
Abstract
The synovial tissue in rheumatoid arthritis (RA) patients is enriched with mature antigen presenting cells (APCs) and many T lymphocytes. Interactions between APCs and T cells are essential for the initiation and amplification of T-cell-dependent immune responses, and may therefore play an important role in the chronic inflammatory processes in the synovium. The nature of the antigen(s) involved in RA still remains elusive. However, interactions and signaling through the costimulatory molecules CD28-CD80/86 and CD40-CD40L are critical during APC–T cell interaction for optimal cell activation. This review discusses how such costimulatory signals can be involved in the initiation and amplification of the inflammatory reactions in the synovium. Blocking of the signaling pathways involved in APC–T cell interactions might provide a specific immuno-therapeutic approach for the treatment of RA.
Similar content being viewed by others
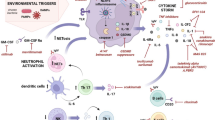

Introduction
It is believed that the pathology and etiology of rheumatoid arthritis involves abnormal presentation of self antigen(s) by APCs and activation of autoreactive T cells [1]. T lymphocytes have therefore been proposed to play a central role in the disease process [2*], the most powerful evidence for which is the association of the disease with certain human leukocyte antigen (HLA)-DR chains that are shared between various alleles of HLA-DR4 and HLA-DR1 [3,4]. The rheumatoid synovial membrane is rich in major histocompatibility complex class II+ APCs, and CD4+ T cells make clusters with such HLA-DR-positive cells [5]. The T cells present in the RA synovium express the activation markers HLA-DR, VLA-4 and CD69. However, it is not clear whether T cell activation occurs before entry to the tissue, during transendothelial migration, or in the synovium. APCs require signals from activated T cells for their differentiation and maturation [6]; this subsequently enables APCs to activate newly arrived T cells in a specific or unspecific manner in the local inflammation (Figure 1). Activated T cells promote the disease progression by inducing the secretion of pro-inflammatory cytokines (in particular, tumor necrosis factor [TNF]-α) from macrophages and synovial cells in a contact-dependent manner [7,8].

Antigen presenting cell-T cell interaction in the synovium. Dendritic cells (DCs) in the synovium present an external or local antigen to T cells. T cell receptor interaction and signaling via CD28-CD80/86 are essential for initial T cell activation leading to upregulation of CD40L on the T cells. Activated T cells are then capable of inducing further differentiation of DCs, as well as T cell help to rheumatoid factor B cells that have taken up antigen-Ig complexes. CD40-CD40L interaction between DCs, B cells and T cells may play a critical role in repeated activation of memory T cells in the synovium and, thus, maintenance of the inflammatory reactions.
Several costimulatory molecules are involved during APC–T cell interactions, including CD28/CD80-86 and CD40-CD40L. Some of these molecules are thought to be critical in initiation of the immune response (CD28/CD80/86), while CD40-CD40L are required for the amplification of the inflammatory response [9**]. This paper reviews the costimulatory molecules and types of APCs involved in local APC–T cell interactions and their possible roles in the disease pathology.
Costimulatory molecules and T cell activation in the synovium
Initial T cell activation is dependent on T cell receptor triggering and costimulatory signaling through CD28-CD80/CD86 [10,11]. APCs such as dendritic cells (DCs), macrophages, B cells and follicular DCs can provide both of these signals. APCs that cannot provide costimulatory signals are unable to activate naïve T cells unless a second signal is provided by accessory APCs. The costimulatory molecules CD80/CD86 are expressed at varying levels by APCs in the synovium. Expression of CD80 is generally low, while the expression of CD86 is relatively high and is expressed on several APCs including DCs, B cells and macrophages [12,13]. It has been suggested that CD80/CD86 interactions influence the development of T helper (Th)1 and Th2 cells, where Th2 polarization is dependent on signals through the CD80 molecule [14*,15]. T cells in the synovium are of Th1 type, and even though the secretion of IFNγ is low, the Th1 cytokine IL-17 can be found in high levels [16,17]. Expression of CD86 and lack of CD80 then correlates with the dominant population of Th1 cells in the synovium [16,18]. APCs in the synovium might therefore be capable of the initial activation of T cells through the Th1 pathway by the expression of CD86 and secretion of IL-12. It has been shown in various models that blocking the CD28 signaling pathway can prevent or treat autoimmune diseases [19,20]. Studies in MLR/lpr mice lacking CD28 suggest that CD28 is most probably a regulator in the induction of autoimmune diseases. Arthritis is abolished and autoantibody production is suppressed in these mice, but the accumulation of abnormal T cells is almost unchanged [21].
The memory population of Th1 cells in the synovium has an increased number of CD28-negative cells [22]. Memory T cells, however, have a lower requirement for CD28 signaling compared with naïve T cells and can respond to T cell receptor signaling alone [23]. A much greater range of APCs can therefore stimulate memory T cells than that which stimulates naïve T cells. In addition, effector T cells are even less dependent on costimulatory signals and can respond to many types of APC including resting B cells and macrophages [24]. APCs in the synovium generally express low levels of CD80/86, but still might function as APCs for memory/effector T cells and maintain the inflammatory response by continuously activating synovial memory T cells.
In addition to CD28/CD86-80 costimulation, CD40-CD40L has been shown to contribute to T cell activation, both by independently costimulating T cells and, at least in part, by upregulating CD80/CD86 molecules on APCs [14*,25]. The CD40 pathway is particularly thought to be important for amplification of the T cell response, as blocking the CD40 signaling pathway has been shown to inhibit Th1 differentiation and maintenance of the immune response [26]. DCs, B cells and macrophages express CD40 in the synovium, and these APCs may be of particular importance for the disease pathogenesis.
DC-T cell interaction
Fully mature DCs are found in the perivascular area in close association with T cells and with B-cell follicles, and express markers associated with mature DCs, including major histocompatibility complex class II molecules, nuclear RelB, adhesion molecules and STAT 4 [5,27]. DCs migrate from peripheral blood, probably under the influence of a chemotactic attractant to the synovial tissue [28]. DCs in the synovium receive differentiation signals from monocyte derived cytokines and through CD40L interaction with T cells. DCs are located adjacent to endothelial cells surrounding blood vessels in the synovium, implying that DCs derived from circulating precursors may have differentiated during a short time after transendothelial migration. If this is so, DCs could function as potent antigen presenting cells, either for external or local antigen(s) released by the ongoing synovial inflammation. Several cytokines are produced in RA synovium, including TNFα, IL-1 and granulocyte macrophage-colony stimulating factor, that can upregulate the ability of DCs to present antigen. The major DC differentiation signal is achieved through ligation of CD40, which triggers high levels of IL-12 secretion and upregulation of CD80, CD86 and ICAM-1 [29,30]. T cells in the synovium express CD40L and, upon interaction with DCs, play an important role in upregulation of the antigen presenting capacity of synovial tissue DCs [31]. A high number of mature DCs are then able to polarize the T cell response into the Th1 pathway, giving the Th1 cells an irreversible phenotype [32].
Antigen presentation by rheumatoid factor B cells and T cell interaction
B cells are another major source of APCs in the synovium. Rheumatoid factor (RF) B cells, in particular, are believed to play an important role in antigen presentation [33**]. RF B cells can take up antigen–Ig immune complexes via their membrane Ig receptors, which have RF specificity and thus react with the IgG Fc part of antibody molecules. The B cells then process and present peptides from the antigen, and thus induce both T cell activation and T cell help for multiple antigens, as well as T cell help for RF secretion (Figure 1). High affinity RFs accumulate in the synovium and contribute to the disease pathogenesis by forming immune complexes and activation of complement [34]. Low affinity RFs are abundant in lymph nodes in healthy individuals, and high affinity RFs are deleted in mice [35,36]. The reason for the persistence of high affinity RFs in RA patients but not in healthy individuals is not fully understood. However, in normals there are mechanisms that prevent replacement mutations which can lead to affinity maturation. Thus, the R:S ratio in the CDR regions are found to be extrememely low [37]. In the synovium, RF B cells produce RFs with multiple replacement mutations in their variable regions, and many undergo class switching and affinity maturation. This indicates an antigen and T cell driven process [38,39]. CD40L expressing, activated T cells [31] can, upon binding with CD40 on B cells, induce proliferation, antibody production and isotype switch. CD40L is upregulated on T cells after activation by DCs, and this suggests that recently primed T cells are recruited for B cell help and affinity maturation, in a similar mechanism as occurs in the lymph node. The crucial role of CD40 signaling for RF production has been demonstrated in RF transgenic mice that lack mature T and B cells but have rearranged RF B cells. Anti-CD40 antibody could stimulate RF production as efficiently as T cells in these mice. Deletion of RF B cells by human IgG could also be blocked using anti-CD40 antibody [40*]. The importance of CD40 signaling in autoantibody production has also been shown by studies in lupus-prone mice where IgG autoantibodies are reduced after blocking CD40 signaling [41]. Synoviocytes can, in addition, support terminal B cell survival and differentiation, even though the involvement of CD40 in these interactions is not clear [42]. T cell signaling through CD40/CD40L appears to be a major factor in the inhibition of apoptosis of high affinity RF B cells, as well as in the production of RF.
Monocytes as APCs
Most synovial macrophages are located in the lining layer, where they are thought to be major contributors to cytokine production and joint destruction. The synovial macrophages are in an activated stage and have an increased expression of HLA-DR, which enhances their antigen presenting capacity [43]. Synovial macrophages, like DCs and B cells, express the costimulatory molecule CD86 [44]. Synovial macrophages have also been shown to express STAT 4, which further demonstrates their activated stage and also indicates a role of these cells in IL-12 secretion and activation/polarization of Th1 cells [45]. IFNγ is known to be a potent activator of macrophages, but the level of IFNγ in the synovium is very low [46]. It has therefore been argued that granulocyte macrophage-colony stimulating factor, which performs many similar roles to IFNγ, may play a more important role in the activation of macrophages. Synovial T cells have also been shown to activate macrophages upon cell-cell interaction where CD40L interaction is important to induce cytotoxicity, HLA-DR expression and cytokine production. Synovial monocytes are shown to be more efficient APCs than their counterparts isolated from peripheral blood [47]. Even so, macrophages may not be important in antigen presentation, as they are mainly located in the synovial lining that is distant from the T cell rich area. However, some macrophages are located in the perivascular area, and these might have specialized APC functions compared with the macrophages in the lining layer, as the former express nuclear RelB and CD68 [48].
Conclusion
There is considerable evidence indicating an ongoing aberrant immune response in the synovial tissues of RA patients. This is critically dependent on the activation of T cells, and much of the resulting tissue destruction is a consequence of the resulting pro-inflammatory cytokine production. The clinical importance of these cytokines has been demonstrated by anti-TNFα therapies in rheumatoid arthritis patients, which have been shown to be powerful treatments for the majority of RA patients, although a substantial minority does not respond [49]. An alternative approach to treatment could be to reduce T cell activation by the blocking of upstream events, particularly the costimulatory signaling pathways involved during APC–T cell interaction.
References
Weyand CM, Goronzy JJ: Pathogenesis of rheumatoid arthritis. Med Clin North Am. 1997, 81: 29-55.
Klimiuk PA, Yang H, Goronzy JJ, Weyand CM: Production of cytokines and metalloproteinases in rheumatoid synovitis is T cell dependent. Clin Immunol. 1999, 90: 65-78. 10.1006/clim.1998.4618.
Nepom GT, Hansen JA, Nepom BS: The molecular basis for HLA class II associations with rheumatoid arthritis. J Clin Immunol. 1987, 7: 1-7.
Gao XJ, Olsen NJ, Pincus T, Stastny P: HLA-DR alleles with naturally occurring amino acid substitutions and risk for development of rheumatoid arthritis. Arthritis Rheum. 1990, 33: 939-946.
Waalen K, Forre O, Natvig JB: Dendritic cells in rheumatoid inflammation. Springer Semin Immunopathol. 1988, 10: 141-156.
Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G: Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T–T help via APC activation. J Exp Med. 1996, 184: 747-752.
Li JM, Isler P, Dayer JM, Burger D: Contact-dependent stimulation of monocytic cells and neutrophils by stimulated human T-cell clones. Immunology. 1995, 84: 571-576.
Isler P, Vey E, Zhang JH, Dayer JM: Cell surface glycoproteins expressed on activated human T cells induce production of interleukin-1 beta by monocytic cells: a possible role of CD69. Eur Cytokine Network. 1993, 4: 15-23.
Larsen CP, Elwood ET, Alexander DZ, Ritchie SC, Hendrix R, Tucker-Burden C, Cho HR, Aruffo A, Hollenbaugh D, Linsley PS, Winn KJ, Pearson TC: Long-term acceptance of skin and cardiac allografts after blocking CD40 and CD28 pathways. Nature. 1996, 381: 434-438.
Janeway CAJ, Bottomly K: Signals and signs for lymphocyte responses. Cell. 1994, 76: 275-285.
Lenschow DJ, Walunas TL, Bluestone JA: CD28/B7 system of T cell costimulation. Annu Rev Immunol. 1996, 14: 233-258.
Thomas R, Quinn C: Functional differentiation of dendritic cells in rheumatoid arthritis: role of CD86 in the synovium. J Immunol. 1996, 156: 3074-3086.
Ranheim EA, Kipps TJ: Elevated expression of CD80 (B7/BB1) and other accessory molecules on synovial fluid mononuclear cell subsets in rheumatoid arthritis. Arthritis Rheum. 1994, 37: 1637-1646.
Kuchroo VK, Das MP, Brown JA, Ranger AM, Zamvil SS, Sobel RA, Weiner HL, Nabavi N, Glimcher LH: B7-1 and B7-2 costimulatory molecules activate differentially the Th1/Th2 developmental pathways: application to autoimmune disease therapy. Cell. 1995, 80: 707-718.
Gause WC, Halvorson MJ, Lu P, Greenwald R, Linsley P, Urban JF, Finkelman FD: The function of costimulatory molecules and the development of IL-4-producing T cells. Immunol Today. 1997, 18: 115-120.
Aarvak T, Chabaud M, Miossec P, Natvig JB: IL-17 is produced by some proinflammatory Th1/Th0 cells but not by Th2 cells. J Immunol. 1999, 162: 1246-1251.
Chabaud M, Fossiez F, Taupin JL, Miossec P: Enhancing effect of IL-17 on IL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritis synoviocytes and its regulation by Th2 cytokines. J Immunol. 1998, 161: 409-414.
Quayle AJ, Chomarat P, Miossec P, Kjeldsen-Kragh J, Forre O, Natvig JB: Rheumatoid inflammatory T-cell clones express mostly Th1 but also Th2 and mixed (Th0-like) cytokine patterns. Scand J Immunol. 1993, 38: 75-82.
Cross AH, Girard TJ, Giacoletto KS, Evans RJ, Keeling RM, Lin RF, Trotter JL, Karr RW: Long-term inhibition of murine experimental autoimmune encephalomyelitis using CTLA-4-Fc supports a key role for CD28 costimulation. J Clin Invest. 1995, 95: 2783-2789.
Webb LM, Walmsley MJ, Feldmann M: Prevention and amelioration of collagen-induced arthritis by blockade of the CD28 co-stimulatory pathway: requirement for both B7-1 and B7-2. Eur J Immunol. 1996, 26: 2320-2328.
Tada Y, Nagasawa K, Ho A, Morito F, Koarada S, Ushiyama O, Suzuki N, Ohta A, Mak TW: Role of the costimulatory molecule CD28 in the development of lupus in MRL/lpr mice. J Immunol. 1999, 163: 3153-3159.
Martens PB, Goronzy JJ, Schaid D, Weyand CM: Expansion of unusual CD4+ T cells in severe rheumatoid arthritis. Arthritis Rheum. 1997, 40: 1106-1114.
London CA, Lodge MP, Abbas AK: Functional responses and costimulator dependence of memory CD4+ T cells. J Immunol. 2000, 164: 265-272.
Croft M, Bradley LM, Swain SL: Naive versus memory CD4 T cell response to antigen. Memory cells are less dependent on accessory cell costimulation and can respond to many antigen-presenting cell types including resting B cells. J Immunol. 1994, 152: 2675-2685.
Grewal IS, Foellmer HG, Grewal KD, Xu J, Hardardottir F, Baron JL, Janeway CAJ, Flavell RA: Requirement for CD40 ligand in costimulation induction, T cell activation, and experimental allergic encephalomyelitis. Science. 1996, 273: 1864-1867.
Howland KC, Ausubel LJ, London CA, Abbas AK: The roles of CD28 and CD40 ligand in T cell activation and tolerance. J Immunol. 2000, 164: 4465-4470.
Pettit AR, MacDonald KP, O'Sullivan B, Thomas R: Differentiated dendritic cells expressing nuclear RelB are predominantly located in rheumatoid synovial tissue perivascular mononuclear cell aggregates. Arthritis Rheum. 2000, 43: 791-800.
Robinson E, Keystone EC, Schall TJ, Gillett N, Fish EN: Chemokine expression in rheumatoid arthritis (RA): evidence of RANTES and macrophage inflammatory protein (MIP)-1 beta production by synovial T cells. Clin Exp Immunol. 1997, 101: 398-407.
Kelsall BL, Stuber E, Neurath M, Strober W: Interleukin-12 production by dendritic cells. The role of CD40–CD40L interactions in Th1 T-cell responses. Ann NY Acad Sci. 1996, 795: 116-126.
Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G: Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T–T help via APC activation. J Exp Med. 1996, 184: 747-752.
MacDonald KP, Nishioka Y, Lipsky PE, Thomas R: Functional CD40 ligand is expressed by T cells in rheumatoid arthritis. J Clin Invest. 1997, 100: 2404-2414.
Aarvak T, Chabaud M, Källberg E, Miossec P, Natvig JB: Change in the Th1/Th2 phenotype of memory T cell clones from RA synovium. Scand J Immunol. 1999, 50: 1-9. 10.1046/j.1365-3083.1999.00581.x.
Roosnek E, Lanzavecchia A: Efficient and selective presentation of antigen-antibody complexes by rheumatoid factor B cells. J Exp Med. 1991, 173: 487-489.
Mageed RA, Borretzen M, Moyes SP, Thompson KM, Natvig JB: Rheumatoid factor autoantibodies in health and disease. Ann NY Acad Sci. 1997, 815: 296-311.
Wang H, Shlomchik MJ: High affinity rheumatoid factor transgenic B cells are eliminated in normal mice. J Immunol. 1997, 159: 1125-1134.
Goodnow CC, Crosbie J, Jorgensen H, Brink RA, Basten A: Induction of self-tolerance in mature peripheral B lymphocytes. Nature. 1989, 342: 385-391.
Borretzen M, Randen I, Zdarsky E, Forre O, Natvig JB, Thompson KM: Control of autoantibody affinity by selection against amino acid replacements in the complementarity-determining regions. Proc Natl Acad Sci U S A. 1994, 91: 12917-12921.
Borretzen M, Chapman C, Natvig JB, Thompson KM: Differences in mutational patterns between rheumatoid factors in health and disease are related to variable heavy chain family and germ-line gene usage. Eur J Immunol. 1997, 27: 735-741.
Randen I, Brown D, Thompson KM, Hughes-Jones N, Pascual V, Victor K, Capra JD, Forre O, Natvig JB: Clonally related IgM rheumatoid factors undergo affinity maturation in the rheumatoid synovial tissue. J Immunol. 1992, 148: 3296-3301.
Kyburz D, Corr M, Brinson DC, Von Damm A, Tighe H, Carson DA: Human rheumatoid factor production is dependent on CD40 signaling and autoantigen. J Immunol. 2000, 163: 3116-3122.
Mohan C, Shi Y, Laman JD, Datta SK: Interaction between CD40 and its ligand gp39 in the development of murine lupus nephritis. J Immunol. 1995, 154: 1470-1480.
Dechanet J, Merville P, Durand I, Banchereau J, Miossec P: The ability of synoviocytes to support terminal differentiation of activated B cells may explain plasma cell accumulation in rheumatoid synovium. J Clin Invest. 1995, 95: 456-463.
Ridley MG, Kingsley G, Pitzalis C, Panayi GS: Monocyte activation in rheumatoid arthritis: evidence for in situ activation and differentiation in joints. Br J Rheumatol. 1990, 29: 84-88.
Liu MF, Kohsaka H, Sakurai H, Azuma M, Okumura K, Saito I, Miyasaka N: The presence of costimulatory molecules CD86 and CD28 in rheumatoid arthritis synovium. Arthritis Rheum. 1996, 39: 110-114.
Frucht DM, Aringer M, Galon J, Danning C, Brown M, Fan S, Centola M, Wu CY, Yamada N, El Gabalawy H, O'Shea JJ: Stat4 is expressed in activated peripheral blood monocytes, dendritic cells, and macrophages at sites of Th1-mediated inflammation. J Immunol. 2000, 164: 4659-4664.
Miossec P: Pro- and antiinflammatory cytokine balance in rheumatoid arthritis. Clin Exp Rheumatol. 1995, 12 (suppl): S13-S16.
Thomas R, Davis LS, Lipsky PE: Rheumatoid synovium is enriched in mature antigen-presenting dendritic cells. J Immunol. 1994, 152: 2613-2623.
Pettit AR, MacDonald KP, O'Sullivan B, Thomas R: Differentiated dendritic cells expressing nuclear RelB are predominantly located in rheumatoid synovial tissue perivascular mononuclear cell aggregates. Arthritis Rheum. 2000, 43: 791-800.
Charles P, Elliott MJ, Davis D, Potter A, Kalden JR, Antoni C, Breedveld FC, Smolen JS, Eberl G, deWoody K, Feldmann M, Maini RN: Regulation of cytokines, cytokine inhibitors, and acute-phase proteins following anti-TNF-alpha therapy in rheumatoid arthritis. J Immunol. 1999, 163: 1521-1528.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Aarvak, T., Natvig, J.B. Cell-cell interactions in synovitis: Antigen presenting cells and T cell interaction in rheumatoid arthritis. Arthritis Res Ther 3, 13 (2000). https://doi.org/10.1186/ar135
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/ar135