
Abstract
Background
The wide spread mosquito Culex pipiens pipiens have two forms molestus and pipiens which frequently hybridize. The two forms have behavioural and physiological differences affecting habitat requirements and host selection, which may affect the transmission dynamic of Cx. p. pipiens-borne diseases.
Methods
During 2013, blood engorged Cx. p. pipiens mosquitoes were captured in urban, rural and natural areas from Southern Spain. In 120 mosquitoes, we identified the blood meal origin at vertebrate species/genus level and the mosquito form. The presence and molecular lineage identity of avian malaria parasites in the head-thorax of each mosquito was also analysed.
Results
Mosquitoes of the form pipiens were more frequently found in natural than in urban areas. The proportion of Cx. pipiens form molestus and hybrids of the two forms did not differ between habitat categories. Any significant difference in the proportion of blood meals on birds between forms was found. Birds were the most common feeding source for the two forms and their hybrids. Among mammals, dogs and humans were the most common hosts. Two Plasmodium and one Haemoproteus lineages were found in mosquitoes, with non-significant differences between forms.
Conclusion
This study supports a differential distribution of Cx. p. pipiens form pipiens between urban and natural areas. Probably due to the similar feeding sources of both mosquito forms and their hybrids here, all of them may frequently interact with avian malaria parasites playing a role in the transmission of Plasmodium.
Similar content being viewed by others


Background
The Culex pipiens complex include species such as the common house mosquito Culex pipiens pipiens that is a wide spread species distributed over extensive areas in Europe and Africa and has been introduced in the Americas and in some temperate areas of Asia and Australia [1, 2]. Two forms or biotypes have been described: the molestus and the pipiens forms [1]. Although morphologically indistinguishable as adults, these two forms present genetic, behavioural and physiological differences. The molestus form is stenogamous and autogenous, that is mosquitoes are able to mate in confined environments and lay their eggs in absence of a previous blood meal, respectively. By contrast, mosquitoes of the pipiens form use open environments for mating (eurygamous) and requires a blood meal, as a nutrient source, for oviposition (anautogenous). In North European countries, the two forms use different habitats with Cx. p. pipiens form molestus frequently living in underground environments in areas with human influence, while mosquitoes of the pipiens form are mainly present in aboveground habitats [3, 4]. In countries of the Mediterranean basin, mosquitoes of the two forms are sympatric and, because molestus and pipiens forms are not completely genetically isolated, hybrids are frequent [5–8]. Hybridization between the two forms has been also occasionally reported in North Europe, in Germany [9], Austria [10], the Netherlands [11, 12] and the UK [13]. Hybridization between forms has also been reported under laboratory conditions [14].
The importance of mosquito species for the transmission of vector-borne pathogens is strongly determined by, among other factors, their feeding behaviour [15]. In addition to the differential competence of each mosquito species to develop a particular pathogen, the feeding behaviour of mosquitoes determines the contact rate between infected and susceptible vertebrate hosts. Culex p. pipiens play a key role in the transmission of numerous vector borne pathogens affecting humans, livestock and wildlife including viruses (e.g. West Nile virus, WNV; [16]), protozoa (e.g. avian malaria; [17]) and metazoa (e.g. filarial worms [18]) parasites (reviewed by [2]). Culex p. pipiens females feed mainly on birds (69–97%) [19], but mammals are also an important fraction of their blood meals, compromising over 20% of the blood meals in some populations [19, 20], also see [2, 21, 22]. However, it has been suggested that Cx. p. pipiens form molestus feed mainly on mammals while Cx. p. pipiens form pipiens feed mainly on birds [23], see also [24]. Hybrids show a higher vectorial competence for the transmission of some pathogens than both pipiens and molestus forms (e.g. WNV, [25]), and it has been suggested that due to their intermediate feeding behaviour [23], they may act as bridge vectors for the transmission of pathogens between birds and humans, as in the case of WNV [4].
Avian malaria parasites of the genus Plasmodium are mosquito-borne parasites commonly found infecting birds [26]. Nowadays, approximately 40 different species have been described [26] but, since the seminar paper by Bensch et al. [27], a high genetic diversity of cytochrome b (cyt b) lineages of Plasmodium parasites have been identified. Several mosquito genera transmit avian malaria parasites, including different species of the genus Culex, which may play a central role in the transmission of avian malaria [17]. In particular, Cx. p. pipiens may play a key role in the transmission dynamic of Plasmodium parasites under natural conditions. Plasmodium infected birds are common hosts of this mosquito species, allowing frequent parasite-Cx. p. pipiens encounters [22, 28]. Furthermore, avian Plasmodium parasites, including P. relictum, successfully develops in Cx. p. pipiens as supported by experimental infections of mosquitoes [17, 29–31], and different avian Plasmodium lineages have been molecularly isolated from mosquitoes trapped in the wild [32–37].
The aims of this study were (1) to investigate the spatial distribution of pipiens and molestus forms and their hybrids in relation to the type of habitats (natural, rural and urban areas), (2) to compare the feeding patterns of both forms in these habitats and (3) to determine avian malaria prevalence and diversity in these mosquitoes. Previous evidence suggest that the two Cx. p. pipiens forms and their hybrids use differential habitats [3, 10], thus a higher frequency of the molestus form is expected in urban areas than in natural ones, while the opposite pattern is expected for mosquitoes of the pipiens form. Furthermore, the molestus form may feed mainly on mammals, blood meals from mosquitoes of the pipiens form should be predominantly avian-derived [38] while their hybrids may show an intermediate feeding behaviour [23, 38]. If the feeding pattern differs between mosquito forms, the prevalence of infection by avian malaria parasites will be higher in the pipiens form due to the expected higher frequency of avian-derived blood meals with a decreased prevalence in mosquitoes of the molestus form and hybrids as the proportion of mammal-derived blood meals increased.
Methods
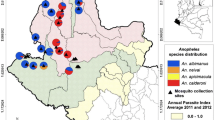
Mosquitoes were captured using BG-sentinel traps baited with BG-lure and dry ice as a source of CO2. Overall, from April to December, 5–6 trapping sessions were conducted at each site. In each trapping session, three traps were operated for 24 h at 45 different sampling sites in Cadiz, Huelva and Seville provinces (15 per each province) once every 45 days (Fig. 1). The sampling sites were grouped in triplets including one natural habitat (conserved landscapes with low density of human and livestock), one rural habitat (farms with livestock) and one urban habitat (urbanized densely populated areas), see [39] for further details of sampling protocol.

Distribution of the 45 mosquito sampling sites in Southern Spain
Adult mosquitoes were preserved in dry ice and stored frozen until morphological identification. Mosquitoes were sorted on a Petri plate on a chill table under a stereomicroscope and morphologically identified to species level following [40, 41]. Culex p. pipiens females with a recent blood meal in the abdomen were selected for this study. The abdomen of engorged mosquitoes was separated from the head-thorax using sterile tips. Genomic DNA of each the abdomen and the head-thorax of each mosquito was extracted using the DNA Kit Maxwell® 16LEV kit [42].
The DNA extracted from the mosquito abdomen was employed to identify the vertebrate origin of the blood meals using a nested PCR following Alcaide et al. [43]. Briefly, this method amplifies a 758-base pairs fragment (excluding primers) of the mitochondrial cytochrome oxidase 1 (COI) gene (barcoding region) of vertebrate species. Amplicons were sequenced in the Macrogen sequencing service (Macrogen Inc., The Netherlands). Vertebrate hosts of mosquitoes were identified by comparisons of the sequences obtained from mosquitoes with those available in public databases (GenBank DNA sequence database, National Center for Biotechnology Information Blast, and/or the Barcode of Life Data Systems).
The DNA extracted from the head-thorax of each engorged mosquito was used to identify the Cx. p. pipiens form and the presence and identity of avian malaria parasite lineages. The mosquito forms were identified following Bahnck and Fonseca [44]. This method is based in the amplification of the 5′ flacking region of CQ11 microsatellite and has been routinely used to identify the Cx. p. pipiens forms in Old-world countries [5, 7, 8], including Spain [45]. The presence and identity of Plasmodium and Haemoproteus parasites in the head-thorax of mosquitoes was determined following Helgreen et al. [46]. Parasite lineages were determined by BLAST comparison of the sequences with those deposited in Genbank and the morphospecies was determined based on the information available in MalAvi [47].
Statistical analyses were conducted using the GLIMMIX procedure in SAS 9.2 with a binomial distribution using a Laplace approximation. Firstly, generalized linear mixed models were used to identify potential differences in the occurrence of each form (molestus or pipiens) and their hybrids between habitats. The triplet (see sampling sites above) was included as a random factor in order to control for geographical pseudo-replication of the samples. Secondly, differences in the presence of avian-derived blood meals between mosquito forms were tested using the presence/absence of avian derived blood meals as the dependent variable, mosquito forms and habitat category as fixed factors and triplet as a random factor. Finally, differences in the prevalence of blood parasites between mosquito forms and habitats were tested by including parasite infection status as the dependent variable, mosquito forms and habitat category as fixed factors and triplet as a random factor.
Results
Overall, the vertebrate origin of 120 mosquito blood meals was identified to the species/genus level, corresponding to 33, 36, 51 individuals trapped in natural, rural and urban areas, respectively. Frequency of mosquitoes of the pipiens form differed between habitats (Fig. 2; F2,103 = 3.96; p = 0.02) being significantly (t = 32.77; p < 0.01) more frequently found in natural (44%, 16/36) than in urban areas (22%, 8/36). Mosquitoes of the molestus form were more frequently found in urban (60%, 27/45) than in natural areas (18%, 8/45), although differences among habitat categories did not reach significance (Fig. 1; F = 2.19; df = 2,103; p = 0.12). Both molestus (22%, 10/45) and pipiens (33%, 12/36) forms showed intermediate percentages in rural environments. Hybrids were similarly distributed between habitat categories (Fig. 2; F2,103 = 0.55; p = 0.58; natural: 23%, 9/39; rural: 36%, 14/39; urban: 41%, 16/39).

Percentage of mosquitoes of the form pipiens, form molestus and their hybrids found in natural (black), rural (grey) and urban (white) areas in Southern Spain. Bars indicate 95% confidence intervals
Most of the blood meals derived from birds (n = 80, 66.7%), while only 40 mosquitoes fed on mammal blood (33.3%) (Table 1). The vertebrate hosts identified from mosquitoes included, at least, 20 bird and 9 mammal species. House sparrows and dogs were the most common bird and mammal hosts of mosquitoes, respectively. Moreover, humans were identified as hosts of pipiens and molestus forms and their hybrids (Table 1). The presence of avian-derived blood meals did not differ between forms nor habitats (mosquito form: F2,101 = 0.16; p = 0.85; habitat: F2,101 = 1.98; p = 0.14).
Parasites were isolated from the head-thorax of eight mosquito females (overall prevalence 6.7%). Two Plasmodium lineages were isolated from seven mosquitoes including the cosmopolitan lineage SGS1 corresponding to Plasmodium relictum (n = 2, overall prevalence: 1.7%) and the lineage SYAT05 (=Rinshi-11) corresponding to Plasmodium vaughani (n = 5; overall prevalence: 4.2%). The Haemoproteus lineage Padom05 belonging to Haemoproteus passeris was isolated from one mosquito of the form pipiens with a house sparrow derived blood meal (Table 2; overall prevalence: 0.8%). The prevalence of blood parasites did not significantly differ between mosquito forms (mosquito form: F2,101 = 0.96; p = 0.39; habitat: F2,101 = 1.51; p = 0.23), although the absence of significant differences could be due to the low prevalence and the low number of mosquitoes analysed.
Discussion
A higher percentage of the pipiens form was found in natural than in urban areas, while the percentage of mosquitoes of the molestus form and hybrids did not differ between the three habitat categories considered in this study. Traditionally, anautogenous mosquitoes, a characteristic typically assigned to the form pipiens, are more commonly found in aboveground and rural habitats, while the autogenous form, corresponding to Cx. p. pipiens form molestus, is usually found in underground urban environments [1, 4, 48, 49]. However, reports of the differential distribution of genetically characterized mosquito forms in areas with different degrees of anthropization are scarce, especially in Southern Europe. In Portugal, the proportion of mosquitoes of the pipiens form was higher in farms located in peri-urban areas without dwellings in the vicinity than indoors in residential areas, supporting a negative association between the frequency of this form and the degree of urbanization [38]. In addition, although the frequency of molestus forms did not significantly differ between urban and peri-urban habitats, the proportion of molestus mosquitoes tended to be higher in indoors traps in areas densely populated by humans rather than in small villages, suggesting the higher presence of this form in urbanized environments [38]. Our results support those from Osorio et al. [38], in spite of the differences between studies regarding the sampling locations (indoors/outdoors).
It is generally assumed that pipiens and molestus forms have a bird and mammal blood feeding preference respectively, while hybrids show an intermediate behaviour [23]. This fact suggest that hybrids may play a central role in the transmission of pathogens such as WNV and USUTU virus (USUV), which circulate between birds but occasionally infect humans. In North America, mosquitoes with a higher ancestry from the molestus form feed more frequently on mammals, including humans [24, 50]. Contrary to this pattern, we did not find any significant association between mosquito forms and their feeding sources. The two forms and their hybrids feed mainly on birds, with a relative high percentage (33%) of blood meals derived from mammals, including humans. A similar feeding pattern of Cx. p. pipiens was reported in Portugal, where mosquitoes of the two forms feed mainly on birds with not significant differences between forms [51], but see [38]. One potential explanation is the different approaches used to identify the mosquito forms between studies, while studies from North America genotyped mosquitoes using seven [50] or ten [24] microsatellite markers, here the mosquito forms were assessed with the CQ11 microsatellite. However, Gomes et al. [51] used six microsatellites and obtained very similar results to our study. Alternatively, mosquito feeding pattern is geographically-dependent [1, 21] and varies seasonally [50, 52]. This variability between areas and seasons may partially explain discrepancies in the feeding source of mosquito forms between studies. The similar feeding behaviour of the two mosquito forms and their hybrids found here supports that all of them could be involved in the transmission of pathogens from birds to humans, as WNV or USUV.
Overall, 6.7% of mosquitoes harboured blood parasites in this study. Glaizot et al. [33] found a similar prevalence (6.6%) in Culex pipiens, although slightly higher values were reported in other previous studies [36, 37]. Differences between studies could be due to the temporal variation in the prevalence of Plasmodium usually found in mosquitoes [35, 36], but also due to methodological differences between studies including the protocol used for parasite detection (e.g. using engorged mosquitoes or mosquitoes with a completely digested blood meal) or the method used for mosquito sampling [53]. Furthermore, the number of mosquitoes with a mammal and avian derived blood meal may differ between trapping methods [54] potentially biasing estimates of parasite prevalence. This fact should be take into account in studies on the epidemiology of vector-borne parasites transmitted by mosquitoes with an opportunistic feeding behaviour. Nevertheless, in this study, the prevalence of avian malaria parasites did not differ between the two forms and their hybrids, which may be due at least in part, to the similar blood feeding behaviour reported between them. However, the absence of significant differences between forms could also be due to the low statistical power owing to the number of mosquitoes analysed and the low prevalence of blood parasites. Avian Plasmodium is a mosquito-borne parasite while the related Haemoproteus is mainly transmitted by Culicoides and louse flies [26]. As occurred here, previous studies have reported the presence of Haemoproteus in mosquitoes [22, 34, 35, 37, 55], however experimental evidence available exclude mosquitoes as vectors of this parasite genus [31, 56]. The Haemoproteus passeris lineage Padom05 was isolated from a mosquito with a recent blood meal on a house sparrow, which is a common avian host of this parasite lineage [31, 57]. This, together with the fact that the mosquito had a recent blood meal from this bird species, suggests that the Haemoproteus DNA derived from rests of undigested blood present in the head-thorax of the mosquito. Alternatively, it is possible that a non-infective form of the parasite present in the mosquito head-thorax was amplified [56], but see [31]. Although molecular isolation does not demonstrate vector competence, the Plasmodium parasites isolated from mosquitoes suggest that the two mosquito forms and their hybrids may frequently interact with birds infected by avian malaria parasites. Moreover, the Plasmodium lineages found in this study have been previously isolated from Cx. pipiens mosquitoes in Europe including Switzerland [33, 36], Portugal [58], Spain [35], France [37] and Italy [22]. Avian Plasmodium develops in laboratory reared Cx. p. pipiens form molestus [29, 30] and these parasites have been molecularly identified in field-collected mosquitoes of this form [32]. Complete development of avian Plasmodium has been reported in mosquitoes of the pipiens form [17]. However, to our knowledge, this is the first study testing the potential differences of the two Cx. p. pipiens forms and their hybrids in the prevalence of avian Plasmodium under natural conditions. Results from this study suggest that, given the similar feeding behaviour of mosquito forms in the study area, the two mosquito forms and their hybrids are similarly involved in the transmission of avian Plasmodium in Southern Spain.
Conclusion
This study supports a differential frequency of mosquitoes of the pipiens form between urban and natural areas. While the hypothesis that both forms and their hybrids differ in their feeding sources was not supported by results from this study, with birds as the most common blood source found in all the cases. Consequently, Cx. p. pipiens molestus and pipiens mosquitoes and their hybrids may frequently interact with WNV, USUV and/or avian malaria parasites playing a role in the transmission of these pathogens.
References
Vinogradova EB. Ecophysiological and morphological variations in mosquitoes of the Culex pipiens complex (Diptera: Culicidae). Acta Soc Zool Bohem. 2003;67:41–50.
Farajollahi A, Fonseca DM, Kramer LD, Kilpatrick AM. “Bird biting” mosquitoes and human disease: a review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect Genet Evol. 2011;11:1577–85.
Byrne K, Nichols RA. Culex pipiens in London underground tunnels: differentiation between surface and subterranean populations. Heredity. 1999;82:7–15.
Fonseca DM, Keyghobadi N, Malcolm CA, Mehmet C, Schaffner F, Mogi M, Fleischer RC, Wilkerson RC. Emerging vectors in the Culex pipiens complex. Science. 2004;303:1535–8.
Amraoui F, Tijane M, Sarih M, Failloux AB. Molecular evidence of Culex pipiens form molestus and hybrids pipiens/molestus in Morocco, North Africa. Parasit Vectors. 2012;5:83.
Gomes B, Kioulos E, Papa A, Almeida APG, Vontas J, Pinto J. Distribution and hybridization of Culex pipiens forms in Greece during the West Nile virus outbreak of 2010. Infect Genet Evol. 2013;16:218–25.
Krida G, Rhim A, Daaboub J, Failloux AB, Bouattour A. New evidence for the potential role of Culex pipiens mosquitoes in the transmission cycle of West Nile virus in Tunisia. Med Vet Entomol. 2015;29:124–8.
Di Luca M, Toma L, Boccolini D, Severini F, La Rosa G, Minelli G, Bongiorno G, Montarsi F, Arnoldi D, Capelli G, Rizzoli A, Romi R. Ecological distribution and CQ11 genetic structure of Culex pipiens complex (Diptera: Culicidae) in Italy. PLoS ONE. 2016;11:e0146476.
Rudolf M, Czajka C, Börstler J, Melaun C, Jöst H, von Thien H, et al. First nationwide surveillance of Culex pipiens complex and Culex torrentium mosquitoes demonstrated the presence of Culex pipiens biotype pipiens/molestus hybrids in Germany. PLoS ONE. 2013;8:e71832.
Zittra C, Flechl E, Kothmayer M, Vitecek S, Rossiter H, Zechmeister T, et al. Ecological characterization and molecular differentiation of Culex pipiens complex taxa and Culex torrentium in eastern Austria. Parasit Vectors. 2016;9:197.
Reusken CBEM, De Vries A, Buijs J, Braks MAH, Den Hartog W, Scholte E-J. First evidence for presence of Culex pipiens biotype molestus in the Netherlands, and of hybrid biotype pipiens and molestus in northern Europe. J Vector Ecol. 2010;35:210–2.
Vogels CB, van de Peppel LJ, van Vliet AJ, Westenberg M, Ibañez-Justicia A, Stroo A, et al. Winter activity and aboveground hybridization between the two biotypes of the West Nile virus vector Culex pipiens. Vector Borne Zoonotic Dis. 2015;15:619–26.
Danabalan R, Ponsonby DJ, Linton Y-M. A critical assessment of available molecular identification tools for determining the status of Culex pipiens s.l. in the United Kingdom. J Am Mosq Control Assoc. 2012;28:68–74.
Duron O, Bernard C, Unal S, Berthomieu A, Berticat C, Weill M. Tracking factors modulating cytoplasmic incompatibilities in the mosquito Culex pipiens. Mol Ecol. 2006;15:3061–71.
Simpson JE, Hurtado PJ, Medlock J, Molaei G, Andreadis TG, Galvani AP, et al. Vector host-feeding preferences drive transmission of multi-host pathogens: West Nile virus as a model system. Proc R Soc Lond B. 2012;279:925–33.
Engler O, Savini G, Papa A, Figuerola J, Groschup MH, Kampen H, et al. European surveillance for West Nile virus in mosquito populations. Int J Environ Res Public Health. 2013;10:4869–95.
Santiago-Alarcon D, Palinauskas V, Schaefer HM. Diptera vectors of avian haemosporidian parasites: untangling parasite life cycles and their taxonomy. Biol Rev Camb Philos Soc. 2012;87:928–64.
Morchón R, Bargues MD, Latorre JM, Melero-Alcíbar R, Pou-Barreto C, Mas-Coma S, et al. Haplotype H1 of Culex pipiens implicated as natural vector of Dirofilaria immitis in an endemic area of Western Spain. Vector Borne Zoonotic Dis. 2007;7:653–8.
Gómez-Díaz E, Figuerola J. New perspectives in tracing vector-borne interaction networks. Trends Parasitol. 2010;26:470–6.
Martínez-de la Puente J, Ruiz S, Soriguer R, Figuerola J. Effect of blood meal digestion and DNA extraction protocol on the success of blood meal source determination in the malaria vector Anopheles atroparvus. Malar J. 2013;12:109.
Muñoz J, Ruiz S, Soriguer R, Alcaide M, Viana DS, Roiz D, et al. Feeding patterns of potential West Nile virus vectors in South-West Spain. PLoS ONE. 2012;7:e39549.
Martínez-de la Puente J, Muñoz J, Capelli G, Montarsi F, Soriguer R, Arnoldi D, et al. Avian malaria parasites in the last supper: identifying encounters between parasites and the invasive Asian mosquito tiger and native mosquito species in Italy. Malar J. 2015;14:32.
Fritz ML, Walker ED, Miller JR, Severson DW, Dworkin I. Divergent host preferences of above- and below-ground Culex pipiens mosquitoes and their hybrid offspring. Med Vet Entomol. 2015;29:115–23.
Huang S, Hamer GL, Molaei G, Walker ED, Goldberg TL, Kitron UD, et al. Genetic variation associated with mammalian feeding in Culex pipiens from a West Nile virus epidemic region in Chicago, Illinois. Vector Borne Zoonotic Dis. 2009;9:637–42.
Ciota AT, Chin PA, Kramer LD. The effect of hybridization of Culex pipiens complex mosquitoes on transmission of West Nile virus. Parasit Vectors. 2013;6:305.
Valkiūnas G. Avian malaria parasites and other haemosporidia. Boca Ratón: CRC Press; 2005.
Bensch S, Stjernman M, Hasselquist D, Ostman O, Hansson B, Westerdahl H, et al. Host specificity in avian blood parasites: a study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc Biol Sci. 2000;267:1583–9.
Spielman A. Structure and seasonality of nearctic Culex pipiens populations. Ann NY Acad Sci. 2001;951:220–34.
Žiegytė R, Bernotienė R, Bukauskaitė D, Palinauskas V, Iezhova T, Valkiūnas G. Complete sporogony of Plasmodium relictum (lineages pSGS1 and pGRW11) in mosquito Culex pipiens pipiens form molestus, with implications to avian malaria epidemiology. J Parasitol. 2014;100:878–82.
Valkiūnas G, Žiegytė R, Palinauskas V, Bernotienė R, Bukauskaitė D, Ilgūnas M, et al. Complete sporogony of Plasmodium relictum (lineage pGRW4) in mosquitoes Culex pipiens pipiens, with implications on avian malaria epidemiology. Parasitol Res. 2015;114:3075–85.
Gutiérrez-López R, Martínez-de la Puente J, Gangoso L, Yan J, Soriguer RC, Figuerola J. Do mosquitoes transmit the avian malaria-like parasite Haemoproteus? An experimental test of vector competence using mosquito saliva. Parasit Vectors. 2016;9:609.
Kim KS, Tsuda Y, Yamada A. Bloodmeal identification and detection of avian malaria parasite from mosquitoes (Diptera: Culicidae) inhabiting coastal areas of Tokyo Bay, Japan. J Med Entomol. 2009;46:1230–4.
Glaizot O, Fumagalli L, Iritano K, Lalubin F, Van Rooyen J, Christe P. High prevalence and lineage diversity of avian malaria in wild populations of great tits (Parus major) and mosquitoes (Culex pipiens). PLoS ONE. 2012;7:e34964.
Inci A, Yildirim A, Njabo KY, Duzlu O, Biskin Z, Ciloglu A. Detection and molecular characterization of avian Plasmodium from mosquitoes in central Turkey. Vet Parasitol. 2012;188:179–84.
Ferraguti M, Martínez-de la Puente J, Muñoz J, Roiz D, Ruiz S, Soriguer R, et al. Avian Plasmodium in Culex and Ochlerotatus mosquitoes from southern Spain: effects of season and host-feeding source on parasite dynamics. PLoS ONE. 2013;8:e66237.
Lalubin F, Delédevant A, Glaizot O, Christe P. Temporal changes in mosquito abundance (Culex pipiens), avian malaria prevalence and lineage composition. Parasit Vectors. 2013;2013(6):307.
Zélé F, Vézilier J, L’Ambert G, Nicot A, Gandon S, Rivero A, Duron O. Dynamics of prevalence and diversity of avian malaria infections in wild Culex pipiens mosquitoes: the effects of Wolbachia, filarial nematodes and insecticide resistance. Parasit Vectors. 2014;7:1.
Osório HC, Zé-Zé L, Amaro F, Nunes A, Alves MJ. Sympatric occurrence of Culex pipiens (Diptera, Culicidae) biotypes pipiens, molestus and their hybrids in Portugal, Western Europe: feeding patterns and habitat determinants. Med Vet Entomol. 2014;28:103–9.
Ferraguti M, Martínez-de la Puente J, Roiz D, Ruiz S, Soriguer R, Figuerola J. Effects of landscape anthropization on mosquito community composition and abundance. Sci Rep. 2016;6:29002.
Schaffner F, Angel G, Geoffroy B, Hervy JP, Rhaiem A, Brunhes J. Les moustiques d’Europe/The mosquitoes of Europe. CD-ROM. Montpellier: Institut de Recherche pour le Développement/EID Méditerranée; 2001.
Becker ND, Petrić D, Zgomba M, Boase C, Dahl C, Madon M, et al. Mosquitoes and their control. 2nd ed. Berlin: Springer Verlag; 2010.
Gutiérrez-López R, Martínez-de la Puente J, Gangoso L, Soriguer RC, Figuerola J. Comparison of manual and semi-automatic DNA extraction protocols for the barcoding characterization of hematophagous louse flies (Diptera: Hippoboscidae). J Vector Ecol. 2015;40:11–5.
Alcaide M, Rico C, Ruiz S, Soriguer R, Muñoz J, Figuerola J. Disentangling vector-borne transmission networks: a universal DNA barcoding method to identify vertebrate hosts from arthropod bloodmeals. PLoS ONE. 2009;4:e7092.
Bahnck CM, Fonseca DM. Rapid assay to identify the two genetic forms of Culex (Culex) pipiens L. (Diptera: Culicidae) and hybrid populations. Am J Trop Med Hyg. 2006;75:251–5.
Brustolin M, Talavera S, Santamaría C, Rivas R, Pujol N, Aranda C, et al. Culex pipiens and Stegomyia albopicta (=Aedes albopictus) populations as vectors for lineage 1 and 2 West Nile virus in Europe. Med Vet Entomol. 2016;30:166–73.
Hellgren O, Waldenstrom J, Bensch S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium and Haemoproteus from avian blood. J Parasitol. 2004;90:797–802.
Bensch S, Hellgren O, Pérez-Tris J. MalAvi: a public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol Ecol Res. 2009;9:1353–8.
Vinogradova EB. Culex pipiens pipiens mosquitoes: taxonomy, distribution, ecology, physiology, genetics, applied importance and control, vol. 2. Sofia, Moscow: Pensoft Publishers; 2000.
Vinogradova EB, Shaikevich EV. Morphometric, physiological and molecular characteristics of underground populations of the urban mosquito Culex pipiens Linnaeus f. molestus Forskål (Diptera: Culicidae) from several areas of Russia. Eur Mosq Bull. 2007;22:17–24.
Kilpatrick AM, Kramer LD, Jones MJ, Marra PP, Daszak P, Fonseca DM. Genetic influences on mosquito feeding behavior and the emergence of zoonotic pathogens. Am J Trop Med Hyg. 2007;77:667–71.
Gomes B, Sousa CA, Vicente JL, Pinho L, Calderón I, Arez E, et al. Feeding patterns of molestus and pipiens forms of Culex pipiens (Diptera: Culicidae) in a region of high hybridization. Parasit Vectors. 2013;6:93.
Kilpatrick AM, Kramer LD, Jones MJ, Marra PP, Daszak P. West Nile Virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLoS Biol. 2006;4:e82.
Carlson JS, Walther E, TroutFryxell R, Staley S, Tell LA, Sehgal RNM, Barker CM, Cornel AJ. Identifying avian malaria vectors: sampling methods influence outcomes. Parasit Vectors. 2015;8:365.
Roiz D, Roussel M, Muñoz J, Ruiz S, Soriguer R, Figuerola J. Efficacy of mosquito traps for collecting potential West Nile mosquito vectors in a natural Mediterranean wetland. Am J Trop Med Hyg. 2012;86:642–8.
Ishtiaq F, Guillaumot L, Clegg SM, Phillimore AB, Black RA, Owens IP, Mundy NI, Sheldon BC. Avian haematozoan parasites and their associations with mosquitoes across Southwest Pacific Islands. Mol Ecol. 2008;17:4545–55.
Valkiūnas G, Kazlauskienė R, Bernotienė R, Palinauskas V, Iezhova TA. Abortive long-lasting sporogony of two Haemoproteus species (Haemosporida, Haemoproteidae) in the mosquito Ochlerotatus cantans, with perspectives on haemosporidian vector research. Parasitol Res. 2013;112:2159–69.
Marzal A, Ricklefs RE, Valkiūnas G, Albayrak T, Arriero E, Bonneaud C, et al. Diversity, loss, and gain of malaria parasites in a globally invasive bird. PLoS ONE. 2011;6:e21905.
Ventim R, Ramos JA, Osório H, Lopes RJ, Pérez-Tris J, Mendes L. Avian malaria infections in western European mosquitoes. Parasitol Res. 2012;111:637–45.
Authors’ contributions
JMP, MF, SR, RS and JF conceived and designed the study. JMP, MF, SR, and DR sampled mosquitoes and/or analysed the samples. JMP drafted the manuscript with valuable contribution of the other authors. All authors read and approved the final manuscript.
Acknowledgements
Special thanks to Antonio Magallanes Martín de Oliva, Juana Moreno Fernández, Esmeralda Pérez and Cristina Pérez for their help during mosquito sampling and identification and to Isabel Martín for their invaluable help with the molecular analyses. Laboratorio de SIG y Teledetección, Estación Biológica de Doñana, CSIC (LAST-EBD) provided logistical support.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Data supporting the conclusions of this article are included within the article. Additional datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
Ethics statement
Mosquito sampling was conducted with all the necessary permits issued by the regional Department of the Environment (Consejería de Medio Ambiente, Junta de Andalucía). Entomological surveys and sampling on private land and in private residential areas were conducted with all the necessary permits and consent, and in the presence of owners. This study did not affect any endangered or protected species.
Funding
This study was funded by projects CGL2015-65055-P from the Spanish Ministry of Science and Innovation and P11-RNM-7038 from the Junta de Andalucía. J.M.P. and M.F. were funded by a Juan de la Cierva contract and a FPU grant, respectively.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Martínez-de la Puente, J., Ferraguti, M., Ruiz, S. et al. Culex pipiens forms and urbanization: effects on blood feeding sources and transmission of avian Plasmodium . Malar J 15, 589 (2016). https://doi.org/10.1186/s12936-016-1643-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-016-1643-5