
Abstract
Food-limited growth of larval fish, defined as growth rates lower than observed in other habitats or from laboratory experiments at a given temperature, is rarely reported in field studies. This would imply that either larval fishes are living in an environment characterized by plenty of food, that nutritional condition selective mortality (i.e., eliminating the weak) is very strong, or this impression is caused by misinterpretation of data concerning e.g., poor taxonomical resolution of potential prey items, i.e., total potential prey abundance is high, but positively selected food is actually scarce. We analyzed RNA:DNA derived growth rates of herring larvae (Clupea harengus L.) and taxonomically differentiated prey field data of six consecutive spring seasons from the Kiel Canal, an artificial waterway in northern Germany, in order to test if food-limited growth in larval fish can occur recurrently in coastal habitats. In all years analyzed, larval growth rates decreased simultaneously with prey abundance at the end of each larval season. Furthermore, larval growth rates were observed to be lower than mean growth rates observed in another herring larvae nursery area at temperatures above 15 °C. Asymptotic relationships between prey abundance and larval growth rates were observed, further supporting the hypothesis of food-limitation. As larval growth was best explained by the abundance of the numerically dominant calanoid copepod Eurytemora affinis, the paramount importance of the dominant prey item is highlighted. We conclude that food limitation can be a severe and re-occurring issue for larval fish in coastal habitats, and that certain prey items play a crucial role in determining larval growth rates, and therefore potentially recruitment.
Similar content being viewed by others

Background
Over a century ago Hjort [35] hypothesized that the nutrition of first feeding larvae is an important recruitment-driving factor. Since then, generations of fishery scientists worked on this issue, but it took decades until they started “emerging from Hjort’s shadow” [39], interpreting the issue of larval nutrition in a broader context. This includes biotic as well as abiotic factors influencing the nutritional situation for fish larvae, and focusing not only on the time of first feeding (e.g., [3, 10, 19, 50]).
Nevertheless, the issue of food limitation in the field has rarely been examined in larval growth analyses, and remains a matter of debate. Numerous field studies reported that only small proportions of marine fish larvae starve (e.g., [15, 18, 21, 67]). Consequently, it was recently hypothesized that intermediate or low prey abundance indirectly affect larval recruitment by enforcing more active foraging and, thus, higher risk of predation by easier detection by predators [43] rather than starvation itself. Accordingly, starvation of larvae would rather be exceptional at very low levels of prey abundance. For example, studies on clupeoid fish larvae showed that lowest threshold values for early survival are ~4–5 copepod nauplii L−1, and that larval growth was increasingly positively affected until upper threshold concentrations of ~50 copepod nauplii L−1 ([55], and references therein). These concentrations are strongly affected by prevailing temperatures, as fish metabolism is temperature dependent [55]. Relatively few studies found evidence that growth rates of marine fish larvae are occasionally food limited [9, 30, 40, 65]. Such observations, however, seem to be exceptions, since food limitation only occurred in certain situations, e.g., in a single year, or in certain water depths, but not on a regular basis.
All the above cited studies were conducted off-shore, providing relatively stable environmental conditions in terms of salinity and showing relatively slow temperature changes. In contrast, conditions in coastal habitats such as lagoons and estuaries are highly variable, including changing salinities and subsequent changes in prey composition (e.g., [8, 29, 42, 61, 64]). Small waterbodies and low water depths are typical for such areas, leading to strong short-term changes in temperature during spring time (e.g., [24, 52]), potentially with qualitative and quantitative implications for prey availability [2].
In previous studies we observed first indications for food limited growth rates of larval herring in the Kiel Canal [53, 54], an artificial waterway in northern Germany, with a limited data set. In this follow-up study we analyzed a 6-years-time series including RNA:DNA derived growth rates of 1177 larval herring and taxonomically differentiated prey field data in order to investigate potential reasons for this phenomenon and to test the hypothesis that food-limited growth of larval fish can occur recurrently in coastal habitats and at inter-annually varying prey levels.
Methods
Sampling
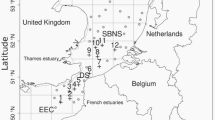
Time series analyses of biochemically derived larval herring growth rates along with prey field and temperature data of six consecutive spring seasons (2007–2012) in the Kiel Canal at a station 13 km inland to the open Baltic Sea (54°20′45N, 9°57′02E, Fig. 1) were conducted. The Kiel Canal has an average width of about 100 m, and is characterized by a mean depth of about 11 m. The water column is constantly mixed due to the heavy shipping traffic (highest frequency of container ship use in the world), and the salinity is low, ranging from 5.5 to 11.5. The Kiel Canal is a suitable habitat to examine prey effects on larval fish, due to its very confined space and weak currents. Larval fish and zooplankton were sampled on a weekly basis. This sampling frequency was chosen according to the response time of the biochemical larval growth indicator used (RNA:DNA), which is several days to a week [17]. Vertical profiles of salinity and temperature were taken (CTD 48 M, Sea & Sun Technology GmbH, Trappenkamp, Germany). To analyze the prey field, a vertical haul was carried out with a WP2-net (200 µm mesh size; the issue of losing the nauplii by using this mesh size is referred to in the discussion) and fixed with 4 % borax buffered formalin solution. Larval herring were sampled by using a bongo net with 500 µm mesh size which was retrieved in an oblique haul with a speed of 2.5 knots. Two hauls were carried out: the first one was preserved with 4 % borax buffered formalin solution and used for the determination of larval abundance. The second one was taken to sample larvae for RNA:DNA analysis. This sample was cooled with crushed ice immediately, and all larvae were sorted out within 30 min after the haul. The larvae were frozen on board at −20 °C, and stored after the return to the institute (about 2–3 h after the haul) at −80 °C. To define and detect food-limitation, larval growth data from Kiel Canal were compared to larval growth data from another important habitat for western Baltic Sea spring spawning herring, the Greifswalder Bodden (Bodden is the German term for an enclosed, shallow and low saline bay at the Baltic Sea coast; Fig. 1; for further information on this site, please see [54]) with samples collected in the spring seasons 2010–2012. The sampling procedure was the same as in the Kiel Canal, apart from the fact that larvae were frozen in liquid nitrogen on board.

Location of the sampling sites in northern Germany: A Kiel Canal and B Greifswalder Bodden
Larval standard lengths were measured to the lower 0.1 mm. Prior to RNA:DNA analysis, the larvae were freeze dried for 24 h (Christ Alpha 1-4 LSC freeze dryer) and then weighed to the nearest 0.1 µg (Sartorius microbalance SC2).
RNA:DNA analysis and instantaneous growth rates (Gi)
RNA and DNA concentrations of whole individual larvae were measured. A detailed description of the analysis is given by Clemmesen [16, 17]. Larvae caught in 2011 however, were defatted before analyzing RNA:DNA and analyzed according to Malzahn et al. [48, 49] to analyze the effects of essential fatty acids on larval growth rates [54]. Comparison with the classical approach by Clemmesen [16, 17] showed no significant differences between RNA:DNA of defatted and non-defatted larvae. We analyzed a minimum of 10 larvae and a maximum of 50 larvae, but on average 20–30 larvae per sampling day. Two size classes were used for further analyses: Length classes smaller and larger than 14 mm were chosen. At 14 mm the dorsal fin is clearly protruded (own observations and [23]). This fin differentiation is assumed as an indicator for an ethological change, due to enhanced swimming and therefore hunting capabilities.
The RNA:DNA ratios were converted to standardized RNA:DNA ratios (sRD; [13]) and used to calculate dry weight related instantaneous growth rates (Gi) of the larvae [11] based on the following relationship:
with Gi being the instantaneous growth rate, sRD the standardized RNA:DNA ratio and T the temperature (°C) determined at the sampling time. Interpretation of Gi-values has to be done in the way that a value of 0 means no growth at all and a value of 1 would be a doubling of the weight of the larva per day. Larval growth data were used in different ways for regression analyses with prey abundance: (1) as individual larval growth data without any size differentiation, (2) mean per sampling day (including all larvae analysed at a given date) and (3) seasonal mean growth rates. The different approaches were chosen in order to (1) display individual variability, (2) visualize sampling date variability, and (3) test for inter-annual variability, which would be completely masked by taking individual data due to the high range of variability of both larval growth data and prey abundance throughout the course of the seasons.
Mesozooplankton analysis
Mesozooplankton samples were divided with a Kott plankton splitter [44] up to the point, where at least 100 individuals (including copepodite and adult stages, excluding nauplii) of the most abundant copepod species were found in a single subsample. If less than 100 individuals were counted, additional subsamples were analyzed until at least 100 individuals were reached. All taxa were recorded and distinguished to the lowest possible taxonomic level using the taxonomic guide by Sars [59].
Further analyses were restricted to calanoid copepods and cirriped nauplii, as these organisms were shown being the major prey items for larval herring in the Kiel Canal [22].
Defining food-limitation
To define and detect food-limitation, negative effects of increasing temperatures on larval growth needed to be excluded. In order to do so, larval herring growth data (means per sampling date) from the Kiel Canal were compared to growth data from the Greifswalder Bodden, where growth rates further increased at temperatures above 15 °C, indicating that the optimal temperature was not reached yet. Trend lines were fitted to larval growth versus temperature data from the Greifswalder Bodden (2010–2012) and the Kiel Canal (2007–2012), respectively, including 95 % confidence intervals (CI). When the upper 95 % CI of larval herring growth data from the Kiel Canal was below the lower 95 % CI of larval growth data from the Greifswalder Bodden, these larvae were assigned to be food-limited, as their growth rates were lower than possible.
Statistics
The statistic package Statistica (version 6) was used to perform statistical analyses. For all statistics, the significance level was set to α = 0.05. By using the Shapiro–Wilk- and the Levene-Test, data were analyzed for normal distribution and homogeneity of variances, respectively. To test for differences in larval growth rates during each spring season, one-way analyses of variance (ANOVA) were conducted and Tukey honestly significant difference (HSD) tests were used for post hoc comparison. Growth data used for this approach were not further differentiated with respect to size classes, but used to generally test for significant declines in larval growth rates towards the end of the season in the Kiel Canal. Mann–Whitney-U-tests were used to test for significant differences between larvae smaller (range 4.8–13.9 mm) and larger than 14 mm (range 14–20 mm) at given dates. In order to test for prey effects on larval growth, individual larval growth data and mean growth per sampling date were correlated to prey abundance observed at that sampling date. Additionally, seasonal mean larval growth data and seasonal mean prey abundance were correlated. As the 2009-season was temporally not fully covered, that year was excluded from the seasonal mean approach. As temperature is included as a term in the multi-species growth model used [11], larval Gi could potentially rather reflect temperature changes than changes in the larvae's nutritional condition. To test in how far larval Gi was determined by the nutritional condition of the larvae, a regression analysis between Gi and sRD was performed. Further, generalized additive models (GAMs) were performed in order to test for explained variability of sRD for both Eurytemora abundance and temperature in combination. This was only possible for larvae <14 mm, as Eurytemora abundance and temperature were significantly negative correlated for the data set of larvae >14 mm. This was based on the fact that larger larvae only occurred from the middle of the season onwards. At that time point, Eurytemora abundances were always at their highest levels, and decreased thereafter.
Results
The zooplankton community in the Kiel Canal was dominated by Eurytemora affinis, Acartia tonsa and cirriped nauplii (Table 1). Eurytemora affinis was the numerically dominant species among the copepods found in the Kiel Canal (Fig. 2; Table 1). On average Eurytemora made up 79 % of the total copepod abundance across all years investigated. Total copepod abundance (all species named in Table 1, including Eurytemora) and Eurytemora abundance alone correlated significantly throughout the investigated seasons (analyzed time frame and sampling frequency is the same as for herring larvae growth analyses; y = 0.9568x − 2859.9, R2 = 0.97, p < 0.001, N = 43).

Inter-annual variability of the copepod abundance in the Kiel Canal and the contribution of Eurytemora affinis. Other copepods comprise calanoid copepods of the genera Acartia, Pseudocalanus, Temora and Centropages. Seasonal means are displayed
Results from regression analyses were significant between larval herring seasonal mean Gi and seasonal mean Eurytemora abundance (R2 = 0.9, p < 0.05, N = 5) and insignificant for the other taxa (p > 0.05; Acartia tonsa R2 = 0.34, Pseudocalanus elongatus R2 = 0.71, Temora longicornis R2 = 0.39, Centropages hamatus R2 = 0.38, cirriped nauplii R2 = 0.56). Therefore, further analyses of prey effects on larval growth rates concentrated on Eurytemora (all stages from CI to CVI).
Generally, significant positive relationships between larval instantaneous growth rates and Eurytemora abundance were observed (Fig. 3a–c). Individual larval growth rates showed a high variability at given Eurytemora abundances (Fig. 3a, p < 0.01, N = 1177), mirrored in a relatively low explained variability of 25 %. Explained variability of larval growth rates increased to 31 % using weekly means of larval growth rates (Fig. 3b, p < 0.01, N = 43), while 90 % of larval growth variability was explained using seasonal means (Fig. 3c, p < 0.05, N = 5).

Relationships between seasonal mean larval herring growth rates (including large and small larvae) versus the dominant copepod species in the Kiel Canal, Eurytemora affinis. a Individual larval instantaneous growth rates [Gi (d−1); N = 1177)] versus the abundance of Eurytemora affinis; data from the years 2007–2012. b Weekly mean larval growth rates versus the Eurytemora abundance observed at the given sampling date (N = 44); data from the years 2007–2012. c Seasonal mean larval growth data versus seasonal mean Eurytemora abundance (N = 5). Error bars indicate standard deviations. Here, the season 2009 is not included, as this season was not fully covered by the sampling
Over the years, a recurring seasonal pattern was observed in the Kiel Canal: during the early season, Eurytemora abundances increased or were on a high level and larval growth rates increased simultaneously (Fig. 4). Larvae larger than 14 mm grew significantly faster than smaller larvae in 2/3 of the cases (Mann–Whitney-U-test, p < 0.05, N = 13), but in 1/3 of the cases (N = 6) growth rates of small and large larvae were not significantly different (Fig. 4). 83.3 % (N = 5) of the insignificant cases were observed at food limited situations. Temporal trends did not differ between large and small larvae.

Seasonal course of larval growth rates and prey availability from 2007 to 2012. Growth rates of herring larvae smaller and larger than 14 mm were differentiated. Circles indicate the median of larval growth rates, error bars show 95 % confidence intervals. Stars denote significant differences of growth rates of small and large larvae. Note the different scales of Eurytemora abundances
In the late season at temperatures above 15 °C Eurytemora abundances decreased and remained at very low levels (Fig. 5), and larval growth rates decreased significantly (ANOVA, p < 0.05). Up to 15 °C larval growth rates in the Kiel Canal were similar to the mean larval growth rates observed in the Greifswalder Bodden in the seasons 2010–2012 (Fig. 6). Thereafter, the upper 95 % confidence interval of the Kiel Canal-larval growth data was constantly below the lower 95 % confidence interval of the Greifswalder Bodden-larval growth data.

Relationship between Eurytemora abundance and temperature (N = 43). Different symbols indicate different years

Definition of food-limitation in the present study. Instantaneous larval growth rates (Gi) versus temperature in the Kiel Canal. Means of sampling dates are displayed (N = 43). The grey trend line allocates to Kiel Canal data and dotted lines indicate 95 % interval. The black regression line allocates to Greifswalder Bodden larval growth data in the seasons 2010–2012 (N = 23), the dashed lines, are indicating 95 % confidence intervals (CI). Larval growth rates were defined to be food-limited, when the upper 95 % CI from Kiel Canal was constantly below the lower 95 % confidence interval of larval growth rates from the Greifswalder Bodden, as their growth performance was lower than possible
82 % of larval Gi variability (all size groups) was explained by the standardized RNA:DNA ratio (sRD), indicating major effects of larval nutritional condition on larval Gi (Fig. 7, p < 0.001, N = 1177). Further statistical analyses on the effects of prey availability and temperature on sRD variability could only be performed on larvae <14 mm due to an autocorrelation between prey and temperature for the period when large larvae occurred in the Kiel Canal. Results from a generalized additive model (GAM) analysis showed that sRD for larvae <14 mm was explained to 44.4 % by Eurytemora abundance and temperature. However, when using Eurytemora exclusively as an independent variable, explained variability of sRD was 40.9 % (Fig. 8a), while temperature alone explained 24.1 % of sRD variability (Fig. 8b).

Regression analysis of larval instantaneous growth rate Gi (d−1) versus standardized RNA:DNA ratio (sRD; N = 1177)

General additive model (GAM) analyses of larvae <14 mm (N = 820) of a standardized RNA:DNA ratio (sRD) versus Eurytemora abundance (explained variability: 40.9 %) and b sRD versus temperature (explained variability: 24.1 %). GAMs could only be performed for larvae <14 mm, as an autocorrelation between prey and temperature existed for the period when large larvae occurred in the Kiel Canal
Abiotic conditions (salinity and temperature) showed considerable differences between the years, especially in terms of salinity, which ranged between 5.5 and 11.5 (Fig. 9b). While salinity was extraordinary high in 2011, it was lowest in 2007. In contrast to the relatively high salinity variability, temperature development was linearly throughout the years, ranging from to 7.5 to 19.4 °C (Fig. 9a). In 2010, temperatures were lower on average compared to the other seasons. Neither Eurytemora abundance, nor larval Gi correlated significantly with temperature or salinity.

Seasonal course of a temperature, and b salinity during the years of investigation in the Kiel Canal. Different symbols indicate different years
Discussion
Based on contradicting results of studies dealing with prey availability and larval fish growth, it was recently hypothesized that fluctuating prey availability affects survival rates not primarily via larval growth rates, but rather indirectly by changes in larval behavior and therefore increasing predation rates [43]. Nevertheless, evidence shows that stage duration and therefore growth rates are of importance for recruitment ([39], and references therein). We demonstrate that larval growth rates can be recurrently food-limited in coastal habitats, which is, to the best of our knowledge, reported for the first time. Variability of mean larval herring growth rates was best explained when the analysis was restricted to the dominant copepod, Eurytemora affinis.
In the present study, food-limited growth was defined as lower growth compared to growth rates of herring larvae observed in a different habitat (Greifswalder Bodden) at the same temperature, which indicates a suboptimal growth performance. Similar approaches have been used by other authors before. For example, growth rates of central Baltic Sea cod larvae were correlated with temperature and compared to growth rates from Georges Bank and mesocosm experiments [40]. Similarly, body weight of field-caught radiated shannies Ulvaria subbifurcata were correlated with prey biomass and compared to larval weight from experimental work [7]. Additionally to the comparison of larval growth data from the Kiel Canal with those from the Greifswalder Bodden, the asymptotic relationship between larval growth and Eurytemora abundance serves as an indicator for food-limitation in itself, as larvae grew at lower rates at low prey abundances. Indirect temperature effects (increasing copepod abundance with increasing temperature) can be excluded, as Eurytemora abundance and temperature were not correlated.
Food-limited growth has rarely been observed in off-shore regions [9, 30, 40, 65]. As recurrently occurring food-limitation was observed in a coastal habitat in this study, the question arises, which basic differences between coastal and off-shore habitats do exist that potentially drive food-limitation? As food-limitation was observed each year, a recurrently occurring mechanism limiting copepod abundance is the most likely explanation. As most coastal habitats such as lagoons, estuaries and canals are characterized by a relatively small water body, they sensitively react to climate and weather events such as rainy seasons and strong short-term temperature changes which typically occur in boreal regions during spring time. In the study at hand, temperatures above 15 °C negatively affected copepod abundance. Even though salinity might not be drastically changed by rain or river-run off, relatively small changes could produce effects on prey availability. For example, small long term-changes in salinity correlated with decreasing copepod biomass caused by an altered species composition (decreasing abundance of large species) in the northern Baltic Sea [66], leading to food-limitation for adult herring [26]. However, Eurytemora abundance was not negatively but rather positively affected by this development [66]. In the Kiel Canal, no significant correlation between salinity and the abundance of the brackish water copepod Eurytemora was observed at salinities between 5.5 and 11.5. Nevertheless, highest Eurytemora abundances were observed when salinity was highest in 2011. Experimental work showed that egg production does not significantly differ at salinities between 5 and 15 [20].
Though larval herring appear to be selective feeders, they seem to be rather plastic concerning the choice of their preferred prey item, with a preference for the dominant species (for references, see below). This might explain the significant correlation between larval growth rates and Eurytemora, but not other copepod species in the present study. It was previously shown that larval herring feed selectively on Pseudocalanus and Oithona [14], while Acartia was negatively selected. Contrary, Busch [12] and Hesse [32] found herring larvae selectively feeding on Acartia spp. Results by Buckley and Durbin [10] showed highest correlation coefficients between prey abundance and growth rates of larval cod (Gadus morhua) and haddock (Melanogrammus aeglefinus), when analyses were restricted to their preferred prey, Pseudocalanus spp.. Similarly, abundances of the copepod Sinocalanus sinensis were observed to significantly affect growth rates of larval Japanese temperate bass (Lateolabrax japonicus; [41]), while this was not observed for other co-occurring copepod species. Peterson and Ausubel [56] found larval Atlantic mackerel (Scomber scombrus) selectively feeding on certain copepod species as well.
As abundances of adult herring remained quite constant in the northern Baltic Sea after changing copepod species composition caused by changing salinities [26], the early life stages of herring were apparently not strongly affected by these changing conditions. However, the situation might be different for other fish species or in different habitats. It was shown that the North Atlantic oscillation has the potential to cause regime shifts, including changing dominance of copepod species and leading to declining or low level cod (Gadus morhua) stocks [1, 5] in the North and Baltic Seas. Further, droughts were observed to cause hypersalinity in estuaries in the tropics and subtropics with severe ecological consequences [60], or increasing salinities were observed in coastal lakes caused by sea-level rise [61]. Effects of changing abiotic conditions on prey composition and therefore larval fish nutrition and survival and ultimately recruitment are expected to be case-dependent. Here, particularly the suitability of new dominant copepod species as prey organisms is important. Factors determining this suitability are primarily temporal occurrence and quality aspects (e.g., size, energy content and biochemical composition [especially essential fatty acid concentrations and C:N)].
Using a theoretical framework with larval fish behavior included [43], it was shown that low prey abundances lead to a more active larval behavior, and therefore better detection by predators and consequently increased predation rates. In that model, behavioral adjustment concerning food availability was included by vertical migration [25]. Due to the shallowness of the canal, the possibility for vertical migration is restricted. Consequently, the opportunity of behavioral adjustment is reduced for fish larvae in shallow nursery areas such as the Kiel Canal, but also lagoons or estuaries serving as nurseries (e.g., [4, 24, 27, 57]), and such areas might be exceptions of the validity of the model.
Generally, larval growth is used as an indirect indicator for mortality rates [39] assuming that a better nutritional situation for the larvae is reflected in better larval growth and lower larval mortality rates, and vice versa [3, 37]. Indeed, results of otolith analyses of the survivors in the Kiel Canal showed that the vast majority of the survivors hatched early in the season 2011, when growth conditions were good [33]. This supports the findings of the present study, were non-food-limited situations in the beginning of the season were observed, defined by increasing larval growth rates. Only very few survivors originated from the mid-season or shortly before food-limitation occurred. Overall, the survivors were larger during the season compared to the mean size of the larvae [33], supporting the Stage-Duration hypothesis and “bigger is better” [3, 37].
Potentially, food-limitation can have strong negative effects on recruitment (e.g., [39, 43]). Consequently, the question arises in how far recruitment might be affected by the food-limitation observed each season in the present study? In the Kiel Canal, food-limitation occurred only temporally during the season, and not consistently throughout the whole season. During the non-food limited period, which includes the particularly sensitive phase of first feeding of the peak-hatch cohort, copepod availability was high. Therefore, food-limitation most likely primarily affects survival of late hatched larvae. This means, that the time point of food limitation is of crucial importance for larval survival. Similarly, one might expect negative selection pressure on late hatching cohorts, when food limitation occurs recurrently at a certain time point of the season. However, herring is a very plastic species and distinct homing, i.e., that most or all herring that hatched in the Kiel Canal return to this sampling site to spawn, is unlikely. In other herring nursery areas, such as the Kiel Fjord and the Greifswalder Bodden, decreasing larval growth rates were not observed at the end of May/beginning of June (unpublished data), which is the period were food-limited growth occurred in the Kiel Canal. These results indicate that there is no uniform prey-induced selection pressure on herring larvae in the investigation area during the season.
Though significant differences in growth rates between larvae smaller and larger than 14 mm existed in the majority of the cases tested, observed trends did not differ between size classes. Results from experimental work indicated that large herring larvae grow faster than small ones (positive linear relationship; [17]). In our study, in one-third of the cases no significant differences between small and larger larvae were found, mostly during the food-limited periods. A possible reason might be that larger larvae have a higher energy demand than smaller ones, which is difficult to meet during food-limitation. This is especially true for the late season, where temperatures were relatively high. Instantaneous larval growth was found to increase approximately 0.01/1 °C increase in temperature [38], and increasing temperatures enhance larval metabolism and therefore energy demand [38], which is critical under food limitation. Hence, it is assumed that increasing temperatures affected larval growth negatively in the late season, i.e., under food limitation.
Eurytemora affinis was the numerically dominant copepod species found in Kiel Canal, contributing 79 % to total copepod abundance across all years investigated. Eurytemora is known to be dominant in brackish waters ([34, 62], and references therein). At a temperature of about 15 °C, the abundance of E. affinis decreased rapidly to negligible values throughout the years. This is consistent with findings by Hirche [34], who reported that the productivity of E. affinis females is directly related to their size, and the size decreases remarkably at temperatures ~15 °C. Eurytemora is reported being outcompeted by A. tonsa at temperatures above 15 °C ([34], and references therein]; the latter species needs relatively high temperatures for the eggs to hatch (≥13 °C; [36]).
High variability of individual larval growth rates was observed at given Eurytemora abundances. Factors causing this variability are especially temperature (increasing metabolism with increasing temperature, see above) and size-dependent growth. The latter phenomenon was shown being dependent on prey concentrations (Paulsen et al. accepted): the higher the prey concentrations, the smaller the differences between growth rates of small and large larvae were in the Kiel Canal during the years of investigation. Additionally, genetic and behavioral components (individual growth capacity and hunting capabilities) might play a role (as seen in experiments, when single larval herring specimens grow at rates far above average compared to same-age conspecifics).
Larval Gi variability was largely explained by standardized RNA:DNA ratios (sRD; [13]). This indicates that larval growth rates were predominantly determined by the nutritional condition of the larvae rather than temperature. This was also supported by results of GAM analyses, which showed major effects of Eurytemora abundance on sRD of larvae <14 mm. Between sRD and Eurytemora abundance an asymptotic relationship was observed. This is in line with findings from other studies, where non-linear to asymptotic relationships between larval growth rates and prey concentrations or prey ingestion rates were observed ([55], and references therein). This means that especially at the lower end increasing prey abundances are strongly beneficial for larval nutrition. It was shown that increasing temperatures negatively affect sRD levels [28], as less RNA is needed to achieve a certain growth rate at higher temperatures. This is explained by the fact that the ribosoms are more active at higher temperatures, which reduces the amount of RNA needed for a certain growth rate. In the present study, however, sRD moderately increased up to 15 °C and then steeply decreased with further increasing temperature levels. This pattern coincides with the seasonal development of Eurytemora abundances and points to the distinct effect of Eurytemora on the nutritional condition of the herring larvae.
A WP2-net with 200 µm mesh-size was used to sample the prey field. Though the smallest copepodite stages might be sampled only semi-quantitatively and nauplii are missed for most parts by using 200 µm mesh size, we are confident to have sampled a representative prey field as copepodite abundance follow nauplii abundances trends [58], and a significant correlation between copepod and nauplii abundance using 100 µm mesh size was observed in the Kiel Canal in 2014 [6].
Why were the slow growing larvae not removed by predators in the Kiel Canal, as observed in Japanese anchovies [63]? Unfortunately, information on predators in the habitat investigated is scarce. Potential predators known to occur in the Kiel Canal are pikeperch (Stizostedion lucioperca L.; [31]), river perch (Perca fluviatilis L.; [47]) and moon jellyfish (Aurelia aurita L.; [51]). While three-spined sticklebacks (Gasterosteus aculeatus L.) heavily prey upon herring eggs [45], they only rarely feed on larval herring [46]. Pikeperch and river perch are occasionally caught by local anglers close to the sampling site. However, the angler's catches start relatively late in the year during summer, which might hint at a temporal mismatch. Similarly, moon jellyfish were observed on the sampling site, but not before mid-June to July, also leading to a temporal mismatch situation between larval herring and predators in Kiel Canal.
We conclude that our hypothesis proposing that in coastal habitats food-limited growth of larval fish can occur recurrently was supported. Larval growth observed in Kiel Fjord was substantially lower compared to the Greifswalder Bodden at temperatures ≥15 °C. Furthermore, growth rates decreased simultaneously with Eurytemora abundance in the late season, and asymptotic relationships between Eurytemora and larval growth were observed. Larval growth was best explained by the abundance of the numerically dominant prey item Eurytemora affinis, highlighting the crucial role of this copepod concerning larval herring nutrition in the Kiel Canal. As food limitation occurs recurrently at the end of the season in the Kiel Canal, the optimal time point for larvae to hatch is early in the season so that larvae can take full advantage of the high copepod production in this period.
Abbreviations
- ANOVA:
-
one way analysis of variance
- CI:
-
confidence interval
- DNA:
-
deoxyribonucleic acid
- GAM:
-
generalized additive model
- Gi :
-
larval instantaneous growth rate
- HSD:
-
honestly significant difference
- mm:
-
millimeter
- RNA:
-
ribonucleic acid
- sRD:
-
standardized RNA:DNA ratio
- T:
-
temperature
References
Alheit J, Möllmann C, Dutz J, Kornilovs G, Loewe P, Mohrholz V, et al. Synchronous ecological regime shifts in the central Baltic and the North Sea in the late 1980s. ICES J Mar Sci. 2005;62:1205–15.
Ambler JW. Seasonal factors affecting egg production and viability of eggs of Acartia tonsa Dana from East Lagoon, Galveston, Texas. Estuar Coast Shelf Sci. 1985;20:743–60.
Anderson JT. A review of size dependent survival during pre-recruit stages of fishes in relation to recruitment. J Northwest Atl Fish Sci. 1988;8:55–66.
Barletta-Bergan A, Barletta M, Saint-Paul U. Structure and seasonal dynamics of larval fish in the Caeté River estuary in North Brazil. Estuar Coast Shelf Sci. 2002;54:193–206.
Beaugrand G, Brander KM, Alistair Lindley J, Souissi S, Reid PC. Plankton effect on cod recruitment in the North Sea. Nature. 2003;426:661–4.
Beckmann R. Effects of microzooplankton on the planktonic food web. Diploma thesis, Kiel University; 2015.
Bochdansky AB, Grønkjær P, Pepin P, Leggett WC. Food limitation in larval fish: ontogenetic variation in feeding scope and its potential effect on survival. Mar Ecol Prog Ser. 2008;367:239–48.
Brucet S, Boix D, Gascón S, Sala J, Quintana XD, Badosa A, et al. Species richness of crustacean zooplankton and trophic structure of brackish lagoons in contrasting climate zones: north temperate Denmark and Mediterranean Catalonia (Spain). Ecography. 2009;32:692–702.
Buckley LJ, Caldarone EM, Lough RG. Optimum temperature and food-limited growth of larval Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) on Georges Bank. Fish Oceanogr. 2004;13:134–40.
Buckley LJ, Durbin EG. Seasonal and inter-annual trends in the zooplankton prey and growth rate of Atlantic cod (Gadus morhua) and haddock (Melanogrammus aeglefinus) larvae on Georges Bank. Deep-Sea Res Part II. 2006;53:2758–70.
Buckley LJ, Caldarone EM, Clemmesen C. Multi-species larval fish growth model based on temperature and fluorometrically derived RNA/DNA ratios: results from a meta-analysis. Mar Ecol Prog Ser. 2008;371:221–32.
Busch A. Nahrungsökologische Untersuchungen an den Larven des Rügenschen Frühjahrsherings (Clupea harengus L.) im Greifswalder Bodden in den Jahren 1990 bis 1992. Dissertation, Rostock University; 1993.
Caldarone EM, Clemmesen CM, Berdalet E, Miller TJ, Folkvord A, Holt GJ, et al. Intercalibration of four spectrofluorometric protocols for measuring RNA/DNA ratios in larval and juvenile fish. Limnol Oceanogr Methods. 2006;4:153–63.
Checkley DM. Selective feeding by Atlantic herring (Clupea harengus) larvae on zooplankton in natural assemblages. Mar Ecol Prog Ser. 1982;9:245–53.
Chicharo MA, Chicharo L, Valdes L, Lopez-Jamar E, Re P. Does the nutritional condition limit survival potential of sardine Sardina pilchardus (Walbaum, 1792) larvae off the north coast of Spain? RNA/DNA ratios and their variability. Fish Res. 1998;39:43–54.
Clemmesen C. A RNA and DNA fluorescence technique to evaluate the nutritional condition of individual marine fish larvae. Meeresforschung. 1988;32:134–43.
Clemmesen C. The effect of food availability, age or size on the RNA/DNA ratio of individually measured herring larvare—laboratory calibration. Mar Biol. 1994;118:377–82.
Clemmesen C, Sanchez R, Wongtschowski C. A regional comparison of the nutritional condition of SW Atlantic anchovy larvae, Engraulis anchoita, based on RNA/DNA ratios. Arch Fish Mar Res. 1997;45:17–43.
Cury P, Roy C. Optimal environmental window and pelagic fish recruitment success in upwelling areas. Can J Fish Aquat Sci. 1989;46:670–80.
Devreker D, Souissi S, Winkler G, Forget-Leray J, Leboulenger F. Effects of salinity, temperature and individual variability on the reproduction of Eurytemora affinis (Copepoda; Calanoida) from the Seine estuary: a laboratory study. J Exp Mar Biol Ecol. 2009;368:113–23.
Diaz MV, Pajaro M, Olivar MP, Martos P, Macchi GJ. Nutritional condition of Argentine anchovy Engraulis anchoita larvae in connection with nursery ground properties. Fish Res. 2011;109:330–41.
Donner M. Wachstum und Kondition von Heringslarven (Clupea harengus L.) in der Kieler Förde und im Nord-Ostsee-Kanal. Diploma thesis, Kiel University; 2006.
Doyle M. A morphological staging system for the larval development of the herring, Clupea harengus L. J Mar Biol Assoc UK. 1977;57:859–67.
Fey DP. Differences in temperature conditions and somatic growth rate of larval and early juvenile spring-spawned herring from the Vistula Lagoon, Baltic Sea manifested in the otolith to fish size relationship. J Fish Biol. 2001;58:1257–73.
Fiksen Ø, Jørgensen C. Model of optimal behaviour in fish larvae predicts that food availability determines survival, but not growth. Mar Ecol Prog Ser. 2011;432:207–19.
Flinkman J, Aro E, Vuorinen I, Viitasalo M. Changes in northern Baltic zooplankton and herring nutrition from 1980s to 1990s: top-down and bottom-up processes at work. Mar Ecol Prog Ser. 1998;165:127–36.
Franco A, Franzoi P, Malavasi S, Riccato F, Torricelli P, Mainardi D. Use of shallow water habitats by fish assemblages in a Mediterranean coastal lagoon. Estuar Coast Shelf Sci. 2006;66:67–83.
Goolish EM, Barron MG, Adelman IR. Thermoacclimatory response of nucleic acid and protein content of carp muscle tissue: influence of growth rate and relationship to glycine uptake by scales. Can J Zool. 1984;62:2164–70.
Greenwald GM, Hurlbert SH. Microcosm analysis of salinity effects on coastal lagoon plankton assemblages. Hydrobiologia. 1993;267:307–35.
Grønkjær P, Clemmesen C, St John M. Nutritional condition and vertical distribution of Baltic cod larvae. J Fish Biol. 1997;51:352–69.
Hansson S, Arrhenius F, Nellbring S. Diet and growth of pikeperch (Stizostedion lucioperca L.) in a Baltic Sea area. Fish Res. 1997;31:163–7.
Hesse J. Quantitative and qualitative analyses of the food consumption of Clupea harengus L. larvae in the Greifswalder Bodden and Strelasund, Baltic Sea, Germany. Diploma thesis, Rostock University; 2010.
Hesse C. Analyses of growth and survival of herring larvae in the Kiel Canal. Diploma thesis, Kiel University; 2012.
Hirche H-J. Egg production of Eurytemora affinis—effect of k-strategy. Estuar Coast Shelf Sci. 1992;35:395–407.
Hjort J. Fluctuations in the great fisheries of northern Europe viewed in the light of biological research. Rapp Proces. 1914;20:1–228.
Holste L, Peck MA. The effects of temperature and salinity on egg production and hatching success of Baltic Acartia tonsa (Copepoda: Calanoida): a laboratory investigation. Mar Biol. 2005;148:1061–70.
Houde ED. Fish early life dynamics and recruitment variability. Am Fish Soc Symp. 1987;2:17–29.
Houde ED. Comparative growth, mortality, and energetics of marine fish larvae: temperature and implied latitudinal effects. Fish Bull. 1989;87:471–95.
Houde ED. Emerging from Hjort’s Shadow. J Northwest Atl Fish Sci. 2008;41:53–70.
Huwer B, Clemmesen C, Grønkjær P, Köster FW. Vertical distribution and growth performance of Baltic cod larvae—field evidence for starvation-induced recruitment regulation during the larval stage? Prog Oceanogr. 2011;91:382–96.
Islam MS, Hibino M, Nakayama K, Tanaka M. Condition of larval and early juvenile Japanese temperate bass Lateolabrax japonicus related to spatial distribution and feeding in the Chikugo estuarine nursery ground in the Ariake Bay, Japan. J Sea Res. 2006;55:141–55.
Jeppesen E, Søndergaard M, Pedersen AR, Jürgens K, Strzelczak A, Lauridsen TL, et al. Salinity induced regime shift in shallow brackish lagoons. Ecosystems. 2007;10:48–58.
Jørgensen C, Opdal AF, Fiksen O. Can behavioural ecology unite hypotheses for fish recruitment? ICES J Mar Sci. 2014;71:909–17.
Kott P. Modified whirling apparatus for the subsampling of plankton. Aust J Mar Freshw Res. 1953;4:387–93.
Kotterba P, Kuehn C, Hammer C, Polte P. Predation of threespine stickleback (Gasterosteus aculeatus) on the eggs of Atlantic herring (Clupea harengus) in a Baltic Sea lagoon. Limnol Oceanogr. 2014;59:578–87.
Kotterba P. Atlantic herring (Clupea harengus) within the food web of a southern Baltic Sea lagoon. Dissertation, University of Hamburg; 2015.
Lappalainen A, Rask M, Koponen H, Vesala S. Relative abundance, diet and growth of perch (Perca fluviatilis) and roach (Rutilus rutilus) at Tvaerminne, northern Baltic Sea, in 1975 and 1997: responses to eutrophication? Boreal Environ Res. 2001;6:107–18.
Malzahn AM, Aberle N, Clemmesen C, Boersma M. Nutrient limitation of primary producers affects planktivorous fish condition. Limnol Oceanogr. 2007;52:2062–71.
Malzahn AM, Clemmesen C, Wiltshire KH, Laakmann S, Boersma M. Comparative nutritional condition of larval dab Limanda limanda and lesser sandeel Ammodytes marinus in a highly variable environment. Mar Ecol Prog Ser. 2007;334:205–12.
Meekan MG, Fortier L. Selection for fast growth during the larval life of Atlantic cod Gadus morhua on the Scotian Shelf. Mar Ecol Prog Ser. 1996;137:25–37.
Möller H. Reduction of a larval herring population by jellyfish predator. Science. 1984;224:621–2.
Newton A, Mudge SM. Temperature and salinity regimes in a shallow, mesotidal lagoon, the Ria Formosa, Portugal. Estuar Coast Shelf Sci. 2003;57:73–85.
Paulsen M, Clemmesen C, Malzahn AM. Essential fatty acid (docosahexaenoic acid, DHA) availability affects growth of larval herring in the field. Mar Biol. 2014;161:239–44.
Paulsen M, Hammer C, Malzahn AM, Polte P, von Dorrien C, Clemmesen C. Nutritional situation for larval Atlantic herring (Clupea harengus L.) in two nursery areas in the western Baltic Sea. ICES J Mar Sci. 2014;71:991–1000.
Peck MA, Reglero P, Takahashi M, Catalán IA. Life cycle ecophysiology of small pelagic fish and climate-driven changes in populations. Prog Oceanogr. 2013;116:220–45.
Peterson WT, Ausubel SJ. Diets and selective feeding by larvae of Atlantic mackerel Scomber scombrus on zooplankton. Mar Ecol Prog Ser. 1984;17:65–75.
Polte P, Kotterba P, Hammer C, Gröhsler T. Survival bottlenecks in the early ontogenesis of Atlantic herring (Clupea harengus, L.) in coastal lagoon spawning areas of the western Baltic Sea. ICES J Mar Sci. 2014;71:982–90.
Postel A, Postel L, Hantke H. Untersuchungen zur räumlich-zeitlichen Verteilung der Heringslarvennahrung von April bis Juni 1988 im südlichen Greifswalder Bodden. Fisch Forsch Rostock. 1989;29:43–55.
Sars GO. An account of the Crustacea of Norway: with short descriptions and figures of all the species, vol 1. Copenhagen: A. Cammermeyer; 1895.
Savenije HHG, Pagès J. Hypersalinity: a dramatic change in the hydrology of Sahelian estuaries. J Hydrol. 1992;135:157–74.
Schallenberg M, Hall CJ, Burns CW. Consequences of climate-induced salinity increases on zooplankton abundance and diversity in coastal lakes. Mar Ecol Prog Ser. 2003;251:181–9.
Schnack D. Nahrungsökologische Untersuchungen an Heringslarven. Ber Dt Wiss Komm Meeresforsch. 1972;22:273–343.
Takasuka A, Aoki I, Mitani I. Evidence of growth-selective predation on larval Japanese anchovy Engraulis japonicus in Sagami Bay. Mar Ecol Prog Ser. 2003;252:223–38.
Telesh IV, Khlebovich VV. Principal processes within the estuarine salinity gradient: a review. Mar Pollut Bull. 2010;61:149–55.
Voss R, Clemmesen C, Baumann H, Hinrichsen HH. Baltic sprat larvae: coupling food availability, larval condition and survival. Mar Ecol Prog Ser. 2006;308:243–54.
Vuorinen I, Hänninen J, Viitasalo M, Helminen U, Kuosa H. Proportion of copepod biomass declines with decreasing salinity in the Baltic Sea. ICES J Mar Sci. 1998;55:767–74.
Yandi I, Altinok I. Defining the starvation potential and the influence on RNA/DNA ratios in horse mackerel (Trachurus mediterraneus) larvae. Helgol Mar Res. 2015;69:25–35.
Authors’ contributions
MP carried out larval herring RNA:DNA ratio analyses from the seasons 2009–2012 (including Greifswalder Bodden samples), sorted the mesozooplankton samples from the seasons 2007, 2008, 2009, 2011 and 2012 and drafted the manuscript. CC, CH, PP and AMM participated in data analyses, interpretation of the data and ms writing/correction. Furthermore, PP sampled larval herring in Greifswalder Bodden. All authors read and approved the final manuscript.
Acknowledgements
We thank both anonymous reviewers for their constructive comments. Many thanks go to Helgi Mempel and Jesco Peschutter, who were involved in the beginning of the time series, kept it running and analyzed RNA:DNA of larval herring from the seasons 2007 and 2008. We like to thank the research vessel crew of FB “Polarfuchs” for their help during sampling. Thanks to Paul Kotterba for creating Fig. 1, and to Martina Stiasny for running the GAMs. The research was partly financed and conducted as part of the Fehmarn Belt Science Provision Project. Part of the study was funded by the German Science Foundation (DFG) in the AQUASHIFT priority program 520 1164 (“RECONN”) with a grant awarded to Catriona Clemmesen.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Data will be made publicly and freely accessible on the database PANGAEA right after publication of this manuscript.
Consent for publication
The manuscript includes no personal data. All authors have read the manuscript and have approved the revised submission.
Ethics approval and consent to participate
Permission to sample and analyze the herring larvae in the Kiel Canal has been granted by the Landessportfischerverband Schleswig–Holstein e.V. and the Animal Welfare Ethics Commission at Kiel University (§4 Notification “Time series—Plankton investigations in the Kiel Fjord and the Kiel Canal” is approved with the internal number 896).
Funding
Part of the study was funded by the German Science Foundation (DFG) in the AQUASHIFT priority program 520 1164 (“RECONN”) with a grant awarded to Catriona Clemmesen. The research was partly financed and conducted as part of the Fehmarn Belt Science Provision Project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Paulsen, M., Clemmesen, C., Hammer, C. et al. Food-limited growth of larval Atlantic herring Clupea harengus recurrently observed in a coastal nursery area. Helgol Mar Res 70, 17 (2017). https://doi.org/10.1186/s10152-016-0470-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s10152-016-0470-y