
Abstract
Background
Migraine is a disabling and chronic neurovascular headache disorder. Trigeminal vascular activation and release of calcitonin gene-related peptide (CGRP) play a pivotal role in the pathogenesis of migraine. This knowledge has led to the development of CGRP(-receptor) therapies. Yet, a substantial proportion of patients do not respond to these treatments. Therefore, alternative targets for future therapies are warranted. The current narrative review provides a comprehensive overview of the pathophysiological role of these possible non-CGRP targets in migraine.
Findings
We covered targets of the metabotropic receptors (pituitary adenylate cyclase-activating polypeptide (PACAP), vasoactive intestinal peptide (VIP), amylin, and adrenomedullin), intracellular targets (nitric oxide (NO), phosphodiesterase-3 (PDE3) and -5 (PDE5)), and ion channels (potassium, calcium, transient receptor potential (TRP), and acid-sensing ion channels (ASIC)). The majority of non-CGRP targets were able to induce migraine-like attacks, except for (i) calcium channels, as it is not yet possible to directly target channels to elucidate their precise involvement in migraine; (ii) TRP channels, activation of which can induce non-migraine headache; and (iii) ASICs, as their potential in inducing migraine attacks has not been investigated thus far.
Drugs that target its receptors exist for PACAP, NO, and the potassium, TRP, and ASIC channels. No selective drugs exist for the other targets, however, some existing (migraine) treatments appear to indirectly antagonize responses to amylin, adrenomedullin, and calcium channels. Drugs against PACAP, NO, potassium channels, TRP channels, and only a PAC1 antibody have been tested for migraine treatment, albeit with ambiguous results.
Conclusion
While current research on these non-CGRP drug targets has not yet led to the development of efficacious therapies, human provocation studies using these targets have provided valuable insight into underlying mechanisms of migraine headaches and auras. Further studies are needed on these alternative therapies in non-responders of CGRP(-receptor) targeted therapies with the ultimate aim to pave the way towards a headache-free future for all migraine patients.
Similar content being viewed by others

Introduction
Migraine is a chronic neurovascular headache disorder, typically characterized by moderate to severe headache attacks which are accompanied by nausea, vomiting, photo- and phonophobia [1]. Approximately one third of migraine patients additionally suffer from transient neurologic symptoms called migraine auras [1]. Migraine poses a large socioeconomic burden and is ranked as the most disabling disorder in women under the age of fifty [2] — in whom prevalence is the highest [3, 4]. While exact pathophysiological mechanisms remain elusive, increased (sub)cortical excitability, trigeminovascular activation, and release of the neuropeptide calcitonin-gene related peptide (CGRP) — a member of the larger calcitonin family [5] — have been consistently demonstrated to play a pivotal and causative role in the pathogenesis of migraine [6, 7, 8]. The involvement of neuronal mechanisms (e.g. cortical spreading depression and sensitization of perivascular sensory nerve terminals) and structures (e.g. the hypothalamus and brainstem) are crucial for the initiation of migraine attacks, including the premonitory and aura phase [9]. CGRP alone seems unable to either activate or sensitize mechanosensitive meningeal nociceptive neurons in rats [10]. Nevertheless, its discovery has made CGRP the primary pharmacological target of recently approved (preventive) treatments that either target CGRP or its receptor, namely monoclonal antibodies and gepants [11, 12]. Despite their efficacy and tolerability in a substantial portion of both episodic and chronic migraine patients [13, 14], a significant percentage of migraine patients are classified as ‘non-responders’ and exhibit insufficient or no response to these CGRP(-receptor) targeted therapies. This emphasizes the need to explore the potential role of alternative substrates, which might serve as targets for future therapies in a variety of migraine patients [15]. Furthermore, the complex pathogenesis of migraine and its heterogeneous manifestations in patients suggest that different signaling pathways (neuropeptides and neurotransmitters) might be involved in different migraine patients [16] — highlighting the need to further explore these targets. These targets can broadly be categorized into (i) metabotropic receptors or G protein-coupled receptors, which include other members of the calcitonin family of peptides, i.e. amylin and adrenomedullin; (ii) intracellular targets; and (iii) ion channels. In this narrative review, a comprehensive overview is provided of the pathophysiological role of these non-CGRP mechanisms in migraine. We aimed to focus on three main questions for each target, namely whether (i) it has the potential to induce migraine-like attacks; (ii) we have drugs that target its receptors; and (iii) drugs against the substance have been tested for migraine treatment. Our primary aim was to provide a summary of current clinical evidence of these targets, considering that they might serve as a target for future pharmacological treatments and a valuable extension of the current therapeutic armamentarium for migraine.
Metabotropic receptors (G protein-coupled receptors)
Pituitary adenylate cyclase-activating polypeptide
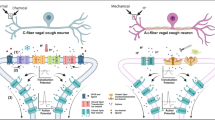
In 1989, pituitary adenylate cyclase-activating peptide (PACAP) was isolated from the hypothalamus [17] and over the years it has become a key molecule of interest in migraine research [18]. PACAP is an endogenous peptide belonging to the vasoactive intestinal polypeptide (VIP), secretin, and glucagon superfamily of peptides, existing in two major isoforms; a 38-amino acid neuropeptide known as PACAP38 and a shorter 27-amino acid truncated version, known as PACAP27 [17]. PACAP38 is more widely expressed, accounting for over 90% of PACAP in mammalian tissues and is detected in the trigeminal sensory and parasympathetic perivascular nerve fibers [18]. PACAP38 binds with equal affinity to the G-protein coupled receptors VPAC1 and VPAC2 [19, 20], but it is also capable of binding to a third G-protein coupled receptor, PAC1 [21] (Table 1). Activation of these receptors triggers the intracellular cyclic adenosine monophosphate (cAMP) pathway (Fig. 1).

A schematic, yet simplified, overview of (the interaction of) several CGRP-related and non-CGRP-related targets that might be of interest as future therapies in migraine. The final action of these targets is the efflux of potassium via the opening of KATP and BKCa channels. The figure was created using Biorender. Abbreviations: AC, Adenylate Cyclase; ADM, Adrenomedullin; AMP, Adenosine Monophosphate; AMY1, Amylin Receptor 1; AMY2, Amylin Receptor 2; BKCa, Big Conductance Calcium-Activated Potassium Channel; cAMP, Cyclic Adenosine Monophosphate; cGMP, Cyclic Guanosine Monophosphate; CGRP, Calcitonin Gene-Related Peptide; sGC, soluble Guanylyl Cyclase; KATP, Adenosine Triphosphate-Sensitive Potassium Channel; NO, Nitric Oxide; PAC1, Pituitary Adenylate Cyclase-Activating Polypeptide Type 1 Receptor; PACAP, Pituitary Adenylate Cyclase-Activating Polypeptide; PKA, Protein Kinase A; PKG, Protein Kinase G; PDE, phosphodiesterase; VIP, Vasoactive Intestinal Polypeptide; VPAC, Vasoactive Intestinal Polypeptide Receptor
Intravenous infusion of PACAP38 [57, 58] as well as PACAP27 [59] induced mild and short-lasting headaches in healthy volunteers. In patients with migraine without aura, intravenous infusion of both peptides additionally induced delayed migraine-like attacks. In double-blind, placebo-controlled, crossover trials, 58% of the patients reported migraine attacks after PACAP38 [57] and 55% after PACAP27 [60].
Given the interest in PACAP as a potential target for migraine therapeutics, several studies were conducted to investigate the migraine-evoking properties of PACAP. Preclinical studies indicated that PACAP-specific active transport systems exist across the blood brain barrier (BBB). However, once transported across the BBB, both isoforms are either rapidly degraded or diffuse back across the BBB into the blood, suggesting that PACAP exerts its effects mainly via peripheral mechanisms [61]. In vitro studies reported that PACAP38 was able to relax the vascular smooth muscle cells after abluminal, but not after luminal application in cerebral arteries [62]. In vivo studies on cerebral hemodynamics demonstrated no effect of intravenous infusion of PACAP38 on regional cerebral blood flow [63]. Moreover, in vivo studies using high-resolution magnetic resonance angiography reported a selective, marked and long-lasting vasodilatory effect of intravenous infusion of PACAP38 [20] and PACAP27 [59] on the extracerebral, but not the middle cerebral arteries. Upon administration of sumatriptan, the headache symptoms were relieved alongside contraction of the middle meningeal artery, but not the middle cerebral artery [58]. These findings suggest that the migraine-inducing effect of at least PACAP38 may be outside of the BBB. The mechanism of action is more complex and involves mast cell degranulation [31] and/or prolonged vasodilation [20]. Degranulation of mast cells causes release of histamine. It has been shown in rats that PACAP38 induces histamine release [64], and in migraine without aura patients, histamine triggers migraine-like attacks [65]. Yet, pretreatment with clemastine, a histamine (H1) antagonist, did not reduce the frequency of PACAP38-induced migraine attacks, suggesting an alternative mechanism for the migraine symptoms [66]. Using a double-blind, crossover design, a head-to-head comparison study of PACAP38 and VIP reported a significantly higher migraine-induction rate after PACAP38 compared to VIP (73% vs 18%, respectively). The study also showed prolonged meningeal artery dilation after PACAP38 but not after VIP infusion [20]. Moreover, sumatriptan was able to prevent PACAP38-induced migraine attacks, potentially via modulation of nociceptive transmission within the trigeminovascular system [67]. Thus, the prolonged meningeal artery dilation may play a role in PACAP38-induced migraine attacks. Receptor-wise, the main difference between PACAP38 and VIP is the over 1000-fold higher affinity of PACAP38 to the PAC1 receptor (Table 1), which positions the PAC1 receptor as a potential novel target for migraine treatment [68]. This led to the development of AMG301, a human monoclonal antibody selective for inhibition of the PAC1 receptor. However, clinical trials on AMG301 failed to prevent migraine [69]. Following the lack of success in targeting the PAC1 receptor, antibodies targeting the PACAP ligands have been proposed. For further details on the role of PACAP (blockade) in headaches, including migraine, we refer the readers to (recently published) reviews [70,71,72].
Vasoactive intestinal polypeptide
VIP is a 28-amino acid peptide with a short half-life of about one minute that is expressed in the intestine, pancreas, tongue, adrenal glands, urogenital tract, and brain (Table 1). VIP is thought to be involved in thermoregulation, cell proliferation, immune response, smooth muscle tone, and nociception [32, 73]. Among the complex scenarios of the trigeminovascular activation in the migraine attack, VIP seems to play an important role along with its homologous peptide PACAP [74]. VIP is released by parasympathetic fibers and exerts its role through the aforementioned receptors VPAC1 and VPAC2, which show a similar affinity to VIP and PACAP. VIP is theoretically able to bind also to PAC1; however, this latter receptor has a 1000-fold higher affinity to PACAP and is, therefore, considered to be PACAP-specific [31, 73] (Table 1, Fig. 1). In recent years, the role of VIP in migraine attacks has been extensively studied. Levels of plasma VIP have been found to be increased in the interictal phase in both adults with episodic and chronic migraine versus controls [75]. Levels of VIP in blood and saliva, though elevated during a spontaneous migraine attack, undergo a significant reduction after triptan administration in adults with migraine [76]. VIP is also involved in craniocervical vasomotor responses, and has a strong vasodilatory effect [76].
In a double-blind placebo-controlled crossover study, Hansen et al. evaluated twelve healthy subjects who received intravenous VIP or placebo over 25 min, which induced a very mild and short-lasting headache in five patients (42%) during VIP infusion compared to only one patient during placebo infusion [77]. In another study performed by Pellesi et al. twelve healthy volunteers were subjected to continuous VIP infusion over two hours, which provoked mild headache in 67% of participants, mainly in the post-infusion period. Interestingly, 25% of the participants reported migraine-like attacks [78]. Rahmann et al. were the first to evaluate the headache occurrence and vasomotor response in migraine patients after infusion of VIP or placebo in a double-blind crossover study [79]. Patients who received VIP showed marked but short-lasting vasodilation of both intracranial and extracranial arteries and a mild immediate headache with respect to placebo but no migraine-like attacks [79]. A more recent study reported that a two-hour infusion of VIP induced migraine attacks in 71% of patients with a history of migraine without aura [80]. The only methodological difference between Rahmann et al. [79] and the recent study from Pellesi et al. [80], was the duration of VIP infusion (i.e., 25 min versus 2 h). This suggests that the long-lasting (probably vascular) effect of VIP is more important for migraine induction. Indeed, the throbbing headache in migraine most likely originates in sensory fibers that transmit pain signals from intra- and extracranial (vasodilated) vessels, in particular arteries [81, 82]. So far, selective blockade of VIP has not been studied as a treatment of migraine. As current results suggest that a prolonged vasodilation due to VIP might provoke migraine-like attacks, VIP blockade might be a potential target in migraine treatment.
Amylin
Amylin belongs to the calcitonin peptides superfamily that includes CGRP, calcitonin, adrenomedullin, and adrenomedullin 2/intermedin [21, 37]. This is a 37-amino acid peptide structurally related to CGRP and is mainly released by the beta cells of islets of Langerhans in the pancreas [83] (Table 1). Amylin, which is secreted along with insulin, is involved in meal-ending satiation and inhibits insulin secretion [83, 84]. A key function of amylin is to maintain glucose homeostasis and reduce the uptake of glucose through its actions on the secretion of glucagon, gastric emptying, and caloric intake (reviewed by [85]). CGRP and amylin share CGRP and amylin 1 (AMY1) receptors, which have a similar structure composed of receptor activity modifying protein 1 (RAMP1) in combination with either calcitonin receptor (CTR) for AMY1, or calcitonin receptor-like receptor (CLR) for the CGRP receptor [5] (Table 1). As a result, the investigations of distribution and action of AMY1 have been challenged by its cross-reactivity with the CGRP receptor [5] (Fig. 1). While CGRP has equal affinity for the CGRP receptor and AMY1, amylin has a much lower affinity than CGRP for the CGRP receptor [83]. Considering these limitations, AMY1 has been posited to be distributed on structures involved in migraine pathophysiology, such as the trigeminal ganglion (TG) and the spinal trigeminal complex [83].
The development of the amylin analog pramlintide has aided research into the involvement of amylin in migraine pathophysiology. Pramlintide is an antidiabetic drug, approved by the U.S. Food and Drug Administration, with headache as a side effect in 13–17% of its users [83, 86]. In clinical trials, headache incidence was higher after pramlintide compared with placebo [83]. In coherence, intravenous infusion of pramlintide induced headache in 17% vs 6% with placebo in healthy subjects [87]. A subsequent provocation trial aimed to investigate the migraine induction rate of the pramlintide. In this head-to-head comparison study between pramlintide and CGRP in 36 patients 88% developed headache. Additionally, a migraine-like attack was triggered in 14 patients (41%) after pramlintide and 19 patients (56%) after CGRP. There was no statistical difference between the induction rates as well as the attack phenotypes were similar on both experimental days [88]. Human pharmacology studies demonstrated that one or more of the CTR/RAMP complexes (AMY1, AMY2, and AMY3 receptors) likely mediate the pramlintide-induced effects and migraine-like attacks, rather than CLR-based receptors (namely, the CGRP receptor or adrenomedullin-responsive receptors (AM1, AM2), which are described in the next section) [88]. Noteworthy, pramlintide produced little arterial vasodilation compared to CGRP (superficial temporal artery dilation, mean maximum change from baseline approximately 8.5% versus 115%, respectively), which could be explained by its lower potency compared to CGRP (> 1000-fold lower potency) for the CGRP receptor and possibly by a different distribution of CTR- and CLR-coupled receptors in the vascular tissue [88]. Further studies showed an increased amylin blood level in chronic migraine with basal level of pain, but not in interictal episodic migraine, suggesting that amylin may serve as a diagnostic biomarker for chronic migraine [89].
There is currently no treatment available to specifically antagonize the AMY1 receptor, although CGRP receptor-targeting treatments erenumab and rimegepant have been shown to antagonize AMY1 with a lower affinity compared with the CGRP receptor [90,91,92].
Adrenomedullin
Adrenomedullin is a 52-amino acid peptide belonging, like amylin, to the calcitonin peptide superfamily. It shares several structural features with CGRP such as a C-terminus amide and a loop structure in the N-terminus [93]. The adrenomedullin receptors AM1 and AM2 have a structure composed of RAMP2 or RAMP3, respectively, in combination with CLR. Interestingly, adrenomedullin acts on the CGRP receptor as well but is ten-fold less potent than CGRP [40] (Table 1). While CGRP and adrenomedullin are both released by sensory C-fibers, adrenomedullin is highly expressed by the vascular endothelium in contrast to CGRP [37, 94] – in particular within the cerebral circulation [38] (Fig. 1). Adrenomedullin stimulates endothelial-induced vasodilation within the cerebral circulation and plays an important role in the regulation of the BBB [95]. Nevertheless, the vasodilatory and hypotensive ability of adrenomedullin is less potent compared with CGRP. Expression of adrenomedullin and its receptors has also been described in neurons of the dorsal root ganglia and TG [37, 39], suggesting a role for adrenomedullin in the pain pathway. With a widespread distribution in various tissues, adrenomedullin plays a role in cardioprotection, reproduction, renal function, and lymphatic system [96]. Its anti-inflammatory, anti-apoptotic, and proliferative properties give adrenomedullin a potent protective role. This raised the interest for its therapeutic applications, notably in pulmonary hypertension and acute myocardial infarction (reviewed by [97]). The cross reaction of adrenomedullin with CGRP has spiked interest in its involvement in migraine pathophysiology. In mice, intrathecal administration of adrenomedullin induced mechanical hyperalgesia and inflammatory pain [98].
Two clinical studies explored the ability of adrenomedullin to provoke a migraine-like attack. A recent placebo-controlled two-way crossover study showed that adrenomedullin administration in 20 migraine patients induced a migraine-like attack in 55% of patients compared with 15% after placebo [99]. Conversely, twelve migraine patients showed in another study the same frequency (33.3%) of migraine-like attack after adrenomedullin administration compared with placebo, but the amount of adrenomedullin infused was slightly lower compared with the aforementioned study [100]. Finally, adrenomedullin was associated with headache induction in other human studies designed to explore the vascular effects of adrenomedullin [5]. Hence, adrenomedullin might induce migraine at possibly supraphysiological concentrations, by also activating the CGRP receptor, but the possibility of the occurrence of an attack at physiological concentrations is unclear to the best of our knowledge. A previous study in migraine patients showed lower plasma levels of adrenomedullin in the ictal and interictal phase compared to controls, suggesting an imbalance between CGRP and adrenomedullin in the migraine pathophysiology [101]. Moreover, which adrenomedullin-responsive receptors (AM1, AM2) or the CGRP receptor mediate these migraine-like responses have yet to be elucidated [99]. AM22-52 is the only adrenomedullin antagonist available but seems to be limited by its weak antagonizing effect on adrenomedullin responses (including adrenomedullin-stimulated cAMP) in rat cells, although it has been shown to have CGRP inhibiting effects [102, 103]. There is currently no treatment available to specifically antagonize adrenomedullin or its receptors, although one in vitro study demonstrated that the CGRP-receptor targeting monoclonal antibody erenumab and the CGRP-receptor small-molecule antagonist telcagepant antagonized not only CGRP, but also adrenomedullin signaling at the CGRP receptor [104].
Intracellular targets
Nitric oxide
Nitric oxide (NO) is a free radical synthesized by isoforms of nitric oxide synthase (NOS): endothelial NOS (eNOS), predominantly expressed in the endothelium and exerting a vasodilating effect; neuronal NOS (nNOS), present in central and peripheral nervous system; and inducible NOS (iNOS), which is involved in the innate immune system [46]. Upon binding of NO to its intracellular receptor, soluble guanylyl cyclase (sGC), levels of the second messenger cyclic guanosine monophosphate (cGMP) increase, and this leads to the opening of adenosine triphosphate-sensitive potassium (KATP) channels and big conductance calcium-activated potassium (BKCa) channels, which are speculated to play a crucial role in generating migraine attacks [105, 106] (Table 1, Fig. 1). The NO donor nitroglycerin, also known as glyceryl trinitrate, is one of the most commonly used experimental triggers for migraine headache [107]. Both headache of a pulsating nature during the administration of the drug and headache with a delayed onset of action were observed, fulfilling the characteristics of migraine without aura. There is also an increased sensitivity to nitroglycerin among migraine patients compared with individuals without a history of headache [108].
There have been studies towards new drugs targeting the arrest of the NO-cGMP cascade. An experiment in mice demonstrated that administration of the sGC stimulator VL-102 produced acute and sustained hyperalgesia. This effect was blocked by an sGC inhibitor (ODQ, 1H-[1,2,4]oxadiazolo[4,3,-a]quinoxalin-1-one), but also by several antimigraine medications (sumatriptan, topiramate, and propranolol) [109].
Additionally, researchers have explored NOS inhibition. Non-selective inhibitors may have important limitations mainly because of the significance of eNOS in regulating blood pressure [110], and therefore, researchers have focused on selective drugs. In a study of seven patients with migraine without aura, a significant increase of iNOS was observed during unprovoked migraine attacks [111]. Nevertheless, the highly selective iNOS inhibitor GW274150 failed to show efficacy both in acute [112] and preventive [113] treatment. In another study NXN-188, a combined nNOS inhibitor and serotonin (5-HT1B/1D) receptor agonist, inhibited the induced release of immunoreactive CGRP (iCGRP) from rat dura mater, TG, and trigeminal nucleus caudalis (TNC). Nevertheless, NXN-413, a selective nNOS inhibitor, inhibited iCGRP release from dura mater, but not from the TG and TNC in the same study [114]. NXN-188 did not reach a significant advantage over placebo in terminating migraine attacks in a small (n = 50) randomized trial [115]. Despite the ambiguous data concerning NOS inhibition as potential therapeutic target, the evidence of involvement of NO-cGMP cascade in the pathophysiology of migraine is constantly growing. Hence, there is a need for more reliable clinical trials, possibly on larger samples, to identify novel drug candidates targeting this molecular pathway.
Phosphodiesterase-3
CGRP activates adenylate cyclase transmembrane receptors in cerebral vascular cells and increases the second messenger cAMP. Intracellular accumulation of cAMP induces migraine attacks [116]. Phosphodiesterase-3 (PDE3) degrades cAMP (Fig. 1). In rats, PDE3 and cyclic nucleotide-gated ion channels are expressed in the trigeminovascular system, which includes the middle cerebral artery, basilar artery, TG, and dura mater [117]. Therefore, modulation of PDE3 in cAMP levels and activation of cyclic nucleotide-gated ion channels may play a role in the pathogenesis of migraine-like attacks [116]. PDE3 is indeed expressed in the TG, vascular smooth muscle cells, and cerebral arteries [118] (Table 1). Trigeminal neurons have been shown to be sensitized through an increase of cAMP [119, 120]. PDE3A and B are found with CGRP in the neuronal part of the trigeminovascular system [116]. Also, PDEs 3 and 4 degrade cAMP in vascular smooth muscle cells [121].
Since cilostazol inhibits PDE3, intracellular cAMP accumulates and the opening of KATP channels is facilitated, causing vascular smooth muscle relaxation and dilatation of cerebral arteries [122, 123] without affecting regional blood flow [116, 121, 124]. Thus, cilostazol leads to activation of the pathway that is further downstream than CGRP and PACAP (Fig. 1), and has a high success-rate to induce migraine-like attacks in 86% of adult patients with migraine without aura [116, 120, 125, 126], but also in healthy individuals [121]. Such is its potential role in migraine headache: modulating nociceptive input in regions where pain is processed in the neuronal parts of the trigeminovascular system [125]. These observations highlight the possible role of PDE3 in pain pathways involved in migraine headache [120, 125]. The effects of sumatriptan were studied in a cross-over study by Falkenberg et al. including migraine without aura patients in whom cilostazol-induced migraine was provoked. The majority of these patients were triptan responders, and the reduction in headache severity in the group given oral 50 mg sumatriptan was not significant at two hours (primary end-point), but rather at four hours when compared with the placebo group [118]. The authors concluded that, therefore, no clear effect was observed of sumatriptan on reducing cilostazol-induced migraine attacks. Hereafter, a double-blind cross-over study was conducted to study migraine headache development after cilostazol administration in migraine without aura patients [125]. The reduction of headache intensity in the group given 6 mg subcutaneous sumatriptan was significant at the second and fourth hour compared with the placebo group [125]. These results indicate that subcutaneous treatment rather than oral administration with sumatriptan is more effective in these cilostazol-induced headaches. These differences are probably related to its different pharmacokinetic properties, namely the faster peak in plasma concentrations, although a stronger placebo effect cannot be ruled out [125]. Khan et al. administered oral and subcutaneous sumatriptan to patients who developed migraine-like headaches after cilostazol administration. They stated that the highest decrease in headache score was after subcutaneous administration, whereas there was a smaller decrease in oral sumatriptan and other rescue drugs [120].
Thus, the modulation of PDE3 in intracellular cAMP levels may play a role in migraine attacks. Subcutaneous sumatriptan rather than oral sumatriptan may be an effective treatment modality for cilostazol-induced migraine attacks and headache intensity, although both exert relatively modest effects. Further studies are warranted to elucidate whether the modest extracellular effects of sumatriptan are due to its inability to completely block intracellular responses, namely the increased cAMP levels, induced by cilostazol [118].
Phosphodiesterase-5
Phosphodiesterase-5 (PDE5) is an intracellular enzyme that breaks down cGMP. Inhibition of PDE5 increases intracellular cGMP levels, leading to smooth muscle cell relaxation and neuronal stimulation [127] (Table 1, Fig. 1). Conversely, CGRP acts via increasing intracellular cAMP levels. PDE5 inhibitors, such as sildenafil and tadalafil, were initially produced for cardiovascular disorders, however, they were later demonstrated to be effective for sexual impotence and to have headache-inducing capacities [127, 128]. Indeed, several experimental studies revealed that up to 83% of migraine patients experience migraine-like headaches after sildenafil administration [129], and up to one third of individuals without migraine may also experience headaches [130].
CGRP and sildenafil act on cAMP and cGMP intracellular signaling pathways, respectively [131]. Nonetheless, patients experience migraine attacks with overlapping clinical features after administration of CGRP and sildenafil [131] suggesting that these two intracellular pathways likely converge in a downstream common denominator responsible for the biological migraine initiating cascade. Accordingly, mounting preclinical evidence supports a reciprocal influence between cAMP and cGMP pathways [47, 131,132,133]. Notably, administration of sildenafil induced more attacks in migraine patients compared to CGRP (89% vs 67%), potentially reflecting a more potent migraine-induction capacity [131].
Whilst the intracellular culprits of PDE5 inhibitors, and headache-inducing substances at large, have been recognized, their biological effects and site of action within the nervous system are still not fully understood. Initial investigations focused on potential cerebrovascular changes mediated by these substances. One study found no large intracranial or extracranial artery dilation after administering PDE5 inhibitors [130]. Another study observed dilation of intradural and extradural middle meningeal artery after CGRP or PDE5 inhibitor infusion, yet with no direct temporal correlation between arterial dilation and pain, suggesting this may reflect the activation of perivascular dural afferents [134]. Therefore, sensitization of perivascular sensory nerve terminals represents arguably a critical biological mechanism of migraine attack initiation [129]. Additionally, a transient increase in glutamate levels in the brainstem has been observed exclusively after PDE5 inhibitor administration, suggesting a possible different effect than CGRP [135]. Yet different pharmacokinetics properties of these molecules, namely the impermeability of the BBB to CGRP, but not to PDE5 inhibitors, may explain these results without discarding the common signaling pathway hypothesis.
PDE5 modulation represents a future unexplored therapeutic landscape. However, at present, no specific drugs which aim to increase PDE5 levels have been investigated in migraine. Thus, emulating the virtuous bench-to-bedside journey of the CGRP system may ensure further outstanding achievements in migraine.
Ion channels
A large range of ion channels are involved in migraine, likely including specific potassium channels and calcium channels. Furthermore, transient receptor potential (TRP) channels and acid-sensing ion channels (ASICs) have been proposed to play a role in migraine, due to their important roles in the trigeminothalamic and nociception systems. Yet, considering the fact that the conduction of human provocation models to study the specific activation of ASICs in humans is lacking, we consider them as two promising targets for future research (Box 1).
Potassium channels
Potassium channels are membrane-spanning protein complexes that are involved in the transportation of potassium ions to mediate or increase the membrane potential. Their activation is dependent on the regulatory domain differentiating the large number of potassium channels. Opening of KATP channels causes relaxation of vascular smooth muscle cells by K+ efflux, a reversible hyperpolarization of the membrane, and decreased intracellular Ca2+ [123, 136]. These phenomena can consequently lead to the induction of a headache attack [137, 138].
High extracellular concentrations of potassium can trigger cortical spreading depression (CSD), which is thought to be a cause of aura in migraine with aura patients [139, 140], activating an inflammatory cascade, which leads to an increase of intracellular calcium and release of proinflammatory peptides (e.g. CGRP).
Genome-wide association studies in migraine patients identified several potassium channel linked genes that were likely susceptible, providing further evidence for their involvement (reviewed in [141]). Mutations in the two-pore-domain potassium channel TRESK have been identified in migraine patients. TRESK inhibits TREK1 and TREK2, thereby increasing excitability of the TG [142]. Reducing the TG excitability using the TREK1/TREK2 agonist ML67-33 reduced an NO donor-induced migraine-like phenotype in mice in a similar manner as the CGRP receptor antagonist olcegepant. Moreover, it completely reversed TG-mediated NO donor-induced facial allodynia in rats [143].
Acting by direct opening of KATP channels, levcromakalim is a potent vasodilator as well as a trigger of headache in healthy subjects and of migraine-like attacks in migraine patients following systemic administration [136, 144]. Some observations revealed that levcromakalim acts downstream from the CGRP receptor and hypersensitivity caused by levcromakalim is independent of direct CGRP release from the TNC and the TG [136, 139]. On the other hand, experimental mice models showed that levcromakalim-induced hypersensitivity was blocked by CGRP neutralization [123].
In vitro studies showed that glibenclamide, a non-specific KATP channel blocker and a widely used anti-diabetic drug, attenuated PACAP-induced dilation of rat cerebellar and human pulmonary arteries [144, 145].
In healthy volunteers, treatment with glibenclamide did not mitigate PACAP38-induced headache and vascular changes [136]. A randomized, placebo-controlled cross-over study in healthy individuals showed that administration of glibenclamide did not induce headache, nor did it prevent headache or changes in blood pressure and heart rate induced by levcromakalim. Yet, the authors found that glibenclamide rather delayed the onset of the induced headaches [146]. More selective KATP channel blockers are needed to clarify potential mechanisms of the KATP channel in pathways involved in migraine, including the PACAP38 signaling pathway.
In contrast to KATP channels, the specific voltage-gated potassium Kv7 channels potentially reduce migraine when opened, as demonstrated in male rats. The Kv7 channel opener retigabine significantly reduced basal and TRP channel-induced CGRP release in vitro [147].
Another investigative target includes the BKCa channels, which are widely expressed in the brain and cardiovascular system and facilitate potassium efflux, hyperpolarization, and thus decreased neuronal excitability [148, 149]. BKCa channels require both membrane depolarisation and binding of Ca2+ for opening, and have been found to be co-localised with voltage-gated Ca2+ channels [150]. Of particular interest here is their role in the activation of vascular smooth muscle cells and related involvement in the trigeminovascular system [151]. Additionally, they modulate neurotransmitter release in peripheral neurons, and their opening has been shown to inhibit Aδ-fiber firing in the rat’s TNC when applied directly [152], whereas genetically ablating them from sensory neurons in mice increased nociceptive behaviors in some but not all pain models [153]. BKCa channels have furthermore been connected with various signaling pathways and factors known to contribute to migraine, such as CGRP and PACAP [154]. This does indeed raise the possibility of downstream involvement in the migraine pathophysiology.
Results of a small pilot study confirmed that pharmacologic opening of BKCa channels using a small vasoactive molecule (MaxiPost) did indeed provoke cephalic vasodilation in 90% of healthy subjects, as compared to 30% with placebo injection [155]. Finally, Al-Karagholi et al. demonstrated that MaxiPost induced migraine-like attacks – and not merely vasodilatory headache – in 21 of 22 migraine patients [156].
Both preclinical and clinical findings, albeit contrasting, might reflect the wide diversity of BKCa channel types [148, 150]. Therefore, molecules selectively targeting the channels or specific protein subunits of BKCa that are involved in migraine pathophysiology should be preferably developed. This in order to avoid non-selective action and related side effects, as is the case with calcium channel blockers and other drugs currently used for migraine therapy [157]. These findings, analogous to earlier research on CGRP, suggest that BKCa channel antagonists could prove to be promising molecules in the pharmacotherapy of migraine.
Calcium channels
Calcium channels are voltage-gated ion channels that regulate the flow of Ca2+ ions, an essential intracellular second messenger. Calcium blockers have long been used as migraine prophylactic drugs that target broad calcium receptors such as nimodipine and flunarizine that, following the cerebral hypoxia hypothesis, aim to reduce excessive intracellular influx of calcium ions that results from increased neuronal excitability [158, 159]. It should, however, be noted that the efficacy of these drugs is likely not mediated by calcium channel action but by other mechanisms, including the suppression of CSD [160].
One of the main calcium channels implicated in migraine is the voltage-gated Cav2.1, found in presynaptic terminals throughout the central nervous system with its main role involving neurotransmitter release and dendritic transients [161, 162]. These channels give rise to P/Q-type calcium currents and its encoding gene, CACNA1A, stands as a classical genetic locus associated with the rare genetic disease familial hemiplegic migraine (FHM) [163].
Several missense mutations have been found in FHM families, including those that code for the voltage sensor and pore structures in the α1 subunit of the channel [163,164,165]. There is evidence that these mutations present gain-of-function changes in cortical excitatory neurotransmission, reducing the threshold for CSD, all the while interneuron inhibition remains intact, potentially driving an excitatory-inhibitory imbalance [166, 167]. Additionally, this mechanism may also play a role in descending pain modulatory pathways, as its blockade has been shown to enhance trigeminal transmission in the rat [168]. Furthermore, Chan et al. [169] assessed trigeminovascular activity in a genetic mouse model of FHM and found responses that reflect desensitization of CGRP receptors. However, most (experimental) treatments for CACNA1A-associated disorders do not specifically target this channel. One possible exception is 4-aminopyridine; more widely known as a potassium channel blocker, pharmacologic activity on calcium channels has been described [170]. To our knowledge, no clinical studies on migraine using this molecule have yet been carried out. Nonetheless, in the context of CACNA1A-associated spinocerebellar ataxia type 6, some preliminary and preclinical possibilities for selective small molecules were reported in 2018 [171].
The role of calcium channels in migraine has been explored to a limited extent, due to its many roles in neurotransmitter release and secondary messenger pathways, which makes it difficult to extract its precise involvement in migraine. Further work on calcium channels and how they interact with other neurotransmitter circuits, especially in the case of genetic migraine, may uncover new potential therapeutic targets of migraine pathophysiology.
Transient receptor potential (TRP) channels
The TRP channel superfamily consists of non-selective cation channels that mediate the transmembrane flow of calcium and are mainly implicated in sensory transduction. They are also thought to modulate pain processing and particularly the vanilloid TRP (TRPV) channel subfamily is considered relevant here [172, 173]. Several TRP channels have been identified as potential antimigraine targets, including the TRPV1 and TRP ankyrin 1 (TRPA1) channels [174] (Fig. 1). Briefly, TRPV1 channels process thermal and pH-related nociception, are involved in calcium release in sensory neurons, and are expressed in the trigeminal pathway [175, 176]. Their relevance in migraine extends further, as activation of these channels induces release of CGRP and increases TG activity, which is reversed by sumatriptan [175, 177]. TRPV1 channel modulators have had moderate success in the treatment of migraine, with part of their antinociceptive activity being affected by topical agonists, such as capsaicin, to desensitize the channel [178]. Capsaicin is a TRPV1 agonist and its intranasal administration, as well as its synthetic isosomer civamide, have shown some efficacy for the treatment of cluster headache and migraine [179, 180]. However, discrepancies in efficacy and safety between preclinical and clinical studies justify the need for further research [175].
On the other hand, the TRPA1 channel has attracted attention due to its ability to transduce a multitude of chemical agents such as formaldehyde and cigarette smoke, many of which are also common migraine triggers [174]. Notably, the “headache tree” extract, umbellulone, is able to trigger headache when inhaled, and has shown to act on the trigeminovascular system to induce CGRP release from trigeminal afferents in the dura [181]. Parthenolide, another herb extract, is a partial TRPA1 agonist and has the ability to desensitize it, effecting anti-migraine responses on CGRP-mediated trigeminal activity [182], and positioning it as a potential migraine target. We refer the readers to several excellent reviews on TRP channels for further details on their role in the pathophysiology of migraine [183, 184].
Future directions
Human provocation studies have contributed to an enhanced understanding of the pathophysiology of migraine and to the identification of possible new targets of treatment for migraine. Despite our growing knowledge on these alternative targets – all leading to vasodilation of intracranial arteries (Fig. 1) – no successful therapies to block CGRP-independent mechanisms have been developed yet. Indeed, an antibody to block PACAP, which is a member of the VIP, secretin, and glucagon superfamily of peptides, has been the first developed alternative therapy, although it was not effective in trials. Further, data on blockade of NO-induced responses have been ambiguous, and treatment with glibenclamide did not mitigate PACAP38-induced and levcromakalim-induced headaches. Yet, TRPV1 agonist capsaicin and civamide have shown some efficacy by their ability to desensitize nerve endings that express these channels. Further research is needed to develop alternative targeted therapies to prevent migraine (i.e. targeting VIP, amylin, adrenomedullin, PDE3, PDE5, calcium channels, and ASICs). Theoretically, blockade of the most downstream targets (e.g. KATP channels) – being the “end-chain” in the cascade – might lead to more efficacious outcomes, but also to possible severe and highly undesired side effects [189]. Whether this explains the highest induction rate of levcromakalim (Fig. 2) remains to be demonstrated.

Mean induction rates (with their standard deviations) of migraine attacks (irrespective of aura symptoms) of non-CGRP targets compared to placebo, as observed in placebo-controlled studies in migraine patients. PACAP38 rate is based on [57] and [20] – the latter is a head-to-head comparison study of PACAP38 and VIP (instead of placebo); PACAP27 rate is based on [60]; VIP rate is based on [20] with an active control (PACAP38) and [79, 80]. Please note that infusion duration in the study of Rahmann et al. [79] was 25 minutes, while it was two hours in the study of Pellesi et al. [80]; pramlintide rate is based on [88] which was compared to CGRP and not to placebo; adrenomedullin rate is based on [99]; nitroglycerin rate is based on [190]; cilostazol (PDE3 inhibitor) rate is based on [116]; sildenafil (PDE5 inhibitor) rate is based on [129], and levcromakalim rate is based on [137, 191]. Please note that [191] only included migraine with aura patients
However, to study such substances as potential candidates for migraine therapy, the presence of selective agonists or stimulating agents are indispensable. While the majority of non-CGRP targets were able to induce migraine-like attacks with relatively high induction rates (Fig. 2), this was not the case for (i) calcium channels, as it is not yet possible to directly target channels to elucidate their precise involvement in migraine; (ii) TRP channels, activation of which can induce non-migraine headache; and (iii) ASICs, as their the potential in inducing migraine attacks has not been investigated thus far. While we emphasize that the current overview is far from complete, we have aimed to provide a comprehensive overview of the (clinically) most relevant future targets for migraine. Yet, further studies on the receptor pharmacology and dual (and overlapping) agonism of several other members of the CGRP/calcitonin peptide family and calcitonin mimetics are warranted, as well as knowledge on clinical consequences of receptor internalization and recycling. As an example, salmon calcitonin was already approved as a treatment of metabolic bone diseases [192] and has been shown to have antinociceptive properties, posing it as a potential treatment migraine option as well [93, 193]. Rat studies have shown that it might ameliorate migraine-like pain through modulation of CGRP release and mast cell degranulation in the dura mater [194]. Salmon calcitonin shows potent AMY3 agonism, but also binds with approximately a similar affinity as human calcitonin to CTR, while amylin/pramlintide can also activate these latter receptors [93, 195].
Further, human provocation studies, in which these (non-CGRP) targets are studied through their ability to induce migraine attacks, are necessary. A limitation of these human provocation studies is that they cannot reveal the initial cause of spontaneous migraine attacks [105], nor are they able to provoke migraine auras consistently [126, 196], except for levcromakalim [137]. In addition, they do not induce attacks in all patients or account for differences in trigger sensitivity between migraine patients and the interplay of different compounds [144]. Indeed, theoretically, it should be considered that blockade of one system might lead to (over)activation of compensatory mechanisms. Therefore, the development of blockers (antagonists) of one (isolated) target or system might not guarantee efficacious headache relief in all migraine patients. Also, in different migraine patients, different pathways might be primarily activated – as demonstrated by the clinical study of Ghanizada et al. who showed that a subgroup of migraine patients only respond to CGRP and a few only to pramlintide [88]. It remains to be demonstrated whether this subgroup of patients are also non-responders to CGRP(-receptor) targeted therapies.
Future translational studies are warranted to further optimize human provocation models that allow the study of novel drug targets in migraine. In addition, basic and clinical studies are needed to improve our understanding on (i) the role of these non-CGRP targets in uncontrolled settings, given the controlled environment of the human provocation studies; (ii) the eventual role of additional targets that might play a role in the pathophysiology of auras; (iii) potential (long-term) side effects, especially considering the physiological role of e.g. adrenomedullin in the cardiovascular system [197]; (iv) the relationship between the relative contribution of different targets (e.g. amylin, adrenomedullin, PACAP) in the ictal and interictal phase in both EM and CM patients; and (v) the role of sex (steroids) on the activity of these different targets, and vice versa, considering the marked influence of sex steroids in migraine [198]. In the longer term, studies should be conducted to investigate the potential efficacy of these alternative therapies in non-responders of CGRP(-receptor) targeted therapies with the ultimate aim to, hopefully, pave the way towards a headache-free future for all migraine patients.
Conclusion
In conclusion, the development of alternatives for CGRP(-receptor) blocking agents is essential, given the amount of non-responders to these drugs and the potential (long-term) side effects. While current research on these non-CGRP drug targets has not led to the development of efficacious therapies yet, studies on their provoking substances in human models have provided valuable insights into underlying mechanisms of migraine headaches and auras. Further research is warranted to understand the interplay of these different agents in the ictal and interictal phase as well as their relative contributions, especially in relation to the heterogeneous manifestations of migraine.
Availability of data and materials
Not applicable.
Abbreviations
- AC:
-
Adenylate Cyclase
- ADM:
-
Adrenomedullin
- AMP:
-
Adenosine Monophosphate
- AMY1 :
-
Amylin Receptor 1
- AMY2 :
-
Amylin Receptor 2
- AMY3 :
-
Amylin Receptor 3
- ASIC:
-
Acid-Sensing Ion Channel
- BBB:
-
Blood Brain Barrier
- BKCa :
-
Big Conductance Calcium-Activated Potassium Channel
- cAMP:
-
Cyclic Adenosine Monophosphate
- cGMP:
-
Cyclic Guanosine Monophosphate
- CGRP:
-
Calcitonin Gene-Related Peptide
- CLR:
-
Calcitonin-Like Receptor
- CSD:
-
Cortical Spreading Depression
- CTR:
-
Calcitonin Receptor
- FMH:
-
Familial Hemiplegic Migraine
- KATP :
-
Adenosine Triphosphate-Sensitive Potassium Channel
- NO:
-
Nitric Oxide
- PAC1 :
-
Pituitary Adenylate Cyclase-Activating Polypeptide Type 1 Receptor
- PACAP:
-
Pituitary Adenylate Cyclase-Activating Polypeptide
- PDE:
-
Phosphodiesterase
- PKA:
-
Protein Kinase A
- PKG:
-
Protein Kinase G
- RAMP1:
-
Receptor Activity-Modifying Protein
- SCA:
-
Spinocerebellar Ataxia
- sGC:
-
Soluble Guanylyl Cyclase
- TG:
-
Trigeminal Ganglion
- TNC:
-
Trigeminal Nucleus Caudalis
- TRP:
-
Transient Receptor Potential
- VIP:
-
Vasoactive Intestinal Polypeptide
- VPAC:
-
Vasoactive Intestinal Polypeptide Receptor
References
Headache Classification Committee of the International Headache Society (IHS) (2018) The International Classification of Headache Disorders, 3rd edition. Cephalalgia 38(1):1–211
GBD 2019 Diseases and Injuries Collaborators (2020) Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet 396(10258):1204–1222
Ashina M, Katsarava Z, Do TP, Buse DC, Pozo-Rosich P, Özge A et al (2021) Migraine: epidemiology and systems of care. Lancet 397(10283):1485–1495. https://doi.org/10.1016/S0140-6736(20)32160-7
Victor TW, Hu X, Campbell JC, Buse DC, Lipton RB (2010) Migraine prevalence by age and sex in the United States: a life-span study. Cephalalgia 30(9):1065–1072
Rees TA, Hendrikse ER, Hay DL, Walker CS (2022) Beyond CGRP: the calcitonin peptide family as targets for migraine and pain. Br J Pharmacol 179(3):381–399
Ashina M, Terwindt GM, Al-Karagholi MA, de Boer I, Lee MJ, Hay DL et al (2021) Migraine: disease characterisation, biomarkers, and precision medicine. Lancet 397(10283):1496–1504. https://doi.org/10.1016/S0140-6736(20)32162-0
Edvinsson L, Haanes KA, Warfvinge K, Krause DN (2018) CGRP as the target of new migraine therapies — successful translation from bench to clinic. Nat Rev Neurol 14(6):338–350
Dodick DW (2018) Migraine. Lancet 391:1315–1330. https://doi.org/10.1016/s0140-6736(18)30478-1
Dodick DW (2018) A phase-by-phase review of migraine pathophysiology. Headache 58(Suppl 1):4–16
Levy D, Burstein R, Strassman AM (2005) Calcitonin gene-related peptide does not excite or sensitize meningeal nociceptors: implications for the pathophysiology of migraine. Ann Neurol 58(5):698–705
Ashina M, Buse DC, Ashina H, Pozo-Rosich P, Peres MFP, Lee MJ et al (2021) Migraine: integrated approaches to clinical management and emerging treatments. Lancet 397(10283):1505–1518. https://doi.org/10.1016/S0140-6736(20)32342-4
Al-Hassany L, Goadsby PJ, Danser AHJ, MaassenVanDenBrink A (2022) Calcitonin gene-related peptide-targeting drugs for migraine: how pharmacology might inform treatment decisions. Lancet Neurol 21(3):284–294
Moreno-Ajona D, Villar-Martínez MD, Goadsby PJ (2022) New generation gepants: migraine acute and preventive medications. J Clin Med Res 11(6):1656
Wang X, Chen Y, Song J, You C (2021) Efficacy and safety of monoclonal antibody against calcitonin gene-related peptide or its receptor for migraine: a systematic review and network meta-analysis. Front Pharmacol 12:649143
Do TP, Al-Saoudi A, Ashina M (2021) Future prophylactic treatments in migraine: beyond anti-CGRP monoclonal antibodies and gepants. Rev Neurol 177(7):827–833
Charles A (2018) The pathophysiology of migraine: implications for clinical management. Lancet Neurol 17(2):174–182
Miyata A, Arimura A, Dahl RR, Minamino N, Uehara A, Jiang L et al (1989) Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates adenylate cyclase in pituitary cells. Biochem Biophys Res Commun 164(1):567–574
Schytz HW, Olesen J, Ashina M (2010) The PACAP receptor: a novel target for migraine treatment. Neurotherapeutics 7(2):191–196
Harmar AJ, Fahrenkrug J, Gozes I, Laburthe M, May V, Pisegna JR et al (2012) Pharmacology and functions of receptors for vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide: IUPHAR review 1. Br J Pharmacol 166(1):4–17
Amin FM, Hougaard A, Schytz HW, Asghar MS, Lundholm E, Parvaiz AI et al (2014) Investigation of the pathophysiological mechanisms of migraine attacks induced by pituitary adenylate cyclase-activating polypeptide-38. Brain 137(Pt 3):779–794
Dominguez-Moreno R, Do TP, Ashina M (2022) Calcitonin gene-related peptide and pituitary adenylate cyclase-activating polypeptide in migraine treatment. Curr Opin Endocrinol Diabetes Obes 29(2):225–231
Kee Z, Kodji X, Brain SD (2018) The role of Calcitonin Gene Related Peptide (CGRP) in neurogenic vasodilation and its cardioprotective effects. Front Physiol 9:1249
Russo AF, Hay DL (2023) CGRP physiology, pharmacology, and therapeutic targets: migraine and beyond. Physiol Rev 103(2):1565–1644
Kraenzlin ME, Ch’ng JL, Mulderry PK, Ghatei MA, Bloom SR (1985) Infusion of a novel peptide, calcitonin gene-related peptide (CGRP) in man Pharmacokinetics and effects on gastric acid secretion and on gastrointestinal hormones. Regul Pept 10(2–3):189–197
Sherwood NM, Krueckl SL, McRory JE (2000) The origin and function of the pituitary adenylate cyclase-activating polypeptide (PACAP)/glucagon superfamily. Endocr Rev 21(6):619–670
Hirabayashi T, Nakamachi T, Shioda S (2018) Discovery of PACAP and its receptors in the brain. J Headache Pain 19(1):28
Ramos-Álvarez I, Mantey SA, Nakamura T, Nuche-Berenguer B, Moreno P, Moody TW et al (2015) A structure–function study of PACAP using conformationally restricted analogs: identification of PAC1 receptor-selective PACAP agonists. Peptides 66:26–42
Bourgault S, Vaudry D, Botia B, Couvineau A, Laburthe M, Vaudry H et al (2008) Novel stable PACAP analogs with potent activity towards the PAC1 receptor. Peptides 29(6):919–932
Banks WA, Kastin AJ, Komaki G, Arimura A (1993) Passage of pituitary adenylate cyclase activating polypeptide1-27 and pituitary adenylate cyclase activating polypeptide1-38 across the blood-brain barrier. J Pharmacol Exp Ther 267(2):690–696
Ganea D, Hooper KM, Kong W (2015) The neuropeptide vasoactive intestinal peptide: direct effects on immune cells and involvement in inflammatory and autoimmune diseases. Acta Physiol 213(2):442–452
Jansen-Olesen I, Hougaard PS (2018) PACAP and its receptors in cranial arteries and mast cells. J Headache Pain 19(1):16
Domschke S, Domschke W, Bloom SR, Mitznegg P, Mitchell SJ, Lux G et al (1978) Vasoactive intestinal peptide in man: pharmacokinetics, metabolic and circulatory effects. Gut 19(11):1049–1053
Dogrukol-Ak D, Banks WA, Tuncel N, Tuncel M (2003) Passage of vasoactive intestinal peptide across the blood–brain barrier. Peptides 24(3):437–444
Hay DL, Chen S, Lutz TA, Parkes DG, Roth JD (2015) Amylin: pharmacology, physiology, and clinical potential. Pharmacol Rev 67(3):564–600
Clodi M, Thomaseth K, Pacini G, Hermann K, Kautzky-Willer A, Waldhüsl W et al (1998) Distribution and kinetics of amylin in humans. Am J Physiol 274(5):E903–E908
Banks WA, Kastin AJ, Maness LM, Huang W, Jaspan JB (1995) Permeability of the blood-brain barrier to amylin. Life Sci 57(22):1993–2001
Edvinsson L, Grell AS, Warfvinge K (2020) Expression of the CGRP family of neuropeptides and their receptors in the trigeminal ganglion. J Mol Neurosci 70(6):930–944
Kis B, Kaiya H, Nishi R, Deli MA, Abrahám CS, Yanagita T et al (2002) Cerebral endothelial cells are a major source of adrenomedullin. J Neuroendocrinol 14(4):283–293
Ma W, Chabot JG, Quirion R (2006) A role for adrenomedullin as a pain-related peptide in the rat. Proc Natl Acad Sci U S A 103(43):16027–16032
Garelja ML, Bower RL, Brimble MA, Chand S, Harris PWR, Jamaluddin MA et al (2022) Pharmacological characterisation of mouse calcitonin and calcitonin receptor-like receptors reveals differences compared with human receptors. Br J Pharmacol 179(3):416–434
Meeran K, O’Shea D, Upton PD, Small CJ, Ghatei MA, Byfield PH et al (1997) Circulating adrenomedullin does not regulate systemic blood pressure but increases plasma prolactin after intravenous infusion in humans: a pharmacokinetic study. J Clin Endocrinol Metab 82(1):95–100
Kis B, Deli MA, Kobayashi H, Abrahám CS, Yanagita T, Kaiya H et al (2001) Adrenomedullin regulates blood-brain barrier functions in vitro. NeuroReport 12(18):4139–4142
Bredt DS (1999) Endogenous nitric oxide synthesis: biological functions and pathophysiology. Free Radic Res 31(6):577–596
Guix FX, Uribesalgo I, Coma M, Muñoz FJ (2005) The physiology and pathophysiology of nitric oxide in the brain. Prog Neurobiol 76(2):126–152
Thomas DD, Liu X, Kantrow SP, Lancaster JR (2001) The biological lifetime of nitric oxide: implications for the perivascular dynamics of NO and O2. Proc Natl Acad Sci U S A 98(1):355–360
Olesen J (2008) The role of nitric oxide (NO) in migraine, tension-type headache and cluster headache. Pharmacol Ther 120(2):157–171
Nordgaard JC, Kruse LS, Møller M, Kruuse C (2014) Phosphodiesterases 3 and 5 express activity in the trigeminal ganglion and co-localize with calcitonin gene-related peptide. Cephalalgia 34(7):503–513
Bender AT, Beavo JA (2006) Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use. Pharmacol Rev 58(3):488–520
Menniti FS, Faraci WS, Schmidt CJ (2006) Phosphodiesterases in the CNS: targets for drug development. Nat Rev Drug Discov 5(8):660–670
Cesarini V, Guida E, Campolo F, Crescioli C, Di Baldassarre A, Pisano C et al (2020) Type 5 phosphodiesterase (PDE5) and the vascular tree: from embryogenesis to aging and disease. Mech Ageing Dev 190:111311
Pershing LK, Reilly CA, Corlett JL, Crouch DJ (2004) Effects of vehicle on the uptake and elimination kinetics of capsaicinoids in human skin in vivo. Toxicol Appl Pharmacol 200(1):73–81
Chaiyasit K, Khovidhunkit W, Wittayalertpanya S (2009) Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. J Med Assoc Thai 92(1):108–113
Kawada T, Watanabe T, Katsura K, Takami H, Iwai K (1985) Formation and metabolism of pungent principle of capsicum fruits: XV. Microdetermination of capsaicin by high-performance liquid chromatography with electrochemical detection. J Chromatogr A 329:99–105
Rollyson WD, Stover CA, Brown KC, Perry HE, Stevenson CD, McNees CA et al (2014) Bioavailability of capsaicin and its implications for drug delivery. J Control Release 196:96–105
Saria A, Skofitsch G, Lembeck F (1982) Distribution of capsaicin in rat tissues after systemic administration. J Pharm Pharmacol 34(4):273–275
Kang YS, Kim JM (1999) Permeability of a capsaicin derivative, [14C]DA-5018 to blood-brain barrier corrected with HPLC method. Arch Pharm Res 22(2):165–172
Schytz HW, Birk S, Wienecke T, Kruuse C, Olesen J, Ashina M (2009) PACAP38 induces migraine-like attacks in patients with migraine without aura. Brain 132(Pt 1):16–25
Amin FM, Asghar MS, Guo S, Hougaard A, Hansen AE, Schytz HW et al (2012) Headache and prolonged dilatation of the middle meningeal artery by PACAP38 in healthy volunteers. Cephalalgia 32(2):140–149
Ghanizada H, Al-Karagholi MAM, Arngrim N, Ghanizada M, Larsson HBW, Amin FM et al (2019) Effect of pituitary adenylate cyclase-activating polypeptide-27 on cerebral hemodynamics in healthy volunteers: A 3T MRI study. Peptides 121:170134
Ghanizada H, Al-Karagholi MAM, Arngrim N, Olesen J, Ashina M (2020) PACAP27 induces migraine-like attacks in migraine patients. Cephalalgia 40(1):57–67
Amin FM, Schytz HW (2018) Transport of the pituitary adenylate cyclase-activating polypeptide across the blood-brain barrier: implications for migraine. J Headache Pain 19(1):35
Grände G, Nilsson E, Edvinsson L (2013) Comparison of responses to vasoactive drugs in human and rat cerebral arteries using myography and pressurized cerebral artery method. Cephalalgia 33(3):152–159
Birk S, Sitarz JT, Petersen KA, Oturai PS, Kruuse C, Fahrenkrug J et al (2007) The effect of intravenous PACAP38 on cerebral hemodynamics in healthy volunteers. Regul Pept 140(3):185–191
Mori T, Kawashima T, Beppu Y, Takagi K (1994) Histamine release induced by pituitary adenylate cyclase activating polypeptide from rat peritoneal mast cells. Arzneimittelforschung 44(9):1044–1046
Krabbe AA, Olesen J (1980) Headache provocation by continuous intravenous infusion of histamine. Clinical results and receptor mechanisms. Pain 8(2):253–259
Vollesen LH, Guo S, Andersen MR, Ashina M (2019) Effect of the H1-antihistamine clemastine on PACAP38 induced migraine. Cephalalgia 39(5):597–607
Wienholtz NKF, Christensen CE, Zhang DG, Coskun H, Ghanizada H, Al-Karagholi MAM et al (2021) Early treatment with sumatriptan prevents PACAP38-induced migraine: a randomised clinical trial. Cephalalgia 41(6):731–748
Vollesen ALH, Amin FM, Ashina M (2018) Targeted pituitary adenylate cyclase-activating peptide therapies for migraine. Neurotherapeutics 15(2):371–376
Ashina M, Doležil D, Bonner JH, Zhou L, Klatt J, Picard H et al (2021) A phase 2, randomized, double-blind, placebo-controlled trial of AMG 301, a pituitary adenylate cyclase-activating polypeptide PAC1 receptor monoclonal antibody for migraine prevention. Cephalalgia 41(1):33–44
Edvinsson L, Tajti J, Szalárdy L, Vécsei L (2018) PACAP and its role in primary headaches. J Headache Pain 19(1):21
Guo S, Jansen-Olesen I, Olesen J, Christensen SL (2023) Role of PACAP in migraine: an alternative to CGRP? Neurobiol Dis 176:105946
Rubio-Beltrán E, Correnti E, Deen M, Kamm K, Kelderman T, Papetti L et al (2018) PACAP38 and PAC1 receptor blockade: a new target for headache? J Headache Pain 19(1):64
White CM, Ji S, Cai H, Maudsley S, Martin B (2010) Therapeutic potential of vasoactive intestinal peptide and its receptors in neurological disorders. CNS Neurol Disord Drug Targets 9(5):661–666
Sokolov AY, Osipchuk AV, Skiba IB, Amelin AV (2022) The role of pituitary adenylate cyclase-activating polypeptide and vasoactive intestinal peptide in migraine pathogenesis. Neurochem J 16(1):31–38
Cernuda-Morollón E, Martínez-Camblor P, Alvarez R, Larrosa D, Ramón C, Pascual J (2015) Increased VIP levels in peripheral blood outside migraine attacks as a potential biomarker of cranial parasympathetic activation in chronic migraine. Cephalalgia 35(4):310–316
Tajti J, Szok D, Majláth Z, Tuka B, Csáti A, Vécsei L (2015) Migraine and neuropeptides. Neuropeptides 52:19–30
Hansen JM, Sitarz J, Birk S, Rahmann AM, Oturai PS, Fahrenkrug J et al (2006) Vasoactive intestinal polypeptide evokes only a minimal headache in healthy volunteers. Cephalalgia 26(8):992–1003
Pellesi L, Al-Karagholi MAM, Chaudhry BA, Lopez CL, Snellman J, Hannibal J et al (2020) Two-hour infusion of vasoactive intestinal polypeptide induces delayed headache and extracranial vasodilation in healthy volunteers. Cephalalgia 40(11):1212–1223
Rahmann A, Wienecke T, Hansen JM, Fahrenkrug J, Olesen J, Ashina M (2008) Vasoactive intestinal peptide causes marked cephalic vasodilation, but does not induce migraine. Cephalalgia 28(3):226–236
Pellesi L, Al-Karagholi MAM, De Icco R, Coskun H, Elbahi FA, Lopez-Lopez C et al (2021) Effect of vasoactive intestinal polypeptide on development of migraine headaches. JAMA Netw Open 4(8):e2118543
Graham JR, Wolff HG (1938) Mechanism of migraine headache and action of ergotamine tartrate. Arch NeurPsych 39(4):737–763
Olesen J, Burstein R, Ashina M, Tfelt-Hansen P (2009) Origin of pain in migraine: evidence for peripheral sensitisation. Lancet Neurol 8(7):679–690
Hay DL (2017) Amylin. Headache 57(Suppl 2):89–96
Martínez A, Weaver C, López J, Bhathena SJ, Elsasser TH, Miller MJ et al (1996) Regulation of insulin secretion and blood glucose metabolism by adrenomedullin. Endocrinology 137(6):2626–2632
Hieronymus L, Griffin S (2015) Role of amylin in Type 1 and Type 2 diabetes. Diabetes Educ 41(1 Suppl):47S-56S
Ryan GJ, Jobe LJ, Martin R (2005) Pramlintide in the treatment of type 1 and type 2 diabetes mellitus. Clin Ther 27(10):1500–1512
Heise T, Heinemann L, Heller S, Weyer C, Wang Y, Strobel S et al (2004) Effect of pramlintide on symptom, catecholamine, and glucagon responses to hypoglycemia in healthy subjects. Metabolism 53(9):1227–1232
Ghanizada H, Al-Karagholi MAM, Walker CS, Arngrim N, Rees T, Petersen J et al (2021) Amylin analog pramlintide induces migraine-like attacks in patients. Ann Neurol 89(6):1157–1171
Irimia P, Martínez-Valbuena I, Mínguez-Olaondo A, Domínguez-Vivero C, Sánchez-Arias JA, Martínez-Vila E et al (2021) Interictal amylin levels in chronic migraine patients: a case-control study. Cephalalgia 41(5):604–612
Pan KS, Siow A, Hay DL, Walker CS (2020) Antagonism of CGRP signaling by Rimegepant at two receptors. Front Pharmacol 20(11):1240
Cour SHL, La Cour SH, Juhler K, Kogelman LJA, Olesen J, Klærke DA et al (2022) Characterization of erenumab and rimegepant on calcitonin gene-related peptide induced responses in Xenopus Laevis oocytes expressing the calcitonin gene-related peptide receptor and the amylin-1 receptor. J Headache Pain 23:59
Garelja ML, Walker CS, Hay DL (2022) CGRP receptor antagonists for migraine. Are they also AMY1 receptor antagonists? Br J Pharmacol 179(3):454–459
Hay DL, Garelja ML, Poyner DR, Walker CS (2018) Update on the pharmacology of calcitonin/CGRP family of peptides: IUPHAR Review 25. Br J Pharmacol 175(1):3–17
Gibbons C, Dackor R, Dunworth W, Fritz-Six K, Caron KM (2007) Receptor activity-modifying proteins: RAMPing up adrenomedullin signaling. Mol Endocrinol 21(4):783–796
Walker CS, Hay DL (2013) CGRP in the trigeminovascular system: a role for CGRP, adrenomedullin and amylin receptors? Br J Pharmacol 170(7):1293–1307
Schönauer R, Els-Heindl S (2017) Adrenomedullin–new perspectives of a potent peptide hormone. J Pept Sci 23(7–8):472–485
Wong HK, Cheung TT, Cheung BMY (2012) Adrenomedullin and cardiovascular diseases. JRSM Cardiovasc Dis 1(5):cvd.2012.012003
Huang H, Wang M, Hong Y (2019) Intrathecal administration of adrenomedullin induces mechanical allodynia and neurochemical changes in spinal cord and DRG. Neurosci Lett 18(690):196–201
Ghanizada H, Al-Karagholi MAM, Arngrim N, Mørch-Rasmussen M, Walker CS, Hay DL et al (2021) Effect of adrenomedullin on migraine-like attacks in patients with migraine: a randomized crossover study. Neurology 96(20):e2488–e2499
Petersen KA, Birk S, Kitamura K, Olesen J (2009) Effect of adrenomedullin on the cerebral circulation: relevance to primary headache disorders. Cephalalgia 29(1):23–30
Akcali A, Geyik S, Dogru I, Dogru A, Yurekli M, Neyal A et al (2016) Plasma adrenomedullin levels in patients with migraine during naturel attack and attack free period. Med Sci Discov 3(4):153–158
Brain SD, Grant AD (2004) Vascular actions of calcitonin gene-related peptide and adrenomedullin. Physiol Rev 84(3):903–934
Hinson JP, Kapas S, Smith DM (2000) Adrenomedullin, a multifunctional regulatory peptide. Endocr Rev 21(2):138–167
Bhakta M, Vuong T, Taura T, Wilson DS, Stratton JR, Mackenzie KD (2021) Migraine therapeutics differentially modulate the CGRP pathway. Cephalalgia 41(5):499–514
Ashina H, Christensen RH, Ashina M (2022) Provoked versus spontaneous migraine attacks: pathophysiological similarities and differences. J Headache Pain 23(1):87
Ashina M (2020) Migraine. N Engl J Med 383(19):1866–1876
Ashina M, Hansen JM, Olesen J (2017) Human models of migraine—short-term pain for long-term gain. Nat Rev Neurol 13(12):713–724
Christiansen I, Daugaard D, Lykke Thomsen L, Olesen J (2000) Glyceryl trinitrate induced headache in migraineurs - relation to attack frequency. Eur J Neurol 7(4):405–411
Ben Aissa M, Tipton AF, Bertels Z, Gandhi R, Moye LS, Novack M et al (2018) Soluble guanylyl cyclase is a critical regulator of migraine-associated pain. Cephalalgia 38(8):1471–1484
Lassen LH, Ashina M, Christiansen I, Ulrich V, Grover R, Donaldson J et al (1998) Nitric oxide synthase inhibition: a new principle in the treatment of migraine attacks. Cephalalgia 18(1):27–32
Sarchielli P, Floridi A, Mancini ML, Rossi C, Coppola F, Baldi A et al (2006) NF-kappaB activity and iNOS expression in monocytes from internal jugular blood of migraine without aura patients during attacks. Cephalalgia 26(9):1071–1079
Palmer JE, Guillard FL, Laurijssens BE, Wentz AL (2009) A randomised, single-blind, placebo-controlled, adaptive clinical trial of GW274150, a selective iNOS inhibitor, in the treatment of acute migraine. Cephalalgia 29(1):124
Høivik HO, Laurijssens BE, Harnisch LO, Twomey CK, Dixon RM, Kirkham AJT et al (2010) Lack of efficacy of the selective iNOS inhibitor GW274150 in prophylaxis of migraine headache. Cephalalgia 30(12):1458–1467
Bhatt DK, Gupta S, Jansen-Olesen I, Andrews JS, Olesen J (2013) NXN-188, a selective nNOS inhibitor and a 5-HT1B/1D receptor agonist, inhibits CGRP release in preclinical migraine models. Cephalalgia 33(2):87–100
Hougaard A, Hauge AW, Guo S, Tfelt-Hansen P (2013) The nitric oxide synthase inhibitor and serotonin-receptor agonist NXN-188 during the aura phase of migraine with aura: a randomized, double-blind, placebo-controlled cross-over study. Scand J Pain 4(1):48–52
Guo S, Olesen J, Ashina M (2014) Phosphodiesterase 3 inhibitor cilostazol induces migraine-like attacks via cyclic AMP increase. Brain 137(Pt 11):2951–2959
Kruse LS, Sandholdt NTH, Gammeltoft S, Olesen J, Kruuse C (2006) Phosphodiesterase 3 and 5 and cyclic nucleotide-gated ion channel expression in rat trigeminovascular system. Neurosci Lett 404(1–2):202–207
Falkenberg K, Dunga BÓÁ, Guo S, Ashina M, Olesen J (2018) Cilostazol induced migraine does not respond to sumatriptan in a double blind trial. J Headache Pain 19(1):11
Ingram SL, Williams JT (1996) Modulation of the hyperpolarization-activated current (Ih) by cyclic nucleotides in guinea-pig primary afferent neurons. J Physiol 492(Pt 1):97–106
Khan S, Deen M, Hougaard A, Amin FM, Ashina M (2018) Reproducibility of migraine-like attacks induced by phosphodiesterase-3-inhibitor cilostazol. Cephalalgia 38(5):892–903
Birk S, Kruuse C, Petersen KA, Tfelt-Hansen P, Olesen J (2006) The headache-inducing effect of cilostazol in human volunteers. Cephalalgia 26(11):1304–1309
Chrissobolis S, Sobey CG (2003) Inwardly rectifying potassium channels in the regulation of vascular tone. Curr Drug Targets 4(4):281–289
Christensen SL, Rasmussen RH, Ernstsen C, La Cour S, David A, Chaker J et al (2021) CGRP-dependent signalling pathways involved in mouse models of GTN- cilostazol- and levcromakalim-induced migraine. Cephalalgia 41(14):1413–1426
Birk S, Kruuse C, Petersen KA, Jonassen O, Tfelt-Hansen P, Olesen J (2004) The phosphodiesterase 3 inhibitor cilostazol dilates large cerebral arteries in humans without affecting regional cerebral blood flow. J Cereb Blood Flow Metab 24(12):1352–1358
Falkenberg K, Bjerg HR, Olesen J (2020) Subcutaneous sumatriptan reduces cilostazol induced headache in migraine patients. Cephalalgia 40(8):842–850
Butt JH, Rostrup E, Hansen AS, Lambertsen KL, Kruuse C (2018) Induction of migraine-like headache, but not aura, by cilostazol in patients with migraine with aura. Brain 141(10):2943–2951
Evans RW, Kruuse C (2004) Phosphodiesterase-5 inhibitors and migraine. Headache 44(9):925–926
Goldstein I, Lue TF, Padma-Nathan H, Rosen RC, Steers WD, Wicker PA (1998) Oral sildenafil in the treatment of erectile dysfunction. Sildenafil Study Group. N Engl J Med 338(20):1397–1404
Kruuse C, Thomsen LL, Birk S, Olesen J (2003) Migraine can be induced by sildenafil without changes in middle cerebral artery diameter. Brain 126(1):241–247
Kruuse C, Thomsen LL, Jacobsen TB, Olesen J (2002) The phosphodiesterase 5 inhibitor sildenafil has no effect on cerebral blood flow or blood velocity, but nevertheless induces headache in healthy subjects. J Cereb Blood Flow Metab 22(9):1124–1131
Younis S, Christensen CE, Toft NM, Søborg T, Amin FM, Hougaard A et al (2019) Investigation of distinct molecular pathways in migraine induction using calcitonin gene-related peptide and sildenafil. Cephalalgia 39(14):1776–1788
Barman SA, Zhu S, Han G, White RE (2003) cAMP activates BKCa channels in pulmonary arterial smooth muscle via cGMP-dependent protein kinase. Am J Physiol Lung Cell Mol Physiol 284(6):L1004–L1011
Jiang H, Colbran JL, Francis SH, Corbin JD (1992) Direct evidence for cross-activation of cGMP-dependent protein kinase by cAMP in pig coronary arteries. J Biol Chem 267(2):1015–1019
Christensen CE, Amin FM, Younis S, Lindberg U, de Koning P, Petersen ET et al (2019) Sildenafil and calcitonin gene-related peptide dilate intradural arteries: a 3T MR angiography study in healthy volunteers. Cephalalgia 39(2):264–273
Younis S, Hougaard A, Christensen CE, Vestergaard MB, Petersen ET, Paulson OB et al (2018) Effects of sildenafil and calcitonin gene-related peptide on brainstem glutamate levels: a pharmacological proton magnetic resonance spectroscopy study at 3.0 T. J Headache Pain 19(1):44
Kokoti L, Al-Karagholi MAM, Elbahi FA, Coskun H, Ghanizada H, Amin FM et al (2022) Effect of KATP channel blocker glibenclamide on PACAP38-induced headache and hemodynamic. Cephalalgia 42(9):846–858
Al-Karagholi MAM, Ghanizada H, Nielsen CAW, Hougaard A, Ashina M (2021) Opening of ATP sensitive potassium channels causes migraine attacks with aura. Brain 144(8):2322–2332
Al-Karagholi MA, Hakbilen CC, Ashina M (2022) The role of high-conductance calcium-activated potassium channel in headache and migraine pathophysiology. Basic Clin Pharmacol Toxicol 131(5):347–354
Pietrobon D, Moskowitz MA (2014) Chaos and commotion in the wake of cortical spreading depression and spreading depolarizations. Nat Rev Neurosci 15(6):379–393
Waliszewska-Prosół M, Nowakowska-Kotas M, Chojdak-Łukasiewicz J, Budrewicz S (2021) Migraine and sleep-an unexplained association? Int J Mol Sci 22(11):5539
Pietrobon D (2018) Ion channels in migraine disorders. Curr Opin Physio 2:98–108
Royal P, Andres-Bilbe A, Prado PÁ, Verkest C, Wdziekonski B et al (2019) Migraine-associated TRESK mutations increase neuronal excitability through alternative translation initiation and inhibition of TREK. Neuron 101(2):232–45.e6
Prado PÁ, Landra-Willm A, Verkest C, Ribera A, Chassot AA, Baron A et al (2021) TREK channel activation suppresses migraine pain phenotype. iScience. 24(9):102961
Ashina M, Hansen JM, Dunga BOá, Olesen J (2017) Human models of migraine — short-term pain for long-term gain. Nat Rev Neurol 13(12):713–724
Koide M, Syed AU, Braas KM, May V, Wellman GC (2014) Pituitary Adenylate Cyclase Activating Polypeptide (PACAP) dilates cerebellar arteries through activation of large-conductance Ca2 -activated (BK) and ATP-sensitive (KATP) K channels. J Mol Neurosci 54(3):443–450
Al-Karagholi MAM, Ghanizada H, Kokoti L, Paulsen JS, Hansen JM, Ashina M (2020) Effect of KATP channel blocker glibenclamide on levcromakalim-induced headache. Cephalalgia 40(10):1045–1054
Citak A, Kilinc E, Torun IE, Ankarali S, Dagistan Y, Yoldas H (2022) The effects of certain TRP channels and voltage-gated KCNQ/Kv7 channel opener retigabine on calcitonin gene-related peptide release in the trigeminovascular system. Cephalalgia 42(13):1375–1386
N’Gouemo P (2014) BKCa channel dysfunction in neurological diseases. Front Physiol 5(373). https://doi.org/10.3389/fphys.2014.00373
Yan J, Dussor G (2014) Ion channels and migraine. Headache 54(4):619–639
Kaczorowski GJ, Knaus HG, Leonard RJ, McManus OB, Garcia ML (1996) High-conductance calcium-activated potassium channels; Structure, pharmacology, and function. J Bioenerg Biomembr 28(3):255–267
Lee US, Cui J (2010) BK channel activation: structural and functional insights. Trends Neurosci 33(9):415–423
Akerman S, Holland PR, Lasalandra MP, Goadsby PJ (2010) Inhibition of trigeminovascular dural nociceptive afferents by Ca2 -activated K (MaxiK/BKCa) channel opening. Pain 151(1):128–136
Lu R, Lukowski R, Sausbier M, Zhang DD, Sisignano M, Schuh CD et al (2014) BKCa channels expressed in sensory neurons modulate inflammatory pain in mice. Pain 155(3):556–565
Al-Karagholi MAM, Gram C, Nielsen CAW, Ashina M (2020) Targeting BKCa channels in migraine: rationale and perspectives. CNS Drugs 34(4):325–335
Al-Karagholi MAM, Ghanizada H, Nielsen CAW, Skandarioon C, Snellman J, Lopez Lopez C et al (2020) Opening of BKCa channels alters cerebral hemodynamic and causes headache in healthy volunteers. Cephalalgia 40(11):1145–1154
Al-Karagholi MAM, Ghanizada H, Waldorff Nielsen CA, Skandarioon C, Snellman J, Lopez-Lopez C et al (2021) Opening of BKCa channels causes migraine attacks: a new downstream target for the treatment of migraine. Pain 162(10):2512–2520
Rau JC, Dodick DW (2019) Other preventive anti-migraine treatments: ACE inhibitors, ARBs, calcium channel blockers, serotonin antagonists, and NMDA receptor antagonists. Curr Treat Options Neurol 21(4):17
Amery WK (1983) Flunarizine, a calcium channel blocker: a new prophylactic drug in migraine. Headache 23(2):70–74
Gelmers HJ (1985) Calcium-channel blockers in the treatment of migraine. Am J Cardiol 55(3):B139–B143
Ayata C, Jin H, Kudo C, Dalkara T, Moskowitz MA (2006) Suppression of cortical spreading depression in migraine prophylaxis. Ann Neurol 59(4):652–661
Catterall WA (2011) Voltage-gated calcium channels. Cold Spring Harb Perspect Biol 3(8):a003947–a003947
Zamponi GW, Striessnig J, Koschak A, Dolphin AC (2015) The physiology, pathology, and pharmacology of voltage-gated calcium channels and their future therapeutic potential. Pharmacol Rev 67(4):821–870
Ducros A, Denier C, Joutel A, Cecillon M, Lescoat C, Vahedi K et al (2001) The clinical spectrum of familial hemiplegic migraine associated with mutations in a neuronal calcium channel. N Engl J Med 345(1):17–24
Ophoff RA, Terwindt GM, Vergouwe MN, van Eijk R, Oefner PJ, Hoffman SMG et al (1996) Familial hemiplegic migraine and episodic ataxia Type-2 are caused by mutations in the Ca2 channel gene CACNL1A4. Cell 87(3):543–552
Pietrobon D (2013) Calcium channels and migraine. Biochim Biophys Acta 1828(7):1655–1665
Hans M, Luvisetto S, Williams ME, Spagnolo M, Urrutia A, Tottene A et al (1999) Functional consequences of mutations in the human α1Acalcium channel subunit linked to familial hemiplegic migraine. J Neurosci 19(5):1610–1619
Tottene A, Conti R, Fabbro A, Vecchia D, Shapovalova M, Santello M et al (2009) Enhanced excitatory transmission at cortical synapses as the basis for facilitated spreading depression in CaV2.1 knockin migraine mice. Neuron 61(5):762–773
Knight YE, Bartsch T, Kaube H, Goadsby PJ (2002) P/Q-type calcium-channel blockade in the periaqueductal gray facilitates trigeminal nociception: a functional genetic link for migraine? J Neurosci 22(5):RC213–RC213
Chan KY, Labastida-Ramírez A, Ramírez-Rosas MB, Labruijere S, Garrelds IM, Danser AH et al (2019) Trigeminovascular calcitonin gene-related peptide function in Cacna1a R192Q-mutated knock-in mice. J Cereb Blood Flow Metab 39(4):718–729
Wu ZZ, Li DP, Chen SR, Pan HL (2009) Aminopyridines potentiate synaptic and neuromuscular transmission by targeting the voltage-activated calcium channel β subunit. J Biol Chem 284(52):36453–36461
Pastor PDH, Du X, Fazal S, Davies AN, Gomez CM (2018) Targeting the CACNA1A IRES as a treatment for spinocerebellar ataxia Type 6. The Cerebellum 17(1):72–77
Venkatachalam K, Montell C (2007) TRP channels. Annu Rev Biochem 76(1):387–417
Clapham DE, Runnels LW, Strübing C (2001) The TRP ion channel family. Nat Rev Neurosci 2(6):387–396
Dussor G, Yan J, Xie JY, Ossipov MH, Dodick DW, Porreca F (2014) Targeting TRP channels for novel migraine therapeutics. ACS Chem Neurosci 5(11):1085–1096
Akerman S, Kaube H, Goadsby PJ (2003) Vanilloid type 1 receptors (VR1) on trigeminal sensory nerve fibres play a minor role in neurogenic dural vasodilatation, and are involved in capsaicin-induced dural dilation. Br J Pharmacol 140(4):718–724
Kárai LJ, Russell JT, Iadarola MJ, Oláh Z (2004) Vanilloid receptor 1 regulates multiple calcium compartments and contributes to Ca2 -induced Ca2 release in sensory neurons. J Biol Chem 279(16):16377–16387
Evans MS, Cheng X, Jeffry JA, Disney KE, Premkumar LS (2012) Sumatriptan inhibits TRPV1 channels in trigeminal neurons. Headache 52(5):773–784
Numazaki M, Tominaga T, Takeuchi K, Murayama N, Toyooka H, Tominaga M (2003) Structural determinant of TRPV1 desensitization interacts with calmodulin. Proc Natl Acad Sci 100(13):8002–8006
Diamond S, Freitag F, Phillips SB, Bernstein JE, Saper JR (2000) Intranasal civamide for the acute treatment of migraine headache. Cephalalgia 20(6):597–602
Fusco BM, Barzoi G, Agrò F (2003) Repeated intranasal capsaicin applications to treat chronic migraine. Br J Anaesth 90(6):812
Nassini R, Materazzi S, Vriens J, Prenen J, Benemei S, De Siena G et al (2012) The “headache tree” via umbellulone and TRPA1 activates the trigeminovascular system. Brain 135(2):376–390
Materazzi S, Benemei S, Fusi C, Gualdani R, De Siena G, Vastani N et al (2013) Parthenolide inhibits nociception and neurogenic vasodilatation in the trigeminovascular system by targeting the TRPA1 channel. Pain 154(12):2750–2758
Shibata M, Tang C (2021) Implications of transient receptor potential cation channels in migraine pathophysiology. Neurosci Bull 37(1):103–116
Iannone LF, De Logu F, Geppetti P, De Cesaris F (2022) The role of TRP ion channels in migraine and headache. Neurosci Lett 18(768):136380
Cadiou H, Studer M, Jones NG, Smith ESJ, Ballard A, McMahon SB et al (2007) Modulation of acid-sensing ion channel activity by nitric oxide. J Neurosci 27(48):13251–13260
Holton CM, Strother LC, Dripps I, Pradhan AA, Goadsby PJ, Holland PR (2020) Acid-sensing ion channel 3 blockade inhibits durovascular and nitric oxide-mediated trigeminal pain. Br J Pharmacol 177(11):2478–2486
Dussor G (2015) ASICs as therapeutic targets for migraine. Neuropharmacology 94:64–71
Karsan N, Gonzales EB, Dussor G (2018) Targeted acid-sensing ion channel therapies for migraine. Neurotherapeutics 15(2):402–414
Clement A, Guo S, Jansen-Olesen I, Christensen SL (2022) ATP-sensitive potassium channels in migraine: translational findings and therapeutic potential. Cells 11(15):2406
Thomsen LL, Kruuse C, Iversen HK, Olesen J (1994) A nitric oxide donor (nitroglycerin) triggers genuine migraine attacks. Eur J Neurol 1(1):73–80
Al-Karagholi MAM, Hansen JM, Guo S, Olesen J, Ashina M (2019) Opening of ATP-sensitive potassium channels causes migraine attacks: a new target for the treatment of migraine. Brain 142(9):2644–2654
Chesnut CH 3rd, Azria M, Silverman S, Engelhardt M, Olson M, Mindeholm L (2008) Salmon calcitonin: a review of current and future therapeutic indications. Osteoporos Int 19(4):479–491
Micieli G, Cavallini A, Martignoni E, Covelli V, Facchinetti F, Nappi G (1988) Effectiveness of salmon calcitonin nasal spray preparation in migraine treatment. Headache 28(3):196–200
Kilinc E, Dagistan Y, Kukner A, Yilmaz B, Agus S, Soyler G et al (2018) Salmon calcitonin ameliorates migraine pain through modulation of CGRP release and dural mast cell degranulation in rats. Clin Exp Pharmacol Physiol 45(6):536–546
Sonne N, Karsdal MA, Henriksen K (2021) Mono and dual agonists of the amylin, calcitonin, and CGRP receptors and their potential in metabolic diseases. Mol Metab 46:101109. https://doi.org/10.1016/j.molmet.2020.101109
Lindblad M, Hougaard A, Amin FM, Ashina M (2017) Can migraine aura be provoked experimentally? A systematic review of potential methods for the provocation of migraine aura. Cephalalgia 37(1):74–88
Cheung BMY, Li CYY, Wong LYF (2004) Adrenomedullin: its role in the cardiovascular system. Semin Vasc Med 4(2):129–134
Al-Hassany L, Haas J, Piccininni M, Kurth T, Maassen Van Den Brink A, Rohmann JL (2020) Giving researchers a headache - sex and gender differences in migraine. Front Neurol 11:549038
Acknowledgements
Not applicable.
Funding
The publication fee for the present article was granted by a full waiver.
Author information
Authors and Affiliations
Consortia
Contributions
All authors on behalf of the European Headache Federation contributed equally to the drafting and critical revisions of the manuscript. Linda Al-Hassany took the lead in writing, drafting and revising the manuscript. Faisal Mohammad Amin supervised the entire process and designed the concept of the review. All authors read and gave final approval and accepted responsibility for the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
Linda Al-Hassany, Deirdre M. Boucherie, Hannah Creeney, Ruben W.A.van Drie, Fatemeh Farham, Silvia Favaretto, Grangeon Lou, Hannah Lyons, Karol Marschollek, Dilara Onan, Umberto Pensato, Emily Stanyer, Marta Waliszewska-Prosół, Hui Zhou Chen declare no conflict of interest. Cédric Gollion reports speaker fees from Teva, Novartis, Lundbeck and Lilly. Wietse Wiels declares to have received an FWO Vlaanderen Fundamental Research scholarship. Faisal Mohammad Amin has received Honoraria or personal fees from Pfizer, Teva, Novartis, Lundbeck and Eli Lilly for lecturing or participating in advisory boards; is principal investigator for phase IV trials sponsored by Novartis and by Teva; serves as president of Danish Headache Society and board member of the European Headache Federation; serves as associate editor for Acta Neurol Scand, Front Neurol, Front Res Pain, and Headache Medicine; serves as junior associate editor for Cephalalgia and Cephalalgia Reports; member of the editorial board of J Headache Pain.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Al-Hassany, L., Boucherie, D.M., Creeney, H. et al. Future targets for migraine treatment beyond CGRP. J Headache Pain 24, 76 (2023). https://doi.org/10.1186/s10194-023-01567-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10194-023-01567-4