
Abstract
Background
Parasites from the genus Plasmodium, the aetiological agent of malaria in humans, can also infect non-human primates (NHP), increasing the potential risk of zoonotic transmission with its associated global public health concerns. In Colombia, there are no recent studies on Plasmodium spp. infecting free-ranging NHP. Thus, this study aimed to determine the diversity of Plasmodium species circulating in fragmented forests in central Colombia, both in Anopheles mosquitoes and in the four sympatric NHP in the region (Ateles hybridus, Cebus versicolor, Alouatta seniculus and Aotus griseimembra), in order to evaluate the risk of infection to humans associated with the presence of sylvatic hosts and vectors infected with Plasmodium spp.
Methods
Overall, there were collected 166 fecal samples and 25 blood samples from NHP, and 442 individuals of Anopheles spp. DNA extraction, nested PCR using mitochondrial (cox3 gene) and ribosomal (18S rDNA) primers, electrophoresis and sequencing were conducted in order to identify Plasmodium spp. from the samples.
Results
Plasmodium falciparum was detected in two fecal samples of Alouatta seniculus, while Plasmodium vivax/simium infected Ateles hybridus, Cebus versicolor and Alouatta seniculus. Co-infections with P. vivax/simium and Plasmodium malariae/brasilianum were found in three individuals. The highest prevalence from blood samples was found for Plasmodium malariae/brasilianum in two Alouatta seniculus while Plasmodium vivax/simium was most prevalent in fecal samples, infecting four individuals of Alouatta seniculus. Seven Anopheles species were identified in the study site: Anopheles (Anopheles) punctimacula, Anopheles (An.) malefactor, Anopheles (Nyssorhynchus) oswaldoi, Anopheles (Nys.) triannulatus, Anopheles (An.) neomaculipalpus, Anopheles (Nys.) braziliensis and Anopheles (Nys.) nuneztovari. Infection with P. vivax/simium was found in An. nuneztovari, An. neomaculipalpus, and An. triannulatus. Furthermore, An. oswaldoi and An. triannulatus were found infected with P. malariae/brasilianum. The effect of fragmentation and distance to the nearest town measured in five forests with different degrees of fragmentation was not statistically significant on the prevalence of Plasmodium in NHP, but forest fragmentation did have an effect on the Minimum Infection Rate (MIR) in Anopheles mosquitoes.
Conclusions
The presence of Plasmodium spp. in NHP and Anopheles spp. in fragmented forests in Colombia has important epidemiological implications in the human–NHP interface and the associated risk of malaria transmission.
Similar content being viewed by others

Background
Parasites belonging to the genus Plasmodium are among the best studied parasites in the world, since they are responsible of causing malaria, the deadliest vector borne disease [1]. Although malaria was targeted for elimination in 2030 in at least 35 countries, and the reduction of its incidence from 2005 until 2014 suggested this goal was achievable, in 2016 nine Latin American countries reported an increase in its incidence [2]. In Colombia, the number of cases increased in 2016, but most importantly, Plasmodium falciparum, the parasite responsible for cerebral malaria had an increase in its incidence, becoming more prevalent than Plasmodium vivax, the predominant species in the country until 2013 [2]. In 2018, the National System of Public Health Surveillance (SIVIGILA by its Spanish acronym) reported 61,339 cases of malaria in Colombia [3], P. vivax being the most prevalent (50%), followed by P. falciparum (48%), and mixed infection P. vivax and P. falciparum (2%).
Changes in malaria eco-epidemiology can be related to shifts in sylvatic transmission cycles, since Plasmodium parasites are also present in non-human primates (NHP) in tropical regions around the world [4]. Until now, 26 Plasmodium species have been formally described in NHP, each found infecting from one to 29 species [4].
In Latin America, NHP are potential reservoirs of Plasmodium brasilianum/Plasmodium malariae [5, 6], and some species have been found infected with P. falciparum [7] or with Plasmodium simium [5, 8], which is closely related to P. vivax [9, 10]. Although the risk of malaria zoonotic infection is of public health concern [11], it still remains largely understudied.
Several studies were carried out between 1930 and 1985 in Brazil, Panama, Venezuela, Peru and Colombia, and blood samples were obtained from the genera Alouatta, Aotus, Cacajao, Callicebus, Callithrix, Brachyteles, Chiropotes, Lagothrix, Pithecia, Saimiri, Saguinus, Cebus, Callicebus and Ateles [5, 8, 12,13,14,15,16,17,18,19,20,21,22,23]. They reported P. simium infecting Alouatta guariba and Brachyteles arachnoides [5, 8], and P. brasilianum infecting Callicebus brunneus, Chiropotes satanas, Saguinus midas, Phitecia monachus, Lagothrix cana, Cebus capucinus, Saimiri sciureus, Saimiri boliviensis, Ateles geoffroyi and Alouatta palliata [8, 16,17,18]. Most recent studies have been conducted in Brazil [24,25,26,27,28,29] and few others in Costa Rica [30] and Venezuela [6]. In Colombia, recent reports of Plasmodium infection on free-ranging primates are not available, and the last studies were conducted between 1952 and 1968. These early studies found evidence of P. brasilianum infecting NHP, including Ateles geoffroyi, Cebus albifrons, Cebus apella, Cebus capucinus, Lagothrix lagotricha and Saimiri sciureus [15, 17, 21].
The zoonotic risk of transmission between humans and NHP involves the presence of insect vectors feeding on both hosts; thus, mosquito feeding behaviour can influence Plasmodium transmission between humans and NHP [31]. In Africa, Makanga et al. documented that certain sylvatic mosquitoes infected with ape parasites also bite humans, being potential bridge vectors between humans and apes [32].
In Colombia, the most important malaria vectors are Anopheles (Nyssorhynchus) albimanus, Anopheles (Nys.) darlingi and Anopheles (Nys.) nuneztovari [33]. Studies on Anopheles mosquitoes in the country have been mainly focused in urban transmission cycles. However, in a context of pervasive transformation of natural areas into agricultural fields and extensive cattle ranches, it is essential to identify malaria vectors in forested areas, as these species may transmit Plasmodium from NHP to humans or vice versa [31].
In a socio-ecological context, the increasing human population coupled with greater demand for agricultural land, has led to an incremental deforestation in tropical countries where malaria is endemic [34]. It has been reported that deforestation and land use changes greatly influence malaria’s incidence [35]. In Colombia, the Middle Magdalena River valley is a region where the natural forest has been reduced to less than 15% of the original coverage due to deforestation and land use change [36]. This pervasive process might increase the contact between humans and NHP as well as affect parasite-host dynamics [37].
In this context, this study aimed to identify the prevalence of Plasmodium in five fragmented forest patches in Colombia in order to infer a potential risk of malaria zoonotic transmission involving NHP. The risk can exist if infected NHP species and infected Anopheles species are present in the study sites. To achieve this goal, the main objectives were: (i) to determine the presence and infection rate of Plasmodium parasites in NHP and Anopheles, (ii) to establish if infection rates vary among infected Anopheles species, and (iii) to evaluate if sites with different degrees of habitat transformation exhibit variation in infection rates.
Methods
Study sites
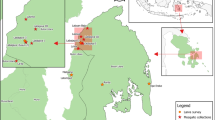
Fieldwork was performed in five forest fragments (San Juan, Lucitania, Rompederos, El Silencio, and Quinchas) located in the Middle Magdalena River valley in Santander, Antioquia and Boyaca Departments, in Colombia (Table 1 and Fig. 1a). In the Department of Santander malaria transmission occurs mainly in two municipalities, Cimitarra and Puerto Wilches. In 2018, P. vivax was the dominant species [38] with 22 cases, while seven were notified of P. falciparum and one co-infection [3]. In the Department of Antioquia in 2018, P. vivax was detected in 4360 of the notified cases, P. falciparum in 975 and 80 co-infections [3]. In the Department of Boyacá fewer cases are known, with only four records in 2017 in the municipality of Puerto Boyacá [39], and none in 2018 [40].

a Study sites. b Number of NHP samples per species and study site. c Plasmodium infection presence in Anopheles and NHP samples (feces and blood) per study site. d Anopheles species collected per study site
Non-human primate samples
Between 2016 and 2018, primates were followed from dawn to dusk and 166 fecal samples were collected from the soil immediately after defecation, and placed in eppendorf tubes with RNAlater solution. Fecal samples from brown spider monkeys (Ateles hybridus), capuchin monkeys (Cebus versicolor), red howler monkeys (Alouatta seniculus) and grey-legged night monkeys (Aotus griseimembra) were collected (Table 2 and Fig. 1b). Additionally, in San Juan, blood samples were collected from 25 primates (Table 2) anesthetized using darts with zolazepam hydrochloride (Zoletil), with specific doses for each species. Once sedated, blood samples were collected by caudal vein puncture, and placed in Vacutainer tubes containing sodium citrate. Plasma and red blood cells were separated by centrifugation and stored in liquid nitrogen tank (Thermo Scientific) until transported to the laboratory. For 16 specimens (three Alouatta seniculus, six Aotus griseimembra, two Ateles hybridus and five Cebus versicolor) blood and fecal samples were obtained from the same individuals in the same field campaign.
Mosquito collection and identification
Adult mosquitoes were sampled in three study sites (Rompederos, San Juan, and Lucitania) during three consecutive nights in each site; due to transport and accessibility restrictions, the remaining two sites were not sampled for mosquitoes (Quinchas and El Silencio). One Shannon trap [41, 42], three BG-Sentinel traps (BioGents, Regensburg, Germany) [43] baited with Octenol and six CDC light-traps [44, 45] (three on the canopy and three on the understorey) were set, close to places frequented by primates.
Mosquitoes were sorted immediately after capture and female Anopheles were preserved in RNAlater buffer. Some females were kept dry for morphologic identification using the keys of González and Carrejo [46] and Forattini [47]. Species identity was confirmed through DNA barcoding with amplification of the 658 bp region from the COI gene [48, 49].
Molecular analyses
DNA from fecal samples was extracted using a ZR fecal DNA MiniPrep Kit (Zymo), according to the manufacturer's protocol. DNA extraction from NHP blood samples was performed individually using High Pure PCR Template Preparation Kit (Roche). Female mosquitoes were pooled by species and study site, and DNA was extracted from pools with up to ten individuals using the ZR Tissue & Insect DNA MiniPrep Kit (Zymo).
To detect P. falciparum, P. vivax/simium and P. malariae/brasilianum, nested PCR using mitochondrial primers (cox3 gene) and nested PCR using ribosomal primers (18S rDNA) were performed in all samples following published methods [50, 51]. All PCR products from second reactions were visualized on an agarose gel and positive samples were sequenced by Big Dye Terminator v3.0 Cycle Sequencing Kit using the ABI-3500 Genetic Analyzer (Life Technologies) for species identity confirmation. Sequences were edited using Geneious Software and compared by BLAST (Basic Local Alignment Search Tool) [52] with publicly available sequences in GenBank (National Center for Biotechnology Information).
Data analyses
The prevalence (infection rate) of Plasmodium spp. in each NHP species and study site was calculated as # infected individuals/total examined individuals * 100. The minimum infection rate (MIR) in each Anopheles species was calculated as the number of positive pools divided by the total of tested specimens, assuming that each positive pool contains at least one infected individual [49].
A General Linear Model (Family: quasibinomial) was performed using the RStudio integrated development environment, in order to evaluate the effect of fragmentation and distance to the nearest town on Plasmodium prevalence in NHP and the MIR. Forest fragment size (ha) was used as a proxy for fragmentation intensity [53].
Results
Non-human primate samples
Plasmodium spp. infection was detected in 20 NHP samples. Plasmodium falciparum was found infecting Alouatta seniculus (n = 2), while P. vivax/simium infected Ateles hybridus (n = 5), Cebus versicolor (n = 2) and Alouatta seniculus (n = 5). Additionally, P. malariae/brasilianum was found infecting the four NHP sampled species (Table 3 and Fig. 1c). Co-infections with P. vivax/simium and P. malariae/brasilianum were found in two individuals of Ateles hybridus in Lucitania and one individual of Alouatta seniculus in San Juan. For those specimens with paired fecal and blood samples, consensus in infection results was not obtained since five individuals were only positive for fecal samples and seven for blood samples.
The higher Plasmodium prevalence was obtained with P. malariae/brasilianum in Alouatta seniculus blood samples and Ateles hybridus fecal samples (Table 3).
Plasmodium prevalence in NHP was not related to fragmentation (General linear model: fecal samples Pr = 0.415, blood samples Pr = 0.272) or distance to nearest town (General linear model: fecal samples Pr = 0.272, blood samples Pr = 0.272).
Mosquito collection
In total, 442 mosquitoes were collected, but due to loss of important taxonomic characters during processing and transport, 26.6% of them could not be identified and were not processed for parasite infection. The remaining 324 mosquitoes were collected mainly in Rompederos (51.8%), followed by San Juan (36.7%) and Lucitania (10.8%).
In this study, the Shannon trap was the most efficient capture method with 430 Anopheles. CDC light-traps caught 12 Anopheles (only one from the canopy) while BG-Sentinel traps did not capture any Anopheles. Seven Anopheles species were identified based on diagnostic morphological characters: Anopheles (Anopheles) punctimacula, Anopheles (An.) malefactor, Anopheles (Nys.) oswaldoi, Anopheles (Nys.) triannulatus, Anopheles (An.) neomaculipalpus, Anopheles (Nys.) braziliensis and Anopheles (Nys.) nuneztovari (Fig. 1d). The species identity of An. punctimacula, An. malefactor, An. oswaldoi, An. triannulatus, and An. neomaculipalpus was confirmed by DNA barcoding, obtaining 99–100% of identity with the reference sequences in GenBank. The remaining two species An. braziliensis and An. nuneztovari did not have successful amplification of the COI gene, and the obtained sequences could not be validated, but morphologic identification was successfully performed based on diagnostic characters. Anopheles triannulatus was found in all three sampling sites, and An. oswaldoi and An. neomaculipalpus were the most abundant species (Table 4).
Infection with P. vivax/simium was found in An. nuneztovari, An. neomaculipalpus, and An. triannulatus. Furthermore, An. oswaldoi and An. triannulatus were found infected with P. malariae/brasilianum (Table 4). The MIR was related to fragmentation (General linear model: Pr = 0.002), but not to proximity to the nearest town (General linear model: Pr = 0.056).
Discussion
In this study, three Plasmodium species were found infecting NHP in the Magdalena River valley. As expected, the most prevalent parasite species was P. malariae/P. brasilianum that naturally infects different NHP species in Central and South America [6, 25, 30]. Interestingly, P. brasilianum has been reported infecting humans living in close proximity with NHP in the Venezuelan Amazon [6], which highlights the risk of parasite transmission from NHP to humans.
The finding of P. vivax/P. simium in NHP is of great interest given that P. vivax has been the most prevalent species historically recorded in the country. In 2017, after analysing 28 human blood samples from an outbreak in the Atlantic Forest coastal region of Brazil, P. simium was detected infecting humans [24]. Authors suggested that this species could be circulating in humans before, but was misdiagnosed as P. vivax due to the absence of adequate diagnostic techniques to perform species identification. Additionally, Grigg and Snounou (2017) consider Brazilian monkeys as reservoirs for P. vivax [54]. The presence of sylvatic reservoirs is relevant as it can potentially threaten successful malaria elimination campaigns [54]. In the Colombian context the presence of infected monkeys should be taken into account when elucidating the potential risk of human infection.
In our study, the unexpected presence of P. falciparum infecting Alouatta seniculus open very relevant questions and concerns. Although this species has been found in New World primates [7, 55] it is not as prevalent as P. brasilianum. The presence of P. falciparum in NHP suggests parasite transmission from humans to NHP, which is relevant when evaluating human contribution to emerging infectious diseases in sylvatic NHP. Other vector borne diseases transmitted from humans to primates such as Yellow Fever in South America, have negatively impacted populations of Alouatta guariba clamitans and Alouatta caraya in Argentina and Brazil, reassessing their conservation status to Critically Endangered and Near Threatened respectively [56]. Regarding the establishment of P. falciparum in the wild, Araújo et al. considered that, due to the ability of the parasite to develop in NHP, mosquito infection from infected NHP is likely and so is the establishment of a sylvatic transmission cycle [7].
Although infection with Plasmodium species has been reported in the genera Alouatta, Cebus, Aotus and Ateles [7, 8, 17], results obtained in this study constitute new records at the species level for Cebus versicolor, Ateles hybridus and Aotus griseimembra.
Fecal samples have been used in different studies mainly in Africa and Asia [57, 58] for Plasmodium detection as a simple, non-invasive and inexpensive alternative to blood samples. However, they degrade quickly [58] and the presence of bacteria and polysaccharides from plant diet, which are potential inhibitors of PCR, makes it difficult to use these samples for diagnosis by PCR [59]. To solve this inconveniences, there was used RNAlater for sample preservation and BSA in the PCR mix in order to stabilize the DNA [60]. Since Plasmodium detection from blood is more sensitive than from fecal samples [57, 61], prevalence rates found in this study may be underestimated, given that blood samples could not be obtained for all sampled primates. Also, those differences in detection sensitivity according to the type of sample could partially explain the lack of coincidence in the obtained results for the tested paired fecal and blood samples. For future studies, in order to confirm Plasmodium species circulating in zoonotic cycles, whole genome sequencing should be performed. In this way, it is possible to determine if Plasmodium infections are caused by P. simium or P. vivax, and P. malariae or P. brasilianum.
Regarding mosquito collections, all the species found in this study were known records for the study sites: An. nuneztovari, An. triannulatus, An. neomaculipalpus and An. oswaldoi have been previously reported in Santander Department [62, 63], and An. punctimacula, An. neomaculipalpus and An. triannulatus in Antioquia Department [33], as well as An. oswaldoi [64].
Plasmodium vivax was found infecting three species, An. neomaculipalpus, An. triannulatus and An. nuneztovari, which is concordant with previous records [65, 66]. Anopheles neomaculipalpus is known to be highly anthropophilic [65] while An. triannulatus has been collected resting on cattle and is known to colonize transformed environments and become very abundant [67]. Lucitania, Rompederos and San Juan exhibit the presence of cattle and altered ecosystems mainly due to the progressive introduction of oil palm plantations. It has been reported that deforestation related to monoculture favours the presence of ponds which are frequent breeding sites of An. nuneztovari [68], a species recognized as primary malaria vector in Colombia [69].
Infection with P. malariae was found in An. triannulatus and An. oswaldoi as has also been previously recorded [70, 71]. Anopheles triannulatus has been reported with zoophilic and anthropophilic activities [33] and An. oswaldoi has been incriminated as secondary vector in the country [69]. Anopheles triannulatus and An. oswaldoi were found in this study, supporting previous reports and confirming their presence in Santander and Antioquia Departments.
The most abundant species were An. oswaldoi and An. neomaculipalpus found infected with P. vivax and P. malariae, respectively; this suggests their potential role as vectors in the study sites. Rompederos and Lucitania were the localities with highest MIR. Anopheles vectors and NHP exhibit different selection strategies; while the lifespan of NHP is long, probably supporting a long course of infection, mosquitoes have a short lifespan and marked fluctuation in population densities related to environmental variables [72, 73]. Possibly the sampling time of this study coincided with low Anopheles local abundances thus the number of collected individuals was below the detection threshold. Long-term studies including seasonal variation would allow a better understanding of hosts’ population dynamics and the parasite transmission system in the studied environment.
Regarding the diversity of Anopheles species it was higher in Rompederos and San Juan, compared to Lucitania. This could be explained partially by the fact that those fragments belong to flooded forests which could provide optimal mosquito breeding sites, while Lucitania is a terra firme fragment forest.
The analyses on the effect of habitat fragmentation and distance to nearest town over the prevalence of Plasmodium in NHP didn’t show any significant association, while the fragmentation but not the distance to nearest town had an effect over the MIR. However, when evaluating malaria risk, it is important to consider that multiple factors and processes interact e.g. the environment (land cover use/change, landscape transformations), human populations (host susceptibility, movement patterns, forest-related activity), vector biology (vector activity and life cycle, mosquito species distribution) [74]. This study only focused in some of those factors (e.g.: Plasmodium infection rates), but the complexity of the malaria cycle is a fact that must be taken into account, and for further studies it is suggested to involve as many factors as possible.
Conclusions
The results of this study provide evidence for a potential risk of zoonotic malaria transmission in terms of Plasmodium species infecting NHP and Anopheles, which can have negative effect on both human and NHP populations. Fragmentation and proximity to the nearest town did not show a statistically significant effect on the prevalence of Plasmodium in NHP, while fragmentation had an effect over the MIR.
Availability of data and materials
All data generated during this study are included in this published article.
Abbreviations
- BLAST:
-
Basic Local Alignment Search Tool
- COI:
-
cytochrome c oxidase subunit I
- DNA:
-
deoxyribonucleic acid
- MIR:
-
minimum infection rate
- NHP:
-
non-human primates
- PCR:
-
polymerase chain reaction
References
WHO. Vector Borne Diseases. Geneva: World Health Organization; 2018. https://www.who.int/heli/risks/vectors/vector/en/. Accessed 11 Mar 2019.
PAHO. Actualización Epidemiológica. Aumento de malaria en las Américas. Pan American Health Organization. 2018. http://www.paho.org/hq/index.php?option=com_docman&task=doc_view&Itemid=270&gid=43437&lang=es. Accessed 11 Mar 2019.
Instituto Nacional de Salud. Boletín Epidemiológico Semanal. 2018. https://www.ins.gov.co/buscador-eventos/BoletinEpidemiologico/2018Boletínepidemiológicosemana52.pdf. Accessed 25 June 2019.
Faust C, Dobson AP. Primate malarias: diversity, distribution and insights for zoonotic Plasmodium. One Health. 2015;1:66–75.
Deane L, Ferreira Neto J, Sitonio J. Novo hospedeiro natural do Plasmodium simium e do Plasmodium brasilianum: o mono, Brachyteles arachnoides. Rev Inst Med Trop Sao Paulo. 1968;10:287–8.
Lalremruata A, Magris M, Vivas-Martínez S, Koehler M, Esen M, Kempaiah P, et al. Natural infection of Plasmodium brasilianum in humans: man and monkey share quartan malaria parasites in the Venezuelan Amazon. EBioMedicine. 2015;2:1186–92.
Araújo MS, Messias MR, Figueiró MR, Gil LH, Probst CM, Vidal N, et al. Natural Plasmodium infection in monkeys in the state of Rondônia (Brazilian Western Amazon). Malar J. 2013;12:180.
Deane L, Ferreira Neto J, Okumura M, Ferreira M. Malaria parasites of Brazilian monkeys. Rev Inst Med Trop Sao Paulo. 1969;11:71–86.
Escalante A, Barrio E, Ayala F. Evolutionary origin of human and primate malarias: evidence from the circumsporozoite protein gene. Mol Biol Evol. 1995;12:616–26.
Camargos D, Pereira G, De Souza F, Araújo F, De Souza J, Braga Z, et al. Plasmodium simium, a Plasmodium vivax-related malaria parasite: genetic variability of Duffy binding protein II and the Duffy antigen/receptor for chemokines. PLoS One. 2015;10:e0131339.
Baird J. Malaria zoonoses. Travel Med Infect Dis. 2009;7:269–77.
Garnham P. Malaria parasites and other haemosporidia. Oxford: Blackwell Scientific Publications; 1966.
Porter J, Johnson C, De Sousa L. Prevalence of malaria in Panamanian primates. J Parasit. 1966;52:669–70.
Serrano J. Infección natural de un araguato, Alouatta seniculus straminea, por Plasmodium brasilianum en Venezuela [Natural infection of a monkey, Alouatta seniculus straminea, by Plasmodium brasilianum in Venezuela]. Acta Cient Venez. 1967;18:13–5.
Renjifo S, Sanmartín C, De Zulueta J. A survey of the blood parasites of vertebrates in Eastern Columbia. Acta Trop. 1952;9:151–69.
De Arruda M. Presença do Plasmodium brasilianum em macacos capturados na área de enchimento do reservatório da Usina Hidrelética de Tucuruí, Pará. Mem Inst Oswaldo Cruz. 1985;80:367–9.
Marinkelle J, Grose E. Plasmodium brasilianum in Colombian monkeys. Trop Geogr Med. 1968;20:276–80.
Clark H. Progress in the survey for blood parasites of the wild monkeys of Panama. Am J Trop Med. 1931;11:11–20.
Clark H. A preliminary report on some parasites in the blood of wild monkey in Panama. Am J Trop Med Hyg. 1930;10:25–41.
Deane L. Epidemiology of simian malaria in the American continent. Pan Am Health Organ. 1976;317:144–63.
Dunn FL, Lambrecht F. The hosts of Plasmodium brasilianum Gonder and von Berenberg-Gossler, 1908. J Parasit. 1963;49:316–9.
Fonseca F. Plasmódio de primata do Brasil. Mem Inst Oswaldo Cruz. 1951;49:543–51.
Garnham P. Parasites in South American monkeys. Trans R Soc Trop Med Hyg. 1960;54:1–2.
Brasil P, Zalis M, Pina-costa A, Machado A, Júnior C, Silva S, et al. Outbreak of human malaria caused by Plasmodium simium in the Atlantic Forest in Rio de Janeiro: a molecular epidemiological investigation. Lancet Glob Health. 2017;5:E1038–46.
Bueno M, Rohe F, Kirchgatter K, di Santi S, Guimaraes L, Witte C, et al. Survey of Plasmodium spp. in free-ranging neotropical primates from the Brazilian Amazon Region impacted by anthropogenic actions. Ecohealth. 2013;10:48–53.
de Alvarenga D, Culleton R, Pina-Costa A, Fonseca D, Junior C, Silva S, et al. An assay for the identification of Plasmodium simium infection for diagnosis of zoonotic malaria in the Brazilian Atlantic Forest. Sci Rep. 2018;8:86.
de Alvarenga D, Pina-costa A, De Sousa T, Pissinatti A, Zalis M, Suárez M, et al. Simian malaria in the Brazilian Atlantic forest: first description of natural infection of capuchin monkeys (Cebinae subfamily) by Plasmodium simium. Malar J. 2015;14:81.
Figueiredo M, di Santi S, Gómez W, André M, Zacarias R. Identification of Plasmodium spp in Neotropical primates of Maranhense Amazon in Northeast Brazil. PLoS One. 2017;12:0182905.
Figueiredo M, Di Santi S, Figueiredo T, Machado R. Natural Plasmodium infection in neotropical primates in the island of São Luís, state of Maranhão, Brazil. Braz J Vet Parasitol. 2015;24:122–8.
Fuentes-Ramírez A, Jiménez-Soto M, Castro R, Romero-Zuñiga J, Dolz G. Molecular detection of Plasmodium malariae/Plasmodium brasilianum in non-human primates in captivity in Costa Rica. PLoS One. 2017;12:e0170704.
Verhulst N, Smallegange R, Takken W. Mosquitoes as potential bridge vectors of malaria parasites from non-human primates to humans. Front Physiol. 2012;3:197.
Makanga B, Yangari P, Rahola N, Rougeron V, Elguero E, Boundenga L, et al. Ape malaria transmission and potential for ape-to-human transfers in Africa. Proc Natl Acad Sci USA. 2016;113:5329–34.
Gutiérrez L, González J, Gómez G, Castro M, Rosero D, Luckhart S, et al. Species composition and natural infectivity of anthropophilic Anopheles (Diptera: Culicidae) in Córdoba and Antioquia states in northwestern Colombia. Mem Inst Oswaldo Cruz. 2009;104:1117–24.
Ramasamy R. Zoonotic malaria—global overview and research and policy needs. Front Public Health. 2014;2:123.
Yasuoka J, Levins R. Impact of deforestation and agricultural development on anopheline ecology and malaria epidemiology. Am J Trop Med Hyg. 2007;76:450–60.
Roncancio-Duque N, Benavides J. Parásitos intestinales en poblaciones pequeñas y aisladas de mono aullador rojo (Alouatta seniculus) y mono araña café (Ateles hybridus), Atelidae—primates en el Magdalena Medio, Colombia. Vet y Zootecnia. 2013;7:71–89.
Trejo-macías G, Estrada A, Ángel M, Cabrera M. Survey of Helminth Parasites in Populations of Alouatta palliata mexicana and A. pigra in continuous and in fragmented habitat in southern Mexico. Int J Primatol. 2007;28:931–45.
Gobernación de Santander. Boletín Epidemiológico Semanal. 2018. http://www.santander.gov.co/index.php/gobernacion/documentacion/send/938-boletines/16135-boletin-epidemiologico-semanal-no-50-de-2018. Accessed 11 Mar 2019.
Gobernación de Boyacá. Boletín Epidemiológico de Boyacá. 2017. http://www.boyaca.gov.co/SecSalud/component/easyfolderlistingpro/?view=download&format=raw&data=eNpVkGFugzAMhe_iCxAY65D5N63niAwYaikhCAepU9W7lzSgab9sP-t9enqEpcGH4gVhDG7gFVrFukIQTxNr8RP6zfMcgxbfwXGUmfW6yMBegguT9EGToywRNuU1M7KEYM7XtLHGP_4XgrVvLV1Nts7kOZ0GIY1LVmWAVtBk08puoXhL70-EojJlc1wwRnF8AuoaoctZLf-LarvwSz1ZZU8zWVPZE_GBwPd4bMswHin5vsjKekbYqwKKkfpbqgTablefL80aaI8. Accessed 11 Mar 2019.
Gobernación de Boyacá. Boletín Epidemiológico de Boyacá. 2018. http://www.boyaca.gov.co/SecSalud/component/easyfolderlistingpro/?view=download&format=raw&data=eNpVkGFugzAMhe_iCxAY65D5N63niAwYaikhCAepU9W7lzSgab9sP-t9enqEpcGH4gVhDG7gFVrFukIQTxNr8RP6zfMcgxbfwXGUmfW6yMBegguT9EGToywRNuU1M7KEYM7XtLHGP_4XgrVvLV1Nts7kOZ0GIY1. Accessed 11 Mar 2019.
Shannon R. Methods for collecting and feeding mosquitoes in jungle yellow fever studies. Am J Trop Med Hyg. 1939;19:131–40.
Neves A, Urbinatti P, Malafronte R, Fernandes A, Da Silva W, Natal D. Malaria outside the Amazon region: natural Plasmodium infection in anophelines collected near an indigenous village in the Vale do Rio Branco, Itanhaém, SP, Brazil. Acta Trop. 2013;125:102–6.
Schmied W, Takken W, Killeen G, Knols B, Smallegange R. Evaluation of two counterflow traps for testing behaviour-mediating compounds for the malaria vector Anopheles gambiae s.s. under semi-field conditions in Tanzania. Malar J. 2008;7:230.
Sudia W, Chamberlain R. Battery-operated light trap, an improved model. Mosq News. 1962;22:126–9.
Makanga B, Costantini C, Rahola N, Yangari P, Rougeron V, Ayala D, et al. “Show me which parasites you carry and I will tell you what you eat”, or how to infer the trophic behavior of hematophagous arthropods feeding on wildlife. Ecol Evol. 2017;7:7578–84.
González R, Carrejo N. Introducción al estudio taxonómico de Anopheles de Colombia: claves y notas de distribución. 2nd ed. Cali: Universidad del Valle; 2009.
Forattini O. Entomología Médica. Culicini: Culex, Aedes e Psorophora. São Paulo, Brasil: Faculdade de Higiene e Saúde Pública, Depto. de Parasitologia; 1965.
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotechnol. 1994;3:294–9.
González C, Molina A, León C, Salcedo N, Rondón S, Paz A, et al. Entomological characterization of malaria in northern Colombia through vector and parasite species identification, and analyses of spatial distribution and infection rates. Malar J. 2017;16:431.
Snounou G, Viriyakosol S, Zhu XP, Jarra W, Pinheiro L, Rosario V, et al. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol. 1993;61:315–20.
Isozumi R, Fukui M, Kaneko A, Chan C, Kawamoto F, Kimura M. Improved detection of malaria cases in island settings of Vanuatu and Kenya by PCR that targets the Plasmodium mitochondrial cytochrome c oxidase III (cox3) gene. Parasitol Int. 2015;64:304–8.
Altschul S, Madden T, Schäffer A, Zhang J, Zhang Z, Miller W, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–402.
Rimbach R, Link A, Heistermann M, Gómez-Posada C, Galvis N, Heymann E. Effects of logging, hunting, and forest fragment size on physiological stress levels of two sympatric ateline primates in Colombia. Conserv Physiol. 2013;1:1–11.
Grigg M, Snounou G. Plasmodium simium: a Brazilian focus of anthropozoonotic vivax malaria? Lancet Glob Health. 2017;5:E961–2.
Duarte A, Malafronte R, Cerutti C, Curado I, de Paiva B, Maeda A, et al. Natural Plasmodium infections in Brazilian wild monkeys: reservoirs for human infection? Acta Trop. 2008;107:179–85.
Bicca-Marques J, Calegaro-Marques C. Parasite sharing between humans and nonhuman primates and the hidden dangers to primate conservation. Zoologia. 2014;31:313–5.
Liu W, Li Y, Learn G, Rudicell R, Robertson J, Keele B, et al. Origin of the human malaria parasite Plasmodium falciparum in gorillas. Nature. 2010;467:420–5.
Siregar J, Faust C, Murdiyarso L, Rosmanah L, Saepuloh U, Dobson A, et al. Non-invasive surveillance for Plasmodium in reservoir macaque species. Malar J. 2015;14:404.
Monteiro L, Bonnemaison D, Vekris A, Petry K, Bonnet J, Vidal R, et al. Complex polysaccharides as PCR inhibitors in feces: Helicobacter pylori model. J Clin Microbiol. 1997;35:995–8.
Assis G, De Alvarenga D, Camargos D, de Souza J, Braga Z, Satiko F, et al. Detection of Plasmodium in faeces of the New World primate Alouatta clamitans. Mem Inst Oswaldo Cruz. 2016;111:570–6.
Nwakanma D, Gomez-Escobar N, Walther M, Crozier S, Dubovsky F, Malkin E, et al. Quantitative detection of Plasmodium falciparum DNA in saliva, blood, and urine. J Infect Dis. 2009;199:1567–74.
Parra-Henao G, Delgado D, Alarcón E. Especies de Anopheles en el Canón del Río Sogamoso, Santander, Colombia, al inicio de la puesta en marcha del proyecto hidroeléctrico. Boletín Malariol y Salud Ambient. 2012;2:287–94.
Brochero H, Pareja P, Ortiz G, Olano V. Sitios de cría y actividad de picadura de especies de Anopheles en el municipio de Cimitarra, Santander, Colombia. Biomédica. 2006;26:269–77.
Ruiz-Lopez F, Wilkerson R, Ponsonby D, Herrera M, Mureb M, Velez I, et al. Systematics of the Oswaldoi Complex (Anopheles, Nyssorhynchus) in South America. Parasit Vectors. 2013;6:324.
Moreno J, Rubio-Palis Y, Páez E, Pérez E, Sánchez V, Vaccari E. Anopheles (Anopheles) neomaculipalpus: a new malaria vector in the Amazon basin? Med Vet Entomol. 2005;19:329–32.
Naranjo-Diaz N, Rosero D, Rua-Uribe G, Luckhart S, Correa M. Abundance, behavior and entomological inoculation rates of anthropophilic anophelines from a primary Colombian malaria endemic area. Parasit Vectors. 2013;6:61.
Rosero D, Naranjo-Diaz N, Alvarez N, Cienfuegos A, Torres C, Luckhart S, et al. Colombian Anopheles triannulatus (Diptera: Culicidae) naturally infected with Plasmodium spp. ISRN Parasitol. 2013;2013:927453.
Montoya-Lerma J, Solarte Y, Giraldo-Calderón G, Quiñones M, Ruiz-López F, Wilkerson R, et al. Malaria vector species in Colombia—a review. Mem Inst Oswaldo Cruz. 2011;106:223–38.
Montoya C, Bascuñán P, Rodríguez-Zabala J, Correa M. Abundance, composition and natural infection of Anopheles mosquitoes from two malaria-endemic regions of Colombia. Biomédica. 2017;37:98–105.
Tadei W, Thatcher D. Malaria vectors in the Brazilian Amazon: Anopheles of the subgenus Nyssorhynchus. Rev Inst Med Trop Sao Paulo. 2000;42:87–94.
Branquinho M, Taipe C, Rocha R, Natal D, Barata J, Cochrane A, et al. Anophelines in the state of Acre, Brazil, infected with Plasmodium falciparum, P. vivax, the variant P. vivax VK247 and P. malariae. Trans R Soc Trop Med Hyg. 1993;87:391–4.
Bashar K, Tuno N. Seasonal abundance of Anopheles mosquitoes and their association with meteorological factors and malaria incidence in Bangladesh. Parasit Vectors. 2014;7:442.
Rossati A, Bargiacchi O, Kroumova V, Zaramella M, Caputo A, Garavelli P. Climate, environment and transmission of malaria. Infez Med. 2016;24:93–104.
Tucker J, Vittor A, Rifai S, Valle D. Does deforestation promote or inhibit malaria transmission in the Amazon? A systematic literature review and critical appraisal of current evidence. Philos Trans R Soc B Biol Sci. 2017;372:20160125.
Acknowledgements
We thank the Centers for Disease Control and Prevention for providing financial support, and Centro de Investigaciones en Microbiología y Parasitología Tropical (CIMPAT) at Universidad de los Andes as well as Fundación Proyecto Primates for providing all logistic support.
Funding
This research was partly funded by the Office of Infectious Disease, Bureau for Global Health, U.S. Agency for International Development, under the terms of an Interagency Agreement with Centers for Disease Control and Prevention (CDC). Additionally, SR was awarded a full scholarship from the Department of Biological Sciences, Faculty of Sciences, Uniandes.
Author information
Authors and Affiliations
Contributions
SR and CG designed the study, SR and AL conducted primate and entomological sampling, SR and CL performed molecular biology procedures. SR, CG, AL drafted and edited the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval for the collection of fecal and blood samples as well as the collection of vectors were obtained by Universidad de los Andes, the National Environmental Licensing Authority of Colombia (ANLA) and the Centers for Disease Control and Prevention. Permits Nº: 2017025578-1-000, 2017043863-1-000, 2017065795-1-000, 2017013727-1-000, 2017052943-1-000, 2017081458-1-000, 2017108650-1-000.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Rondón, S., León, C., Link, A. et al. Prevalence of Plasmodium parasites in non-human primates and mosquitoes in areas with different degrees of fragmentation in Colombia. Malar J 18, 276 (2019). https://doi.org/10.1186/s12936-019-2910-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-019-2910-z