
Abstract
Breast cancer (BC) represents aggressive cancer affecting most women’s lives globally. Metastasis and recurrence are the two most common factors in a breast cancer patient's poor prognosis. Cancer stem cells (CSCs) are tumor cells that are able to self-renew and differentiate, which is a significant factor in metastasis and recurrence of cancer. Long non-coding RNAs (lncRNAs) describe a group of RNAs that are longer than 200 nucleotides and do not have the ability to code for proteins. Some of these lncRNAs can be mainly produced in various tissues and tumor forms. In the development and spread of malignancies, lncRNAs have a significant role in influencing multiple signaling pathways positively or negatively, making them promise useful diagnostic and prognostic markers in treating the disease and guiding clinical therapy. However, it is not well known how the interaction of lncRNAs with CSCs will affect cancer development and progression.
Here, in this review, we attempt to summarize recent findings that focus on lncRNAs affect cancer stem cell self-renewal and differentiation in breast cancer development and progression, as well as the strategies and challenges for overcoming lncRNA's therapeutic resistance.
Similar content being viewed by others


Introduction
Breast cancer (BC) is the most frequently diagnosed cancer in women worldwide, and it is ranked as second in terms of cancer related death in women [1]. Advances in early diagnosis and therapy techniques have reduced BC death and to some extent, improved patient prognosis and survival rate [2]. However, the emergence of drug resistance in BC patients has decreased the success rate of systemic therapies [3]. Cancer stem cells (CSCs), also known as cancer-producing cells, are characterized by the unique feature of self-renewal and differentiation into tumor-propagating cells [4, 5]. Approximately CSCs make up about 0.01–2% of all tumor cells in the body, which play a critical role in cancer initiation, progression, apoptotic endurance, and therapeutic resistance [6, 7]. CSCs first described in 2003 by Visvader and Linderman, describing CSCs role in acute myeloid leukemia (AML) [8].
Breast cancer stem cells (BrCSCs) can be formed from differentiated mammary cells due to disease-related mutations. The specific origin of BrCSCs has been a contentious issue for decades, and it's unclear where they come from. Exposure to environmental stimuli such as radiation causes genetic defects in non-malignant somatic cells and causes dedifferentiation of non-malignant somatic cells. Microenvironmental stimuli also can trigger a malignant transformation of differentiated cells into BrCSCs. Removing CSCs from the body is seen as a novel approach for the complete elimination of cancerous tumors and their prevention from recurrence [9].
It is estimated that 90% of the human genome is translated into RNAs [10,11,12], and less than two % of these transcripts are used to produce proteins [13]. Noncoding RNAs (ncRNAs) are a class of genes that have limited or no ability to code for proteins and are linked to cellular functions and disease development. Based on their length, non-coding RNAs (ncRNAs) are classified into two main classes based on their sizes: long ncRNAs (lncRNAs) more than 200nt in length and small ncRNAs less than 200nt in length [14].
According to certain studies, CSCs' activity is intimately linked to the abnormal expression of lncRNAs in malignant tumors. For example, OCT4 [15], SOX2 [16], KLF4 [17], and other stem cell-related pathways are regulated by many lncRNAs, which in turn affect CSC activities [18]. Furthermore, the control of miR-34a by lncRNAs, which were involved in tumor growth, was already reported in several studies to modulate CSC-like features in a range of malignancies [19, 20]. Recently, cancer-related research on lncRNA and CSCs is gaining growing attention. This study provides a comprehensive analysis of the current research status on lncRNAs and BrCSCs, their mechanisms of action, and the contribution of lncRNAs to tumorigenesis in BC by regulating CSCs, which could be used as potential diagnostic biomarkers and therapeutic strategies. This review can motivate further research to validate the highlighted lncRNAs-CSCs role as potential diagnostic biomarkers and therapeutic strategies in BC patients.
LncRNAs in cancer biology
The first regulatory non-coding RNAs (ncRNAs) were discovered in bacteria in the 1980s, and then in most eukaryotic organisms. A few long noncoding RNAs (lncRNAs) such as H19 and Xist (X-inactive specific transcript) were identified prior to the genomic era, but they remained exceptions until the early 2000s [21]. They were later discovered to have five distinct origins: (A) a protein-coding gene undergoes structural damage and is transformed into a lncRNA; (B) chromosomal rearrangement brings two non-transcribed regions together, resulting in a lncRNA with multiple exons; (C) retrotranspositional duplication of a noncoding gene produces either a functional retrogene or a nonfunctional retropseudogene, both without encoding proteins; (D) two tandem duplication events result in adjacent repeats within a noncoding RNA; and (E) transposon insertion results in a functional lncRNA [22].
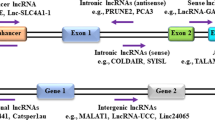
The presence of 18,805 LncRNAs has been established by GENCODE (release 40 (GRCh38.p13)), with about 39% of them found between genes and referred to as long intergenic ncRNAs (lincRNAs) [23]. Intronic lncRNAs, overlapping, and antisense are among the others [24]. They control a wide range of gene functions at the transcriptional [25], post-transcriptional [26], and epigenetic levels [27] by interacting with mRNAs, miRNAs, genomic DNA, and proteins. Thus, lncRNAs play a role in gene expression regulation, histone modification, chromatin remodeling, transcription and post-transcription splicing regulation, and translation.
Their relation to tumorigenesis is dependent on abnormal expression in a variety of cancers, especially BC. Guide, signal, scaffold, and decoy are the main functions of lncRNAs [28] (Fig. 1). A number of lncRNAs can help regulate gene expression by directing the recruitment of transcriptional activators and suppressors to specific genomic locations [29, 30]. Guide lncRNAs are essential for the organization and localization of components at certain genomic loci in order to regulate the genome [31]. For example, HOTTIP, XIST, MEG3, COLDAIR, KCNQ1OT1, ANRIL, and TUG1 are all lncRNAs that can act as guides for epigenetic modifier recruitment to their specific loci [32]. Additionally, signal lncRNAs are expressed in response to external stimuli at a specified time and location within the cell. An example of this paradigm is that certain lncRNAs have regulatory functions, whereas others are essentially by-products of transcribing. Signaling lncRNAs have been shown to interact with chromatin-modifying enzymes such as histone methyltransferases to quiet their target genes by preventing their transcription or by forming heterochromatin [28]. Many lncRNAs may also function as scaffolding proteins, attracting chromatin remodeling complexes such as the PRC1 (polycomb repressive complex 1) and the PRC2 to inhibit specific target gene [33, 34]. In addition, several lncRNAs such as lncRNA ROR, DAPALR, and PANDA can serve as enhancer RNAs, have a role in stabilizing the looping and recruitment of regulatory transcriptional RNAs, cofactors, and RNA Pol II [35, 36]. LncRNA decoys inhibit transcription by preventing a specific effector from interacting with its intrinsic target [28]. By trapping regulatory components including miRNAs, altering chromatin subunits, and TF and restricting their availability, these lncRNAs indirectly regulate transcription process [37].

Transcriptional control via LncRNA
Since lncRNAs regulate gene expression, they have been connected to cellular mechanisms and diseases like stem cell maintenance and cancer metastasis [38, 39]. Perhaps this is because they can attach to DNA, RNA, or proteins.
Long non-coding RNAs and BrCSC renewal
The ability to self-renew through mitotic cell division and the ability to differentiate into specialized cell types are two of the most fundamental characteristics of stem cells. They can produce more undifferentiated stem cells due to the latter potential. The latter feature, on the other hand, allows the development of differentiated cell types within an organ.
According to the CSC hypothesis, malignancies are caused by the transformation of stem or progenitor cells capable of multilineage differentiation [40]. Although CSCs make up a small proportion out of the total tumor mass, their survival is dependent on this small cell population [41].
These cells, which have been given the designation of CSCs, exhibit characteristics of both stem cells and cancer cells. Self-renewal, differentiation, and the ability to form tumors are some of the characteristics of these cells. Firstly, Lapidot et al. revealed that human AML is organized as a hierarchical structure that is derived from primitive hematopoietic cells [42]. A decade later, this paradigm was used to study BC in NOD-SCID mice by Al-Hajj et al., who uncovered a tumorigenic subpopulation in the mice [43]. Published studies in the following years have demonstrated that CSCs are present in a wide range of cancers [44]. Later, Rosen and Jordan, showed that the Wnt, Notch, and Hedgehog (Hh) signaling pathways, which drive stem cell self-renewal can also control the functions of CSCs [45].
As cancer progresses, several factors are hypothesized to influence the characteristics of CSCs such as lncRNAs. Interestingly, lncRNAs have the potential to influence the CSCs to self-renew by modulating key cancer-related pathways through overexpression, deficiency, or mutation. These pathways include the epithelial-mesenchymal transition (EMT) [46], the Notch pathway [47], the Rho-GTPase system [48], and the NF-B cascade [49]. For example, EMT and BrCSC self-renewal has been shown to be induced by overexpression of linc00617, which decreases E-cadherin levels while elevating N-cadherin and Vimentin levels [50]. Furthermore, due to the enrichment of the CSC fraction, BC cell populations that overexpress lincRNA are more likely to form mammospheres and become tumorigenic. It binds to the promoter of the Sox2 gene and activates its transcription by recruiting hnRNP-K to the promoter which promotes self-renewal ability of CSCs [51]. Figure 1 describes the roles of lncRNAs in BC cells that can either promote or inhibit gene expression.
Prognostic role of LncRNAs in BrCSCs
In stem cell biology, lncRNAs are emerging as key contributors. LncRNAs have been identified to modulate pluripotency and differentiation of embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs), according to several studies. In addition, lncRNAs are becoming more significant adult stem cell regulators. OCT4, SOX2, KLF4, and PcG, all of which are critical in ESCs, have been found to be active in CSCs [52]. Depending on investigations on the existence and development of BC, BrCSCs appear to be resistant to chemotherapy, radiation, and hypoxia [53,54,55]. Additionally, the tumorigenicity and invasiveness of BrCSCs play a significant role in the initiation, proliferation, metastasis, and recurrence of BC [56].
In cancers dysregulation of lncRNAs [38, 57] contemplate the role of lncRNA in controlling stem cell signaling in cancer cells [58]. Further exploration of versatile functions of lncRNAs in CSC may unravel hidden and novel therapeutic strategies for overcoming chemotherapy resistance a major obstacle in BC treatment. LncRNAs, including MALAT-1, HOTAIR, and H19, are commonly found in patients with BC because they affect key signaling pathways involved in tumor promotion and suppression [38]. As a result of the research of deregulation mechanisms, it has become clear that lncRNAs might be used as biomarkers in the diagnosis, prognosis, and therapeutic strategies. Interestingly, lncRNAs play a pivotal role in BrCSCs by engaging in several critical pathways [59] and significantly impact BC progression. LncRNAs can promote or impede BC cell invasion and metastasis by activating or repressing the EMT during the initiation and progression of BC [60].
A variety of LncRNAs have been reported to be overexpressed in BrCSCs, including HOTAIR [61], MALATI [62,63,64], H19, DANCR [65,66,67], NR2F1-AS1 (NAS1) [68], NEAT1 [69, 70], NRAD1 [71], LINC-ROR [72, 73], linc00617 [50], CCAT1 [74], RP1-5O6.5 (RP1) [75], and lncRNA-Hh [76], and they all play a role in various cancer hallmarks, by the modulation of several proteins and miRNAs (Table 1).
LncRNAs regulate BrCSCs through epigenetic modifications
There are several ways in which the DNA sequence is not altered but epigenetic regulation affects gene expression; they include histone modifications, methylation of DNA, and genomic imprinting [81, 82]. LncRNAs are essential modulators of the epigenetic state of the human genome (Fig. 1). Chromatin remodeling and modification complexes are attracted to particular places by certain lncRNAs [83]. Examples of lncRNAs that can influence carcinogenesis include EPB41L4A-AS2 [84], BLAT1 [85], BCLIN25 [86], ANRASSF1 [87], H19 [88], and piR-823 [89] which can control DNA methylation. Han and his group found that BLAT1 expression is controlled by lowering the degree of promoter DNA methylation of CpG islands [90]. They showed that cancer patients with tumors that are BLAT1-hypomethylated have a decreased chance of surviving long-term. The high BLAT1 expression with hypomethylation at CpG sites in BC may be associated with the aggressiveness of the disease. BCLIN25 reduces miR-125b expression and elevates BC risk by increasing CpG methylation at the miR-125b promoter region, increasing ERBB2 expression [86]. Calanca et al. found that the tumor suppressor RASSF1A's epigenetic suppression has been linked to the lncRNA ANRASSF1. They proved that ANRASSF1 also is a therapeutic target with the potential to restore the restrictive chromatin changes generated by PRC2 at the promoter of RASSF1A, potentially leading to upregulation of RASSF1A in BC [91]. In addition, a lncRNA known as H19 suppresses the maternal allele at the H19/IGF2 gene via the methylation process in BC, resulting in a more aggressive phenotype [88]. A recent study demonstrated that lncRNA piR-823 overexpression in luminal BC cells activated Wnt signaling and induced cancer cell stemness by increasing DNMTs (DNMT1, DNMT3A, DNMT3B) expression and promoting adenomatous polyposis coli (APC) methylation [89].
In addition, lncRNA decreases gene transcription by recruiting proteins that modify histones or remodel chromatin. Histone regulators interact with the lncRNA HOTAIR, which results in transcriptional gene suppression in chromatin dynamics [92]. For example, HOTAIR contributes to the silencing of miR-205 by altering the balance of histone modifications on the miR-205 promoter between H3K4me3 and H3K27me3, which controls the production of cyclin J (CCNJ) [93]. Furthermore, HOTAIR inhibits miR-7 expression, leading to increased SETDB1 expression in BC stem cells, inducing EMT [94]. Moreover, HOTAIR promotes EMT by acetylating histone H3K27 to methylate the E-cadherin promoter, inhibiting E-cadherin production [95].
One of the most well-studied lncRNAs is Xist, active in the early phases of X chromosomal inactivation in female embryos. First instances of lncRNA directly engaged in the production of repressive chromatin are found in Xist [96]. Furthermore, Xist through particular RNA sequences, organizes the anchoring of chromatin modifiers to one of the two X chromosomes, enabling transcriptional silencing [97].
Several noncoding RNA molecules function as ligands and form complexes with transcription factors to regulate gene transcription [98, 99]. cis and trans regulation are two mechanisms used by lncRNAs to control gene transcription. Classifying lncRNAs into four categories by Wang and Chang namely; (decoy molecules, guiding molecules, scaffolding molecules, and signal molecules) contributed significantly to the progress of the lncRNA research area in the past few years [100]. The involvement of lncRNAs as a decoy, guiding, scaffolding, and signal molecules at the transcriptional level promotes or suppresses gene expression in BC metastasis (Fig. 1). For example, lncRNAs Lethe, NORAD, and PANDA work as "decoys," mimicking and competing with miRNAs or proteins in the nucleus [28, 98, 101] (Fig. 1a). Furthermore, the “guiding lncRNAs,” such as HOTAIR [102] and lincRNA-p21 [103], may directly act on transcriptional factors or chromatin modifiers and protein complexes and attract them to find particular target gene(s) to alter the transcription process (Fig. 1b) [104]. Moreover, LncRNAs MALAT1 [105], HOTAIR [106], and LINP1 [107] can form ribonucleoprotein complexes and regulate gene expression as scaffold molecules (Fig. 1c). In addition, to limit the activities of regulatory miRNAs, they also function as decoy microRNA-binding sites, like ceRNAs [108]. Additionally, various studies have revealed that lncRNAs can be used as molecular signals since they are transcribed at very particular times and locations, allowing cells to integrate developmental clues, read the cellular environment, or respond to a variety of stimuli as chemical signals [100, 109] (Fig. 1d). According to Wang and his colleagues, the lncTCF7 promotes TCF7 expression by recruiting SWI/SNF to TCF7's promoter. These steps may cause Wnt signaling to be activated, which might lead to cancer stem cells self-renewing and tumor cell proliferation [110]. Besides, the specific targets of guide lncRNAs are promoted by RNA–RNA, RNA–protein, and RNA–DNA interactions (Table 2).
Regulation of transcription in BrCSCs by LncRNAs
LncRNAs influence the translation of mRNAs and regulate their integrity at the posttranscriptional stage by creating double-stranded RNA with mRNAs or via binding to proteins. Tang and his group found that proliferation, colony formation, and development of orthotopic xenograft tumors were all reduced by the depletion of PVT1. Through KLF5/beta-catenin signaling, they also showed that lncRNA PVT1 controls TNBC [111]. Antisense lncRNAs are the most common lncRNAs engaged in mRNA post-transcriptional regulation. There are various ways in which lncRNAs might alter the splicing process of pre-mRNA, either alone or with other splicing factors. For example, the lncRNA RP1-506.5 interacts with eIF4E and inhibits eIF4E from binding to eIF4G, inhibiting p27kip1 translation and adversely regulating Snail levels in BC cells [75]. Likewise, Beta-catenin signaling is increased due to lncRNA PVT1 binding to KLF5 and increasing its stability via BAP1, which increases BC tumorigenesis [111]. The biological processes of malignancies are influenced by lncRNA dysfunction, which has been linked to tumor prognosis. The lncRNA encoded by metallothionein 1 J (MT1JP) has been linked to carcinogenesis, and its expression is downregulated in different types of cancer [112,113,114]. Meanwhile, in vivo and in vitro studies recently showed that MT1JP inhibition increases miRNA-214 gene expressions in BC cells by modulating miRNA-214/RUNX3 Axis [115]. Moreover, it was also shown that lncRNA treRNA interacts with ribonucleoproteins (RNPs) (PUF60, SF3B3, FXR1, FXR2, PUF60, and hnRNP K) to produce a treRNA-RNP complex that inhibits E-cadherin translation by targeting eIF4G1 [116] (Fig. 2).

LncRNAs and Post-transcriptional regulation in BrCSCs. a LncRNAs inhibit the initiation factor that involved in RNA translation such as elF4G which decreases the level of cyclin-dependent kinase inhibitor and promote cell proliferation. b Due to lncRNA PVT1 binding to KLF5 and promoting its stability through BAP1, beta-catenin signaling is accelerated, resulting in an increase in the development of beta-catenin tumors in BC patients. c LncRNA MT1JP suppression enhances miRNA-214 gene transcription through altering the miRNA-214/RUNX3 Axis, which results in Bim expression defect and suppresses the apoptosis process. d lncRNA treRNA forms complexes with ribonucleoproteins (RNPs) that inhibit E-cadherin translation by targeting eIF4G1 and increase the expression of EMT genes, resulting in increased cell migration and invasion
LncRNAs regulate BrCSCs through ceRNAs mechanism
It has been found that RNAs can interact with each other in a novel way (ceRNA networks), and that these interactions are critical to the formation of cancers. They have the potential to act as therapeutic targets in addition to being diagnostic and prognostic markers. According to literature studies, numerous lncRNAs act as ceRNAs in BrCSCs, targeting miRNAs and altering the EMT process. For instance, linc-ROR affects the expression of the stemness factors Nanog, Oct4, and SOX2 and regulates the maintenance of human embryonic stem cells (hESCs) via sponging miR-145 [72]. Further, upregulation of linc-ROR in BC resulted in a higher stem cell phenotype and greater mammosphere development, indicating that the Linc-ROR-miR-145 pathway is critical in cancer stemness [117, 118]. In addition, one of the most well-known and investigated ceRNA mechanisms is the lncRNA H19/Let-7 miRNA [119]. BrCSCs and breast tumors had a significant level of lncRNA H19 expression. Increasing the amount of H19 in tumor cells permits them to survive treatments [120], also H19 is involved in controlling BrCSCs' division by sponging microRNA let-7 which provide potential strategies for stem cell invasion and mammosphere formation [121]. Likewise, lncRNA HOTAIR (homeobox antisense transcript antisense RNA) is associated with the growth and spread of BC cells [122]. BrCSC derived from MCF7 or MB231 have high levels of HOTAIR up-regulation, which affects BrCSC growth, migration, and self-renewal. HOTAIR is also known to influence miR-34a, which in turn promotes Sox2 expression, according to a recent study [61]. HOTAIR can reduce the expression of miR-7 by regulating HoxD10 in MCF-7 and MDA-MB-231 BrCSCs [94]. Additionally, LUCAT1 promotes BrCSC stemness by sponging miR-5582-3p and promoting TCF7L2 and the Wnt/β-catenin pathway. On the other hand, BrCSCs self-renewal was hindered by LUCAT1 downregulation, which enhanced miR-5582-3p expression [123]. Moreover, lncCCAT1 is highly expressed in BrCSCs and is linked to poor patient consequences. LncCCAT1 overexpression promotes BrCSC proliferation, stemness, migration, and invasion capabilities. Depends on the above results, lncRNAs can be used as a marker of the patient's prognosis and the risk of their tumor recurrence which may apply in clinical translation. Meanwhile, miR-204/211, miR-148a/152, and ANXA2 can all interact with LncCCAT1, which in turn can activate TCF4 or promote the translocation of β-catenin to the nucleus and activate Wnt signaling [124]. Additionally, in a recent study, Song and his team found that lncRNA SPRY4-IT1 enhances BC cell proliferation and stemness, as well as BrCSC renewal and stemness maintenance, via promoting the expression of TCF7L2 through targeting miR-6882-3p [125].
Besides, LincK also controls ZEB1 through the sponging of miR-200, which contributes to breast tumorigenesis [126]. In a recent study, lncRNA LSINCT5 has increased cell motility by sponging miR-30a from the Wnt/β-catenin pathway. TCF4 and c-Myc expression was likewise downregulated in cells in which LSINCT5 was knocked out, resulting in decreased proliferation, motility, and EMT [127]. Furthermore, by promoting the miR-185-3p/E2F1/Nanog axis, lncRNA LINC00511 leads to BC tumorigenesis and stemness [128]. Similarly, In TNBC models, LINC01133 acts as a direct mediator of the MSC-triggered miR-199a-FOXP2 axis, enhancing phenotypic and growth characteristics of CSC-like cells [129]. Lastly, evidence supports that the lncRNA, HOTTIP (HOXA transcript at the distal tip), is upregulated in a variety of malignancies, including BC. HOTTIP is also involved in several biological activities, such as maintaining stem cell viability. As a molecular sponge for miR-148a-3p, HOTTIP controls the CSC-like features of BrCSCs, enhancing WNT1 transcription and providing a novel therapeutic target for BC patients [130] (Fig. 3).

Different signaling pathways in which lncRNAs regulate BrCSCs through ceRNAs mechanism (miRNAs gene inhibition) which increases the mamosphere development, EMT and cancer stemness
Table 2 provides additional examples of lncRNA that regulate BrCSCs through ceRNAs mechanism.
Overall, in terms of lncRNA aberrant expression and the causative factors, lncRNAs may be useful prognostic biomarkers in BrCSCs. Additionally, lncRNAs have the potential to be targeted to reverse the process of carcinogenesis, making them valuable therapeutic targets for treatment of cancer.
Since most of lncRNAs/miRNAs axes (ceRNA axes) have been only assessed in one paper, there is no way to compare the results of studies. However, it is possible that a single lncRNA acts as a molecular sponge for more than one miRNA. In fact, it is possible to identify several lncRNAs/miRNAs axes in which some of the contributing ncRNAs have common roles.
LncRNAs participate in different pathways for modulating BrCSCs
Because lncRNAs are highly abundant in BrCSCs, they can play a variety of regulatory functions. In recent years, the importance of lncRNAs has been recognized in BrCSCs, with their functional significance and clinical consequences being highlighted recently. Here, the regulatory functions of lncRNAs on BrCSCs are discussed in terms of their many modes of action.
LncRNAs regulate BrCSCs through HIF signaling pathway
As it turns out, hypoxia influences a specific subset of long noncoding RNAs. Hypoxia-responsive lncRNAs (HRLs) such as NORAD, LncHIFCAR, AC020978, KB-1980E6.3, RAB11B-AS1 may have a role in cancer cell survival and disease progression [101, 136, 137]. HRLs may be classified into two categories: those dependent on hypoxia-inducible factors (HIF) and those not dependent on HIF. Specifically, the hypoxia response element (HRE), located in the promoter regions of HRLs, may directly bind to HIF in hypoxic tumor microenvironments and control their expression by modulating their transcription. Conversely, the HRLs that are excessively expressed may trigger a tumor-specific molecular profile and contribute to the development of tumor characteristics [138]. The molecular properties of HRLs in BC, however, remain unsolved.
Recently, in an in vitro and in vivo studies undertaken in a hypoxic condition, Zhu and colleagues found that the KB-1980E6.3 enhanced BrCSC self-renewal and tumorigenesis [139].
They observed that HIF-1α might interact directly with the HRE in the KB-1980E6.3 promoter to control its transcription activities in hypoxia conditions. Furthermore, Because of its capacity to bind to IGF2BP1, lncRNA KB-1980E6.3 was critical for BrCSC stemness because it increases the stability of c-Myc mRNA and enhances it BC cells' responsiveness to hypoxia [139].
LncRNAs regulate BrCSCs through Hh, Wnt, Hippo signaling pathways
The estrogen receptor (ER) regulates the Hh, Wnt, and Hippo pathways. The dysregulation of these pathways is linked to the fundamental hallmarks of BrCSCs, such as self-renewal, cancer resistance, metastasis, and recurrence [140, 141]. In addition, components in the tumor microenvironment, including hypoxia, angiogenesis, and inflammatory responses, have a main role in modulating BrCSCs [142, 143]. The Hh pathway components (PTCH1, Gli1, and Gli2) are abundantly expressed in normal mammary stem/progenitor cells and stimulated in BrCSCs. For the self-renewal of human BrCSCs, the Hh pathway and Bmi-1 are found to play critical roles highlighting the relevance of the Hh pathway and Bmi-1 in the controlling CSCs, hence applying techniques focused on blocking both pathways provide a helpful therapeutic strategy [144]. Chemotherapy has been demonstrated to be ineffective against BrCSCs, and the stemness of CSCs can be preserved through the Hh pathway. Using MCF-7 cells from BC patients as a model, Miao and his team discovered that the Hh signaling pathway is involved in drug responsiveness [145]. The Hh ligand secreted by CSCs can control the response of cancer-associated fibroblasts (CAFs). Indeed, the CAFs provide elements that encourage CSC growth and self-renewal [146]. Thus, inhibiting this signaling system might be a unique treatment method for BC therapy. Furthermore, SMO protein, transmembrane receptors (PTCH 1), (GLI 1–3), and extracellular Hh ligands are components of this complicated signaling cascade. Also, the Hh ligands activate signal transduction events in the cell, which interact with PTCH. To activate the transcription factor and allow it entry into the cell nucleus, the Hh ligand binds to PTCH, causing changes in its physical conformation and removing the inhibition of SMO. This results in increased cell function, growth, and differentiation [147, 148]. According to recent data, there is a clear relationship between lncRNAs and the cellular proliferation of BrCSCs through different signaling pathways (Table 2). For instance, LINC00617 increases the fraction of a phenotypic stem cell CD44( +)/CD24(-) subpopulation, enhancing BC metastasis. LINC00617 promotes EMT via rising the SOX2 expression in BC cells [50, 149]. Furthermore, lncRNA XIST expression in BC patients is linked to a higher risk of brain metastases. Reduced XIST expression promotes tumor cell stemness via stimulating EMT transition and activating c-Met through moesin MSN-mediated protein stabilization. In mouse mammary glands, knocking down XIST promotes the formation of primary tumors and brain metastases [150]. In addition, there is a clear relationship between EMT and the cellular proliferation of tumor cells. lncRNA-Hh, a lncRNA related to the Hh pathway, has been shown to directly bind GAS1 (a hedgehog signaling enhancer) and promote the activities of SOX2 and OCT4 [59]. Furthermore, lncROPM is a marker for BC patients' malignant grade, stage, and poor prognosis. Studies on the function gain and loss of lncROPM have shown that it is necessary to regulate BrCSC characteristics in vitro and in vivo studies. Through directly binding to the 3'-UTR of PLA2G16 (Group XVI phospholipase A2), lncROPM controls the expression of PLA2G16 by increasing the mRNA stability. Higher PLA2G16 enhances phospholipid metabolism and the generation of free fatty acids [151], particularly arachidonic acid, in BrCSCs, promoting PI3K/AKT, Wnt/-catenin, and Hippo/YAP pathways and ultimately contributing to the regulation of BrCSC stemness [152]. An animal investigation in BrCSCs found that H19 increased stemness via sponging Let-7c and activating its ESR1 and Wnt pathway targets [153]. Additionally, inhibition of symmetric division of BrCSCs by H19 or Let-7c miRNA increases non-CSCs. lncRNA-Hh is linked with the Shh-GLI1 pathway and is transcriptionally controlled by Twist, directly targets GAS1 to enhance Hh signaling activation. Increased GLI1 expression and increased SOX2 and OCT4 transcription are all associated with the activation of Hh, which is thought to play a regulatory function in CSC development [154]. Additionally, lncRNA-Hh stimulates the Hh pathway protein Hh to increase OCT4 and SOX2 expression for BrCSC homeostasis [59]. Similarly, Han et al. showed that lncRNA HOTTIP modulates the miR-148a-3p/WNT1 axis, which allows BrCSCs retain their CSC-like features and promote BC development [130]. In contrast, overexpression of LINC00968 in BC cells reduces the development of resistance by decreasing the stimulation of the Wnt2-β-catenin pathway by suppressing Wnt2 [155] (Fig. 4).

LncRNAs regulate BrCSCs by promoting EMT, β-catenin, and survival genes, which enhances proliferation, EMT, metastasis, and cancer stemness
Altogether, these data imply that the lncRNAs (lncRNA-Hh, H19, LINC00617, lncROPM, XIST, PLA2G16) play a critical role in the regulation of CSCs and are associated with the invasive and migration characteristics of BC cells. They showed that these molecules might be useful in the prognostic and diagnostic method in metastatic BC patients.
LncRNAs involved in cancer stemness and therapeutic resistance
Resistance to therapy is a hallmark of aggressive tumors, which leads to reduced survival and a high mortality rate. Patients with advanced or metastatic BC are often treated with chemotherapy, but some of patients acquire resistance to subsequent treatments [156]. Beside chemotherapy, hormonal therapies like tamoxifen are usually given in BC patients with estrogen-positive tumors. Loss of ER altered expression of co-regulatory proteins (CRPs), as well as increases in the number of ER-negative CSCs in the ER+ tumor, and lncRNAs mediates and contributes to tamoxifen resistance in BC [157,158,159].
Although cancer drugs effectively eliminate or kill the majority of cancerous cells, CSCs are able to evade their effects via a variety of biological strategies [160]. CSCs with slow division or quiescence, for example, are less affected by chemotherapy that targets rapidly dividing cells. Another critical resistance mechanism is aldehyde dehydrogenase (ALDH) overexpression in CSCs [161]. Enzymes in this family catalyze the oxidation of aldehydes to carboxylic acids, which is a step in the detoxification process for both internal and external aldehydes [162].
Induced iPSCs were shown to retain their pluripotency via lncRNA Peblr20, which suggests that lncRNAs may play a role in retaining cancer stemness [163]. EMT-associated stemness is controlled by lncRNAs, which have emerged as new participants in BrCSC stemness [124]. As an example, numerous lncRNAs, including LINC01638 [164], lncRNA RP1-506.5 [165], LINC-ZNF469-3 [166] and LINC-ROR [167, 168], have been shown to protect or promote EMT properties and CSC-properties of BC cells (Fig. 3). A recent study by Tang et al. found a direct link between lncRNA-regulated EMT and BC therapy resistance. According to their findings, DCST1-AS1 interacts effectively with ANXA1 to increase EMT and improve resistance in BC cells to doxorubicin and paclitaxel [169, 170].
In addition, it was determined that numerous pluripotency regulators, such as LIN28, SOX2, OCT3/4, KLF4, and CSC biomarkers, such as ALDH1A3, can stimulate stemness in BrCSCs [171,172,173]. By interacting with CSC-associated genes, lncRNAs play crucial role in BrCSC formation and can be used as a marker and applied for clinical translation. For instance, when CCAT2 lncRNA is expressed in breast cancer stem cells during targeted knockdown, it increases the levels of Nanog, OCT4, and KLF4, as well as the ALDH + CSC population which can serve as a marker for the presence cancer stem cells [174]. In particular, deregulation and circulating lncRNAs in physiological fluids of cancer patients may serve as useful diagnostic and prognostic markers in the treatment of the disease and can guide clinical therapy. For example, urine PCA3 (lncRNA PCA3) has been authorized by the FDA as a urine marker for prostate cancer due to its high sensitivity and specificity over PSA (prostate-specific antigen) [175, 176].
In addition, lncRNAs are reliable indicators of the patient's prognosis and the risk of their tumor recurrence. In clinical studies of breast cancer patients, hormone negativity, tumor size, and nodal status are all linked to high H19 expression. Disease-free survival is considerably worse in patients who has H19 expression than in other patients [66]. Furthermore, high expression of HOTAIR in BC patients is strongly correlated with lymph node metastasis, recurrent and poor prognosis of breast cancer [77, 177].
Likewise, many lncRNAs regulate the expression of pluripotency factors and CSC markers by functioning as ceRNAs that compete with the limited number of miRNAs (Fig. 3). According to Peng et al., the lncRNA H19 in BrCSCs acts as a sponge for Let-7, resulting in the increased expression of Let-7's target LIN28, which supports the maintenance of BrCSCs [67]. They found that LIN28 reduces Let-7 expression and activation, ultimately suppressing H19 transcription in BrCSCs. Treatment resistance may be restored by disruption of the H19-Let-7/LIN28 pathway, which is responsible in part for stemness of BrCSCs, and it presents a new strategy for treating BC. Similarly, the lncRNA LINC01133 induced by MSCs promotes BrCSC's morphological and growth features via directly regulating KLF4 [129].
NRAD1 (LINC00284) is notable for being the first lncRNA to be triggered by a CSC marker. In addition, it was shown that retinoic acid, a byproduct of ALDH1A3, has a beneficial effect on the expression of NRAD1, which promotes cell survival and increases BrCSCs [71]. Interestingly, the Knockdown of ALDH1A3 reduced the activity of ALDH, and ALDH1A1 knockdown suppressed metastatic features and therapy resistance in human BC cells [178].
While long noncoding RNAs play a significant role in maintaining BrCSCs and the potential development of intrinsic therapeutic resistance, further research into the underlying processes and potential clinical applications is needed.
Challenges and strategies to overcome lncRNAs therapeutic resistance in BrCSCs
Challenges to overcoming lncRNA therapeutic resistance
Despite advances in cancer treatment, drug resistance continues to be a significant challenge for people with BC. A comprehensive evaluation of off-target effects, possible toxicity, and drug delivery/precision targeting is necessary for the lncRNA silencing approaches to be considered effective therapeutic strategies. Many lncRNA transcript variations also present a considerable challenge for developing techniques to target molecules. It is possible that not all transcript variations will be targeted by the lncRNA silencing technique, which may reduce the treatment's effectiveness if that variant is functional. Moreover, CRISPR/Cas9 may have challenges or limited utility for the knockout of non-coding genes [179, 180]. Despite their equivalent molecular weight, there are no open reading frames (ORPs) in lncRNAs, unlike protein-coding genes. Furthermore, the roles of most lncRNAs remain unknown, making the creation of efficient medicines and delivery methods much more difficult. A comparison study in clinical trials would be required to determine whether or not lncRNAs are better targets than protein-coding genes [181].
Strategies for reducing therapeutic resistance to lncRNAs
As previously stated, CSCs have responsibility for drug resistance and recurrence, all of which have an impact on anticancer therapeutic potential. The use of lncRNAs or related pathways to target CSCs as a possible therapy for cancer is a novel approach [182]. In BC, down-regulation of the oncogenic lncRNA CCAT1 is linked to an increase in radiosensitivity through altering the miR-148b, which is responsible for cancer progression [183]. Interestingly, Dendrosomal curcumin (DNC), a natural chemical, can be used to treat cancer by reducing the expression of tumor suppressor genes Tusc7 and GAS5 lncRNA in BC cells MCF7, MDA-MB231, and SKBR3. Furthermore, Esmatabadi and his team found that down-regulating GAS5 in BC cells can reduce many characteristics of DNC's anti-cancer activities. This suggests that combining DNC with GAS5 over-expression could be a clinically effective tool for drug-resistance in BC cells [184].
Recent study reveals that MEG3 interacts directly with miRNA-421 to regulate a variety of CSC characteristics, including self-renewal and invading abilities [185]. However, as a result of the methylation that occurs in its promoter region, the expression of the tumor suppressor lncRNA MEG3 is downregulated in cancer cells. A new nanotechnology-based preparation of natural curcumin known as dendrosomal curcumin increased the expression of MEG3 by down-regulating DNA methyltransferase (1A, 3A, and 3B) expression through promoting of miR-29a and miR-185 [186].
Chemoprevention is the focus of new strategies to enhance clinical outcomes and reduce the toxicity of anticancer medications. In the case of advanced ovarian cancer and BC, for example, Oxaliplatin (Oxa) is a platinum medication of the third generation that is used either alone or in combination with other treatments [187, 188]. However, several mechanisms contribute to resistance to Oxa, including DNA damage repair, inhibition of apoptosis, deregulation of signaling pathways, and increased detoxifying efficiency. Combination therapy has been proposed as an emerging approach to solving this problem [189]. It was found that DNC alone or in combination with Oxa has synergistically downregulated several oncogenic lncRNAs such as GAS5, MALAT1, FAL1, ANRIL, ABO73614, CCAT2, LSINCT5 in different types of cancers such as BC, NSCLC, and ovarian cancer through suppressing cell proliferation, prompting cell death, and reducing therapeutic resistance [184, 188, 190, 191]. They also found that DNC or curcumin combined with anti-cancer drugs had a more significant inhibitory effect than monotherapy. Curcumin reduces the lncRNA H19-induced EMT [192].
In addition, CRISPR interference (CRISPRi) or knockout (KO), a technology that may be applied to any genomic site, can be used to inhibit the activity of lncRNAs, which are crucial for the survival or self-renewal of cancer cells [193]. Complete excision of the whole gene, excision of the promoter and transcriptional start point, removing exon/exon junctions, or excision of the transcriptional terminal site are all strategies for lncRNAs knocking out [193]. For instance, CRISPR/Cas9 technology was used by Peng and colleagues to modify LncROR expression in BC cell lines. Researchers found that lncROR increased estrogen production through this new approach and triggered the MAPK/ERK axis in BC. CRISPR/cas9 technique was used in this study to investigate lncRNA loss of function and gives evidence for lncROR might be a possible target in ER+ BC patients [194]. Thus, it is a promising technique to reduce drug resistance, such as tamoxifen resistance of ER+ BC, and can be used as a therapeutic strategy in BC patients. In addition, lncRNA BC200 functions as an oncogene and has an important role in cancer proliferation and drug resistance [195]. Singh and colleagues published a notable study elucidating that oncogenic LncBC200 is elevated in BC [196]. The amount of LncBC200 in breast tumor tissues is greater in ER+ tumors than in ER− tumors. They used CRISPR/Cas9 technology to knock off LncBC200 expression to understand more about the function of ER-regulated LncBC200 expression. According to the findings LncBC200 was shown to have a significant role in the development of cancer. Ultimately, oncogenic lncAK023948 which is essential for cancer cell survival has been used as a target for CRISPR/Cas9 to reduce tumor growth in different types of BC cells [197]. These findings offer a scientific basis for the concept that lncRNAs play an important part in the progression of cancer and could be used as therapeutic targets.
Conclusions
It is predicted that therapies targeting CSC will prevent cancer metastasis and recurrence, since these cells have unique features that enhance anti-cancer therapy resistance. For this reason, identifying molecules that control their function is important in practice. The activity and growth of CSCs have been demonstrated to be influenced by lncRNAs. The effect of these regulatory transcripts on CSCs were mainly studied in BC. The activity and growth of CSCs were demonstrated to be influenced by lncRNAs. The effect of these regulatory transcripts on CSCs has mainly been studied in BC. LncRNAs may help BC patients not only as diagnostic indicators for identifying patient stage and prognosis but also as targets for precision treating cancer, thereby offering new pathways for eliminating CSCs. However, because of the complexity of CSC biology and the interrelationships among its various subtypes and their various cellular activities and signaling processes in health and illness, it is challenging to develop effective therapies that specifically target these cells. LncRNA-based research will help us achieve a new knowledge of the physiology of cancer stem cells. Furthermore, new studies are necessary to uncover and establish the wide range of regulatory mechanisms and the complexity of pathways that lncRNAs may control. Despite its limitations, understanding CSC-lncRNA interactions may usher in a new era of cancer therapy associated with lower drug resistance and increased anti-metastatic efficacy, thereby improving patient prognosis.
Availability of data and materials
Not applicable.
Abbreviations
- lncRNAs:
-
Long non-coding RNAs
- CSCs:
-
Cancer stem cells
- BrCSCs:
-
Breast Cancer stem cells
- AML:
-
Acute myeloid leukemia
- OCT4:
-
Octamer-binding transcription factor 4
- Sox2:
-
Sex determining region Y-box 2
- KLF4:
-
Kruppel Like Factor 4
- RNA:
-
Ribonucleic acid
- iPSCs:
-
Induced Pluripotent stem cells
- ESCs:
-
Embryonic stem cells
- MALAT1:
-
Metastasis-associated lung carcinoma transcript 1
- HOTAIR:
-
Homeobox antisense transcript antisense RNA
- EMT:
-
Epithelial-to-mesenchymal transition
- NR2F1-AS1:
-
NR2F1 Antisense RNA 1
- Xist:
-
X-inactive specific transcript
- MT1JP:
-
Metallothionein 1 J
- RNPs:
-
Ribonucleoproteins
- HRE:
-
Hypoxia response element
- HIF:
-
Hypoxia-inducible factors
- CAFs:
-
Cancer-associated fibroblasts
- PLA2G1:
-
Phospholipase A2, group XVI
- Hh:
-
Hedgehog signaling
- ER:
-
Estrogen receptor
- CRPs:
-
Co-regulatory proteins
- ORPs:
-
Open reading frames
- Oxa:
-
Oxaliplatin
- CRISPRi:
-
CRISPR interference
- KO:
-
Knockout
- CD 44:
-
Transmembrane adhesion glycoprotein
References
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–49.
Sharma GN, Dave R, Sanadya J, Sharma P, Sharma KK. Various types and management of breast cancer: an overview. J Adv Pharm Technol Res. 2010;1(2):109–26.
Bukowski K, Kciuk M, Kontek R. Mechanisms of multidrug resistance in cancer chemotherapy. Int J Mol Sci. 2020;21(9):3233.
Rycaj K, Tang DG. Cell-of-origin of cancer versus cancer stem cells: assays and interpretations. Cancer Res. 2015;75(19):4003–11.
Taurin S, Alkhalifa H. Breast cancers, mammary stem cells, and cancer stem cells, characteristics, and hypotheses. Neoplasia. 2020;22(12):663–78.
Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang J, et al. Targeting cancer stem cell pathways for cancer therapy. Signal Transduct Target Ther. 2020;5(1):8.
Wang C, Yan FH, Zhang JJ, Huang H, Cui QS, Dong W, et al. OV6(+) cancer stem cells drive esophageal squamous cell carcinoma progression through ATG7-dependent β-catenin stabilization. Cancer Lett. 2017;391:100–13.
Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat Rev Cancer. 2008;8(10):755–68.
Rasheed ZA, Kowalski J, Smith BD, Matsui W. Concise review: emerging concepts in clinical targeting of cancer stem cells. Stem Cells. 2011;29(6):883–7.
Djebali S, Davis CA, Merkel A, Dobin A, Lassmann T, Mortazavi A, et al. Landscape of transcription in human cells. Nature. 2012;489(7414):101–8.
Costa FF. Non-coding RNAs: meet thy masters. BioEssays. 2010;32(7):599–608.
Mattick JS. Non-coding RNAs: the architects of eukaryotic complexity. EMBO Rep. 2001;2(11):986–91.
Hartford CCR, Lal A. When long noncoding becomes protein coding. Mol Cell Biol. 2020. https://doi.org/10.1128/MCB.00528-19.
Zhang P, Wu W, Chen Q, Chen M. Non-Coding RNAs and their integrated networks. J Integr Bioinform. 2019;16(3):20190027.
Kumar SM, Liu S, Lu H, Zhang H, Zhang PJ, Gimotty PA, et al. Acquired cancer stem cell phenotypes through Oct4-mediated dedifferentiation. Oncogene. 2012;31(47):4898–911.
Herreros-Villanueva M, Zhang JS, Koenig A, Abel EV, Smyrk TC, Bamlet WR, et al. SOX2 promotes dedifferentiation and imparts stem cell-like features to pancreatic cancer cells. Oncogenesis. 2013;2(8): e61.
Yu F, Li J, Chen H, Fu J, Ray S, Huang S, et al. Kruppel-like factor 4 (KLF4) is required for maintenance of breast cancer stem cells and for cell migration and invasion. Oncogene. 2011;30(18):2161–72.
Eades G, Zhang Y-S, Li Q-L, Xia J-X, Yao Y, Zhou Q. Long non-coding RNAs in stem cells and cancer. World J Clin Oncol. 2014;5(2):134–41.
Zhang Q, Zhuang J, Deng Y, Yang L, Cao W, Chen W, et al. miR34a/GOLPH3 Axis abrogates urothelial bladder cancer chemoresistance via reduced cancer stemness. Theranostics. 2017;7(19):4777–90.
Wang L, Bu P, Ai Y, Srinivasan T, Chen HJ, Xiang K, et al. A long non-coding RNA targets microRNA miR-34a to regulate colon cancer stem cell asymmetric division. Elife. 2016. https://doi.org/10.7554/eLife.14620.
Jarroux J, Morillon A, Pinskaya M. History, discovery, and classification of lncRNAs. Adv Exp Med Biol. 2017;1008:1–46.
Smolle MA, Pichler M. The role of long non-coding RNAs in osteosarcoma. Non-Coding RNA. 2018;4(1):7.
Frankish A, Diekhans M, Jungreis I, Lagarde J, Loveland Jane E, Mudge JM, et al. GENCODE 2021. Nucleic Acids Res. 2020;49(D1):D916–23.
Harrow J, Frankish A, Gonzalez JM, Tapanari E, Diekhans M, Kokocinski F, et al. GENCODE: the reference human genome annotation for The ENCODE Project. Genome Res. 2012;22(9):1760–74.
Statello L, Guo CJ, Chen LL, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118.
Pisignano G, Ladomery M. Post-Transcriptional Regulation through Long Non-Coding RNAs (lncRNAs). Noncoding RNA. 2021;7(2):29.
Garratt H, Ashburn R, Sopić M, Nogara A, Caporali A, Mitić T. Long Non-Coding RNA regulation of epigenetics in vascular cells. Noncoding RNA. 2021;7(4):62.
Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–14.
Mercer TR, Dinger ME, Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009;10(3):155–9.
Mercer TR, Mattick JS. Structure and function of long noncoding RNAs in epigenetic regulation. Nat Struct Mol Biol. 2013;20(3):300–7.
Vance KW, Ponting CP. Transcriptional regulatory functions of nuclear long noncoding RNAs. Trends Genet. 2014;30(8):348–55.
Ahmad P, Bensaoud C, Mekki I, Rehman MU, Kotsyfakis M. Long Non-Coding RNAs and their potential roles in the vector–host–pathogen triad. Life. 2021;11(1):56.
Fang Y, Fullwood MJ. Roles, functions, and mechanisms of long non-coding RNAs in cancer. Genom Proteomics Bioinform. 2016;14(1):42–54.
Li Y, Wang Z, Shi H, Li H, Li L, Fang R, et al. HBXIP and LSD1 Scaffolded by lncRNA Hotair Mediate Transcriptional Activation by c-Myc. Cancer Res. 2016;76(2):293–304.
Fan J, Xing Y, Wen X, Jia R, Ni H, He J, et al. Long non-coding RNA ROR decoys gene-specific histone methylation to promote tumorigenesis. Genome Biol. 2015;16(1):139.
Perez CAG, Adachi S, Nong QD, Adhitama N, Matsuura T, Natsume T, et al. Sense-overlapping lncRNA as a decoy of translational repressor protein for dimorphic gene expression. PLoS Genet. 2021;17(7): e1009683.
Kotake Y, Nakagawa T, Kitagawa K, Suzuki S, Liu N, Kitagawa M, et al. Long non-coding RNA ANRIL is required for the PRC2 recruitment to and silencing of p15(INK4B) tumor suppressor gene. Oncogene. 2011;30(16):1956–62.
Ghafouri-Fard S, Tamizkar KH, Hussen BM, Taheri M. An update on the role of long non-coding RNAs in the pathogenesis of breast cancer. Pathol Res Practice. 2021;219: 153373.
Liu L, Zhang Y, Lu J. The roles of long noncoding RNAs in breast cancer metastasis. Cell Death Dis. 2020;11(9):749.
O’Flaherty JD, Barr M, Fennell D, Richard D, Reynolds J, O’Leary J, et al. The cancer stem-cell hypothesis: its emerging role in lung cancer biology and its relevance for future therapy. J Thorac Oncol. 2012;7(12):1880–90.
Yu Z, Pestell TG, Lisanti MP, Pestell RG. Cancer stem cells. Int J Biochem Cell Biol. 2012;44(12):2144–51.
Lapidot T, Sirard C, Vormoor J, Murdoch B, Hoang T, Caceres-Cortes J, et al. A cell initiating human acute myeloid leukaemia after transplantation into SCID mice. Nature. 1994;367(6464):645–8.
Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA. 2003;100(7):3983–8.
Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, et al. Identification of human brain tumour initiating cells. Nature. 2004;432(7015):396–401.
Rosen JM, Jordan CT. The increasing complexity of the cancer stem cell paradigm. Science. 2009;324(5935):1670–3.
Hussen BM, Shoorei H, Mohaqiq M, Dinger ME, Hidayat HJ, Taheri M, et al. The Impact of Non-coding RNAs in the epithelial to mesenchymal transition. Front Mol Biosci. 2021;8: 665199.
Ghafouri-Fard S, Glassy MC, Abak A, Hussen BM, Niazi V, Taheri M. The interaction between miRNAs/lncRNAs and Notch pathway in human disorders. Biomed Pharmacother. 2021;138: 111496.
Saliani M, Mirzaiebadizi A, Mosaddeghzadeh N, Ahmadian MR. RHO GTPase-related long noncoding RNAs in human cancers. Cancers (Basel). 2021;13(21):5386.
Ghafouri-Fard S, Abak A, Fattahi F, Hussen BM, Bahroudi Z, Shoorei H, et al. The interaction between miRNAs/lncRNAs and nuclear factor-κB (NF-κB) in human disorders. Biomed Pharmacother. 2021;138: 111519.
Li H, Zhu L, Xu L, Qin K, Liu C, Yu Y, et al. Long noncoding RNA linc00617 exhibits oncogenic activity in breast cancer. Mol Carcinog. 2017;56(1):3–17.
Castro-Oropeza R, Melendez-Zajgla J, Maldonado V, Vazquez-Santillan K. The emerging role of lncRNAs in the regulation of cancer stem cells. Cell Oncol (Dordr). 2018;41(6):585–603.
Ayob AZ, Ramasamy TS. Cancer stem cells as key drivers of tumour progression. J Biomed Sci. 2018;25(1):20.
Kabakov AE, Yakimova AO. Hypoxia-induced cancer cell responses driving radioresistance of hypoxic tumors: approaches to targeting and radiosensitizing. Cancers. 2021;13(5):1102.
Schöning JP, Monteiro M, Gu W. Drug resistance and cancer stem cells: the shared but distinct roles of hypoxia-inducible factors HIF1α and HIF2α. Clin Exp Pharmacol Physiol. 2017;44(2):153–61.
Emami Nejad A, Najafgholian S, Rostami A, Sistani A, Shojaeifar S, Esparvarinha M, et al. The role of hypoxia in the tumor microenvironment and development of cancer stem cell: a novel approach to developing treatment. Cancer Cell Int. 2021;21(1):62.
Owens T, Naylor M. Breast cancer stem cells. Front Physiol. 2013. https://doi.org/10.3389/fphys.2013.00225.
Hussen BM, Azimi T, Abak A, Hidayat HJ, Taheri M, Ghafouri-Fard S. Role of lncRNA BANCR in human cancers: an updated review. Front Cell Dev Biol. 2021. https://doi.org/10.3389/fcell.2021.689992.
Schwerdtfeger M, Desiderio V, Kobold S, Regad T, Zappavigna S, Caraglia M. Long non-coding RNAs in cancer stem cells. Transl Oncol. 2021;14(8): 101134.
Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du YE, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem Cells. 2016;34(1):55–66.
Ye X, Tam WL, Shibue T, Kaygusuz Y, Reinhardt F, Ng Eaton E, et al. Distinct EMT programs control normal mammary stem cells and tumour-initiating cells. Nature. 2015;525(7568):256–60.
Deng J, Yang M, Jiang R, An N, Wang X, Liu B. Long Non-Coding RNA HOTAIR regulates the proliferation, self-renewal capacity, tumor formation and migration of the Cancer Stem-Like Cell (CSC) subpopulation enriched from breast cancer cells. PLoS ONE. 2017;12(1): e0170860.
Jin C, Yan B, Lu Q, Lin Y, Ma L. Reciprocal regulation of Hsa-miR-1 and long noncoding RNA MALAT1 promotes triple-negative breast cancer development. Tumour Biol. 2016;37(6):7383–94.
Latorre E, Carelli S, Raimondi I, D’Agostino V, Castiglioni I, Zucal C, et al. The ribonucleic complex HuR-MALAT1 represses CD133 expression and suppresses epithelial-mesenchymal transition in breast cancer. Cancer Res. 2016;76(9):2626–36.
Samir A, Tawab RA, El Tayebi HM. Long non-coding RNAs XIST and MALAT1 hijack the PD-L1 regulatory signaling pathway in breast cancer subtypes. Oncol Lett. 2021;22(2):593.
Zhang K-J, Tan X-L, Guo L. The long non-coding RNA DANCR regulates the inflammatory phenotype of breast cancer cells and promotes breast cancer progression via EZH2-dependent suppression of SOCS3 transcription. Mol Oncol. 2020;14(2):309–28.
Shima H, Kida K, Adachi S, Yamada A, Sugae S, Narui K, et al. Lnc RNA H19 is associated with poor prognosis in breast cancer patients and promotes cancer stemness. Breast Cancer Res Treat. 2018;170(3):507–16.
Peng F, Li TT, Wang KL, Xiao GQ, Wang JH, Zhao HD, et al. H19/let-7/LIN28 reciprocal negative regulatory circuit promotes breast cancer stem cell maintenance. Cell Death Dis. 2017;8(1): e2569.
Liu Y, Zhang P, Wu Q, Fang H, Wang Y, Xiao Y, et al. Long non-coding RNA NR2F1-AS1 induces breast cancer lung metastatic dormancy by regulating NR2F1 and ΔNp63. Nat Commun. 2021;12(1):5232.
Li X, Wang S, Li Z, Long X, Guo Z, Zhang G, et al. The lncRNA NEAT1 facilitates cell growth and invasion via the miR-211/HMGA2 axis in breast cancer. Int J Biol Macromol. 2017;105(Pt 1):346–53.
Shin VY, Chen J, Cheuk IW-Y, Siu M-T, Ho C-W, Wang X, et al. Long non-coding RNA NEAT1 confers oncogenic role in triple-negative breast cancer through modulating chemoresistance and cancer stemness. Cell Death Dis. 2019;10(4):270.
Vidovic D, Huynh TT, Konda P, Dean C, Cruickshank BM, Sultan M, et al. ALDH1A3-regulated long non-coding RNA NRAD1 is a potential novel target for triple-negative breast tumors and cancer stem cells. Cell Death Differ. 2020;27(1):363–78.
Wang Y, Xu Z, Jiang J, Xu C, Kang J, Xiao L, et al. Endogenous miRNA sponge lincRNA-RoR regulates Oct4, Nanog, and Sox2 in human embryonic stem cell self-renewal. Dev Cell. 2013;25(1):69–80.
Zhou Q, Guo J, Huang W, Yu X, Xu C, Long X. Linc-ROR promotes the progression of breast cancer and decreases the sensitivity to rapamycin through miR-194-3p targeting MECP2. Mol Oncol. 2020;14(9):2231–50.
Han C, Li X, Fan Q, Liu G, Yin J. CCAT1 promotes triple-negative breast cancer progression by suppressing miR-218/ZFX signaling. Aging (Albany NY). 2019;11(14):4858–75.
Jia X, Shi L, Wang X, Luo L, Ling L, Yin J, et al. KLF5 regulated lncRNA RP1 promotes the growth and metastasis of breast cancer via repressing p27kip1 translation. Cell Death Dis. 2019;10(5):373.
Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du Y-e, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem cells (Dayton, Ohio). 2016;34(1):55–66.
Hussen BM, Hidayat HJ, Ghafouri-Fard S. Identification of expression of CCND1-related lncRNAs in breast cancer. Pathol Res Practice. 2022;236: 154009.
Rakhshan A, Gholipour M, Hussen BM, Taheri M, Eslami S, Ghafouri-Fard S, et al. Expression analysis of CDKN2C-related lncRNAs in breast cancer. Human Gene. 2022;33: 201070.
Peng F, Wang JH, Fan WJ, Meng YT, Li MM, Li TT, et al. Glycolysis gatekeeper PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene. 2018;37(8):1062–74.
Song Z, Zhang X, Lin Y, Wei Y, Liang S, Dong C. LINC01133 inhibits breast cancer invasion and metastasis by negatively regulating SOX4 expression through EZH2. J Cell Mol Med. 2019;23(11):7554–65.
Deshpande S, Nair S, Balasinor NH. Chapter 2 - Genomic imprinting. In: Tollefsbol T, editor. Epigenetics and Reproductive Health. Cambridge: Academic Press; 2021. p. 27–46.
Kan RL, Chen J, Sallam T. Crosstalk between epitranscriptomic and epigenetic mechanisms in gene regulation. Trends Genet. 2022;38(2):182–93.
Morlando M, Fatica A. Alteration of epigenetic regulation by long Noncoding RNAs in cancer. Int J Mol Sci. 2018;19(2):570.
Pang B, Wang Q, Ning S, Wu J, Zhang X, Chen Y, et al. Landscape of tumor suppressor long noncoding RNAs in breast cancer. J Exp Clin Cancer Res. 2019;38(1):79.
Li J, Sasane M, Zhang J, Zhao J, Ricculli ML, Yao Z, et al. Is time to progression associated with post-progression survival in previously treated metastatic non-small cell lung cancer with BRAF V600E mutation? A secondary analysis of phase II clinical trial data. BMJ Open. 2018;8(8): e021642.
Xu S, Liu H, Wan L, Zhang W, Wang Q, Zhang S, et al. The MS-lincRNA landscape reveals a novel lincRNA BCLIN25 that contributes to tumorigenesis by upregulating ERBB2 expression via epigenetic modification and RNA-RNA interactions in breast cancer. Cell Death Dis. 2019;10(12):920.
Calanca N, Paschoal A, Munhoz É, Galindo L, Barbosa B, Caldeira J, et al. The long non-coding RNA ANRASSF1 in the regulation of alternative protein-coding transcripts RASSF1A and RASSF1C in human breast cancer cells: implications to epigenetic therapy. Epigenetics. 2019;14:741.
MacDonald WA, Mann MRW. Long noncoding RNA functionality in imprinted domain regulation. PLoS Genet. 2020;16(8):e1008930.
Ding X, Li Y, Lü J, Zhao Q, Guo Y, Lu Z, et al. piRNA-823 is involved in cancer stem cell regulation through altering DNA methylation in association with luminal breast cancer. Front Cell Dev Biol. 2021;9:641052.
Han YJ, Boatman SM, Zhang J, Du XC, Yeh AC, Zheng Y, et al. LncRNA BLAT1 is upregulated in basal-like breast cancer through epigenetic modifications. Sci Rep. 2018;8(1):15572.
Calanca N, Paschoal AP, Munhoz ÉP, Galindo LT, Barbosa BM, Caldeira JRF, et al. The long non-coding RNA ANRASSF1 in the regulation of alternative protein-coding transcripts RASSF1A and RASSF1C in human breast cancer cells: implications to epigenetic therapy. Epigenetics. 2019;14(8):741–50.
Pawłowska E, Szczepanska J, Blasiak J. The long noncoding RNA HOTAIR in breast cancer: does autophagy play a role? Int J Mol Sci. 2017;18(11):2317.
Sun X, Du P, Yuan W, Du Z, Yu M, Yu X, et al. Long non-coding RNA HOTAIR regulates cyclin J via inhibition of microRNA-205 expression in bladder cancer. Cell Death Dis. 2015;6(10): e1907.
Zhang H, Cai K, Wang J, Wang X, Cheng K, Shi F, et al. MiR-7, inhibited indirectly by lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of breast cancer stem cells by downregulating the STAT3 pathway. Stem Cells. 2014;32(11):2858–68.
Song Y, Wang R, Li LW, Liu X, Wang YF, Wang QX, et al. Long non-coding RNA HOTAIR mediates the switching of histone H3 lysine 27 acetylation to methylation to promote epithelial-to-mesenchymal transition in gastric cancer. Int J Oncol. 2019;54(1):77–86.
Lee JT, Bartolomei MS. X-inactivation, imprinting, and long noncoding RNAs in health and disease. Cell. 2013;152(6):1308–23.
Naciri I, Lin B, Webb C-H, Jiang S, Carmona S, Liu W, et al. Linking chromosomal silencing with Xist expression from autosomal integrated transgenes. Front Cell Dev Biol. 2021. https://doi.org/10.3389/fcell.2021.693154.
Statello L, Guo C-J, Chen L-L, Huarte M. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118.
Di Liegro CM, Schiera G, Di Liegro I. Regulation of mRNA transport, localization and translation in the nervous system of mammals (Review). Int J Mol Med. 2014;33(4):747–62.
Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–14.
Li H, Wang X, Wen C, Huo Z, Wang W, Zhan Q, et al. Long noncoding RNA NORAD, a novel competing endogenous RNA, enhances the hypoxia-induced epithelial-mesenchymal transition to promote metastasis in pancreatic cancer. Mol Cancer. 2017;16(1):169.
Gao N, Li Y, Li J, Gao Z, Yang Z, Li Y, et al. Long Non-Coding RNAs: the regulatory mechanisms, research strategies, and future directions in cancers. Front Oncol. 2020;10:598817.
Huarte M, Guttman M, Feldser D, Garber M, Koziol MJ, Kenzelmann-Broz D, et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response. Cell. 2010;142(3):409–19.
Ahmad P, Bensaoud C, Mekki I, Rehman MU, Kotsyfakis M. Long Non-Coding RNAs and their potential roles in the vector-host-pathogen triad. Life (Basel). 2021;11(1):56.
Arun G, Aggarwal D, Spector DL. MALAT1 Long Non-Coding RNA: functional implications. Noncoding RNA. 2020;6(2):22.
Xin X, Li Q, Fang J, Zhao T. LncRNA HOTAIR: a potential prognostic factor and therapeutic target in human cancers. Front Oncol. 2021. https://doi.org/10.3389/fonc.2021.679244.
Sakthianandeswaren A, Liu S, Sieber OM. Long noncoding RNA LINP1: scaffolding non-homologous end joining. Cell Death Discovery. 2016;2(1):16059.
Chan JJ, Tay Y. Noncoding RNA:RNA regulatory networks in cancer. Int J Mol Sci. 2018;19(5):1310.
Hussen BM, Azimi T, Hidayat HJ, Taheri M, Ghafouri-Fard S. Long Non-coding RNA RMRP in the pathogenesis of human disorders. Front Cell Dev Biol. 2021. https://doi.org/10.3389/fcell.2021.676588.
Wang Y, He L, Du Y, Zhu P, Huang G, Luo J, et al. The long noncoding RNA lncTCF7 promotes self-renewal of human liver cancer stem cells through activation of Wnt signaling. Cell Stem Cell. 2015;16(4):413–25.
Tang J, Li Y, Sang Y, Yu B, Lv D, Zhang W, et al. LncRNA PVT1 regulates triple-negative breast cancer through KLF5/beta-catenin signaling. Oncogene. 2018;37(34):4723–34.
Ma J, Yan H, Zhang J, Tan Y, Gu W. Long-Chain Non-Coding RNA (lncRNA) MT1JP suppresses biological activities of lung cancer by regulating miRNA-423-3p/Bim Axis. Med Sci Monit. 2019;25:5114–26.
Yu H, Wang S, Zhu H, Rao D. LncRNA MT1JP functions as a tumor suppressor via regulating miR-214–3p expression in bladder cancer. J Cell Physiol. 2019;234:16160.
Zhang Y, Sui R, Chen Y, Liang H, Shi J, Piao H. Long noncoding RNA MT1JP inhibits proliferation, invasion, and migration while promoting apoptosis of glioma cells through the activation of PTEN/Akt signaling pathway. J Cell Physiol. 2019;234(11):19553–64.
Ouyang Q, Cui Y, Yang S, Wei W, Zhang M, Zeng J, et al. lncRNA MT1JP suppresses biological activities of breast cancer cells in vitro and in vivo by regulating the miRNA-214/RUNX3 Axis. Onco Targets Ther. 2020;13:5033–46.
Gumireddy K, Li A, Yan J, Setoyama T, Johannes G, Orom U, et al. Identification of a long non-coding RNA-associated RNP complex regulating metastasis at the translational step. EMBO J. 2013;32:2672.
Venkatesh J, Wasson M-CD, Brown JM, Fernando W, Marcato P. LncRNA-miRNA axes in breast cancer: novel points of interaction for strategic attack. Cancer Lett. 2021;509:81–8.
Eades G, Wolfson B, Zhang Y, Li Q, Yao Y, Zhou Q. lincRNA-RoR and miR-145 regulate invasion in triple-negative breast cancer via targeting ARF6. Mol Cancer Res. 2015;13(2):330–8.
Kallen AN, Zhou XB, Xu J, Qiao C, Ma J, Yan L, et al. The imprinted H19 lncRNA antagonizes let-7 microRNAs. Mol Cell. 2013;52(1):101–12.
Raveh E, Matouk IJ, Gilon M, Hochberg A. The H19 Long non-coding RNA in cancer initiation, progression and metastasis—a proposed unifying theory. Mol Cancer. 2015;14:184.
Flores-Huerta N, Silva-Cázares MB, Arriaga-Pizano LA, Prieto-Chávez JL, López-Camarillo C. LncRNAs and microRNAs as essential regulators of stemness in breast cancer stem cells. Biomolecules. 2021;11(3):380.
Huang QY, Liu GF, Qian XL, Tang LB, Huang QY, Xiong LX. Long non-coding RNA: dual effects on breast cancer metastasis and clinical applications. Cancers (Basel). 2019;11(11):1802.
Zheng A, Song X, Zhang L, Zhao L, Mao X, Wei M, et al. Long non-coding RNA LUCAT1/miR-5582-3p/TCF7L2 axis regulates breast cancer stemness via Wnt/β-catenin pathway. J Exp Clin Cancer Res. 2019;38(1):305.
Tang T, Guo C, Xia T, Zhang R, Zen K, Pan Y, et al. LncCCAT1 promotes breast cancer stem cell function through activating WNT/β-catenin signaling. Theranostics. 2019;9(24):7384–402.
Song X, Zhang X, Wang X, Chen L, Jiang L, Zheng A, et al. LncRNA SPRY4-IT1 regulates breast cancer cell stemness through competitively binding miR-6882-3p with TCF7L2. J Cell Mol Med. 2020;24(1):772–84.
Li J, Hao Y, Mao W, Xue X, Xu P, Liu L, et al. LincK contributes to breast tumorigenesis by promoting proliferation and epithelial-to-mesenchymal transition. J Hematol Oncol. 2019;12(1):19.
Zhang G, Song W. Long non-coding RNA LSINCT5 inactivates Wnt/β-catenin pathway to regulate MCF-7 cell proliferation and motility through targeting the miR-30a. Ann Transl Med. 2020;8(24):1635.
Lu G, Li Y, Ma Y, Lu J, Chen Y, Jiang Q, et al. Long noncoding RNA LINC00511 contributes to breast cancer tumourigenesis and stemness by inducing the miR-185-3p/E2F1/Nanog axis. J Exp Clin Cancer Res. 2018;37(1):289.
Tu Z, Schmöllerl J, Cuiffo BG, Karnoub AE. Microenvironmental regulation of long noncoding RNA LINC01133 promotes cancer stem cell-like phenotypic traits in triple-negative breast cancers. Stem Cells. 2019;37(10):1281–92.
Han L, Yan Y, Zhao L, Liu Y, Lv X, Zhang L, et al. LncRNA HOTTIP facilitates the stemness of breast cancer via regulation of miR-148a-3p/WNT1 pathway. J Cell Mol Med. 2020;24(11):6242–52.
Deng J, Yang M, Jiang R, An N, Wang X, Liu B. Long Non-Coding RNA HOTAIR regulates the proliferation, self-renewal capacity, tumor formation and migration of the cancer stem-like cell (CSC) subpopulation enriched from breast cancer cells. PLoS ONE. 2017;12: e0170860.
Keshavarz M, Asadi MH. Long non-coding RNA ES1 controls the proliferation of breast cancer cells by regulating the Oct4/Sox2/miR-302 axis. Febs j. 2019;286(13):2611–23.
Lua Z-D, Jiao D-C, Qiao J-H, Yang S, Liu Z-Z. Long non-coding RNA SOX21-AS1 modulates breast cancer stem cells properties and carcinogenesis via targeting SOX2. Oncotarget. 2017. https://doi.org/10.18632/oncotarget.23593.
Zhang Z, Sun L, Zhang Y, Lu G, Li Y, Wei Z. Long non-coding RNA FEZF1-AS1 promotes breast cancer stemness and tumorigenesis via targeting miR-30a/Nanog axis. J Cell Physiol. 2018;233(11):8630–8.
Wang D, Wang Z, Zhang L, Sun S. LncRNA PDCD4-AS1 alleviates triple negative breast cancer by increasing expression of IQGAP2 via miR-10b-5p. Transl Oncol. 2021;14: 100958.
Niu Y, Bao L, Chen Y, Wang C, Luo M, Zhang B, et al. HIF2-Induced Long Noncoding RNA RAB11B-AS1 promotes hypoxia-mediated angiogenesis and breast cancer metastasis. Cancer Res. 2020;80(5):964–75.
Hua Q, Mi B, Xu F, Wen J, Zhao L, Liu J, et al. Hypoxia-induced lncRNA-AC020978 promotes proliferation and glycolytic metabolism of non-small cell lung cancer by regulating PKM2/HIF-1α axis. Theranostics. 2020;10(11):4762–78.
Kuo TC, Kung HJ, Shih JW. Signaling in and out: long-noncoding RNAs in tumor hypoxia. J Biomed Sci. 2020;27(1):59.
Zhu P, He F, Hou Y, Tu G, Li Q, Jin T, et al. A novel hypoxic long noncoding RNA KB-1980E63 maintains breast cancer stem cell stemness via interacting with IGF2BP1 to facilitate c-Myc mRNA stability. Oncogene. 2021;40(9):1609–27.
Wu L, Yang X. Targeting the hippo pathway for breast cancer therapy. Cancers. 2018;10(11):422.
Crabtree JS, Miele L. Breast cancer stem cells. Biomedicines. 2018;6(3):77.
Aaliyari-Serej Z, Ebrahimi A, Barazvan B, Ebrahimi-Kalan A, Hajiasgharzadeh K, Kazemi T, et al. Recent advances in targeting of breast cancer stem cells based on biological concepts and drug delivery system modification. Adv Pharm Bull. 2020;10(3):338–49.
Dittmer J. Breast cancer stem cells: features, key drivers and treatment options. Semin Cancer Biol. 2018;53:59–74.
Liu S, Dontu G, Mantle ID, Patel S, Ahn NS, Jackson KW, et al. Hedgehog signaling and Bmi-1 regulate self-renewal of normal and malignant human mammary stem cells. Cancer Res. 2006;66(12):6063–71.
He M, Fu Y, Yan Y, Xiao Q, Wu H, Yao W, et al. The Hedgehog signalling pathway mediates drug response of MCF-7 mammosphere cells in breast cancer patients. Clin Sci (Lond). 2015;129(9):809–22.
Valenti G, Quinn HM, Heynen G, Lan L, Holland JD, Vogel R, et al. Cancer stem cells regulate cancer-associated fibroblasts via activation of hedgehog signaling in mammary gland tumors. Cancer Res. 2017;77(8):2134–47.
Yang L, Shi P, Zhao G, Xu J, Peng W, Zhang J, et al. Targeting cancer stem cell pathways for cancer therapy. Signal Transduct Target Ther. 2020;5(1):8.
Najafi M, Farhood B, Mortezaee K. Cancer stem cells (CSCs) in cancer progression and therapy. J Cell Physiol. 2019;234(6):8381–95.
Jin H, Du W, Huang W, Yan J, Tang Q, Chen Y, et al. lncRNA and breast cancer: progress from identifying mechanisms to challenges and opportunities of clinical treatment. Mol Therapy Nucleic Acids. 2021;25:613–37.
Xing F, Liu Y, Wu S-Y, Wu K, Sharma S, Mo Y-Y, et al. Loss of XIST in breast cancer activates MSN-c-Met and reprograms microglia via exosomal miRNA to promote brain metastasis. Can Res. 2018;78(15):4316–30.
Li L, Liang S, Wasylishen AR, Zhang Y, Yang X, Zhou B, et al. PLA2G16 promotes osteosarcoma metastasis and drug resistance via the MAPK pathway. Oncotarget. 2016;7(14):18021–35.
Liu S, Sun Y, Hou Y, Yang L, Wan X, Qin Y, et al. A novel lncRNA ROPM-mediated lipid metabolism governs breast cancer stem cell properties. J Hematol Oncol. 2021;14(1):178.
Wang M, Li Y, Xiao GD, Zheng XQ, Wang JC, Xu CW, et al. H19 regulation of oestrogen induction of symmetric division is achieved by antagonizing Let-7c in breast cancer stem-like cells. Cell Prolif. 2019;52(1): e12534.
Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du Y-E, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem cells (Dayton, Ohio). 2015;34:55.
Xiu D-H, Liu G-F, Yu S-N, Li L-Y, Zhao G-Q, Liu L, et al. RETRACTED ARTICLE: Long non-coding RNA LINC00968 attenuates drug resistance of breast cancer cells through inhibiting the Wnt2/β-catenin signaling pathway by regulating WNT2. J Exp Clin Cancer Res. 2019;38(1):94.
Lainetti PF, Leis-Filho AF, Laufe-Amorim R, Battazza A, Fonseca-Alves CE. Mechanisms of resistance to chemotherapy in breast cancer and possible targets in drug delivery systems. Pharmaceutics. 2020;12(12):1193.
Liu Y, Li M, Yu H, Piao H. lncRNA CYTOR promotes tamoxifen resistance in breast cancer cells via sponging miR-125a-5p. Int J Mol Med. 2020;45(2):497–509.
Shi Q, Li Y, Li S, Jin L, Lai H, Wu Y, et al. LncRNA DILA1 inhibits Cyclin D1 degradation and contributes to tamoxifen resistance in breast cancer. Nat Commun. 2020;11(1):5513.
Ojo D, Wei F, Liu Y, Wang E, Zhang H, Lin X, et al. Factors promoting tamoxifen resistance in breast cancer via stimulating breast cancer stem cell expansion. Curr Med Chem. 2015;22(19):2360–74.
Li Y, Atkinson K, Zhang T. Combination of chemotherapy and cancer stem cell targeting agents: preclinical and clinical studies. Cancer Lett. 2017;396:103–9.
Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007;1(5):555–67.
Sládek NE. Human aldehyde dehydrogenases: potential pathological, pharmacological, and toxicological impact. J Biochem Mol Toxicol. 2003;17(1):7–23.
Wang C, Jia L, Wang Y, Du Z, Zhou L, Wen X, et al. Genome-wide interaction target profiling reveals a novel Peblr20-eRNA activation pathway to control stem cell pluripotency. Theranostics. 2020;10(1):353–70.
Luo L, Tang H, Ling L, Li N, Jia X, Zhang Z, et al. LINC01638 lncRNA activates MTDH-Twist1 signaling by preventing SPOP-mediated c-Myc degradation in triple-negative breast cancer. Oncogene. 2018;37(47):6166–79.
Brown JM, Wasson M-CD, Marcato P. The missing Lnc: the potential of targeting triple-negative breast cancer and cancer stem cells by inhibiting. Cells. 2020;9(3):763.
Hochmair MJ, Morabito A, Hao D, Yang CT, Soo RA, Yang JC, et al. Sequential treatment with afatinib and osimertinib in patients with EGFR mutation-positive non-small-cell lung cancer: an observational study. Future oncology (London, England). 2018;14(27):2861–74.
Zhou Q, Guo J, Huang W, Yu X, Xu C, Long X. Linc-ROR promotes the progression of breast cancer and decreases the sensitivity to rapamycin through miR-194-3p targeting MECP2. Mol Oncol. 2020;14(9):2231–50.
Fedele M, Sgarra R, Battista S, Cerchia L, Manfioletti G. The epithelial-mesenchymal transition at the crossroads between metabolism and tumor progression. Int J Mol Sci. 2022;23(2):800.
Tang L, Chen Y, Chen H, Jiang P, Yan L, Mo D, et al. DCST1-AS1 Promotes TGF-β-Induced epithelial-mesenchymal transition and enhances chemoresistance in triple-negative breast cancer cells via ANXA1. Front Oncol. 2020;10:280.
Taheri M, Mahmud Hussen B, Tondro Anamag F, Shoorei H, Dinger ME, Ghafouri-Fard S. The role of miRNAs and lncRNAs in conferring resistance to doxorubicin. J Drug Target. 2022;30(1):1–21.
Hamanaka S, Yamaguchi T, Kobayashi T, Kato-Itoh M, Yamazaki S, Sato H, et al. Generation of germline-competent rat induced pluripotent stem cells. PLoS ONE. 2011;6(7): e22008.
Zou H, Luo J, Guo Y, Liu Y, Wang Y, Deng L, et al. RNA-binding protein complex LIN28/MSI2 enhances cancer stem cell-like properties by modulating Hippo-YAP1 signaling and independently of Let-7. Oncogene. 2022;41:1657.
Zhang X, Powell K, Li L. Breast cancer stem cells: biomarkers, identification and isolation methods, regulating mechanisms, cellular origin, and beyond. Cancers (Basel). 2020;12(12):3765.
Xu Z, Liu C, Zhao Q, Lü J, Ding X, Luo A, et al. Long non-coding RNA CCAT2 promotes oncogenesis in triple-negative breast cancer by regulating stemness of cancer cells. Pharmacol Res. 2020;152: 104628.
Cui Y, Cao W, Li Q, Shen H, Liu C, Deng J, et al. Evaluation of prostate cancer antigen 3 for detecting prostate cancer: a systematic review and meta-analysis. Sci Rep. 2016;6(1):25776.
Chen J, Miao Z, Xue B, Shan Y, Weng G, Shen B. Long non-coding rnas in urologic malignancies: functional roles and clinical translation. J Cancer. 2016;7(13):1842–55.
Cantile M, Di Bonito M, Cerrone M, Collina F, De Laurentiis M, Botti G. Long Non-Coding RNA HOTAIR in breast cancer therapy. Cancers (Basel). 2020;12(5):1197.
Croker AK, Rodriguez-Torres M, Xia Y, Pardhan S, Leong HS, Lewis JD, et al. Differential functional roles of ALDH1A1 and ALDH1A3 in mediating metastatic behavior and therapy resistance of human breast cancer cells. Int J Mol Sci. 2017;18(10):2039.
Ho T-T, Zhou N, Huang J, Koirala P, Xu M, Fung R, et al. Targeting non-coding RNAs with the CRISPR/Cas9 system in human cell lines. Nucleic Acids Res. 2015;43(3):e17.
Rasul MF, Hussen BM, Salihi A, Ismael BS, Jalal PJ, Zanichelli A, et al. Strategies to overcome the main challenges of the use of CRISPR/Cas9 as a replacement for cancer therapy. Mol Cancer. 2022;21(1):64.
Brown JM, Wasson M-CD, Marcato P. The Missing Lnc: the potential of targeting triple-negative breast cancer and cancer stem cells by inhibiting long non-coding RNAs. Cells. 2020;9(3):763.
Khashkhashi Moghadam S, Bakhshinejad B, Khalafizadeh A, Mahmud Hussen B, Babashah S. Non-coding RNA-associated competitive endogenous RNA regulatory networks: novel diagnostic and therapeutic opportunities for hepatocellular carcinoma. J Cell Mol Med. 2022;26(2):287–305.
Lai Y, Chen Y, Lin Y, Ye L. Down-regulation of LncRNA CCAT1 enhances radiosensitivity via regulating miR-148b in breast cancer. Cell Biol Int. 2018;42(2):227–36.
Esmatabadi MJD, Motamedrad M, Sadeghizadeh M. Down-regulation of lncRNA, GAS5 decreases chemotherapeutic effect of dendrosomal curcumin (DNC) in breast cancer cells. Phytomedicine. 2018;42:56–65.
Chen PY, Hsieh PL, Peng CY, Liao YW, Yu CH, Yu CC. LncRNA MEG3 inhibits self-renewal and invasion abilities of oral cancer stem cells by sponging miR-421. J Formos Med Assoc. 2021;120(4):1137–42.
Zamani M, Sadeghizadeh M, Behmanesh M, Najafi F. Dendrosomal curcumin increases expression of the long non-coding RNA gene MEG3 via up-regulation of epi-miRs in hepatocellular cancer. Phytomedicine. 2015;22(10):961–7.
Polyzos A, Gogas H, Markopoulos C, Tsavaris N, Papadopoulos O, Polyzos K, et al. Salvage chemotherapy with oxaliplatin and capecitabine for breast cancer patients pretreated with anthracyclines and taxanes. Anticancer Res. 2009;29(7):2851.
Seyed Hosseini E, Alizadeh Zarei M, Babashah S, Nakhaei Sistani R, Sadeghizadeh M, Haddad Kashani H, et al. Studies on combination of oxaliplatin and dendrosomal nanocurcumin on proliferation, apoptosis induction, and long non-coding RNA expression in ovarian cancer cells. Cell Biol Toxicol. 2019;35(3):247–66.
Martinez-Balibrea E, Martínez-Cardús A, Ginés A, Ruiz de Porras V, Moutinho C, Layos L, et al. Tumor-related molecular mechanisms of oxaliplatin resistance. Mol Cancer Ther. 2015;14(8):1767–76.
Ghafouri-Fard S, Shoorei H, Bahroudi Z, Hussen BM, Talebi SF, Taheri M, et al. Nrf2-related therapeutic effects of curcumin in different disorders. Biomolecules. 2022;12(1):82.
Wang H, Zhang K, Liu J, Yang J, Tian Y, Yang C, et al. Curcumin regulates cancer progression: focus on ncRNAs and molecular signaling pathways. Front Oncol. 2021;11:660712.
Cai J, Sun H, Zheng B, Xie M, Xu C, Zhang G, et al. Curcumin attenuates lncRNA H19-induced epithelial-mesenchymal transition in tamoxifen-resistant breast cancer cells. Mol Med Rep. 2021;23(1):13.
Rosenlund IA, Calin GA, Dragomir MP, Knutsen E. CRISPR/Cas9 to Silence Long Non-Coding RNAs. Methods Mol Biol. 2021;2348:175–87.
Peng WX, Huang JG, Yang L, Gong AH, Mo YY. Linc-RoR promotes MAPK/ERK signaling and confers estrogen-independent growth of breast cancer. Mol Cancer. 2017;16(1):161.
Gao BB, Wang SX. LncRNA BC200 regulates the cell proliferation and cisplatin resistance in non-small cell lung cancer via PI3K/AKT pathway. Eur Rev Med Pharmacol Sci. 2019;23(3):1093–101.
Singh R, Gupta SC, Peng WX, Zhou N, Pochampally R, Atfi A, et al. Regulation of alternative splicing of Bcl-x by BC200 contributes to breast cancer pathogenesis. Cell Death Dis. 2016;7(6): e2262.
Koirala P, Huang J, Ho T-T, Wu F, Ding X, Mo Y-Y. LncRNA AK023948 is a positive regulator of AKT. Nature Commun. 2017;8:14422.
Acknowledgements
Not applicable.
Funding
Open Access funding enabled and organized by Projekt DEAL. This study was financially supported by Grant from Medical School of Shahid Beheshti University of Medical Sciences.
Author information
Authors and Affiliations
Contributions
SGF and BMH wrote the manuscript and revised it. MT supervised and designed the study. RKK, HJH, HSR, AS and STA collected the data and designed the figures and tables. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent of publication
Not applicable.
Competing interests
The authors declare they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hussen, B.M., Kheder, R.K., Abdullah, S.T. et al. Functional interplay between long non-coding RNAs and Breast CSCs. Cancer Cell Int 22, 233 (2022). https://doi.org/10.1186/s12935-022-02653-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-022-02653-4