
Abstract
Introduction
The prolactin-Janus-kinase-2-signal transducer and activator of transcription-5 (JAK2-STAT5) pathway is essential for the development and functional differentiation of the mammary gland. The pathway also has important roles in mammary tumourigenesis. Prolactin regulated target genes are not yet well defined in tumour cells, and we undertook, to the best of our knowledge, the first large genetic screen of breast cancer cells treated with or without exogenous prolactin. We hypothesise that the identification of these genes should yield insights into the mechanisms by which prolactin participates in cancer formation or progression, and possibly how it regulates normal mammary gland development.
Methods
We used subtractive hybridisation to identify a number of prolactin-regulated genes in the human mammary carcinoma cell line SKBR3. Northern blotting analysis and luciferase assays identified the gene encoding heat shock protein 90-alpha (HSP90A) as a prolactin-JAK2-STAT5 target gene, whose function was characterised using apoptosis assays.
Results
We identified a number of new prolactin-regulated genes in breast cancer cells. Focusing on HSP90A, we determined that prolactin increased HSP90A mRNA in cancerous human breast SKBR3 cells and that STAT5B preferentially activated the HSP90A promoter in reporter gene assays. Both prolactin and its downstream protein effector, HSP90α, promote survival, as shown by apoptosis assays and by the addition of the HSP90 inhibitor, 17-allylamino-17-demethoxygeldanamycin (17-AAG), in both untransformed HC11 mammary epithelial cells and SKBR3 breast cancer cells. The constitutive expression of HSP90A, however, sensitised differentiated HC11 cells to starvation-induced wild-type p53-independent apoptosis. Interestingly, in SKBR3 breast cancer cells, HSP90α promoted survival in the presence of serum but appeared to have little effect during starvation.
Conclusions
In addition to identifying new prolactin-regulated genes in breast cancer cells, we found that prolactin-JAK2-STAT5 induces expression of the HSP90A gene, which encodes the master chaperone of cancer. This identifies one mechanism by which prolactin contributes to breast cancer. Increased expression of HSP90A in breast cancer is correlated with increased cell survival and poor prognosis and HSP90α inhibitors are being tested in clinical trials as a breast cancer treatment. Our results also indicate that HSP90α promotes survival depending on the cellular conditions and state of cellular transformation.
Similar content being viewed by others

Introduction
The proliferation and functional differentiation of mammary epithelial cells is highly dependent on the action of prolactin [1]. These effects of prolactin are mainly mediated through the prolactin receptor Janus kinase-2-signal transducers and activators of transcription-5 (JAK2-STAT5) pathway, and results in mammary epithelial cell proliferation and the differentiation of alveolar cells during pregnancy. On weaning, a large proportion of these alveolar cells die off in a massive wave of apoptosis and tissue remodelling [2]. Generally speaking, prolactin and STAT5 are thought to induce genes for survival in the differentiated cells, while STAT3 is thought to induce genes required for apoptosis [3].
Prolactin signalling has also been implicated in mammary and breast cancer, including invasive [4] and non-invasive breast cancer [5–7]. The transgenic expression of prolactin results in increased tumour formation in mice [8–10]. Crossing prolactin-deficient mice with oncogenic polyoma middle-T antigen transgenic mice demonstrated that prolactin decreased the latency of tumour formation and increased tumour growth [11]. Using a cross with SV-40T oncogene transgenics and prolactin receptor null mice, and transplant of the epithelium to endocrine normal mice, the prolactin receptor was demonstrated to increase neoplasia and positively impact the transition to invasive carcinoma [12].
In humans, high serum prolactin levels increase the risk of breast cancer for women [4, 7, 13]. Although expression of the prolactin receptor is more often found in oestrogen receptor-positive breast tumours, which tend to have a better prognosis, it is also found in many oestrogen receptor-negative breast tumours [14]. The gene encoding the prolactin receptor is also highly expressed in a subset of breast tumours with poor prognosis and is part of a set of prognostic gene markers [15]. Prolactin is not only secreted by the pituitary gland, but is also produced locally in the majority of breast tumours and is thought to act in an autocrine and/or paracrine fashion [6]. Although prolactin can transduce signals through multiple pathways, the activation of STAT family members, which are downstream of prolactin and other growth factors has also been implicated in tumourigenesis [16, 17]. Prolactin-regulated target genes are not yet well defined in breast cancer cells. Only recently have large-scale attempts at identifying prolactin target genes been performed using the mammary gland and these have been limited to normal mammary epithelial cells [18–24].
The heat shock protein 90-alpha (HSP90α) protein is referred to as the cancer chaperone [25, 26], a molecular chaperone of proteins involved in essential signal transduction pathways regulating proliferation, differentiation, apoptosis, angiogenesis, metastasis, oncogenesis [27–29], genetic variation [30, 31], invasion [32] and cellular transformation [33]. It is distinct from HSP90β [34], a protein encoded by a related gene, which is constitutively expressed. HSP90α gene expression is elevated in breast cancers [35–37] and is correlated with decreased patient survival [35–37]. HSP90 inhibitors bind specifically and preferentially to HSP90α and β in cancerous cells versus normal cells [38]. Inhibition of HSP90 results in the proteosomal degradation of many HSP90 client proteins. Client proteins include erythroblastic leukaemia viral oncogene homolog 2 (ERBB2) and AKT/protein kinase B [39] and their loss results in apoptosis. HSP90 inhibitors, such as geldanamycin derivatives like 17-allylamino-17-demethoxygeldanamycin (17-AAG), have entered clinical trials for the treatment of breast and prostate cancer and melanoma [27, 40, 41].
Prolactin-regulated target genes in breast cancer cells are likely to also function in normal cells, but under appropriate regulation. We hypothesised that prolactin-regulated genes in breast cancer cells would also have important functions in non-transformed mammary epithelial cells and may potentially contribute to cancer progression when deregulated. Using a subtractive hybridisation approach, we identified a number of cancer-related genes whose expression is modified by the addition of prolactin in the human breast adenocarcinoma cell line, SKBR3. We specifically identified the therapeutically important gene, HSP90A as a STAT5 regulated target gene. HSP90A appears to regulate survival differentially depending on the cellular levels of its protein product HSP90α, the presence of survival factors and the status of cellular transformation.
Materials and methods
Antibodies
Polyclonal rabbit antibodies against HSP90α were acquired from StressGen (Victoria, BC, Canada), and rabbit anti-STAT5B from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Mouse anti-phospho-histone 2A.X (Ser139) clone JBW301 and anti-GRB2 antibodies (Upstate, Charlottesville, USA) and mouse anti-GRB2 (BD Biosciences, Ontario, Canada) were also acquired. GRB2 was used as a loading control [42].
Plasmids
The HSP90A-luciferase reporter gene contains about 1.8 kb of promoter sequences (Xho I-Hind III) of the human HSP90A gene cloned into pLux F3 (KS89 α XL Lux). Expression constructs for the β-galactosidase gene, STAT5A and STAT5B [43], STAT1 and STAT3 (pME18S) and prolactin-receptor [44] have been described. The DNA encoding human HSP90A (2199 bp, accession number [GenBank:X15183]) was amplified by PCR from cDNA prepared from SKBR3 cells (SMART cDNA synthesis kit, Clontech Laboratories, Mountain View, CA, USA). The resulting DNA was cut with Bam HI and Not I (New England Biolabs, Ipswich, MA, USA) (adaptors on the primers), subcloned into pcDNA 3.1/Zeo(+) behind the constitutively active cytomegalovirus immediate early promoter (CMV) and the expected sequence verified.
Cell culture and cell lines
SKBR3 cells, a human breast cancer cell line, were grown in Dulbecco's Modified Eagle Medium with L-glutamine and 10% fetal bovine serum. Undifferentiated HC11 cells, a mouse mammary epithelial cell line [45], were cultured in RPMI with fetal bovine serum and maintained in 0.01 μg/ml epidermal growth factor (EGF) and 5 μg/ml insulin. Confluent cells became competent to respond to lactogenic hormones after incubation in 0.01 μg/ml EGF for one to four days and then 5 μg/ml insulin and 1 × 10-7 M dexamethasone for one day (option if treating with EGF for only one day). Differentiation was then induced (for three days if insulin and dexamethasone pre-treatment was used, or four days if not) with 1 × 10-7 M dexamethasone, 5 μg/ml insulin and 5 μg/ml prolactin. Undifferentiated HC11 cells were transfected with HSP90A-pcDNA 3.1/Zeo(+) (HC11-HSP90α) or the empty vector pcDNA3.1/Zeo(+) (HC11-EV) using Lipofectamine 2000 (catalogue 11668-019, Invitrogen Corporation, Carlsbad, California, USA) according to the manufacturer's instructions. Transfected cells were selected as a pool with zeocin treatment and constitutive expression was verified by both northern and western blotting.
Subtractive hybridisation libraries
RNA was prepared (RNeasy, Clontech, Heidelberg, Germany) from SKBR3 cells seeded at 1 × 106 and 2 × 106 cells per 15 cm plate, starved of fetal bovine serum the following day for 16 hours and then treated with or without 5 μg/ml prolactin for 60 minutes in the presence of 10 nM cycloheximide. The subtraction hybridisation libraries were prepared using the SMART PCR cDNA Synthesis Kit and the PCR Select Subtractive Hybridization kit (Clontech, Heidelberg, Germany), and probed using the PCR Select Differential Screening kit (Clontech, Heidelberg, Germany). Positive clones were sequenced to obtain their identity.
Northern blotting
Total RNA was extracted using peqGOLD Trifast (peqLab, Erlangen, Germany) and resolved on a formaldehyde agarose gel. The blot was blocked using ExpressHyb (Clontech, Heidelberg, Germany), and hybridised with radioactively labelled probes (Strip-EZ DNA, Ambion, AMS Biotechnology Ltd). A DNA probe encoding exon 3 of the human CIS gene [46] was amplified using the forward primer 5'-GCT GGT ATT GGG GTT CC-3' and the reverse primer 5'-TGA GGG CTC TGT ACA TGA AAG-3' The fragments were gel purified (Qiaquick Gel Extraction Kit, Qiagen GmbH, Hilden, Germany).
Electrophoretic mobility shift assays
Electrophoretic mobility shift assays using a radioactively labelled fragment of the bovine β-casein promoter were performed as described [47]. Essentially the double strand STAT responsive element from the β-casein promoter was prepared and radioactively labelled before incubation with protein extracts. The complexes were resolved on a non-reducing gel and autoradiographed.
Transfection and luciferase assays
Transfection using calcium chloride and luciferase assays using HeLa cells was performed as described [48] using overnight treatments with 5 μg/ml prolactin. A β-galactosidase gene was included to compare transfection efficiencies in individual experiments.
Western blotting
Soluble protein extracts were prepared in a buffer containing 1% Nonidet-P-40, 50 mM Tris pH 7.5, 5 mM ethylene glycol tetraacetic acid and 200 mM sodium chloride, with freshly added protease and phosphatase inhibitors: 1 mM sodium vanadate, 20 μM phenylarsine oxide, 1 μg/ml leupeptin, 0.5 μg/ml aprotinin, 100 μM phenylmethylsulphonyl fluoride and 1 mM DTT. After protein concentrations were measured with the Bio-Rad Assay, 50 μg of each lysate was resolved by 15% SDS-PAGE and then transferred to Hybond-P PVDF transfer membrane (catalogue RPN303F, Amersham, GE Healthcare, Baie d'Urfé, Québec, Canada). The membrane was blocked in 5% non-fat milk in tris-buffered saline with 0.05% Tween 20 and incubated with 1 μg/ml of primary antibody mouse anti-phospho-histone H2A.X (Ser139), clone JBW301 (Upstate, Millipore, Billerica, MA, USA) followed with horseradish peroxidase (HRP)-conjugated goat anti-mouse secondary antibody. The signal was developed by solutions prepared with 250 mM luminal solution, 90 mM p-coumaric solution, 1 M Tris pH 8.5 and 30% hydrogen peroxide.
Apoptosis assay
HC11 cells transfected with either the empty vector HC11-EV or with the HSP90α expression construct (HC11-HSP90α) were plated at 130,000 cells/well of a 96-well plate and differentiated as above, then starved by the absence of serum and hormones. SKBR3 cells were plated at 10,000 cells/well. The presence of mono- and oligo-nucleosomes in the cytoplasm were qualitatively measured using the Cell Death Detection Elisa Plus Kit (Roche, Mississauga, ON, Canada). Essentially, protein extracts were incubated with anti-DNA (HRP-coupled) and anti-histone (biotin coupled) antibodies, before incubation in streptavidin-coated 96-well plates. Colorimetric detection was performed at the absorbance wavelength of 405 to 490 nm.
Results
SKBR3 human breast carcinoma cells are responsive to prolactin through STAT5-mediated gene transcription
To investigate the role of prolactin in breast cancer, we set out to identify prolactin responsive genes in the breast cancer cell line, SKBR3. We first examined the prolactin-based activation of STAT5 and the induction kinetics of previously identified STAT5-dependent genes. SKBR3 cells were treated with different doses of prolactin and the activated DNA-binding form of STAT5 was visualised in electrophoretic mobility shift assays (Figure 1a). The experiment shows that prolactin is able to activate STAT5 in SKBR3 cells in a dose-dependent manner. A STAT5 specific antibody was used to confirm the specificity of the protein-DNA complex (Figure 1a). Prolactin stimulation of 60 minutes resulted in activation and binding of STAT5 to DNA response elements present in the β-casein promoter. Of note is the lack of STAT5 activation in the absence of prolactin stimulation. SKBR3 cells have previously been shown to express the prolactin gene [49], but perhaps the endogenous levels of prolactin are not sufficient to induce activation of the JAK2-STAT5 pathway.

Prolactin-STAT5-mediated signal transduction in SKBR3 cells. (a) Electrophoretic mobility shift assays were carried out with a radioactively labelled β-casein gene promoter probe and protein extracts of SKBR3 cells treated for one hour with the indicated amounts of prolactin (PRL). The arrows indicate the positions of the signal transducers and activators of transcription (STAT5) DNA complex and of a supershifted complex formed in the presence of a STAT5 specific antibody (Ab). (b) SKBR3 cells were seeded one day, the next day starved of serum overnight and the following day were stimulated (or not) with 5 μg/ml prolactin for the times indicated in the absence of cycloheximide (CHX) unless indicated. In each lane 20 μg of RNA were applied. The northern blot was first hybridised with labelled DNA encoding cytokine inducible SH2 containing protein (CIS) and then rehybridised with a glyceraldehyde 3-phosphate dehydrogenase (GAPDH) specific probe.
We next followed the kinetics of cytokine-inducible-SH2-containing protein (CIS) mRNA induction as a function of prolactin treatment of SKBR3 cells. The CIS gene [50] is a known target of the JAK2-STAT5 pathway activated by interleukin-2 or erythropoietin in lymphoid cells and is important for feedback inhibition [51, 52]. The amount of CIS mRNA increased within 60 minutes of treatment and was further enhanced in the presence of cycloheximide, an inhibitor of protein synthesis (Figure 1b). The level of CIS mRNA reached a maximum at four hours and remained high for at least 18 hours. This confirmed that SKBR3 cells are appropriate to study the early induction of prolactin-JAK2-STAT5-regulated genes and that a time point of 60 minutes would result in the production of early target genes.
Prolactin-regulated genes in SKBR3 breast cancer cells
To identify additional genes regulated by prolactin in SKBR3 cells, we prepared subtraction hybridisation libraries. Based on the above observations, SKBR3 cells were treated for 60 minutes with 5 μg/ml prolactin and the RNA was used for preparation of the subtractive hybridisation libraries. Cycloheximide was added to cell preparations, both untreated and treated with prolactin in order to avoid identification of secondary targets. We prepared subtraction libraries for the genes differentially expressed on prolactin treatment (forward) and for genes expressed in the absence of prolactin (reverse). The two libraries were used in the differential screen for prolactin-regulated genes. About 1200 gene fragments were screened from the forward library and 770 from the reverse library. Genes were screened using Southern blotting in batches of 100 genes using the forward and reverse libraries as probes. The genes with the most intense differential signal on each blot, as measured using a Biorad phosphoimager (Bio-Rad, Munich, Germany), were selected for sequencing. Seventy-two positive clones selected on the basis of high expression in the forward (Table 1) or reverse libraries (Table 2) were sequenced and identified. Some genes were observed more than once (Tables 1 and 2), also indicating high differential expression. Of the 51 genes represented, 19 genes with the highest expression levels were rescreened in dot-blots using both the unsubtracted probes (cDNA) and subtracted probes and the expression patterns of 16 of the 19 genes were confirmed (86%) (Tables 1 and 2).
Although there were no pre-existing large studies of prolactin-regulated genes in breast cancer cells, we compared our results with other prolactin-related studies. Of note, 11 of the genes we identified overlapped with those previously identified as downstream targets of the prolactin pathway. In one study, prolactin gene targets were identified by gene array using regenerated mammary glands from prolactin-receptor-/- and cyclin D-/- mammary epithelial cell transplants. Cyclin D1 was thought to represent a secondary target of prolactin, and therefore genes identified in this screen would represent effectors downstream of prolactin and upstream of cyclin D1 [18]. Although the transplants were nontransformed cells, unlike SKBR3 cells, we noted some similarities. Genes that were identical in this study and our report include the genes encoding the mouse ferritin heavy chain gene, HSPs 70, 71 and 84 (HSP90B). We also found overlap with prolactin-regulated genes identified in the rat Nb2-11c lymphoma cell line [53], including HSP70 and HSP86 (HSP90A). Genes found to be similar (either functionally related or different subunits of a complex) between these two studies and ours include Sec 23 [18] and Sec 22 [53] (similar to Sec 24 in our study), elongation factor 2 [18, 53] (similar to elongation factor 1 alpha), lactate dehydrogenase 1 A chain [18] (similar to lactate dehydrogenase H chain), myosin heavy chain (similar to myosin regulatory light chain), T-complex protein 1 e and h subunits [53] (similar to T-complex b subunit). The degree of overlap with the prolactin-regulated genes of these two studies is comparable with the level previously described between other prolactin target gene studies [19]. The diversity of the gene lists found in each study is thought to derive from the differences between experimental conditions, cell types and methods.
HSP90Ais a prolactin-induced gene in SKBR3 human breast cancer cells
The gene HSP90A was identified five times in the initial group selected for sequencing; and based on its high representation, differential expression and its function in cancer cells, was used for further analysis.
We used northern blotting analysis to independently verify the induction of the HSP90A gene in SKBR3 cells by prolactin. We plated cells at two different confluences (about 40% and 60% confluent on the day of stimulation) as was performed for the preparation of the library. Both populations responded to 60 minutes of prolactin treatment with the increase in HSP90A mRNA, but the dose response of HSP90A induction in the two-cell population was distinguishable. Cells at the higher confluence exhibited a maximal response at 1.5 μg/ml, whereas cell at lower confluence required 5 μg/ml for maximal induction (Figure 2a). Higher concentrations of prolactin reduce the maximal response. The observed lower level of total RNA in high confluence cells treated with 10 μg/ml prolactin, and potentially the inhibitory presence of CIS, may explain the reduction in signal observed in higher confluence cells at 10 μg/ml. Another alternative is that the high concentration of prolactin induced a refractory state of prolactin signal transduction [54]. These results confirm that the HSP90A gene is a prolactin-regulated gene in the human mammary carcinoma cell line, SKBR3.

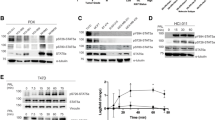
HSP90α is a prolactin regulated target gene. (a) Dose response of the heat shock protein 90 alpha (HSP90A) gene to prolactin in SKBR3 cells assessed by northern analysis. SKBR3 cells were seeded to achieve low and higher confluence before treatment for one hour with the indicated doses of prolactin (PRL). Total RNA was transferred to a nylon membrane, and the blot probed with the HSP90A gene fragment. (b) Western blot analysis: HSP90α protein is increased about two-fold in SKBR3 cells in response to 5 ug/mL prolactin. (c) Northern analysis: HSP90α mRNA is increased about four-fold in HC11 cells in response to a one hour treatment of lactogenic hormones, prolactin, dexamethasone and insulin (DIP) compared with undifferentiated (U) HC11 cells treated with epidermal growth factor and insulin. (d) Western blot of HSP90α protein extracts from undifferentiated, competent (C) HC11 cells or after DIP treatment for the time indicated. Production of beta-casein is observed after 96 hours when the cells are fully differentiated. Longer exposures show a small amount of beta-casein after 72 hours.
We then investigated whether there was an increase in the encoded protein HSP90α in prolactin-treated SKBR3 cells. SKBR3 cells were cultured in the absence or presence of prolactin and protein extracts were resolved by SDS-PAGE. Western blotting using antibodies directed against HSP90α indicated that there is a two-fold increase in the amount of HSP90α protein in prolactin treated cells (Figure 2b). Prolactin therefore induces both the expression of HSP90A mRNA and increases HSP90α protein in breast cancer cells.
We also investigated the response of the HSP90A gene in mouse mammary epithelial HC11 cells. HC11 cells exist in an undifferentiated state until competent cells are stimulated appropriately with lactogenic hormones, dexamethasone, insulin and prolactin, to differentiate and produce milk proteins [55]. We compared the levels of HSP90A mRNA in HC11 cells in their undifferentiated state and after a one-hour treatment of competent cells with lactogenic hormones, including prolactin (Figure 2c), simulating the time point used in SKBR3 cells. Lactogenic hormone induction resulted in a rapid four-fold increase of HSP90A mRNA in HC11 cells, as quantified by phosphoimager analysis of northern blots using actin as a loading control. We also observed up to a two-fold increase in HSP90α protein, peaking at 48 hours of lactogenic hormone induction of HC11 cells (Figure 2d). This demonstrated that the HSP90A mRNA and protein are elevated during early mammary epithelial cell differentiation in response to lactogenic hormones.
The promoter of the HSP90Agene is preferentially activated by STAT5B
Inspection of the human HSP90A gene upstream sequence indicated the presence of at least two potential STAT-binding DNA elements that could bind STAT1, STAT3 or STAT5 [56] (nucleotides 1611-1603 and 1177–1185, [GenBank:U25822]). Although STAT1 and STAT3 have been reported to respond to prolactin, the prolactin signal is mainly conferred through the activation of STAT5A and STAT5B, two highly homologous members of the STAT family [57]. In order to investigate the prolactin responsiveness of the HSP90A gene promoter, we conducted reporter assays using a gene construct containing about 1.8 kb of the human HSP90A upstream regulatory sequence fused to a luciferase reporter gene. We transfected HeLa cells with expression vectors for the long form of the human prolactin-receptor, the HSP90A-luciferase reporter construct and various STATs.
Prolactin activation of cells transfected with STAT5B, or STAT5A and STAT5B together, caused over a four-fold or an over two-fold increase in luciferase activity, respectively, when compared with cells without exogenous STAT5 expression (Figure 3a). As the induction in the presence of exogenous STAT5A alone was not statistically significant, the significance of the STAT5A/5B result may be due to the presence of STAT5B. The smaller effect of the combination of STAT5A/5B over STAT5B alone may be due to the sequestering of STAT5B through the formation of heterodimers. We also performed reporter assays in both COS-7 as well as SKBR3 cells and obtained similar results with respect to the preferential transcription of the reporter by STAT5B rather than STAT5A (data not shown). In HeLa cells, a small effect of STAT1 and no effect of STAT3 on luciferase activity were observed (Figure 3b). STAT5B is the predominant form of STAT5 in breast tumour cell lines including SKBR3 [58] and most likely contributes to the elevated expression of HSP90α in breast cancer cells. These reporter assays confirm that HSP90A is a prolactin-STAT5 regulated target gene.

Prolactin-STAT5 induction of a HSP90A promoter-luciferase reporter gene. HeLa cells were transiently cotransfected with DNA encoding the prolactin-receptor, the indicated signal transducers and activators of transcription (STATs), the heat shock protein 90 alpha (HSP90A)-luciferase reporter and β-galactosidase. Luciferase assays were performed 48 hours post transfection and the luciferase activity values normalised with β-galactosidase levels. Fold induction was calculated using the normalised luciferase activity from transfected cells in the absence of prolactin. An asterix indicates that the results are significantly different (p < 0.05 t-test) compared with the sample with no STAT proteins. Each bar represents the average of three to five experiments with standard deviation. (a) STAT5A and/or STAT5B. (b) STAT1 and/or STAT3.
Prolactin acts as a survival factor in differentiated HC11 cells
Following the line of reasoning that prolactin acts as a survival factor in breast cancer cells [59–63] and that STAT5 promotes survival in normal mammary epithelial cells [64, 65], we tested whether the addition of prolactin to starved, untransformed mammary epithelial HC11 cells would rescue differentiated cells from apoptosis. HC11 cells were induced to differentiate after they reached confluence by the addition of the lactogenic hormones prolactin, dexamethasone and insulin. Differentiated HC11 cells were then starved of serum and lactogenic hormones with individual hormones returned as indicated for 72 hours, followed by analysis of mono- and oligo-nucleosomes as an indicator of apoptosis (Figure 4). Serum and hormone withdrawal of HC11 cells is known to induce apoptosis [66]. Each of the lactogenic hormones, including prolactin, greatly protects HC11 cells equally well from apoptosis when serum and other lactogenic hormones are removed.

Prolactin is a survival factor for HC11 cells. Differentiated HC11 cells were starved of serum and hormones and with specific hormones added back alone as indicated. After 72 hours, cytoplasmic extracts were prepared and analysed by sandwich ELISA for apoptotic mono- and oligo-nuclesomes. All rescue treatments of prolactin (P), dexamethasone (D) or insulin (I) significantly reduced apoptosis caused by starvation of hormones and serum (pos), for example, between prolactin rescue and positive control, p = 0.001 in a t-test. Negative control (neg) are cells in differentiation medium (DIP) without starvation. Each bar represents the average of three experiments with standard deviation.
Constitutive HSP90Aexpression sensitises mammary epithelial cells to apoptosis in starved HC11 cells
We reasoned that HSP90α would have important functions downstream of prolactin not only in cancerous breast cells, but also in untransformed mammary epithelial cells, such as HC11. For this purpose, we created HC11 cell lines that either constitutively expressed the gene for human HSP90α (HC11-HSP90α) or carried the empty vector (HC11-EV). There is a two-fold increase in HSP90α in the HC11-HSP90α line. Given that we showed that prolactin is a survival factor in HC11 cells, we then investigated whether the HC11-HSP90α cells were susceptible to apoptosis induced by the removal of prolactin and other survival factors. We used two independent methods.
First, we used an antibody against phosphorylated-histone 2A.X as a marker of the apoptotic DNA damage [67] that occurs in response to starvation. As expected, there was little to no indication of phosphorylated-histone 2A.X in the undifferentiated, competent or differentiated cells, which are cultured in the presence of serum and hormones (Figure 5a). Differentiated cells were then starved of serum and all lactogenic hormones for up to 48 hours to induce apoptosis. The HC11-HSP90α cell lines were more sensitive to starvation than the parental HC11 cells or cells expressing the empty vector. After 24 hours of serum and hormone withdrawal, phospho-histone 2A.X is easily detected by western blot in the HC11-HSP90α cells, but not as easily in the control HC11 cells (Figure 5b) or cells carrying the empty vector (HC11-EV) (not shown as the response was similar).

Constitutive HSP90A expression sensitizes cells to apoptosis. (a) Protein extracts were prepared from control parental HC11 or heat shock protein 90 alpha (HSP90α)-HC11 (line 2) cells that were cultured as undifferentiated (U), competent (C) or differentiated (D) cells. (b, c) Differentiated parental HC11 and two different pools of HC11-HSP90α cells (lines 1 and/or 2) were induced to differentiate and then starved of lactogenic hormones and serum for the time indicated. Equal amounts of protein were resolved by SDS-PAGE (15%). An antibody was used to detect phosphorylated-histone 2A.X, and an antibody against GRB2 was used as a loading control. (d) Differentiated control HC11 and HC11-HSP90α cells (line 2) were starved or not apoptosis was assessed by the relative quantities of mono- and oligo-nucleosomes (apoptotic index). Each bar represents the average of three experiments with standard deviation. t-test, *p = 0.02 at 24 hours, **p = 0.00004 at 48 hours.
We then investigated whether the amount of HSP90α differs between the differentiated and starved cells. When cells are starved for 24 hours, the amount of HSP90α in control cells decreases slightly, but remains more constant in the cells constitutively expressing HSP90A, HC11-HSP90α cells (Figure 5c). We can not be certain that the effect on survival we observed (shown in Figure 5b) is due to the differential protein levels between the differentiated and starved cells in the two cell lines, or to the overall two-fold elevated levels of HSP90α in the HC11-HSP90α line. The greater amounts of phospho-histone 2A.X at 24 hours of starvation indicate that constitutive expression of the gene encoding HSP90α increases levels of the marker for DNA damage, phospho-histone 2A.X, and may sensitise the cells to apoptosis.
To confirm that the phosphorylation of histone 2A.X represented events that occur during apoptosis, we also qualitatively assayed the mono- and oligo-nucleosomes generated due to apoptotic DNA nuclease activation. As both HC11-HSP90α cell lines behaved similarly, we used only line two for further characterisation. The HC11 cell lines were treated as shown in Figure 5b, and equal amounts of the cytoplasmic extracts were assessed for the presence of mono- and oligo-nucleosomes. HC11-HSP90α cells clearly had higher levels of nucleosomes than control cells HC11-EV (empty vector) after 24 and 48 hours of starvation, indicating greater levels of apoptosis (Figure 5d). These results support that although prolactin acts as a survival factor in the absence of prolactin or other survival factors, HC11 cells constitutively expressing HSP90α are sensitised to starvation-induced apoptosis.
We used the HSP90 inhibitor 17-AAG to confirm our results. First, we confirmed that HSP90α promotes survival in the presence of prolactin and serum. Differentiated HC11-HSP90α (Figure 6a) or HC11-EV cells (Figure 6b) were untreated or treated with 1 μg/mL 17-AAG either in differentiation medium (dexamethasone, insulin and prolactin plus serum) or starvation medium (no serum or hormones) for a total of 24 hours before measuring apoptosis. Consistent with the role of HSP90α promoting survival, inhibition of HSP90 by 17-AAG induced apoptosis in differentiated HC11-HSP90α (Figure 6a) and HC11-EV cells (Figure 6b) in the presence of survival factors such as prolactin and serum. HSP90α also promotes survival in the control HC11-EV cell line in starvation medium, as demonstrated by the increase in apoptosis with the addition of the inhibitor 17-AAG (Figure 6b).

Inhibition of HSP90α by 17-AAG defines roles for survival as well as apoptosis in HC11 cells. The presence of mono- and oligo-nucleosomes was measured as an indication of apoptosis (apoptotic index). (a) HC11-heat shock protein 90 alpha (HSP90α) cells or (b) HC11-EV cells were differentiated and left untreated or treated with 1 μM of 17-allylamino-17-demethoxygeldanamycin (17-AAG) in either differentiation medium (prolactin, dexamethasone and insulin (DIP)) or starvation medium for 24 hours. Comparison of untreated to 17-AAG-treated HC11-HSP90α (t-test *p = 0.003) or HC11-EV (t-test *p = 0.006) cells after differentiation. Comparison of untreated to 17-AAG-treated HC11-HSP90α (t-test *p = 0.04) or HC11-EV (t-test *p = 0.001) cells in the absence of serum and hormones. Each bar represents the average of three experiments with standard deviation. Each cell line was tested independently and so the absolute levels of nucleosomes can be compared only within each panel.
Second, we confirmed that constitutive expression of HSP90α promotes apoptosis in starved cells. Consistent with the effect of constitutive HSP90α expression on the sensitisation of HC11 cells, 17-AAG reduced the starvation-induced apoptosis (Figure 6a). Overall, starvation enhanced the general level of apoptosis in differentiated cells of both cell lines, although the effects of the HSP90 inhibitor were different. Together this confirms our initial observations and indicates that HSP90α functions to promote survival in differentiated cells in the presence of survival factors, but that constitutive expression sensitises these immortal mammary epithelial HC11 cells to starvation-induced apoptosis.
HSP90α promotes survival in breast cancer cells
Increased expression of HSP90α has been reported in breast cancer, including SKBR3 cells [35], and cytotoxicity has also been reported for the use of HSP90α inhibitors in breast cancer cells, including SKBR3 [68–70]. To test the role of HSP90α in SKBR3 cells, we assessed oligo-nucleasome formation in the presence and absence of 17-AAG and in the presence or absence of serum. HSP90α promotes survival, in the presence of serum, as indicated by the increase in apoptosis after treatment with 17-AAG (Figure 7). Interestingly, overall there was less apoptosis in the absence of serum and the amount of apoptosis was independent of 17-AAG. In general, cancer cells are known to be resistant to apoptotic stimuli, but the role of HSP90α in the absence of serum seems to be minimal. HSP90α promotes survival of SKBR3 cells in the presence of serum.

HSP90α promotes survival only in the presence of serum in SKBR3 breast cancer cells. Apoptosis was assessed by measuring the presence of mono- and oligo-nucleosomes as an indication of apoptosis (apoptotic index). Addition of 500 nM 17-allylamino-17-demethoxygeldanamycin (17-AAG) to SKBR3 cells increases apoptosis (t-test *p = 0.01). Starvation alone reduces the overall level of apoptosis observed in SKBR3 cells with serum (untr) (t-test p = 0.004). Each bar represents the average of three experiments with standard deviation. HSAP90α = heat shock protein 90 alpha.
Discussion
We used a subtractive hybridisation approach to identify a number of prolactin-regulated target genes in the human breast cancer cell line SKBR3. By focusing on the HSP90A gene in particular, we determined that the HSP90α protein has the potential to regulate survival differently in normal (immortal) mammary epithelial cells depending on the context of the hormonal milieu and the constitutive expression of the gene encoding HSP90α. As HSP90α is a central therapeutic target for breast cancer treatment, this also elevates the importance of prolactin and specifically identifies one mechanism for its contribution to breast cancer.
Prolactin regulated genes in breast cancer cells
We identified a number of genes in our screen of breast cancer SKBR3 cells whose expression either increased or decreased in response to prolactin. Some of the genes have been previously associated with cancer progression (T complex protein-1 beta [71], tetratricopeptide repeat protein-4 [72]), cancer survival (Bax inhibitor 1 [73], mitofusion 2 [74, 75]), heterogeneous nuclear riboprotein A1 [76]), drug resistance (T complex protein-1, HSP70 [77]), or cancer cell migration (Hs1 binding protein [78, 79]). Prolactin signalling has been implicated in each of these phenomena in breast cancer cells though its role in drug resistance has not yet been thoroughly examined.
Prolactin-JAK2-STAT5-regulates HSP90A, a therapeutically important gene
In this study, we also identified HSP90A as a prolactin-induced STAT5-activated target gene. HSP90α is a molecular chaperone of a large number of proteins involved in critical signal transduction pathways. The role of HSP90α downstream of prolactin helps explain the multiple effects of prolactin in normal cells and emphasises the significant contribution of prolactin-JAK2-STAT5 signal transduction to breast cancer.
HSP90 in cancer cells is present in an active form, in a multi-chaperone complex with high ATPase activity, in contrast to the HSP90 in normal cells, which is in an inactive, uncomplexed form. It is thought that these differences account for the high affinity of cancer-associated HSP90α for the inhibitory ATP mimetic drugs such as 17-AAG [38]. Chemotherapeutic drugs, such as 17-AAG, inhibit HSP90 and usually result in the degradation of HSP90 client proteins [80]. There are multiple client proteins of HSP90α, including steroid hormone receptors such as oestrogen receptor, protein kinases, cell cycle proteins and transcription factors that are essential targets in cancerous cell growth, survival, immortalisation, angiogenesis and metastasis [81]. The prolactin-mediated induction of HSP90α implicates prolactin in the acquisition or maintenance of each of those cancer-related traits.
Prolactin is a survival factor in HC11 cells
We also determined that prolactin, dexamethasone and insulin, each act as survival factors in differentiated mammary epithelial HC11 cells. Insulin [66, 82] and the glucocorticoid receptor have previously been identified as survival factors [83, 84]. There is existing evidence showing a survival role for prolactin in breast cancer cells [59–63], and for STAT5 in normal mammary epithelial cells [64, 65]. The survival function of prolactin is partially due to the prolactin-mediated activation of AKT/protein kinase B [85, 86]. AKT/protein kinase B is a survival factor and a regulator of mammary gland involution, as transgenic mice expressing constitutively active AKT/protein kinase B in the mammary gland showed delayed involution and a delayed onset of apoptosis [87, 88]. AKT/protein kinase B is also a client protein of HSP90 [89, 90].
HSP90α promotes survival depending on the cellular context
The prolactin-JAK2-STAT5 target gene, HSP90α, can contribute to survival in the presence of prolactin, but sensitises untransformed mammary epithelial HC11 cells to starvation-induced apoptosis when constitutively expressed. We confirmed these results with the use of 17-AAG.
There is evidence for the pro-apoptotic function of HSP90α in other cell types [91–93]. We can also hypothesise that the pro-apoptotic function is due to the action of one of the client proteins either stabilised under these conditions, such as mutant p53, or disengagement from one of its client proteins such as AKT/protein kinase B. Although it is known that HSP90 stabilises mutant p53 forms [94], many of these mutant forms contribute to cellular immortalisation and transformation. The mutant forms of p53 in HC11 cells are thought to contribute to their immortalisation [95], but it is not known if, under certain conditions, mutant p53 could contribute to the sensitisation of cells to starvation-induced apoptosis as does wild-type p53 [96]. The fact that HSP90 can stabilise mutant p53 forms that can contribute to immortalisation and transformation emphasises a contribution of prolactin to these functions, as one of its upstream inducers.
We propose that the switch from survival to apoptosis in untransformed cells involves the loss of survival factors and the availability of HSP90α. In contrast, the switch is absent in SKBR3 breast cancer cells, which do not respond to 17-AAG during serum starvation. HSP90 is important for proliferation and survival, as SKBR3 cells have been shown to respond to 17-AAG by a reduction in proliferation [68, 70] and an increase in apoptosis [68, 69]. This latter observation is consistent with our results in this report. Possible mechanisms involved in loss of HSP90-mediated survival after 17-AAG treatment include the loss of AKT/protein kinase B [97] or ERBB2 [69, 98, 99].
Together with our results, this indicates that in addition to a role in survival, HSP90α also has a pro-apoptotic role that may be cell-type specific, specific to the hormone milieu in the environment or specific to the cellular state of transformation and complement of tumour-suppressor proteins. We hypothesise that HSP90α together with prolactin-mediated events support survival in differentiated or cancerous cells, whereas HSP90α alone may sensitise differentiated mammary cells to wild-type p53-independent apoptosis depending on the cellular context.
Conclusion
The evidence for a contribution of prolactin and STAT5 to breast cancer cell survival, breast cancer progression and to chemotherapeutic response is strengthened by our observations that prolactin treatment of human breast cancer cells regulates a number of genes associated with cancer progression, including the therapeutically important target gene, HSP90A. HSP90α is important for malignant progression in breast cancer, but when elevated in untransformed mammary epithelial cells may participate in a switch between survival and apoptosis.
Abbreviations
- 17-AAG:
-
17-allylamino-17-demethoxygeldanamycin
- bp:
-
base pairs
- CIS :
-
cytokine inducible SH2 containing protein
- EGF:
-
epidermal growth factor
- ERBB2:
-
erythroblastic leukaemia viral oncogene homolog 2
- HRP:
-
horseradish peroxidase
- HSP90:
-
heat shock protein 90
- JAK2:
-
Janus kinase-2
- STAT:
-
signal transducers and activators of transcription.
References
Shemanko CS, Groner B: Transcription factors, cofactors and target genes mediating prolactin action. Prolactin. Edited by: Horseman ND. 2001, USA: Kluwer Academic Publishers, 381-404.
Watson CJ: Involution: apoptosis and tissue remodelling that convert the mammary gland from milk factory to a quiescent organ. Breast Cancer Res. 2006, 8: 203-10.1186/bcr1401.
Clarkson RW, Boland MP, Kritikou EA, Lee JM, Freeman TC, Tiffen PG, Watson CJ: The genes induced by signal transducer and activators of transcription (STAT)3 and STAT5 in mammary epithelial cells define the roles of these STATs in mammary development. Mol Endocrinol. 2006, 20: 675-685. 10.1210/me.2005-0392.
Tworoger SS, Eliassen AH, Rosner B, Sluss P, Hankinson SE: Plasma prolactin concentrations and risk of postmenopausal breast cancer. Cancer Res. 2004, 64: 6814-6819. 10.1158/0008-5472.CAN-04-1870.
Nouhi Z, Chughtai N, Hartley S, Cocolakis E, Lebrun JJ, Ali S: Defining the role of prolactin as an invasion suppressor hormone in breast cancer cells. Cancer Res. 2006, 66: 1824-1832. 10.1158/0008-5472.CAN-05-2292.
Clevenger CV, Furth PA, Hankinson SE, Schuler LA: The role of prolactin in mammary carcinoma. Endocr Rev. 2003, 24: 1-27. 10.1210/er.2001-0036.
Tworoger SS, Hankinson SE: Prolactin and breast cancer etiology: An epidemiologic perspective. J Mammary Gland Biol Neoplasia. 2008, 13: 41-53. 10.1007/s10911-008-9063-y.
Arendt LM, Rose-Hellekant TA, Sandgren EP, Schuler LA: Prolactin potentiates transforming growth factor alpha induction of mammary neoplasia in transgenic mice. Am J Pathol. 2006, 168: 1365-1374. 10.2353/ajpath.2006.050861.
Rose-Hellekant TA, Arendt LM, Schroeder MD, Gilchrist K, Sandgren EP, Schuler LA: Prolactin induces ERalpha-positive and ERalpha-negative mammary cancer in transgenic mice. Oncogene. 2003, 22: 4664-4674. 10.1038/sj.onc.1206619.
Wennbo H, Gebre-Medhin M, Gritli-Linde A, Ohlsson C, Isaksson OG, Tornell J: Activation of the prolactin receptor but not the growth hormone receptor is important for induction of mammary tumors in transgenic mice. J Clin Invest. 1997, 100: 2744-2751. 10.1172/JCI119820.
Vomachka AJ, Pratt SL, Lockefeer JA, Horseman ND: Prolactin gene-disruption arrests mammary gland development and retards T-antigen-induced tumor growth. Oncogene. 2000, 19: 1077-1084. 10.1038/sj.onc.1203348.
Oakes SR, Robertson FG, Kench JG, Gardiner-Garden M, Wand MP, Green JE, Ormandy CJ: Loss of mammary epithelial prolactin receptor delays tumor formation by reducing cell proliferation in low-grade preinvasive lesions. Oncogene. 2007, 26: 543-553. 10.1038/sj.onc.1209838.
Tworoger SS, Eliassen AH, Sluss P, Hankinson SE: A prospective study of plasma prolactin concentrations and risk of premenopausal and postmenopausal breast cancer. J Clin Oncol. 2007, 25: 1482-1488. 10.1200/JCO.2006.07.6356.
Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R, Ghosh D, Barrette T, Pandey A, Chinnaiyan AM: ONCOMINE: a cancer microarray database and integrated data-mining platform. Neoplasia. 2004, 6: 1-6.
Bertucci F, Nasser V, Granjeaud S, Eisinger F, Adelaide J, Tagett R, Loriod B, Giaconia A, Benziane A, Devilard E, Jacquemier J, Viens P, Nguyen C, Birnbaum D, Houlgatte R: Gene expression profiles of poor-prognosis primary breast cancer correlate with survival. Hum Mol Genet. 2002, 11: 863-872. 10.1093/hmg/11.8.863.
Turkson J: STAT proteins as novel targets for cancer drug discovery. Expert Opin Ther Targets. 2004, 8: 409-422. 10.1517/14728222.8.5.409.
Wagner KU, Rui H: Jak2/Stat5 signaling in mammogenesis, breast cancer initiation and progression. J Mammary Gland Biol Neoplasia. 2008, 13: 93-103. 10.1007/s10911-008-9062-z.
Brisken C, Ayyannan A, Nguyen C, Heineman A, Reinhardt F, Tan J, Dey SK, Dotto GP, Weinberg RA, Jan T: IGF-2 is a mediator of prolactin-induced morphogenesis in the breast. Dev Cell. 2002, 3: 877-887. 10.1016/S1534-5807(02)00365-9.
Gass S, Harris J, Ormandy C, Brisken C: Using gene expression arrays to elucidate transcriptional profiles underlying prolactin function. J Mammary Gland Biol Neoplasia. 2003, 8: 269-285. 10.1023/B:JOMG.0000010029.85796.63.
Harris J, Stanford PM, Sutherland K, Oakes SR, Naylor MJ, Robertson FG, Blazek KD, Kazlauskas M, Hilton HN, Wittlin S, Alexander WS, Lindeman GJ, Visvader JE, Ormandy CJ: Socs2 and elf5 mediate prolactin-induced mammary gland development. Mol Endocrinol. 2006, 20: 1177-1187. 10.1210/me.2005-0473.
Hou Z, Bailey JP, Vomachka AJ, Matsuda M, Lockefeer JA, Horseman ND: Glycosylation-dependent cell adhesion molecule 1 (GlyCAM 1) is induced by prolactin and suppressed by progesterone in mammary epithelium. Endocrinology. 2000, 141: 4278-4283. 10.1210/en.141.11.4278.
Naylor MJ, Ginsburg E, Iismaa TP, Vonderhaar BK, Wynick D, Ormandy CJ: The neuropeptide galanin augments lobuloalveolar development. J Biol Chem. 2003, 278: 29145-29152. 10.1074/jbc.M303746200.
Naylor MJ, Oakes SR, Gardiner-Garden M, Harris J, Blazek K, Ho TW, Li FC, Wynick D, Walker AM, Ormandy CJ: Transcriptional changes underlying the secretory activation phase of mammary gland development. Mol Endocrinol. 2005, 19: 1868-1883. 10.1210/me.2004-0254.
Ormandy CJ, Naylor M, Harris J, Robertson F, Horseman ND, Lindeman GJ, Visvader J, Kelly PA: Investigation of the transcriptional changes underlying functional defects in the mammary glands of prolactin receptor knockout mice. Recent Prog Horm Res. 2003, 58: 297-323. 10.1210/rp.58.1.297.
Neckers L: Heat shock protein 90: the cancer chaperone. J Biosci. 2007, 32: 517-530. 10.1007/s12038-007-0051-y.
Workman P, Burrows F, Neckers L, Rosen N: Drugging the cancer chaperone HSP90: combinatorial therapeutic exploitation of oncogene addiction and tumor stress. Ann N Y Acad Sci. 2007, 1113: 202-216. 10.1196/annals.1391.012.
Beliakoff J, Whitesell L: Hsp90: an emerging target for breast cancer therapy. Anticancer Drugs. 2004, 15: 651-662. 10.1097/01.cad.0000136876.11928.be.
Cullinan SB, Whitesell L: Heat shock protein 90: a unique chemotherapeutic target. Semin Oncol. 2006, 33: 457-465. 10.1053/j.seminoncol.2006.04.001.
Workman P, Maloney A: HSP90 as a new therapeutic target for cancer therapy: the story unfolds. Expert Opin Biol Ther. 2002, 2: 3-24. 10.1517/14712598.2.1.3.
Queitsch C, Sangster TA, Lindquist S: Hsp90 as a capacitor of phenotypic variation. Nature. 2002, 417: 618-624. 10.1038/nature749.
Rutherford SL, Lindquist S: Hsp90 as a capacitor for morphological evolution. Nature. 1998, 396: 336-342. 10.1038/24550.
Eustace BK, Sakurai T, Stewart JK, Yimlamai D, Unger C, Zehetmeier C, Lain B, Torella C, Henning SW, Beste G, Scroggins BT, Neckers L, Ilag LL, Jay DG: Functional proteomic screens reveal an essential extracellular role for hsp90 alpha in cancer cell invasiveness. Nat Cell Biol. 2004, 6: 507-514. 10.1038/ncb1131.
Teng SC, Chen YY, Su YN, Chou PC, Chiang YC, Tseng SF, Wu KJ: Direct activation of HSP90A transcription by c-Myc contributes to c-Myc-induced transformation. J Biol Chem. 2004, 279: 14649-14655. 10.1074/jbc.M308842200.
Csermely P, Schnaider T, Soti C, Prohaszka Z, Nardai G: The 90-kDa molecular chaperone family: structure, function, and clinical applications. A comprehensive review. Pharmacol Ther. 1998, 79: 129-168. 10.1016/S0163-7258(98)00013-8.
Pick E, Kluger Y, Giltnane JM, Moeder C, Camp RL, Rimm DL, Kluger HM: High HSP90 expression is associated with decreased survival in breast cancer. Cancer Res. 2007, 67: 2932-2937. 10.1158/0008-5472.CAN-06-4511.
Yano M, Naito Z, Tanaka S, Asano G: Expression and roles of heat shock proteins in human breast cancer. Jpn J Cancer Res. 1996, 87: 908-915.
Yano M, Naito Z, Yokoyama M, Shiraki Y, Ishiwata T, Inokuchi M, Asano G: Expression of hsp90 and cyclin D1 in human breast cancer. Cancer Lett. 1999, 137: 45-51. 10.1016/S0304-3835(98)00338-3.
Kamal A, Thao L, Sensintaffar J, Zhang L, Boehm MF, Fritz LC, Burrows FJ: A high-affinity conformation of Hsp90 confers tumour selectivity on Hsp90 inhibitors. Nature. 2003, 425: 407-410. 10.1038/nature01913.
Powers MV, Workman P: Targeting of multiple signalling pathways by heat shock protein 90 molecular chaperone inhibitors. Endocr Relat Cancer. 2006, 13 (Suppl 1): S125-135. 10.1677/erc.1.01324.
Modi S, Stopeck AT, Gordon MS, Mendelson D, Solit DB, Bagatell R, Ma W, Wheler J, Rosen N, Norton L, Cropp GF, Johnson RG, Hannah AL, Hudis CA: Combination of trastuzumab and tanespimycin (17-AAG, KOS-953) is safe and active in trastuzumab-refractory HER-2 overexpressing breast cancer: a phase I dose-escalation study. J Clin Oncol. 2007, 25: 5410-5417. 10.1200/JCO.2007.11.7960.
Sharp S, Workman P: Inhibitors of the HSP90 molecular chaperone: current status. Adv Cancer Res. 2006, 95: 323-348. 10.1016/S0065-230X(06)95009-X.
Tuna M, Chavez-Reyes A, Tari AM: HER2/neu increases the expression of Wilms' Tumor 1 (WT1) protein to stimulate S-phase proliferation and inhibit apoptosis in breast cancer cells. Oncogene. 2005, 24: 1648-1652. 10.1038/sj.onc.1208345.
Liu X, Robinson GW, Gouilleux F, Groner B, Hennighausen L: Cloning and expression of Stat5 and an additional homologue (Stat5b) involved in prolactin signal transduction in mouse mammary tissue. Proc Natl Acad Sci USA. 1995, 92: 8831-8835. 10.1073/pnas.92.19.8831.
Wakao H, Gouilleux F, Groner B: Mammary gland factor (MGF) is a novel member of the cytokine regulated transcription factor gene family and confers the prolactin response. Embo J. 1994, 13: 2182-2191.
Ball RK, Friis RR, Schoenenberger CA, Doppler W, Groner B: Prolactin regulation of beta-casein gene expression and of a cytosolic 120-kd protein in a cloned mouse mammary epithelial cell line. Embo J. 1988, 7: 2089-2095.
Uchida K, Yoshimura A, Inazawa J, Yanagisawa K, Osada H, Masuda A, Saito T, Takahashi T, Miyajima A: Molecular cloning of CISH, chromosome assignment to 3p21.3, and analysis of expression in fetal and adult tissues. Cytogenet Cell Genet. 1997, 78: 209-212. 10.1159/000134658.
Wakao H, Schmitt-Ney M, Groner B: Mammary gland-specific nuclear factor is present in lactating rodent and bovine mammary tissue and composed of a single polypeptide of 89 kDa. J Biol Chem. 1992, 267: 16365-16370.
Gouilleux F, Wakao H, Mundt M, Groner B: Prolactin induces phosphorylation of Tyr694 of Stat5 (MGF), a prerequisite for DNA binding and induction of transcription. Embo J. 1994, 13: 4361-4369.
Shaw-Bruha CM, Pirrucello SJ, Shull JD: Expression of the prolactin gene in normal and neoplastic human breast tissues and human mammary cell lines: promoter usage and alternative mRNA splicing. Breast Cancer Res Treat. 1997, 44: 243-253. 10.1023/A:1005879103367.
Yoshimura A, Ohkubo T, Kiguchi T, Jenkins NA, Gilbert DJ, Copeland NG, Hara T, Miyajima A: A novel cytokine-inducible gene CIS encodes an SH2-containing protein that binds to tyrosine-phosphorylated interleukin 3 and erythropoietin receptors. Embo J. 1995, 14: 2816-2826.
Matsumoto A, Masuhara M, Mitsui K, Yokouchi M, Ohtsubo M, Misawa H, Miyajima A, Yoshimura A: CIS, a cytokine inducible SH2 protein, is a target of the JAK-STAT5 pathway and modulates STAT5 activation. Blood. 1997, 89: 3148-3154.
Verdier F, Rabionet R, Gouilleux F, Beisenherz-Huss C, Varlet P, Muller O, Mayeux P, Lacombe C, Gisselbrecht S, Chretien S: A sequence of the CIS gene promoter interacts preferentially with two associated STAT5A dimers: a distinct biochemical difference between STAT5A and STAT5B. Mol Cell Biol. 1998, 18: 5852-5860.
Bole-Feysot C, Perret E, Roustan P, Bouchard B, Kelly PA: Analysis of prolactin-modulated gene expression profiles during the Nb2 cell cycle using differential screening techniques. Genome Biol. 2000, 1: RESEARCH0008-10.1186/gb-2000-1-4-research0008.
Djiane J, Houdebine LM, Kelly PA: Correlation between prolactin-receptor interaction, down-regulation of receptors, and stimulation of casein and deoxyribonucleic acid biosynthesis in rabbit mammary gland explants. Endocrinology. 1982, 110: 791-795.
Desrivières S, Prinz T, Castro-Palomino Laria N, Meyer M, Boehm G, Bauer U, Schäfer J, Neumann T, Groner B, Shemanko CS: Comparative proteomic analysis of proliferating and functionally differentiated mammary epithelial cells. Molecular and Cellular Proteomics. 2003, 2: 1039-1054. 10.1074/mcp.M300032-MCP200.
Decker T, Kovarik P, Meinke A: GAS elements: a few nucleotides with a major impact on cytokine-induced gene expression. J Interferon Cytokine Res. 1997, 17: 121-134.
Calo V, Migliavacca M, Bazan V, Macaluso M, Buscemi M, Gebbia N, Russo A: STAT proteins: from normal control of cellular events to tumorigenesis. J Cell Physiol. 2003, 197: 157-168. 10.1002/jcp.10364.
Weaver AM, Silva CM: Modulation of signal transducer and activator of transcription 5b activity in breast cancer cells by mutation of tyrosines within the transactivation domain. Mol Endocrinol. 2006, 20: 2392-2405. 10.1210/me.2005-0418.
Abdelmagid SA, Too CK: Prolactin and Estrogen Upregulate Carboxypeptidase-D to Promote Nitric Oxide Production and Survival of MCF-7 Breast Cancer Cells. Endocrinology. 2008
Chakravarti P, Henry MK, Quelle FW: Prolactin and heregulin override DNA damage-induced growth arrest and promote phosphatidylinositol-3 kinase-dependent proliferation in breast cancer cells. Int J Oncol. 2005, 26: 509-514.
Chen WY, Ramamoorthy P, Chen N, Sticca R, Wagner TE: A human prolactin antagonist, hPRL-G129R, inhibits breast cancer cell proliferation through induction of apoptosis. Clin Cancer Res. 1999, 5: 3583-3593.
Ginsburg E, Vonderhaar BK: Prolactin synthesis and secretion by human breast cancer cells. Cancer Res. 1995, 55: 2591-2595.
Perks CM, Keith AJ, Goodhew KL, Savage PB, Winters ZE, Holly JM: Prolactin acts as a potent survival factor for human breast cancer cell lines. Br J Cancer. 2004, 91: 305-311.
Cui Y, Riedlinger G, Miyoshi K, Tang W, Li C, Deng CX, Robinson GW, Hennighausen L: Inactivation of Stat5 in mouse mammary epithelium during pregnancy reveals distinct functions in cell proliferation, survival, and differentiation. Mol Cell Biol. 2004, 24: 8037-8047. 10.1128/MCB.24.18.8037-8047.2004.
Iavnilovitch E, Eilon T, Groner B, Barash I: Expression of a carboxy terminally truncated Stat5 with no transactivation domain in the mammary glands of transgenic mice inhibits cell proliferation during pregnancy, delays onset of milk secretion, and induces apoptosis upon involution. Mol Reprod Dev. 2006, 73: 841-849. 10.1002/mrd.20479.
Merlo GR, Basolo F, Fiore L, Duboc L, Hynes NE: p53-dependent and p53-independent activation of apoptosis in mammary epithelial cells reveals a survival function of EGF and insulin. J Cell Biol. 1995, 128: 1185-1196. 10.1083/jcb.128.6.1185.
Rogakou EP, Nieves-Neira W, Boon C, Pommier Y, Bonner WM: Initiation of DNA fragmentation during apoptosis induces phosphorylation of H2AX histone at serine 139. J Biol Chem. 2000, 275: 9390-9395. 10.1074/jbc.275.13.9390.
Munster PN, Marchion DC, Basso AD, Rosen N: Degradation of HER2 by ansamycins induces growth arrest and apoptosis in cells with HER2 overexpression via a HER3, phosphatidylinositol 3'-kinase-AKT-dependent pathway. Cancer Res. 2002, 62: 3132-3137.
Solit DB, Basso AD, Olshen AB, Scher HI, Rosen N: Inhibition of heat shock protein 90 function down-regulates Akt kinase and sensitizes tumors to Taxol. Cancer Res. 2003, 63: 2139-2144.
Zsebik B, Citri A, Isola J, Yarden Y, Szollosi J, Vereb G: Hsp90 inhibitor 17-AAG reduces ErbB2 levels and inhibits proliferation of the trastuzumab resistant breast tumor cell line JIMT-1. Immunol Lett. 2006, 104: 146-155. 10.1016/j.imlet.2005.11.018.
Coghlin C, Carpenter B, Dundas SR, Lawrie LC, Telfer C, Murray GI: Characterization and over-expression of chaperonin t-complex proteins in colorectal cancer. J Pathol. 2006, 210: 351-357. 10.1002/path.2056.
Poetsch M, Dittberner T, Cowell JK, Woenckhaus C: TTC4, a novel candidate tumor suppressor gene at 1p31 is often mutated in malignant melanoma of the skin. Oncogene. 2000, 19: 5817-5820. 10.1038/sj.onc.1203961.
Grzmil M, Thelen P, Hemmerlein B, Schweyer S, Voigt S, Mury D, Burfeind P: Bax inhibitor-1 is overexpressed in prostate cancer and its specific down-regulation by RNA interference leads to cell death in human prostate carcinoma cells. Am J Pathol. 2003, 163: 543-552.
Brooks C, Wei Q, Feng L, Dong G, Tao Y, Mei L, Xie ZJ, Dong Z: Bak regulates mitochondrial morphology and pathology during apoptosis by interacting with mitofusins. Proc Natl Acad Sci USA. 2007, 104: 11649-11654. 10.1073/pnas.0703976104.
Neuspiel M, Zunino R, Gangaraju S, Rippstein P, McBride H: Activated mitofusin 2 signals mitochondrial fusion, interferes with Bax activation, and reduces susceptibility to radical induced depolarization. J Biol Chem. 2005, 280: 25060-25070. 10.1074/jbc.M501599200.
Patry C, Bouchard L, Labrecque P, Gendron D, Lemieux B, Toutant J, Lapointe E, Wellinger R, Chabot B: Small interfering RNA-mediated reduction in heterogeneous nuclear ribonucleoparticule A1/A2 proteins induces apoptosis in human cancer cells but not in normal mortal cell lines. Cancer Res. 2003, 63: 7679-7688.
Verrills NM, Liem NL, Liaw TY, Hood BD, Lock RB, Kavallaris M: Proteomic analysis reveals a novel role for the actin cytoskeleton in vincristine resistant childhood leukemia – an in vivo study. Proteomics. 2006, 6: 1681-1694. 10.1002/pmic.200500417.
Radhika V, Onesime D, Ha JH, Dhanasekaran N: Galpha13 stimulates cell migration through cortactin-interacting protein Hax-1. J Biol Chem. 2004, 279: 49406-49413. 10.1074/jbc.M408836200.
Ramsay AG, Keppler MD, Jazayeri M, Thomas GJ, Parsons M, Violette S, Weinreb P, Hart IR, Marshall JF: HS1-associated protein X-1 regulates carcinoma cell migration and invasion via clathrin-mediated endocytosis of integrin alphavbeta6. Cancer Res. 2007, 67: 5275-5284. 10.1158/0008-5472.CAN-07-0318.
Whitesell L, Lindquist SL: HSP90 and the chaperoning of cancer. Nat Rev Cancer. 2005, 5: 761-772. 10.1038/nrc1716.
Workman P: Combinatorial attack on multistep oncogenesis by inhibiting the Hsp90 molecular chaperone. Cancer Lett. 2004, 206: 149-157. 10.1016/j.canlet.2003.08.032.
Merlo GR, Graus-Porta D, Cella N, Marte BM, Taverna D, Hynes NE: Growth, differentiation and survival of HC11 mammary epithelial cells: diverse effects of receptor tyrosine kinase-activating peptide growth factors. Eur J Cell Biol. 1996, 70: 97-105.
Wu W, Zou M, Brickley DR, Pew T, Conzen SD: Glucocorticoid receptor activation signals through forkhead transcription factor 3a in breast cancer cells. Mol Endocrinol. 2006, 20: 2304-2314. 10.1210/me.2006-0131.
Schorr K, Furth PA: Induction of bcl-xL expression in mammary epithelial cells is glucocorticoid-dependent but not signal transducer and activator of transcription 5-dependent. Cancer Res. 2000, 60: 5950-5953.
al-Sakkaf KA, Dobson PR, Brown BL: Prolactin induced tyrosine phosphorylation of p59fyn may mediate phosphatidylinositol 3-kinase activation in Nb2 cells. J Mol Endocrinol. 1997, 19: 347-350. 10.1677/jme.0.0190347.
Bailey JP, Nieport KM, Herbst MP, Srivastava S, Serra RA, Horseman ND: Prolactin and transforming growth factor-beta signaling exert opposing effects on mammary gland morphogenesis, involution, and the Akt-forkhead pathway. Mol Endocrinol. 2004, 18: 1171-1184. 10.1210/me.2003-0345.
Hutchinson J, Jin J, Cardiff RD, Woodgett JR, Muller WJ: Activation of Akt (protein kinase B) in mammary epithelium provides a critical cell survival signal required for tumor progression. Mol Cell Biol. 2001, 21: 2203-2212. 10.1128/MCB.21.6.2203-2212.2001.
Schwertfeger KL, Richert MM, Anderson SM: Mammary gland involution is delayed by activated Akt in transgenic mice. Mol Endocrinol. 2001, 15: 867-881. 10.1210/me.15.6.867.
Basso AD, Solit DB, Chiosis G, Giri B, Tsichlis P, Rosen N: Akt forms an intracellular complex with heat shock protein 90 (Hsp90) and Cdc37 and is destabilized by inhibitors of Hsp90 function. J Biol Chem. 2002, 277: 39858-39866. 10.1074/jbc.M206322200.
Sato S, Fujita N, Tsuruo T: Modulation of Akt kinase activity by binding to Hsp90. Proc Natl Acad Sci USA. 2000, 97: 10832-10837. 10.1073/pnas.170276797.
Galea-Lauri J, Richardson AJ, Latchman DS, Katz DR: Increased heat shock protein 90 (hsp90) expression leads to increased apoptosis in the monoblastoid cell line U937 following induction with TNF-alpha and cycloheximide: a possible role in immunopathology. J Immunol. 1996, 157: 4109-4118.
Sapozhnikov AM, Ponomarev ED, Tarasenko TN, Telford WG: Spontaneous apoptosis and expression of cell surface heat-shock proteins in cultured EL-4 lymphoma cells. Cell Prolif. 1999, 32: 363-378. 10.1111/j.1365-2184.1999.tb01354.x.
Wu YP, Kita K, Suzuki N: Involvement of human heat shock protein 90 alpha in nicotine-induced apoptosis. Int J Cancer. 2002, 100: 37-42. 10.1002/ijc.10449.
Whitesell L, Sutphin PD, Pulcini EJ, Martinez JD, Cook PH: The physical association of multiple molecular chaperone proteins with mutant p53 is altered by geldanamycin, an hsp90-binding agent. Mol Cell Biol. 1998, 18: 1517-1524.
Merlo GR, Venesio T, Taverna D, Marte BM, Callahan R, Hynes NE: Growth suppression of normal mammary epithelial cells by wild-type p53. Oncogene. 1994, 9: 443-453.
Blagosklonny MV, Wu GS, Somasundaram K, E S, W D: Wild-type p53 is not sufficient for serum starvation-induced apoptosis in cancer cells but accelerates apoptosis in sensitive cells. International Journal of Oncology. 1997, 11: 1165-1170.
Basso AD, Solit DB, Munster PN, Rosen N: Ansamycin antibiotics inhibit Akt activation and cyclin D expression in breast cancer cells that overexpress HER2. Oncogene. 2002, 21: 1159-1166. 10.1038/sj.onc.1205184.
Wang K, Ma Q, Ren Y, He J, Zhang Y, Zhang Y, Chen W: Geldanamycin destabilizes HER2 tyrosine kinase and suppresses Wnt/beta-catenin signaling in HER2 overexpressing human breast cancer cells. Oncol Rep. 2007, 17: 89-96.
Xu W, Yuan X, Jung YJ, Yang Y, Basso A, Rosen N, Chung EJ, Trepel J, Neckers L: The heat shock protein 90 inhibitor geldanamycin and the ErbB inhibitor ZD1839 promote rapid PP1 phosphatase-dependent inactivation of AKT in ErbB2 overexpressing breast cancer cells. Cancer Res. 2003, 63: 7777-7784.
Acknowledgements
We would like to thank Margot Landerz for sequencing the large selection of candidate clones and Sabina Vogel for assistance in the initial screening of the library, as well as Drs E. Hickey and Lee Weber for the HSP90A-luciferase reporter gene. The work of CSS, CP and RL was supported by grants from the Natural Sciences and Engineering Research Council of Canada, the Alberta Cancer Foundation and the Alberta Cancer Board.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
CP provided data regarding HSP90α protein levels. RL also participated in characterising HSP90α levels and the response of cells to prolactin. CTP helped with luciferase assays. NB performed EMSA and northern blots. JS and PB provided bioinformatics support. CSS prepared and screened the library, contributing luciferase and apoptosis assays. EP, CSS and BG contributed to early project design. BG and CSS provided funding. CSS provided further project development and wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Perotti, C., Liu, R., Parusel, C.T. et al. Heat shock protein-90-alpha, a prolactin-STAT5 target gene identified in breast cancer cells, is involved in apoptosis regulation. Breast Cancer Res 10, R94 (2008). https://doi.org/10.1186/bcr2193
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/bcr2193