is mediated by a single regulatory system since different sets of phenolic compounds induced the expression of faeA and faeB [75, 88]. As FaeA is only found in Aspergilli and related species (see below), it is currently unclear to which extent XlnR orthologs in other fungi are involved in activating expression of FAE encoding genes.
Native fungal FAEs are produced mainly through two types of cultivation techniques: submerged/liquid fermentation in which fungi are grown in liquid medium often with vigorous aeration; and solid-state fermentation in which they grow on moist solid substrates such as lignocellulosic biomass. Although FAE production from native sources can reach high levels, e.g., >106 mU/mL for submerged fermentation of Aspergillus awamori [54] and >103 mU/g for solid-state fermentation of Penicillium brasilianum [89], production of FAEs from native sources faces considerable complications e.g., the choices of suitable substrates, control of fermentation conditions, up-scaling and the purification process [90]. Over the past decade, FAE production has shifted more towards heterologous mainly using two expression hosts, i.e., Escherichia coli and Pichia pastoris under the isopropyl β-d-1-thiogalactopyranoside (IPTG) or methanol inducible promoters, respectively, for Academia (Additional file 1: Table S1). For industry, the established platforms of the company are being used. Heterologous production offers several advantages over native production, such as well-established cultivation conditions for up-scaling, fusion of affinity tags for downstream processing and possibilities for enzyme engineering.
Classification of FAEs
The initial classification of FAEs was based on the induction and substrate specificity of AnFaeA and AnFaeB [15, 91]. Subsequently, based on the substrate specificity towards four model substrates (methyl ferulate, sinapate, caffeate, and p-coumarate) and the ability to release diferulic acid, FAEs were classified into four types (A, B, C and D) [92]. Type A FAEs prefer substrates containing methoxy substitutions at C-3 and/or C-5 as found in ferulic and sinapic acids, and are active towards methyl p-coumarate. They are also capable of releasing 5,5′- and 8-O-4′-diferulic acids. Type B FAEs prefer substrates containing one or two hydroxyl substitutions, as found in p-coumaric and caffeic acids, respectively. Hydrolytic rates of type B FAEs are significantly reduced when a methoxy group is present and they are not active against methyl sinapate. In addition, type B FAEs cannot release diferulic acid. Type C and D FAEs possess broader substrate specificity with activity towards all four model substrates, but only type D can release diferulic acid from plant cell walls [92].
The ABCD classification was very useful and initially was supported by phylogenetic analysis because a limited number of amino acid sequences of FAEs were available at that time. As more FAEs were characterized, it no longer adequately reflects the wealth of putative FAEs encoded by microbial/fungal genomes. Hence, a refined classification was introduced based on phylogenetic analysis of available fungal genomes, which separated FAEs into seven subfamilies (SF1-7) [51]. This classification demonstrated that FAEs evolved from highly divergent esterase families (tannases (SF1-4), acetyl xylan esterases (SF6), and lipases (SF7)) and do not have a common ancestor, even though they all contain a conserved Ser-His-Asp catalytic triad [51]. The availability of fungal genome sequences also enabled a more detailed comparison of the diversity and prevalence of putative FAEs [93, 94]. Although FAEs are carbohydrate-active enzymes, they are only partially included in CAZy database [49] as some FAEs (SF5, SF6) belong to carbohydrate esterase family 1 (CE1) together with acetyl xylan esterases. More recently, a further refined classification was proposed by clustering 365 FAE-related amino acid sequences using descriptor-based computational analysis and machine learning algorithms [52]. At the same time, pharmacophore models for specific FAE subgroups were also developed, which will be useful for production of FAE-based biosynthetic compounds. The descriptor-based classification separated the FAEs into 12 families; however, some of these families were further divided into subgroups (A–D) to distinguish the substrate specificity of characterized FAEs within the family.
Update on the classification of fungal FAEs
New phylogenetic tree based on all published fungal genomes
Based on the previously reported phylogenetic analysis [51], we reconstructed a novel phylogenetic tree using 20 amino acid sequences from characterized FAEs (Table 1) and a BLASTP search against 247 published fungal genomes (Additional file 1: Table S2). All resulting amino acid sequences with an expect value lower than 1E−40 were collected. Duplicate and incomplete sequences as well as sequences with ambiguous amino acids (X) were discarded. Signal peptides were predicted using SignalP [95] and removed from all candidate sequences. This analysis resulted in 1251 putative FAE sequences, which were aligned using Multiple Sequence Comparison by Log-Expectation (MUSCLE). Phylogenetic analysis was performed using the neighbor-joining method with pairwise deletion of gaps and the Poisson correction distance of substitution rates (statistical support for phylogenetic grouping was estimated by 1000 bootstrap re-samplings) of the Molecular Evolutionary Genetics Analysis (MEGA 6) program [96]. A few characterized acetyl xylan esterases, lipases, and tannases were included in the analysis to reveal the relationships of FAEs with those enzymes. In this analysis, the xylanase-related FAEs (e.g., FAEs from Aspergillus terreus (AtFAE-2, AtFAE-3) [56]) were not included in the similarity search because they showed similarity to GH10 and GH11 xylanases. Including these xylanase-related FAEs also recognized other non-FAE members of these two families, which could not be screened out because only two fungal xylanase-related FAEs were identified. Also, a putative FAE from Xylaria polymorpha (XpoGH78) [97] was not included in the phylogenetic tree because it showed no relationship with other FAE sequences.
Previously, the phylogenetic analysis classified the FAEs into seven subfamilies [51] (Table 1; Additional file 1: Table S1). SF1 contained FAEs from A. niger (AnFaeB [75]) and A. oryzae (AoFaeB, AoFaeC [98]) which are closely related to tannases. SF5 contained FAEs from A. nidulans (AN5267 [99]) and Neurospora crassa (NcFaeD [91]) and some members of this subfamily belong to CE1 in the CAZy database. SF6 contained FAEs from Chaetomium sp. CQ31 (ChaeFAE [100]) and Talaromyces funiculosus (FaeB [69]) which also belong to CE1 and are closely related to acetyl xylan esterases. SF7 contained exclusively FAEs from Aspergillus spp., e.g., A. niger (AnFaeA [45]), Aspergillus flavus (AfFaeA [101]), and A. oryzae (AoFaeA [102]), which are closely related to lipases. SF2–SF4 only contained putative FAEs, which showed sequence similarity to SF1 and tannases (Table 1). Our new phylogenetic analysis classified the putative FAEs into 13 subfamilies (Fig. 2; Additional file 2: Figure S1). In comparison with the previous phylogenetic analysis [51], members of SF1–SF3 and SF5–SF7 remain classified to the same subfamilies. In addition, a FAE from Fusarium oxysporium (FoFaeC [103]) has been characterized, which belongs to SF2 and SF7 members that were expanded to cover other fungi than Aspergillus spp. (e.g., Jaapia argillacea, Penicillium rubens, and Armillaria mellea). Subfamily SF8 contains FAEs from Auricularia auricular-judae (EstBC [104]), Anaeromyces mucronatus (Fae1A [48]), and Orpinomyces sp. (OrpFaeA [105]), while SF12 contains Pleurotus sapidus (Est1 [106]) and Pleurotus eryngii (PeFaeA [57]) FAEs, for which there were no homologs found in the previous study [51]. The new subfamily SF9 separated from SF4 which previously contained a putative FAE from A. oryzae (BAE66413). Three tannases (A. fumigatus (XP_748839 [107]), A. niger (ABX89592 [108]), and A. oryzae (BAA09656 [109])) were positioned in SF11, indicating that the enzymes of this subfamily may actually possess tannase activity or potentially dual-activity and may not be true FAEs. The study also resulted in new subfamilies SF10 and SF13. By contrast, no closely related homologs were found for the FAEs from e.g., Piromyces equi (PeEstA [110]), Piromyces sp. (FaeA [47]), and Coprinopsis cinerea (CcEst1 [111]) and together with the other sequences which are not classified in any group, these are referred to as ungrouped sequences (U1–U10, Fig. 2; Additional file 2: Figure S1). These ungrouped sequences may develop into new subfamilies if homologs for them are discovered.

Phylogenetic relationships among the (putative) fungal FAEs. Glucuronoyl esterases (GEs, green-filled circles) were used as an outgroup. AXE, acetyl xylan esterase (blue-filled circles); LIP, lipase (yellow-filled squares); SF, subfamily; TAN, tannase (purple-filled circles). FAEs from previously reported phylogenetic analysis [51] were marked with magenta open triangles for SF1, magenta open rhombuses for SF2-4, magenta-filled triangles for SF5, magenta-filled rhombuses for SF6, light blue-filled squares for SF7, brown-filled circles for AtFAE2 and AtFAE3, and magenta-filled circles for ungrouped ones. Light blue-filled circles indicate ungrouped sequences which numbering indicates different groups
Reflection on origin of the different types of FAEs and comparison with ABCD classification
As mentioned before, FAEs evolved from a diverse class of enzymes (e.g., tannases, acetyl xylan esterases, lipases, and choline esterases). Most FAEs have evolved from tannases, as enzymes belonging to the subfamilies SF1-4 and SF9-11 are related to tannases. SF5 and SF6 enzymes show relationship with acetyl xylan esterases, whereas SF7 enzymes are related to lipases. SF12 and SF13 are related to both lipases and choline esterases. Some FAEs are also related to xylanases (GH10 and GH11) and α-l-rhamnosidases (GH78), whereas some show no similarity to any of the above enzymes (Additional file 1: Table S1). Having evolved from different types of enzymes may explain why different FAEs target different hydroxycinnamic acids. While the ABCD classification system provides hints for the specificity of putative FAEs [92], it no longer reflects the evolutionary relationships among different FAEs [51, 52]. In comparison with the ABCD system, SF6 and SF7 contain solely type B and A FAEs, respectively, whereas SF1 contains both type B and C FAEs, and SF5 a mix of type A and D FAEs (Table 1). SF1 and SF5 may be further divided to support ABCD classification when more FAEs from these subfamilies are characterized. In addition, the two new subfamilies SF8 and SF12, which are distantly related to SF7, also contain type A FAEs, whereas FAEs from Ustilago maydis (SF13) which are distantly related to SF6 also possess type B activity. Therefore, the ABCD system needs to be revisited and combined with the phylogeny-based classification to provide a well-based system that will help in the identification of different types of FAEs and predict the properties of newly discovered FAEs.
Prevalence of different types of FAEs in fungal genomes
From the 247 published fungal genomes in early 2015, 155 of them contained putative FAEs (Tables 2a, b; Additional file 2: Figure S2). Approximately 10% of genomes had only one putative FAE and, surprisingly, almost 25 and 5% of the analyzed fungal genomes contained more than 10 and 20 putative FAEs, respectively. The basidiomycetes Auricularia subglabra and Moniliophthora roreri possessed more than 30 putative FAEs followed by the ascomycetes A. niger, Aspergillus luchuensis (formerly A. kawachii), Oidiodendron maius, Colletotrichum gloeosporioides with more than 20 putative FAEs. This variation in FAE content could be related to the different abilities of the fungi to degrade feruloylated substrates, which in turn may be related to the presence of such substrates in their natural habitat. However, the multiplicity of putative FAEs identified here could include pseudogenes and the similarity-based method could result in the inclusion of other FAE-related enzymes, e.g., SF11 may also contain tannases. We summarized the prevalence of putative FAEs in industrially and ecologically important fungi in Table 2. Most of these fungi produce more than one type of FAEs. It should be noted that our findings are in agreement with the earlier study reporting that Trichoderma reesei (syn. Hypocrea jecorina) does not have any putative FAEs in its genome [112], and therefore supplementation of FAEs can significantly increase the saccharification efficiency of an enzyme cocktail from T. reesei [30, 31, 67]. However, two other species of this genus, T. atroviride and T. virens, contain three putative FAEs in their genomes.
Industrial applications of FAEs
With the ability to remove hydroxycinnamic acids from plant cell walls, FAEs have considerable roles in biotechnological processes for various industrial applications. Earlier Fazary and Ju [113] excellently reviewed the early industrial use of FAEs through patents. To date the patents on FAE applications and discovery are almost doubled compared to 2008. In this section, we update the patents on FAEs presented in European Parliament documents (EP) and World Intellectual Property Organization-Patent Cooperation Treaty (WIPOPCT) databases (Additional file 1: Table S3), and highlight the applications in five major fields: (1) biomass processing, (2) ferulic acid and related fine chemicals production, (3) pulp and paper, (4) feed and (5) seasonings and alcoholic beverages (Fig. 3).

Schematic overview of industrial applications of FAEs (modified from [51])
Applications in biomass processing
FAEs are considered to be essential accessory enzymes to complete hydrolysis of lignocellulosic biomass for bioethanol and other biorefineries. To date more than 150 patents have been filed on applications of FAEs towards biomass processing (Additional file 1: Table S3, both discovery and saccharification). Activity of FAEs on plant biomass in combination with other hydrolases and oxidases not only significantly increases the breakdown of plant materials and enhances the availability of fermentable carbohydrates, but it also releases phenolic compounds and toxic esters which inhibit the fermentation process of pretreated lignocellulosic materials (e.g., [114]). For this reason, fusions of FAEs and other enzymes/proteins have also been created aiming to increase the catalytic efficiency and/or substrate affinity [115–117]. Different strategies have been applied to create FAE mutants which can tolerate the high temperatures in bioprocesses [118–120]. Furthermore, transgenic plants have been manipulated specifically for biofuel production to reduce recalcitrance of cell walls prior to saccharification, which also enhance the digestibility and biomass conversion for livestock (e.g., [32–35, 121]). Besides, FAEs are not only used for complete hydrolysis of lignocellulosic materials, but they can also be applied for manipulating the structure of oligosaccharides e.g., in production of xylo-oligosaccharides [122] which are industrially important functional food additives with prebiotic properties [123].
Applications in production of ferulic acid and related fine chemicals
Ferulic acid and other hydroxycinnamic acids are phenolic phytochemicals which are widely used in food and cosmetic industries because of their unique and potent properties as, e.g.,
-
1.
Antioxidant—they are able to neutralize free radicals, e.g., reactive oxygen species which are implicated to cause DNA damage, cancer, and accelerated cell aging [124–126].
-
2.
Sun protection factor—they are able to absorb UV radiation by the presence of conjugated double bonds, e.g., in an aromatic structure [124, 127].
-
3.
Depigmenting agent—they are tyrosinase inhibitors because their chemical structures resemble those of tyrosine and are suggested to prevent the formation of melanin by competitive inhibition with tyrosine [128, 129].
-
4.
Precursor for synthesis of flavor compounds, such as vanillin and guaiacol (e.g., 4-vinyl guaiacol)—intermediates of ferulic acid degradation pathway. These intermediates are of great interest in the food and fragrance industry [130, 131].
Ferulic acid and other hydroxycinnamic acids can be used as a carrier of vitamin C and E, which double their skin photoprotection with stronger lipophilicity allowing better penetration into the stratum corneum [132]. Furthermore, they show pharmaceutical and health beneficial functions, e.g., antimicrobial, anti-inflammatory, anti-diabetic, anti-thrombosis, anti-cancer, and cholesterol-lowering agents [11, 12, 133]. Although commercially ferulic acid is mainly produced from rice oil (as γ-oryzanol), modern processes are focusing on production of ferulic acid by FAEs in combination with other hydrolases in a biorefinery process (e.g., [121, 122]).
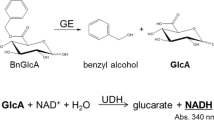
Apart from hydrolysis, FAEs can be used for synthesis of ester-linked hydroxycinnamic acids through a transesterification reaction by exchanging the organic group of an ester (donor) with the organic group of an alcohol (acceptor) (Fig. 4), to obtain products with altered chemical and biological properties. The first report on transesterification activity of FAE was investigated on FAE from Sporotrichum thermophile (StFaeC) using arabinose and arabinobiose as acceptors [134]. Containing both hydrophobic ferulic acid and hydrophilic oligosaccharide moieties, feruloylated arabinose and oligosaccharides possess the physiological functions of both. This includes antioxidant activity, probiotic effects, and inhibition against glycation which are of interest by a wide range of applications in food, pharmaceutical, and cosmetic industries [135]. The advantage of using transesterification over hydrolases or transferases is the flexibility of their acceptor molecules, which can vary from different carbohydrates [66, 136, 137], aliphatic and aromatic alcohols [138, 139], and glycerol [140, 141] to propolis [142]. In the latter case, FAEs can also be used for impoverishing the allergenicity of propolis by specifically removing esters of caffeic acid under hydrolytic conditions [143].

Transferuloylation reaction (modified from [64])
Applications in pulp and paper industry
To produce high-quality paper, whiteness is an important characteristic of wood pulp. Discoloration of the pulp is caused by lignin remaining in the pulp and bleaching is the key step to whiten the pulp by removing the residual lignin. This process uses hazardous and expensive chemicals; mainly chlorine dioxide and hydrogen peroxide or ozone in elemental chlorine-free (ECF) and in totally chlorine-free (TCF) chemical processes, respectively. In the environmentally friendly biobleaching process, FAEs can be used in combination with xylanases and lignin-oxidizing enzymes [144–146] particularly in a bi-sequential process reported by Record et al. [144], which the delignification rates were comparable to the results obtained with hazardous chemicals. The enzymatic process also resulted in lower energy consumption and a significant reduction of the chemical oxygen demand (COD) value of the pulping waste water [147].
Applications in feed industry
Fiber digestibility is an essential criterion for animal feed. Suffering from improper digestion can hamper animal growth and cause immunological stress which results in reduction of the feed conversion ratio in livestock, and hence restricts profitability of farmers. Ferulic and hydroxycinnamic acids themselves can promote health in animals [148, 149]; however, feruloylation in plant cell walls particularly in a high forage diet is among the major inhibitory factors for the ruminant digestive system. Addition of FAEs or enzyme cocktails containing FAEs can improve the access of main chain degrading enzymes resulting in improved fiber digestion and bioavailability of phytonutrients, accelerating animal growth (e.g., [150, 151]), as well as reducing immunological stress [152].
Applications in seasonings and alcoholic beverage industry
Surprisingly, FAEs have been used for both removing off-flavors/odors as well as enhancing the aroma in several seasonings and alcoholic beverages. Flavor and odor are the crucial ingredients for success in the premium fermented seasonings and alcoholic beverage industries in particular Japanese rice wine and cooking liquor—sake and mirin. The major flavor component from these products is ferulic acid as well as its derivatives including 4-vinyl guaiacol, vanillic acid, and vanillin. FAEs can be applied in the saccharification process as a FAE-producing koji (rice-fungal culture starter) or an additive together with xylanases and cellulases to increase the release of ferulic acids from the cell wall of rice and other cereal grains, which then can be converted to the aromatic derivatives during the fermentation and aging process [54, 153–155].
Other applications
Apart from the above-mentioned applications (Additional file 1: Table S3), FAEs can also be used (1) in a form of live FAE-producing Lactobacilli supplement which can reduce triglyceride concentrations, hepatic inflammation and insulin resistance in medical applications [133, 156]; (2) in the milling process for starch production, where FAE is used during the wet milling together with e.g., cellulase and proteases providing an increase in production yield [157]; and (3) in detergent applications, where FAE-containing multi-enzyme system is used to improve the performance of liquid laundry detergents particularly at low temperature (e.g., [158]).
Conclusion
In this review, we provide insight into biodiversity, biochemical properties, production, and discovery of FAEs, a highly diverse group of plant cell wall degrading enzymes. Although FAEs generally play a role in catalyzing the release of ferulic acid and other hydroxycinnamic acids from plant cell wall polysaccharides, they possess diverse specificities towards different feruloylated poly- and oligosaccharides and monomeric hydroxycinnamates. FAEs have evolved from different types of enzymes (e.g., tannases, acetyl xylan esterases, and lipases), which is reflected by their amino acid sequences. Classification based on phylogenetic analysis divided FAEs into distinct groups and also resulted in discovery of novel putative FAEs. These new FAE candidates may possess different substrate specificities and/or biochemical properties which may be useful in different applications. It is clear that more biochemical characterization of FAEs is needed for better understanding of substrate specificity and mode of action of FAEs from different subfamilies. The range of industrial applications of FAEs has been broadened over the past years with emphasis on the conversion of agro-industrial waste materials into valuable products and the synthesis of novel ester-linked hydroxycinnamic products in particular for health and cosmetic applications. The industrial uses of FAEs are still limited to only a few enzymes. Here, we provided the phylogenetic-based classification and putative FAEs resulting from genome mining as a guideline for exploration of FAEs towards the specific applications.
Abbreviations
- CAZy:
-
carbohydrate-active enzymes database
- FAE:
-
feruloyl esterase
- SF:
-
subfamily
References
Aguilar A, Magnien E, Thomas D. Thirty years of European biotechnology programmes: from biomolecular engineering to the bioeconomy. Nat Biotechnol. 2013;30:410–25.
Carroll A, Somerville C. Cellulosic biofuels. Annu Rev Plant Biol. 2009;60:165–82.
Sanchez OJ, Cardona CA. Trends in biotechnological production of fuel ethanol from different feedstocks. Bioresour Technol. 2008;99:5270–95.
Selig M, Knoshaug E, Decker S, Baker J, Himmel M, Adney W. Heterologous expression of Aspergillus niger β-d-xylosidase (XlnD): characterization on lignocellulosic substrates. Appl Biochem Biotechnol. 2008;146:57–68.
Mortimer JC, Miles GP, Brown DM, Zhang Z, Segura MP, Weimar T, Yu X, Seffen KA, Stephens E, Turner SR, Dupree P. Absence of branches from xylan in Arabidopsis gux mutants reveals potential for simplification of lignocellulosic biomass. Proc Natl Acad Sci USA. 2010;107:17409–14.
Silveira RL, Stoyanov SR, Gusarov S, Skaf MS, Kovalenko A. Plant biomass recalcitrance: effect of hemicellulose composition on nanoscale forces that control cell wall strength. J Am Chem Soc. 2013;135:19048–51.
Himmel ME, Ding S-Y, Johnson DK, Adney WS, Nimlos MR, Brady JW, Foust TD. Biomass recalcitrance: engineering plants and enzymes for biofuels production. Science. 2007;315:804–7.
Mäkelä MR, Hildén KS, de Vries RP. Degradation and modification of plant biomass by fungi. In: Nowrousian M, editor. Mycota vol XIII fungal genomics. Berlin: Springer-Verlag; 2014. p. 175–208.
Wong DWS. Feruloyl esterase—a key enzyme in biomass degradation. Appl Biochem Biotechnol. 2006;133:87–112.
Faulds CB. What can feruloyl esterases do for us? Phytochem Rev. 2010;9:121–32.
Ou S, Kwok K-C. Ferulic acid: pharmaceutical functions, preparation and applications in foods. J Sci Food Agr. 2004;84:1261–9.
Kumar N, Pruthi V. Potential applications of ferulic acid from natural sources. Biotechnol Rep. 2014;4:86–93.
Gopalan N, Rodríguez-Duran LV, Saucedo-Castaneda G, Nampoothiri KM. Review on technological and scientific aspects of feruloyl esterases: a versatile enzyme for biorefining of biomass. Bioresour Technol. 2015;193:534–44.
Harris PJ. Diversity in plant cell walls. In: Henry RJ, editor. Plant diversity and evolution: genotypic and phenotypic variation in higher plants. Oxon: CABI Publishing; 2005. p. 201–28.
Kroon PA, Garcia-Conesa MT, Fillingham IJ, Hazlewood GP, Williamson G. Release of ferulic acid dehydrodimers from plant cell walls by feruloyl esterases. J Sci Food Agr. 1999;79:428–34.
de Buanafina MMO. Feruloylation in grasses: current and future perspectives. Mol Plant. 2009;2:861–72.
Carpita NC. Structure and biogenesis of the cell walls of grasses. Annu Rev Plant Physiol Plant Molec Biol. 1996;47:445–76.
Harris PJ, Trethewey JAK. The distribution of ester-linked ferulic acid in the cell walls of angiosperms. Phytochem Rev. 2010;9:19–33.
Waldron KW, Parr AJ, Ng A, Ralph J. Cell wall esterified phenolic dimers: identification and quantification by reverse phase high performance liquid chromatography and diode array detection. Phytochem Anal. 1996;7:305–12.
Levigne S, Ralet MC, Quemener B, Thibault JF. Isolation of diferulic bridges ester-linked to arabinan in sugar beet cell walls. Carbohydr Res. 2004;339:2315–9.
Levigne SV, Ralet MCJ, Quemener BC, Pollet BNL, Lapierre C, Thibault JFJ. Isolation from sugar beet cell walls of arabinan oligosaccharides esterified by two ferulic acid monomers. Plant Physiol. 2004;134:1173–80.
Ralet MC, Thibault JF, Faulds CB, Williamson G. Isolation and purification of feruloylated oligosaccharides from cell walls of sugar-beet pulp. Carbohydr Res. 1994;263:227–41.
Iiyama K, Lam TBT, Stone BA. Covalent cross-links in the cell-wall. Plant Physiol. 1994;104:315–20.
Ralph J, Quideau S, Grabber JH, Hatfield RD. Identification and synthesis of new ferulic acid dehydrodimers present in grass cell walls. J Chem Soc-Perkin Trans. 1994;1:3485–98.
Vismeh R, Lu F, Chundawat SPS, Humpula JF, Azarpira A, Balan V, Dale BE, Ralph J, Jones AD. Profiling of diferulates (plant cell wall cross-linkers) using ultrahigh-performance liquid chromatography-tandem mass spectrometry. Analyst. 2013;138:6683–92.
Waldron KW, Faulds CB. 1.05—Cell wall polysaccharides: composition and structure. In: Kamerling H, editor. Comprehensive glycoscience. Oxford: Elsevier; 2007. p. 181–201.
de Oliveira DM, Finger-Teixeira A, Rodrigues Mota T, Salvador VH, Moreira-Vilar FC, Correa Molinari HB, Craig Mitchell RA, Marchiosi R, Ferrarese-Filho O, dos Santos WD. Ferulic acid: a key component in grass lignocellulose recalcitrance to hydrolysis. Plant Biotechnol J. 2015;13:1224–32.
Carnachan SM, Harris PJ. Ferulic acid is bound to the primary cell walls of all gymnosperm families. Biochem Syst Ecol. 2000;28:865–79.
van den Brink J, de Vries RP. Fungal enzyme sets for plant polysaccharide degradation. Appl Microbiol Biotechnol. 2011;91:1477–92.
Tabka MG, Herpoël-Gimbert I, Monod F, Asther M, Sigoillot JC. Enzymatic saccharification of wheat straw for bioethanol production by a combined cellulase xylanase and feruloyl esterase treatment. Enzyme Microb Technol. 2006;39:897–902.
Gottschalk LMF, Oliveira RA, Bon EPDS. Cellulases, xylanases, β-glucosidase and ferulic acid esterase produced by Trichoderma and Aspergillus act synergistically in the hydrolysis of sugarcane bagasse. Biochem Eng J. 2010;51:72–8.
Buanafina M, Langdon T, Hauck B, Dalton S, Morris P. Expression of a fungal ferulic acid esterase increases cell wall digestibility of tall fescue (Festuca arundinacea). Plant Biotechnol J. 2008;6:264–80.
Buanafina MMD, Langdon T, Hauck B, Dalton S, Timms-Taravella E, Morris P. Targeting expression of a fungal ferulic acid esterase to the apoplast, endoplasmic reticulum or golgi can disrupt feruloylation of the growing cell wall and increase the biodegradability of tall fescue (Festuca arundinacea). Plant Biotechnol J. 2010;8:316–31.
Buanafina MMD, Langdon T, Hauck B, Dalton SJ, Morris P. Manipulating the phenolic acid content and digestibility of Italian ryegrass (Lolium multiflorum) by vacuolar-targeted expression of a fungal ferulic acid esterase. Appl Biochem Biotechnol. 2006;130:416–26.
Badhan A, Jin L, Wang YX, Han SY, Kowalczys K, Brown DCW, Ayala CJ, Latoszek-Green M, Miki B, Tsang A, McAllister T. Expression of a fungal ferulic acid esterase in alfalfa modifies cell wall digestibility. Biotechnol Biofuels. 2014;7:1.
Dunn-Coleman N, Langdon T, Morris P. Manipulation of the phenolic acid content and digestibility of plant cell walls by targeted expression of genes encoding cell wall degrading enzymes. World Patent. 2002. WO 2002068666 A1, 6 Sep 2002.
Lanahan MB, Basu SS, Batie CJ, Chen W, Craig J, Kinkema M. Self processing plants and plant parts. 2009. US Patent, US 20090320831 A1, 31 Dec 2009.
MacKenzie CR, Bilous D. Ferulic acid esterase activity from Schizophyllum commune. Appl Environ Microbiol. 1988;54:1170–3.
Johnson KG, Harrison BA, Schneider H, Mackenzie CR, Fontana JD. Xylan-hydrolysing enzymes from Streptomyces spp. Enzyme Microb Technol. 1988;10:403–9.
Johnson KG, Silva MC, Mackenzie CR, Schneider H, Fontana JD. Microbial degradation of hemicellulosic materials. Appl Biochem Biotechnol. 1989;20–1:245–58.
McDermid KP, MacKenzie CR, Forsberg CW. Esterase activities of Fibrobacter succinogenes subsp. succinogenes S85. Appl Environ Microbiol. 1990;56:127–32.
Faulds CB, Williamson G. The purification and characterization of 4-hydroxy-3-methoxycinnamic (ferulic) acid esterase from Streptomyces olivochromogenes. J Gen Microbiol. 1991;137:2339–45.
Borneman WS, Ljungdahl LG, Hartley RD, Akin DE. Isolation and characterization of p-coumaroyl esterase from the anaerobic fungus Neocallimastix strain MC-2. Appl Environ Microbiol. 1991;57:2337–44.
Borneman WS, Ljungdahl LG, Hartley RD, Akin DE. Purification and partial characterization of two feruloyl esterases from the anaerobic fungus Neocallimastix strain MC-2. Appl Environ Microbiol. 1992;58:3762–6.
de Vries RP, Michelsen B, Poulsen CH, Kroon PA, van den Heuvel RHH, Faulds CB, Williamson G, van den Hombergh JPTW, Visser J. The faeA genes from Aspergillus niger and Aspergillus tubingensis encode ferulic acid esterases involved in degradation of complex cell wall polysaccharides. Appl Environ Microbiol. 1997;63:4638–44.
Fillingham IJ, Kroon PA, Williamson G, Gilbert HJ, Hazlewood GP. A modular cinnamoyl ester hydrolase from the anaerobic fungus Piromyces equi acts synergistically with xylanase and is part of a multiprotein cellulose-binding cellulase-hemicellulase complex. Biochem J. 1999;343:215–24.
Harhangi HR, Akhmanova AS, Emmens R, van der Drift C, de Laat WTAM, van Dijken JP, Jetten MSM, Pronk JT, Op den Camp HJM. Xylose metabolism in the anaerobic fungus Piromyces sp. strain E2 follows the bacterial pathway. Arch Microbiol. 2003;180:134–41.
Qi M, Wang P, Selinger LB, Yanke LJ, Forster RJ, McAllister TA. Isolation and characterization of a ferulic acid esterase (Fae1A) from the rumen fungus Anaeromyces mucronatus. J Appl Microbiol. 2011;110:1341–50.
Lombard V, Golaconda Ramulu H, Drula E, Coutinho PM, Henrissat B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014;42:D490–5.
Topakas E, Vafiadi C, Christakopoulos P. Microbial production, characterization and applications of feruloyl esterases. Process Biochem. 2007;42:497–509.
Benoit I, Danchin EGJ, Bleichrodt RJ, de Vries RP. Biotechnological applications and potential of fungal feruloyl esterases based on prevalence, classification and biochemical diversity. Biotechnol Lett. 2008;30:387–96.
Udatha DBRKG, Kouskoumvekaki I, Olsson L, Panagiotou G. The interplay of descriptor-based computational analysis with pharmacophore modeling builds the basis for a novel classification scheme for feruloyl esterases. Biotechnol Adv. 2011;29:94–110.
Katsimpouras C, Antonopoulou I, Christakopoulos P, Topakas E. Role and applications of feruloyl esterases in biomass bioconversion. In: Gupta VK, editor. Microbial enzymes in bioconversions of biomass. Berlin: Springer; 2016 (in press).
Kanauchi M, Watanabe S, Tsukada T, Atta K, Kakuta T, Koizumi T. Purification and characteristics of feruloyl esterase from Aspergillus awamori G-2 strain. J Food Sci. 2008;73:C458–63.
Koseki T, Furuse S, Iwano K, Matsuzawa H. Purification and characterization of a feruloyl esterase from Aspergillus awamori. Biosci Biotechnol Biochem. 1998;62:2032–4.
Kumar CG, Kamle A, Kamal A. Purification and biochemical characterization of feruloyl esterases from Aspergillus terreus MTCC 11096. Biotechnol Prog. 2013;29:924–32.
Nieter A, Haase-Aschoff P, Linke D, Nimtz M, Berger RG. A halotolerant type A feruloyl esterase from Pleurotus eryngii. Fungal Biol. 2014;118:348–57.
McAuley KE, Svendsen A, Patkar SA, Wilson KS. Structure of a feruloyl esterase from Aspergillus niger. Acta Crystallogr Sect D Biol Crystallogr. 2004;60:878–87.
Hermoso JA, Sanz-Aparicio J, Molina R, Juge N, Gonzalez R, Faulds CB. The crystal structure of feruloyl esterase a from Aspergillus niger suggests evolutive functional convergence in feruloyl esterase family. J Mol Biol. 2004;338:495–506.
Faulds CB, Molina R, Gonzalez R, Husband F, Juge N, Sanz-Aparicio J, Hermoso JA. Probing the determinants of substrate specificity of a feruloyl esterase, AnFaeA, from Aspergillus niger. FEBS J. 2005;272:4362–71.
Benoit I, Asther M, Sulzenbacher G, Record E, Marmuse L, Parsiegla G, Gimbert I, Asther M, Bignon C. Respective importance of protein folding and glycosylation in the thermal stability of recombinant feruloyl esterase A. FEBS Lett. 2006;580:5815–21.
Suzuki K, Hori A, Kawamoto K, Thangudu RR, Ishida T, Igarashi K, Samejima M, Yamada C, Arakawa T, Wakagi T, et al. Crystal structure of a feruloyl esterase belonging to the tannase family: a disulfide bond near a catalytic triad. Proteins. 2014;82:2857–67.
Kraut J. Serine proteases: structure and mechanism of catalysis. Ann Rev Biochem. 1977;46:331–58.
Prates JAM, Tarbouriech N, Charnock SJ, Fontes CMGA, Ferreira LSMA, Davies GJ. The structure of the feruloyl esterase module of xylanase 10B from Clostridium thermocellum provides insights into substrate recognition. Structure. 2001;9:1183–90.
Nieter A, Haase-Aschoff P, Kelle S, Linke D, Krings U, Popper L, Berger RG. A chlorogenic acid esterase with a unique substrate specificity from Ustilago maydis. Appl Environ Microbiol. 2015;81:1679–88.
Kelle S, Nieter A, Krings U, Zelena K, Linke D, Berger RG. Heterologous production of a feruloyl esterase from Pleurotus sapidus synthesizing feruloyl-saccharide esters. Biotechnol Appl Biochem. 2015. doi:10.1002/bab.1430.
Faulds CB, Williamson G. Purification and characterization of a ferulic acid esterase (FAE-III) from Aspergillus niger: specificity for the phenolic moiety and binding to microcrystalline cellulose. Microbiology. 1994;140:779–87.
Kroon PA, Williamson G. Release of ferulic acid from sugar-beet pulp by using arabinanase, arabinofuranosidase and an esterase from Aspergillus niger. Biotechnol Appl Biochem. 1996;23:263–7.
Kroon PA, Williamson G, Fish NM, Archer DB, Belshaw NJ. A modular esterase from Penicillium funiculosum which releases ferulic acid from plant cell walls and binds crystalline cellulose contains a carbohydrate binding module. Eur J Biochem. 2000;267:6740–52.
Mastihuba V, Kremnický L, Mastihubová M, Willett JL, Côté GL. A spectrophotometric assay for feruloyl esterases. Anal Biochem. 2002;309:96–101.
Juge N, Williamson G, Puigserver A, Cummings NJ, Connerton IF, Faulds CB. High-level production of recombinant Aspergillus niger cinnamoyl esterase (FAEA) in the methylotrophic yeast Pichia pastoris. FEMS Yeast Res. 2001;1:127–32.
Dobberstein D, Bunzel M. Separation and detection of cell wall-bound ferulic acid dehydrodimers and dehydrotrimers in cereals and other plant materials by reversed phase high-performance liquid chromatography with ultraviolet detection. J Agric Food Chem. 2010;58:8927–35.
Hingse SS, Digole SB, Annapure US. Method development for simultaneous detection of ferulic acid and vanillin using high-performance thin layer chromatography. J Anal Sci Technol. 2014;5:1–9.
Liu L-J, Gao X, Zhang P, Feng S-L, Hu F-D, Li Y-D, Wang C-M. Ultrasensitive detection of ferulic acid using poly(diallyldimethylammonium chloride) functionalized graphene-based electrochemical sensor. J Anal Methods Chem. 2014;2014:424790.
de Vries RP, VanKuyk PA, Kester HCM, Visser J. The Aspergillus niger faeB gene encodes a second feruloyl esterase involved in pectin and xylan degradation and is specifically induced in the presence of aromatic compounds. Biochem J. 2002;363:377–86.
Faulds CB, de Vries RP, Kroon PA, Visser J, Williamson G. Influence of ferulic acid on the production of feruloyl esterases by Aspergillus niger. FEMS Microbiol Lett. 1997;157:239–44.
Delmas S, Pullan ST, Gaddipati S, Kokolski M, Malla S, Blythe MJ. Uncovering the genome-wide transcriptional responses of the filamentous fungus Aspergillus niger to lignocellulose using RNA sequencing. PLOS Genet. 2012;8:e1002875.
Pullan ST, Daly P, Delmas S, Ibbett R, Kokolski M, Neiteler A, van Munster JM, Wilson R, Blythe MJ, Gaddipati S, et al. RNA-sequencing reveals the complexities of the transcriptional response to lignocellulosic biofuel substrates in Aspergillus niger. Fungal Biol Biotechnol. 2014;1:1–14.
Vanden Wymelenberg A, Gaskell J, Mozuch M, Splinter BonDurant S, Sabat G, Ralph J, Skyba O, Mansfield SD, Blanchette RA, Grigoriev IV, et al. Significant alteration of gene expression in wood decay fungi Postia placenta and Phanerochaete chrysosporium by plant species. Appl Environ Microbiol. 2011;77:4499–507.
Wang B, Cai P, Sun W, Li J, Tian C, Ma Y. A transcriptomic analysis of Neurospora crassa using five major crop residues and the novel role of the sporulation regulator rca-1 in lignocellulose production. Biotechnol Biofuels. 2015;8:1–15.
De Vries RP, Visser J. Regulation of the feruloyl esterase (faeA) gene from Aspergillus niger. Appl Environ Microbiol. 1999;65:5500–3.
van Peij NNME, Gielkens MMC, de Vries RP, Visser J, de Graaff LH. The transcriptional activator XlnR regulates both xylanolytic and endoglucanase gene expression in Aspergillus niger. Appl Environ Microbiol. 1998;64:3615–9.
Marui J, Kitamoto N, Kato M, Kobayashi T, Tsukagoshi N. Transcriptional activator, AoXlnR, mediates cellulose-inductive expression of the xylanolytic and cellulolytic genes in Aspergillus oryzae. FEBS Lett. 2002;528:279–82.
Tamayo EN, Villanueva A, Hasper AA, Graaff LH, Ramon D, Orejas M. CreA mediates repression of the regulatory gene xlnR which controls the production of xylanolytic enzymes in Aspergillus nidulans. Fungal Genet Biol. 2008;45:984–93.
Dowzer CE, Kelly JM. Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol Cell Biol. 1991;11:5701–9.
Dowzer CE, Kelly JM. Cloning of the creA gene from Aspergillus nidulans: a gene involved in carbon catabolite repression. Curr Genet. 1989;15:457–9.
de Vries RP, Visser J, de Graaff LH. CreA modulates the XlnR-induced expression on xylose of Aspergillus niger genes involved in xylan degradation. Res Microbiol. 1999;150:281–5.
Mäkelä MR, Jiménez Barboza LA, de Vries RP, Hildén KS. Production of feruloyl esterases by Aspergillus species. In: de Vries RP, Benoit I, Andersen M, editors. Aspergillus and penicillium in the post-genomic era. Norfolk: Caister Academic Press; 2016.
Panagiotou G, Granouillet P, Olsson L. Production and partial characterization of arabinoxylan-degrading enzymes by Penicillium brasilianum under solid-state fermentation. Appl Microbiol Biotechnol. 2006;72:1117–24.
Koyani RD, Rajput KS. Solid state fermentation: comprehensive tool for utilization of lignocellulosic through biotechnology. J Bioprocess Biotech. 2015;5:1000258.
Crepin VF, Faulds CB, Connerton IF. Identification of a type-d feruloyl esterase from Neurospora crassa. Appl Microbiol Biotechnol. 2003;63:567–70.
Crepin VF, Faulds CB, Connerton IF. Functional classification of the microbial feruloyl esterases. Appl Microbiol Biotechnol. 2004;63:647–52.
Coutinho PM, Andersen MR, Kolenova K, vanKuyk PA, Benoit I, Gruben BS. Post-genomic insights into the plant polysaccharide degradation potential of Aspergillus nidulans and comparison to Aspergillus niger and Aspergillus oryzae. Fungal Genet Biol. 2009;46:S161–9.
Benoit I, Culleton H, Zhou M, DiFalco M, Aguilar-Osorio G, Battaglia E, Bouzid O, Brouwer CPJM, El-Bushari HBO, Coutinho PM, et al. Closely related fungi employ diverse enzymatic strategies to degrade plant biomass. Biotechnol Biofuels. 2015;8:107.
Petersen TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat Meth. 2011;8:785–6.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30:2725–9.
Nghi DH, Bittner B, Kellner H, Jehmlich N, Ullrich R, Pecyna MJ, Nousiainen P, Sipilä J, Huong LM, Hofrichter M, Liers C. The wood rot ascomycete Xylaria polymorpha produces a novel GH78 glycoside hydrolase that exhibits α-l-rhamnosidase and feruloyl esterase activities and releases hydroxycinnamic acids from lignocelluloses. Appl Environ Microbiol. 2012;78:4893–901.
Koseki T, Hori A, Seki S, Murayama T, Shiono Y. Characterization of two distinct feruloyl esterases, AoFaeB and AoFaeC, from Aspergillus oryzae. Appl Microbiol Biotechnol. 2009;83:689–96.
Debeire P, Khoune P, Jeltsch JM, Phalip V. Product patterns of a feruloyl esterase from Aspergillus nidulans on large feruloyl-arabino-xylo-oligosaccharides from wheat bran. Bioresour Technol. 2012;119:425–8.
Yang S-Q, Tang L, Yan Q-J, Zhou P, Xu H-B, Jiang Z-Q, Zhang P. Biochemical characteristics and gene cloning of a novel thermostable feruloyl esterase from Chaetomium sp. J Mol Catal B Enzyme. 2013;97:328–36.
Zhang SB, Zhai HC, Wang L, Yu GH. Expression, purification and characterization of a feruloyl esterase A from Aspergillus flavus. Protein Expr Purif. 2013;92:36–40.
Zeng Y, Yin X, Wu M-C, Yu T, Feng F, Zhu T-D, Pang Q-F. Expression of a novel feruloyl esterase from Aspergillus oryzae in Pichia pastoris with esterification activity. J Mol Catal B Enzym. 2014;110:140–6.
Moukouli M, Topakas E, Christakopoulos P. Cloning, characterization and functional expression of an alkalitolerant type C feruloyl esterase from Fusarium oxysporum. Appl Microbiol Biotechnol. 2008;79:245–54.
Haase-Aschoff P, Linke D, Nimtz M, Popper L, Berger RG. An enzyme from Auricularia auricula-judae combining both benzoyl and cinnamoyl esterase activity. Process Biochem. 2013;48:1872–8.
Blum DL, Kataeva IA, Li XL, Ljungdahl LG. Feruloyl esterase activity of the Clostridium thermocellum cellulosome can be attributed to previously unknown domains of XynY and XynZ. J Bacteriol. 2000;182:1346–51.
Linke D, Matthes R, Nimtz M, Zorn H, Bunzel M, Berger RG. An esterase from the basidiomycete Pleurotus sapidus hydrolyzes feruloylated saccharides. Appl Microbiol Biotechnol. 2013;97:7241–51.
Nierman WC, Pain A, Anderson MJ, Wortman JR, Kim HS, Arroyo J, Berriman M, Abe K, Archer DB, Bermejo C, et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature. 2005;438:1151–6.
Fuentes-Garibay JA, Aguilar CN, Rodríguez-Herrera R, Guerrero-Olazarán M, Viader-Salvadó JM. Tannase sequence from a xerophilic Aspergillus niger strain and production of the enzyme in Pichia pastoris. Mol Biotechnol. 2015;57:439–47.
Hatamoto O, Watarai T, Kikuchi M, Mizusawa K, Sekine H. Cloning and sequencing of the gene encoding tannase and a structural study of the tannase subunit from Aspergillus oryzae. Gene. 1996;175:215–21.
Poidevin L, Levasseur A, Paes G, Navarro D, Heiss-Blanquet S, Asther M, Record E. Heterologous production of the Piromyces equi cinnamoyl esterase in Trichoderma reesei for biotechnological applications. Lett Appl Microbiol. 2009;49:673–8.
Hashimoto K, Kaneko S, Yoshida M. Extracellular carbohydrate esterase from the basidiomycete Coprinopsis cinerea released ferulic and acetic acids from xylan. Biosci Biotechnol Biochem. 2010;74:1722–4.
Martinez D, Berka RM, Henrissat B, Saloheimo M, Arvas M, Baker SE, Chapman J, Chertkov O, Coutinho PM, Cullen D, et al. Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea jecorina). Nat Biotechnol. 2008;26:1193.
Fazary AE, Ju YH. The large-scale use of feruloyl esterases in industry. Biotechnol Mol Biol Rev. 2008;3:95–110.
West SI, William HG. Use of type c and d feruloyl esterases in the manufacture of biofuels. 2010. US Patent, US 20100256353 A1, 7 Oct 2010.
Wu M, Gong Y, Yin X, Zeng Y, Li J. Yeast system for co-expressing faeA and xyn11A. 2013. Chinese Patent, CN 102994542 A, 27 Mar 2013.
Levasseur A, Saloheimo M, Navarro D, Andberg M, Monot F, Nakari-Setälä T, Asther M, Record E. Production of a chimeric enzyme tool associating the Trichoderma reesei swollenin with the Aspergillus niger feruloyl esterase A for release of ferulic acid. Appl Microbiol Biotechnol. 2006;73:872–80.
Levasseur A, Navarro D, Punt PJ, Belaïch J-P, Asther M, Record E. Construction of engineered bifunctional enzymes and their overproduction in Aspergillus niger for improved enzymatic tools to degrade agricultural by-products. Appl Environ Microbiol. 2005;71:8132–40.
Yin X, Li J-F, Wang C-J, Hu D, Wu Q, Gu Y, Wu M-C. Improvement in the thermostability of a type A feruloyl esterase, AuFaeA, from Aspergillus usamii by iterative saturation mutagenesis. Appl Microbiol Biotechnol. 2015;99:10047–56.
Zhang SB, Pei XQ, Wu ZL. Multiple amino acid substitutions significantly improve the thermostability of feruloyl esterase A from Aspergillus niger. Bioresour Technol. 2012;117:140–7.
Cao L-C, Chen R, Xie W, Liu Y-H. Enhancing the thermostability of feruloyl esterase EstF27 by directed evolution and the underlying structural basis. J Agric Food Chem. 2015;63:8225–33.
Dhugga KD, Dolde D, Gupta R, Sandhu AP, Simmons CR. Methods to alter plant cell wall composition for improved biofuel production and silage digestibility. 2013. World Patent, WO 2013063006 A1, 2 May 2013.
Li X, Fang B, Cheng S, Zhang L. Method for simultaneously preparing ferulic acid, xylo-oligosaccharides and ethanol from spent grains. 2013. Chinese Patent, CN 102864180 A, 9 Jan 2013.
Aachary AA, Prapulla SG. Xylooligosaccharides (XOS) as an emerging prebiotic: microbial synthesis, utilization, structural characterization, bioactive properties, and applications. Compr Rev Food Sci Food Saf. 2011;10:2–16.
Graf E. Antioxidant potential of ferulic acid. Free Radic Biol Med. 1992;13:435–48.
RiceEvans CA, Miller NJ, Paganga G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radic Biol Med. 1996;20:933–56.
Lombard DB, Chua KF, Mostoslavsky R, Franco S, Gostissa M, Alt FW. DNA repair, genome stability, and aging. Cell. 2005;120:497–512.
Nappa A, Lorenzini F, Sanhueza A. Compositions and methods to add value to plant products, increasing the commercial quality, resistance to external factors and polyphenol content thereof. 2007. US Patent, US 20070232495 A1, 4 Oct 2007.
Kim HJ, Seo SH, Lee BG, Lee YS. Identification of tyrosinase inhibitors from Glycyrrhiza uralensis. Planta Med. 2005;71:785–7.
Takahashi T, Miyazawa M. Tyrosinase inhibitory activities of cinnamic acid analogues. Pharmazie. 2010;65:913–8.
Di Gioia D, Luziatelli F, Negroni A, Ficca AG, Fava F, Ruzzi M. Metabolic engineering of Pseudomonas fluorescens for the production of vanillin from ferulic acid. J Biotechnol. 2011;156:309–16.
Priefert H, Rabenhorst J, Steinbuchel A. Biotechnological production of vanillin. Appl Microbiol Biotechnol. 2001;56:296–314.
Lin F-H, Lin J-Y, Gupta RD, Tournas JA, Burch JA, Angelica Selim M, Monteiro-Riviere NA, Grichnik JM, Zielinski J, Pinnell SR. Ferulic acid stabilizes a solution of vitamins C and E and doubles its photoprotection of skin. J Invest Dermatol. 2005;125:826–32.
Lorca GL. Lactobacillus supplement for alleviating type 1 diabetes. 2015. US Patent, US 20150182567 A1, 2 Jul 2015.
Topakas E, Vafiadi C, Stamatis H, Christakopoulos P. Sporotrichum thermophile type C feruloyl esterase (StFaeC): purification, characterization, and its use for phenolic acid (sugar) ester synthesis. Enzyme Microb Technol. 2005;36:729–36.
Ou J, Sun Z. Feruloylated oligosaccharides: structure, metabolism and function. J Funct Foods. 2014;7:90–100.
Sakamoto R, Murata S, Tsuchiyama M: 2005. New method for producing ferulic acid ester. Japanese Patent, JP2005224177 A, issued date Aug 25, 2005.
Vafiadi C, Topakas E, Christakopoulos P, Faulds CB. The feruloyl esterase system of Talaromyces stipitatus: determining the hydrolytic and synthetic specificity of TsFaeC. J Biotechnol. 2006;125:210–21.
Togawa M, Masuda H, Kishimoto N: 2003. Method for producing ester of cinnamic acid. Japanese Patent, JP2003033196 A, issued date Feb 04, 2003.
Topakas E, Stamatis H, Biely P, Kekos D, Macris BJ, Christakopoulos P. Purification and characterization of a feruloyl esterase from Fusarium oxysporum catalyzing esterification of phenolic acids in ternary water—organic solvent mixtures. J Biotechnol. 2003;102:33–44.
Kawasaki H, Sakamoto R, Murata S, Tsuchiyama M. Method for producing ferulic acid ester compound with enzymatic method. 2007. Japanese Patent, JP2007000010 A, 11 Jan 2007.
Vafiadi C, Topakas E, Nahmias VR, Faulds CB, Christakopoulos P. Feruloyl esterase-catalysed synthesis of glycerol sinapate using ionic liquids mixtures. J Biotechnol. 2009;139:124–9.
Nagase Y, Yamada Y, Suzuki K, Sakamoto T, Araki Y, Mishima S. Esterified propolis, method for production of the same, anticancer agent, and food formulation. 2007. Japanese Patent, JP2007053947 A, 8 Mar 2007.
Ricchiuto GM, Gardana CS, Guglielmetti SD. Process for obtaining non-allergic propolis. 2013. US Patent, US 20130006011 A1, 3 Jan 2013.
Record E, Asther M, Sigoillot C, Pagès S, Punt PJ, Delattre M, Haon M, van den Hondel CAMJJ, Sigoillot JC, Lesage-Meessen L, Asther M. Overproduction of the Aspergillus niger feruloyl esterase for pulp bleaching application. Appl Microbiol Biotechnol. 2003;62:349–55.
Sigoillot C, Camarero S, Vidal T, Record E, Asther M, Pérez-Boada M, Martínez MJ, Sigoillot J-C, Asther M, Colom JF, Martínez ÁT. Comparison of different fungal enzymes for bleaching high-quality paper pulps. J Biotechnol. 2005;115:333–43.
Tapin S, Sigoillot J-C, Asther M, Petit-Conil M. Feruloyl esterase utilization for simultaneous processing of nonwood plants into phenolic compounds and pulp fibers. J Agric Food Chem. 2006;54:3697–703.
Xu Z, Ma J, Men Z, Chen Z, Wang X, Tan F, Hu UY. Complex enzyme preparation and technique for preparing dissolving pulp by using same. 2013. Chinese Patent, CN 103061181 A, 24 Apr 2013.
Aimutis R, Casper JL, Finnie SM, Campayo VG. Increasing the free phenolic acid in grain-based foods. 2015. World Patent, WO 2015048727 A1, 2 Apr 2015.
Cao B-B, Jin X, Yang H-J, Li S-L, Jiang L-S. Microbial release of ferulic and p-coumaric acids from forages and their digestibility in lactating cows fed total mixed rations with different forage combinations. J Sci Food Agric. 2016;96:650–5.
Jayaraman S, Mukkalil R, Chirakkal H. Use of ferulic acid esterase to improve performance in monogastric animals. 2015. US Patent, US20150118361 A1, 30 Apr 2015.
Lynch JP, Prema D, Van Hamme JD, Church JS, Beauchemin KA. Fiber degradability, chemical composition and conservation characteristics of alfalfa haylage ensiled with exogenous fibrolytic enzymes and a ferulic acid esterase-producing inoculant. Can J Anim Sci. 2014;94:697–704.
Anderson DM, Hsiao HY, Liu L. Enzymes for reduced immunological stress. 2011. US Patent, US 20110177195 A1, 21 Jul 2011.
Hiroyuki Y, Kyoko H, HideakiI K, Yuto Y, Kumiko K. Fermented alcoholic beverage with high content of ferulic acid. 2010. Japanese Patent, JP2010148485 A, 8 Jul 2010.
Uno T, Itoh A, Miyamoto T, Kubo M, Kanamaru K, Yamagata H, Yasufuku Y, Imaishi H. Ferulic acid production in the brewing of rice wine (Sake). J Inst Brew. 2009;115:116–21.
Kanauchi M. Characteristics and role of feruloyl esterase from Aspergillus awamori in Japanese spirits, ‘Awamori’ production. In: Valdez B, editor. Scientific, health and social aspects of the food industry. Rijeka: InTech; 2012. p. 145–62.
Prakash S, Bhathena JR. Oral polymeric membrane feruloyl esterase producing bacteria formulation. 2010. US Patent, US 20100047320 A1, 25 Feb 2010.
Han W, Mclaughlin SR, Long Z. Milling process. 2014. World Patent, WO 2014135063 A1, 12 Sep 2014.
Brooker AT, Somerville RNP, Miracle GS, Lant NJ, Souter PF, Forrest M, Ure C. A catalytic laundry detergent composition comprising relatively low levels of water-soluble electrolyte. 2011. World Patent, WO 2011005730 A1, 13 Jan 2011.
Shin HD, Chen RRZ. A type B feruloyl esterase from Aspergillus nidulans with broad pH applicability. Appl Microbiol Biotechnol. 2007;73:1323–30.
Sakamoto T, Nishimura S, Kato T, Sunagawa Y, Tsuchiyama M, Kawasaki H. Efficient extraction of ferulic acid from sugar beet pulp using the culture supernatant of Penicillium chrysogenum. J Appl Glycosci. 2005;52:115–20.
Garcia-Conesa M-T, Crepin VF, Goldson AJ, Williamson G, Cummings NJ, Connerton IF, Faulds CB, Kroon PA. The feruloyl esterase system of Talaromyces stipitatus: production of three discrete feruloyl esterases, including a novel enzyme, TsFaeC, with a broad substrate specificity. J Biotechnol. 2004;108:227–41.
Damasio ARL, Braga CMP, Brenelli LB, Citadini AP, Mandelli F, Cota J, de Almeida RF, Salvador VH, Paixao DAA, Segato F, et al. Biomass-to-bio-products application of feruloyl esterase from Aspergillus clavatus. Appl Microbiol Biotechnol. 2013;97:6759–67.
Kühnel S, Pouvreau L, Appeldoorn MM, Hinz SWA, Schols HA, Gruppen H. The ferulic acid esterases of Chrysosporium lucknowense C1: purification, characterization and their potential application in biorefinery. Enzyme Microb Technol. 2012;50:77–85.
Knoshaug EP, Selig MJ, Baker JO, Decker SR, Himmel ME, Adney WS. Heterologous expression of two ferulic acid esterases from Penicillium funiculosum. Appl Biochem Biotechnol. 2008;146:79–87.
Topakas E, Moukouli M, Dimarogona M, Christakopoulos P. Expression, characterization and structural modelling of a feruloyl esterase from the thermophilic fungus Myceliophthora thermophila. Appl Microbiol Biotechnol. 2012;94:399–411.
Crepin VF, Faulds CB, Connerton IF. A non-modular type B feruloyl esterase from Neurospora crassa exhibits concentration-dependent substrate inhibition. Biochem J. 2003;370:417–27.
Fazary AE, Ismadji S, Ju YH. Studies on temperature dependent kinetics of Aspergillus awamori feruloyl esterase in water solutions. Kinet Catal. 2010;51:31–7.
Koseki T, Takahashi K, Fushinobu S, Iefuji H, Iwano K, Hashizume K, Matsuzawa H. Mutational analysis of a feruloyl esterase from Aspergillus awamori involved in substrate discrimination and pH dependence. Biochim Biophys Acta. 2005;1722:200–8.
Gong YY, Yin X, Zhang HM, Wu MC, Tang CD, Wang JQ, Pang QF. Cloning, expression of a feruloyl esterase from Aspergillus usamii E001 and its applicability in generating ferulic acid from wheat bran. J Ind Microbiol Biotechnol. 2013;40:1433–41.
Authors’ contributions
RPDV coordinated the manuscript. AD wrote the first draft of the manuscript. MRM, IBG, KSH, RPDV contributed to manuscript revision and approved the final version. AD, MVAP, RPDV performed bioinformatics analysis. All authors read and approved the final manuscript.
Acknowledgements
The authors would like to thank Miaomiao Zhou for her support with the initial bioinformatics work.
Competing interests
The authors declare that they have no competing interests.
Funding
This work was supported by the European Union, Grant Agreement No: 613868 (OPTIBIOCAT). MVAP was supported by a grant of the Dutch Technology Foundation STW, Applied Science division of NWO and the Technology Program of the Ministry of Economic Affairs 016.130.609 to RPDV.
Author information
Authors and Affiliations
Corresponding author
Additional files
13068_2016_651_MOESM1_ESM.xls
Additional file 1: Table S1. Substrate specificity, biochemical properties, induction conditions, classifications based on the amino acid sequences, Table S2. The prevalence of the FAE families in all fungal genomes used in this review article, Table S3. Patents and the inventions related to FAE applications, Table S4. Sequences of characterized and putative FAEs used in this study.
13068_2016_651_MOESM2_ESM.pdf
Additional file 2: Figure S1. Phylogenetic tree of the (putative) fungal FAEs. FAEs from previously reported phylogenetic analysis [51] were marked with magenta open triangles for SF1, magenta open rhombuses for SF2-4, magenta filled triangles for SF5, magenta filled rhombuses for SF6, light blue filled squares for SF7, and magenta filled circles for ungrouped ones. AtFAE2 and AtFAE3 are marked with brown filled circles, acetyl xylan esterases are marked with blue filled circles, lipases are marked with yellow filled squares, tannases are marked with purple filled circles, glucuronoyl esterases (as an outgroup) were marked with green filled circles. The same symbols are used in Fig. 2.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Dilokpimol, A., Mäkelä, M.R., Aguilar-Pontes, M.V. et al. Diversity of fungal feruloyl esterases: updated phylogenetic classification, properties, and industrial applications. Biotechnol Biofuels 9, 231 (2016). https://doi.org/10.1186/s13068-016-0651-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-016-0651-6