
Abstract
Histaminergic afferent system of the cerebellum, having been considered as an essential component of the direct hypothalamocerebellar circuits, originates from the tuberomammillary nucleus in the hypothalamus. Unlike the mossy fibers and climbing fibers, the histaminergic afferent fibers, a third type of cerebellar afferents, extend fine varicose fibers throughout the cerebellar cortex and nuclei. Histamine receptors, belonging to the family of G protein-coupled receptors, are widely present in the cerebellum. Through these histamine receptors, histamine directly excites Purkinje cells and granule cells in the cerebellar cortex, as well as the cerebellar nuclear neurons. Therefore, the histaminergic afferents parallelly modulate these dominant components in the cerebellar circuitry and consequently influence the final output of the cerebellum. In this way, the histaminergic afferent system actively participates in the cerebellum-mediated motor balance and coordination and nonsomatic functions. Accordingly, histaminergic reagents may become potential drugs for clinical treatment of cerebellar ataxia and other cerebellar disease. On the other hand, considering the hypothalamus is a high regulatory center for autonomic and visceral activities, the hypothalamocerebellar histaminergic fibers/projections, bridging the nonsomatic center to somatic structure, may play a critical role in the somatic-nonsomatic integration.
Similar content being viewed by others


Introduction
The cerebellum is a well-known important subcortical motor structure, ensuring coordination, precision, and accurate timing of movement, and learning motor skills [1–4]. The cerebellar neuronal circuitry, organized elaborately and modularly, receives two major types of afferent inputs, mossy fibers and climbing fibers [4, 5]. The former originates from nuclei in the spinal cord and brainstem and carries sensory information from the periphery as well as information from the cerebral cortex, while the latter originates from the inferior olivary nucleus and sends error signals sensed from the motor performance of periphery musculatures to the cerebellum. In addition to obtaining specific and discrete information from the mossy and climbing fiber afferent systems, the cerebellum also receives nonspecific signals from the so-called third type of afferents, typically beaded fibers [6], which contain various amines or neuropeptides. Although more than 20 different types of amines and neuropeptides, such as serotonin [7], norepinephrine [8], histamine [9], orexin [10], and CRF [11, 12], have been found in the cerebellum, their functional significance is largely unknown. In general, beaded fibers form varicose contact with Purkinje cells and interneurons in the cerebellar cortex, as well as neurons in the cerebellar nuclei, fastigial (FN), interpositus (IN) and dentate (DN) nuclei, and exert a widespread modulatory role in the cerebellar circuitry [2, 6, 13].
Monoamines are firstly identified neurotransmitters used in the third type of afferents in the cerebellum. Among them, histamine is a newly found one in the cerebellar afferents. Although histamine was isolated from peripheral tissues as a biologically active amine more than a century ago, histamine acting as a neurotransmitter in the brain and the central histaminergic system gained general acceptance only in recent 30 years [14]. Peripheral histamine is well known to be stored primarily in the tissue mast cells and enterochromaffin-like cells, and holds a pivotal position in allergic reaction, gastric acid secretion and contraction of smooth muscle tissues of the lungs, whereas central histamine tends to be considered as a “modulator for whole brain activity” [14–17]. In the cerebellum, different from serotoninergic and norepinephrinergic afferents arising from the brainstem [6, 13], histaminergic fibers originate from the hypothalamus, a higher center for nonsomatic visceral and autonomic regulation [15, 16]. In 1984, the direct hypothalamocerebellar projections were first definitively presented by Dietrichs [18] in his pioneering study on cats. A subsequent series of neuroanatomical investigations from Haines, Dietrichs, and other colleagues [19, 20] on various mammals and nonmammalian vertebrates further substantiated the direct bidirectional connections between the cerebellum and the hypothalamus, the cerebellar-hypothalamic circuits. Since the cerebellar-hypothalamic circuits extensively exist and appear to be stronger in species ascending the phylogenetic scale, the connections may be phylogenetically old pathways [19]. The neurotransmitters in the hypothalamocerebellar projections have not been well known so far, however, a growing body of data has provided strong evidence that histamine is a potential candidate and plays an important functional role in modulating activity of the cerebellar circuitry. In this review, the structure and function of hypothalamic histaminergic projections in the cerebellum are summarized and discussed.
Review
Origination of histaminergic afferents in the cerebellum
In the cerebellar-hypothalamic circuits, the direct hypothalamocerebellar projections arise from widespread nuclei/regions in the hypothalamus, including the lateral, posterior, and dorsal hypothalamic areas, the dorsomedial and ventromedial nuclei, the periventricular zone/nucleus, the lateral mammillary and supramammillary nuclei, as well as the tuberomammillary (TMN) nucleus [17, 19]. Using an immunofluorescence technique, Ericson et al. [21] demonstrated Fast Blue-labeled l-histidine containing neurons in the TMN after cerebellar injections. In fact, series of studies have ascertained that the TMN is not only the origination of hypothalamocerebellar histaminergic afferents (Figure 1), but also the specific sole region of origin for the whole central histaminergic system in the brain [14, 16].

Hypothalamic histaminergic afferents in the cerebellum. Cerebellar histaminergic afferent fibers originate from the tuberomammillary nucleus in the hypothalamus and project to both of the cerebellar cortex and nuclei. They parallelly modulate the Purkinje cells, granule cells and nuclear neurons via H2 and/or H1 receptors and sequentially influence the outputs of the cerebellum. CF, climbing fiber; CN, cerebellar nuclei; GC, granule cell; H1, histamine H1 receptor; H2, histamine H2 receptor; MF, mossy fiber; PC, Purkinje cell; PF, parallel fiber; TMN, tuberomammillary nucleus.
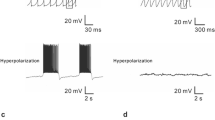
The TMN is a small nucleus located in the posterior hypothalamus. The histaminergic neurons in the TMN mostly have large somata (20–30 μm diameters) with resting potential of about -50 mV. These neurons are spontaneously active with slow regular firing rate at 1–4 Hz and mean mid-amplitude duration of action potential at 1–3 ms [22]. Although hyperpolarization-activated cyclic nucleotide-gated (HCN) channels are expressed in histaminergic neurons, they are not responsible for maintaining the neuronal spontaneous activity as a pacemaker. A complex mechanism involving Na+, K+ and Ca2+ conductances contributes to the pacemaker properties [14–16]. Importantly, the firing rate and pattern of histaminergic neurons varies in different behavioral states, with a ratio of 1.5 between firing rates of histaminergic neurons in active and quiet waking in the cat [23, 24], suggesting the central histaminergic system is closely related to not only wakefulness but also movement.
Innervation of histaminergic afferents in the cerebellum
By means of immunocytochemistry using anti-histidine decarboxylase (HDC, the enzyme catalyzing the reaction that produces histamine) antibody or antiserum against histamine, the detailed distribution of histaminergic fibers in the cerebellum has been successively examined in the guinea pig, rat, tree shrew, and human [9, 25–27]. In the rat cerebellum, HDC-immunoreactive fibers are scattered in all three cerebellar cortical layers, the molecular, Purkinje, and granular layers, rather than concentrated in any specific region [25]. However, other studies did not find any histaminergic afferents in rat cerebellum [28] or very low density in the cerebellar cortex [29]. Similar to those in the rat, the histaminergic fibers are sparsely distributed in all cortical layers in the guinea pig cerebellum, with more denser fiber networks in the vermis and flocculus and less fiber density in the cerebellar nuclei [26]. However, more histamine-immunoreactive fibers innervate cerebellar nuclei in the tree shrew [27]. In human cerebellar samples, a moderate density of histaminergic afferents has also been observed in the molecular layer, and more fibers have been seen in the granular cell layer. Additionally, these fibers run parallelly to the Purkinje cell layer after traversing it perpendicularly [9].The histaminergic fibers share many morphological similarities, including distribution, orientation, branching patterns, and ending sites, with the serotoninergic, noradrenergic and neuropeptidergic axons in the cerebellar cortex (Figure 1). On the basis of these structural properties, the histaminergic afferent fibers in the cerebellar cortex are considered to be classified as multilayered fibers. Furthermore, the most endings of histaminergic fibers do not make typical synaptic specializations but form varicosities. The varicose rather than synaptic contact pattern, together with the dispersive innervation of hypothalamic histaminergic afferents in the cerebellar cortex and nuclei, indicates an extensively modulatory role of histamine in the cerebellar circuitry.
In the TMN neurons, histamine is synthesized from l-histidine through oxidative decarboxylation by HDC. Then, histamine is stored in neuronal somata and especially in axon varicosities, where it is carried into vesicles through the vesicular monoamine transporter VMAT-2 and released in a calcium-dependent manner upon arrival of action potentials [14]. In the targets, histamine is inactivated through transfer of the methyl group from S-adenosylmethionine by histamine N-methyltransferase (HMT) or via oxidative deamination by diamine oxidase (DAO). However, HMT rather than DAO terminates histaminergic transmission in the cerebellum, since only HMT is expressed in the cerebellum [30]. Inhibition of histamine methyltransferase enhance phosphoinositide turnover in the cerebellum [30], which is mediated by histamine H1 receptors.
Expression and distribution of histamine receptors in the cerebellum
Up to date, four histamine receptors, H1-H4 receptors, have been cloned and identified, in which H1, H2 and H3 receptors are richly expressed in the central nervous system [14–16]. Although histamine H4 receptors are detected predominantly in the periphery, recent studies have also reported a functional expression of H4 receptors in human and rodent brain [31–33]. All histamine receptors are metabotropic and belong to the rhodopsin-like family of G protein-coupled receptors [14–16]. Among them, histamine H1, H2 and H4 receptors are postsynaptic and mediate mostly excitatory responses, whereas H3 receptors mediate presynaptic inhibitory actions as auto- or hetero-receptors [14–16]. Owing to autoradiographic mapping, in situ hybridization and immunohistochemistry techniques, expression and distribution of histamine receptors in the cerebellum have been revealed. Accumulating evidence demonstrates that all histamine H1, H2, H3 and H4 receptors exist in the cerebellum with various species difference.
H1 receptor
In situ hybridization studies have revealed that histamine H1 receptor mRNAs are expressed in granular layer and Purkinje cells of the guinea pig cerebellum [34, 35]. Using [3H]mepyramine or [125I]iodobolpyramine as sensitive probe, autoradiographic mapping results have showed a high density of H1 receptors in the molecular layer of the guinea pig cerebellum [36, 37]. Substantial levels of H1 receptors have also been observed in the cerebellum of cats and mice [38, 39]. However, compared with the guinea pig, mouse and cat cerebellum, much lower level of H1 receptors are expressed in rat cerebellum [40].
H2 receptor
Using [125I]iodoaminopotentidine for radioligand binding and a 33P-labelled complementary RNA probe for in situ hybridization, an autoradiographic study have demonstrated that histamine H2 receptor and its mRNAs distribute in the guinea pig cerebellum, especially in Purkinje cell and granular layers [41]. Nevertheless, in the rat brain, only low level of H2 receptor mRNA expression has been detected in the cerebellum by northern blot hybridization [42]. Interestingly, in the mouse cerebellum, from developmental point of view, H2 receptor mRNA levels present an increased tendency with age [43]. The expression and location of H2 receptors have also been observed in the dentate nucleus of human and monkey cerebellum [44].
H3 receptor
Histamine H3 receptor, located on the somata and axon terminals of histaminergic neurons, was identified as a presynaptic autoreceptor in the rat brain by Arrang et al. in 1983 [45]. Besides acting as a presynaptic autoreceptor to modulate histamine synthesis and release, H3 receptor can also exert as a presynaptic heteroreceptor to inhibit the release of various other neurotransmitters [46], such as noradrenaline, acetylcholine, glutamate and GABA. The expression and distribution of H3 receptors in the cerebellum were observed in rodents, pigs and humans [47–49]. In rats, using a 33P-labelled riboprobe for in situ hybridization, a strong mRNA expression of H3 receptor, probably the shorter isoform [50], was found in most Purkinje cells as well as in the cerebellar nuclei, including the FNs and INs [48]. But there was scarce or very low detectable binding of H3 receptors in the Purkinje cells indicated by r-[3H]α-methylhistamine or [125I]iodoproxyfan for autoradiography [48, 49], suggesting H3 receptors are expressed on efferent projections rather than somata or dendrites of the Purkinje cells in rats. Furthermore, immunohistochemical analysis using affinity-enhanced anti-H3 (349–358) antibodies demonstrated that high levels of H3 receptors were detected in Purkinje cell layer but low levels in granule layer of the mouse cerebellum [47], whereas high mRNA expression of the receptors was observed in the guinea pig [51]. By PET, low binding of H3 receptors with [11C]GSK189254 radioligand was also detected in human and pig cerebellum [52, 53]. These observations indicate that H3 receptor expression in the cerebellum varies among species.
H4 receptor
H4 receptor, the newly identified histamine receptor, is expressed predominantly in peripheral tissues and cells, such as blood, lung, gut and liver [14, 54]. However, the expression and localization of H4 receptor in the brain remain controversial in different reports [32, 55, 56]. By using RT-PCR technique, Nakamura et al. reported that expression of H4 receptor mRNAs was not detected in the brain [55]. While, RT-PCR results from other laboratories demonstrated an expression and distribution of H4 receptor mRNAs in various brain regions, including high level expression in rat cerebellum [32] and mouse cerebellar granule layer [31], and low level in human cerebellum [56]. The exact expression and distribution of H4 receptors in the cerebellum still needs to be further studied.
Histaminergic modulation on cerebellar neuronal activities
Innervation of hypothalamic histaminergic afferents on cerebellar cortex and nuclei and expression of histamine receptors in cerebellar neurons strongly suggest that histaminergic afferents may hold a key functional position in the cerebellar neuronal circuitry. In fact, a growing body of data has provided substantial evidence that histamine excites cerebellar neurons [57–63]. Although the distribution of histaminergic afferents in the rat cerebellar cortex seem to be scattered or low, electrophysiological studies show substantial evidence that histamine increases neuronal activities in cerebellar cortical circuit in rats. In 1999, Li et al. first reported that histamine induced an excitation on rat cerebellar granule cells [57], the interneurons relaying mossy fiber inputs via parallel fibers to Purkinje cells. In addition, histamine was found to excite Purkinje cells [63], the principle neurons in cerebellar cortical circuit, as well as neurons in the cerebellar nuclei, including the FN [58, 60], IN [59] and DN [62]. Interestingly, the effects of histamine on these cerebellar neurons are uniform postsynaptic excitation with various underlying receptor mechanisms (Figure 1). H2 receptors mediate the histamine-induced excitation on Purkinje cells and cerebellar nuclear neurons in rats [58–60, 62, 63], whereas H1 and H2 receptors co-mediate the excitatory effect of histamine on granule cells with a predominant contribution of H1 receptors [57]. Activation of H1 receptors in guinea pig cerebellum was also found to increase intracellular Ca2+ concentration in Purkinje cells [64]. Although H3 and H4 receptors are expressed in the cerebellum, role of them in histaminergic modulation on cerebellar neurons remains largely unclear up to date. It is only reported that H3 receptors inhibit and H2 receptors facilitate noradrenaline release in the cerebellum in guinea pigs [65].
It has been well known that histamine H1 receptor is coupled to Gq/11 protein and phospholipase C (PLC), whereas Gs and protein kinase A underlies H2 receptor [14–16]. Following H1 receptor activation in neurons in other brain areas, leak potassium channels are blocked, or Ca2+-activated cation channels and/or Na+-Ca2+ exchangers are activated [14–16]. On the other hand, activation of H2 receptors in dorsal lateral geniculate relay neurons and hippocampal pyramidal cells enhances the hyperpolarization-activated cation current (Ih) and/or inhibits a calcium-activated potassium conductance [14–16]. The whole downstream signal transduction pathways of histamine receptors in different cerebellar cortical and nuclear neurons and the underlying ionic mechanisms have not yet been revealed.
On the other hand, histamine may influence cerebellar neuronal activity through its actions on the cerebellar glial cells. It is reported that H1, H2 and H3 receptors are all expressed in the cerebellar astrocytes [66, 67], including Bergmann glial cells [68]. And histamine elevates several biochemical properties of astrocytes in the cerebellum, such as the activities of ornithine decarboxylase and glutamine synthetase, and incorporation of [3H]thymidine into DNA, and thus regulates growth and development of astrocytes [69]. Moreover, by using fura-2-based Ca2+ imaging, histamine was found to induce calcium entry in rat cerebellar astrocytes [70].
Intriguingly, besides cerebellar neurons, histamine also excites cerebellar target structures, in which vestibular nuclear complex in the brainstem plays a critical role in control of muscle tone and posture [71, 72]. The vestibular nuclear complex comprises four main nuclei, lateral (LVN), medial (MVN), inferior (IVN), and superior (SVN) vestibular nucleus. All of these four nuclei receive direct hypothalamic histaminergic innervations [73–75] and express histamine receptors [41, 48, 76, 77]. In consistent with the effect of histamine on cerebellar neurons, histamine induces an excitatory response of the neurons in vestibular nuclei. Extracellular recordings and whole-cell patch-clamp recordings in vitro showed that histamine directly excited MVN, SVN, and IVN neurons via postsynaptic H1 and H2 receptors [78–80] and depolarized LVN neurons through H2 receptors [81]. Na+-Ca2+ exchangers coupled to H1 receptors and HCN channels linked to H2 receptors contribute to the histamine-induced depolarization on MVN neurons [81, 82]. Presynaptic H3 receptor also holds a key position in vestibular nuclear circuit [83, 84] and even in vestibular compensation [83–85], however, its role in modulation of vestibular nuclear neuronal activity has not been reported.
It is noteworthy that the actions of histamine on cerebellar cortical and nuclear neurons as well as vestibular nuclear neurons are homogeneous excitation. Thus, the hypothalamic histaminergic afferent system acts to uniformly and parallelly excite components in the cerebellar circuitry as well as the cerebellar target structure, vestibular nuclear complex. Due to histaminergic varicose endings and histamine metabotropic receptors, the hypothalamic histaminergic afferent system may not transmit fast signals, but act as a biasing force to influence electrophysiological properties of cerebellar and vestibular neurons and hold their excitability and sensitivity at an appropriate level for responding to inputs coding changes in internal and external environments. In this way, the histaminergic afferent inputs may extensively modulate the sensorimotor integration in the cerebellar and vestibular circuits and sequentially influence cerebellar-related motor behaviors.
Physiological function of histaminergic afferents in the cerebellar-related behaviors
The central histaminergic nervous system has been implicated in many nonsomatic basic physiological functions, such as sleep-waking cycle, energy and endocrine homeostasis, synaptic plasticity, and learning [14–16]. Recently, role of histamine and histaminergic system in somatic motor control receives increasing attention. Intraventricular administration of histamine produced a biphasic effect in spontaneous locomotor activity with an initial transient hypoactivity followed by hyperactivity [86, 87]. Depletion of brain histamine or knockout of histamine receptors influenced motor behaviors [88–90]. The activity levels, such as wheel-running and spontaneous locomotion, in the HDC knock-out mice were lower than those in the wild types [91]. Knockout of H1 receptors in mice altered ambulatory activity and reduced exploratory behavior [89]. The H3 receptor-deficient mice showed a decrease in overall locomotion, wheel-running behavior, and stereotypic responses [90]. Interestingly, bilateral microinjection of histamine into the cerebellar FNs or INs, two final output nuclei of the spinocerebellum, does not influence overground locomotion in rats in an open field [58, 61]. However, microinjection of histamine into the FNs and INs significantly lengthens the endurance time of rats on an accelerating rota-rod (Figure 2) and shortens the time that rats spend traversing a balance beam, which is mediated by H2 receptors [58, 61], indicating a promotion of histamine on cerebellum-mediated motor balance and motor coordination. Furthermore, microinjection of histamine into bilateral FNs narrowed stride width of footprint but did not influence wire suspension, whereas microinjection of histamine into bilateral INs increased stride length and promoted suspension [58] (Figure 3), suggesting that cerebellar histaminergic afferent system may precisely modulate trunk, proximal and distal muscles via biasing the FN and IN.

Histamine promotes motor balance and motor coordination in accelerating rota-rod via H2 receptors in the cerebellar interpositus nuclei. (A) Motor performances of rats microinjected with normal saline, GABA, histamine, ranitidine (antagonist for H2 receptor) and triprolidine (antagonist for H1 receptor) in accelerating rota-rod. (B) Reversal effect of histamine on ranitidine-injected rats. *P < 0.05; **P < 0.01. Modified from Song et al., Neuroscience, 140:33–43, 2006.

Histamine precisely modulates trunk, proximal and distal muscles through the cerebellar fastigial and interpositus nuclei, respectively. (A) Walking track of hindfeet of a normal rat. (B) Microinjection of histamine into the fastigial rather than interpositus nuclei induced a narrower stride width. (C) Microinjection of histamine into the interpositus but not fastigial nuclei induced a longer stride length. (D) Microinjection of histamine into the interpositus but not fastigial nuclei increased the endurance time of suspension. **P < 0.01. SL, stride length; SW, stride width. Modified from He et al., Behav Brain Res., 228:44–52, 2012.
Besides somatic motor control, cerebellum also actively participates in many basic nonsomatic regulations and even high cognitive functions [17, 92]. Interestingly, recently, histamine has been found to be involved in the cerebellar-mediated emotional memory consolidation. Microinjection of histamine into the cerebellar vermis impairs emotional memory consolidation in mice in the elevated plus-maze [93]. The impairment is mediated by H1 rather than H2 receptors [94]. However, via H2 receptors in the cerebellum, histamine enhances memory consolidation of inhibitory avoidance learning in mice [95]. These results indicate that cerebellar histaminergic afferent system may be extensively involved in cerebellar physiological functions.
Histamine and cerebellar ataxia
Cerebellar ataxia, a form of ataxia associated with lesions to the cerebellum, is a complex motor disturbance that involves the planning and execution of movements and reduces movement accuracy and coordination [96]. Cerebellar ataxia presents with symptoms of an inability to coordinate balance, gait, extremity, and eye movements [97]. Since histaminergic afferent system plays an important role in cerebellar functions, histaminergic reagents may become potential drugs for treatment of cerebellar ataxia. Betacerc (betahistidine, an antagonist for H3 receptor and a weak agonist for H1 receptor) ameliorates symptoms of static ataxy in patients with cerebellar ataxia [98]. Ciproxifan, a potent H3 receptor antagonist, enhances MK-801 (dizocilpine, a non-competitive antagonist for NMDA receptor) produced ataxia and motor impairment [99]. Cetirizine, selective H1 receptor antagonist, decreases the falling off latency from the rota-rod and potentiates the effects of ethanol-induced ataxia [100]. The reasons why betacerc and ciproxifan exert opposite effects on ataxias still needs further investigation, Betacerc is also a weak agonist for H1 receptors and different causes of ataxias may be account for it. Although these clinical and experimental results are very preliminary, they provide a new insight and indicate a possibility of using histaminergic reagents to ameliorate symptoms of cerebellar ataxia.
Conclusion
Histaminergic afferent system in the cerebellum, despite being a third type of cerebellar afferents, plays an important modulatory role in the cerebellar circuitry and actively participates in the cerebellar somatic motor and nonsomatic functions. Different from the serotoninergic and noradrenergic fibers originating from lower brainstem, histaminergic afferents in the cerebellum arise from the hypothalamus, a higher center for visceral and autonomic regulation. Thus, the hypothalamocerebellar histaminergic projections bridge nonsomatic center, the hypothalamus, to somatic structure, the cerebellum. These connections and especially the histaminergic modulations may not only endow the cerebellar circuitry with an appropriate functional state, but also form a vital part of the somatic-nonsomatic integration, which is critical for generating an integrated and coordinated behavioral response to changes in internal and external environment.
Although clinical use of histaminergic reagents in the therapy for cerebellar ataxia is still in exploration, intensive studies on function and receptor and ionic mechanisms of the histaminergic modulation on cerebellar circuitry may provide a new target for clinical treatment of cerebellar ataxia.
References
Manto M, Gruol D, Schmahmann J, Koibuchi N, Rossi F: Handbook of the Cerebellum and Cerebellar Disorders. USA: Springer; 2012.
Ito M: Cerebellar circuitry as a neuronal machine. Prog Neurobiol 2006, 78: 272–303. 10.1016/j.pneurobio.2006.02.006
Ito M: The Cerebellum: Brain for an Implicit Self. USA: FT Press; 2012.
Ito M: The Modifiable Neuronal Network of the Cerebellum. Jpn J Physiol 1984, 34: 781–792. 10.2170/jjphysiol.34.781
Bloedel JR: Cerebellar afferent systems: a review. Prog Neurobiol 1973, 2: 3–68.
Ito M: Functional roles of neuropeptides in cerebellar circuits. Neuroscience 2009, 162: 666–672. 10.1016/j.neuroscience.2009.01.019
Bishop GA, Ho RH: The distribution and origin of serotonin immunoreactivity in the rat cerebellum. Brain Res 1985, 331: 195–207. 10.1016/0006-8993(85)91545-8
Olson L, Fuxe K: On the projections from the locus coeruleus noradrealine neurons: the cerebellar innervation. Brain Res 1971, 28: 165–171. 10.1016/0006-8993(71)90533-6
Panula P, Takagi H, Inagaki N, Yamatodani A, Tohyama M, Wada H, Kotilainen E: Histamine-containing nerve-fibers innervate human cerebellum. Neurosci Lett 1993, 160: 53–56. 10.1016/0304-3940(93)90915-8
Yu L, Zhang XY, Zhang J, Zhu JN, Wang JJ: Orexins excite neurons of the rat cerebellar nucleus interpositus via orexin 2 receptors in vitro. Cerebellum 2010, 9: 88–95. 10.1007/s12311-009-0146-0
Cummings S, Sharp B, Elde R: Corticotropin-releasing factor in cerebellar afferent systems: a combined immunohistochemistry and retrograde transport study. J Neurosci 1988, 8: 543–554.
Miyata M, Okada D, Hashimoto K, Kano M, Ito M: Corticotropin-releasing factor plays a permissive role in cerebellar long-term depression. Neuron 1999, 22: 763–775. 10.1016/S0896-6273(00)80735-7
Schweighofer N, Doya K, Kuroda S: Cerebellar aminergic neuromodulation: towards a functional understanding. Brain Res Rev 2004, 44: 103–116. 10.1016/j.brainresrev.2003.10.004
Haas HL, Sergeeva OA, Selbach O: Histamine in the nervous system. Physiol Rev 2008, 88: 1183–1241. 10.1152/physrev.00043.2007
Brown RE, Stevens DR, Haas HL: The physiology of brain histamine. Prog Neurobiol 2001, 63: 637–672. 10.1016/S0301-0082(00)00039-3
Haas H, Panula P: The role of histamine and the tuberomamillary nucleus in the nervous system. Nat Rev Neurosci 2003, 4: 121–130.
Zhu JN, Yung WH, Kwok-Chong Chow B, Chan YS, Wang JJ: The cerebellar-hypothalamic circuits: potential pathways underlying cerebellar involvement in somatic-visceral integration. Brain Res Rev 2006, 52: 93–106. 10.1016/j.brainresrev.2006.01.003
Dietrichs E: Cerebellar autonomic function: direct hypothalamocerebellar pathway. Science 1984, 223: 591–593. 10.1126/science.6198719
Haines DE, Dietrichs E, Mihailoff GA, McDonald EF: The cerebellar-hypothalamic axis: basic circuits and clinical observations. Int Rev Neurobiol 1997, 41: 83–107.
Onat F, Cavdar S: Cerebellar connections: hypothalamus. Cerebellum 2003, 2: 263–269. 10.1080/14734220310016187
Ericson H, Watanabe T, Kohler C: Morphological analysis of the tuberomammillary nucleus in the Rat-brain - delineation of subgroups with antibody against L-histidine decarboxylase as a marker. J Comp Neurol 1987, 263: 1–24. 10.1002/cne.902630102
Haas HL, Reiner PB: Membrane properties of histaminergic tuberomammillary neurones of the rat hypothalamus in vitro. J Physiol 1988, 399: 633–646.
Sakai K, Takahashi K, Anaclet C, Lin JS: Sleep-waking discharge of ventral tuberomammillary neurons in wild-type and histidine decarboxylase knock-out mice. Front Behav Neurosci 2010, 4: 53.
Vanni-Mercier G, Gigout S, Debilly G, Lin JS: Waking selective neurons in the posterior hypothalamus and their response to histamine H3-receptor ligands: an electrophysiological study in freely moving cats. Behav Brain Res 2003, 144: 227–241. 10.1016/S0166-4328(03)00091-3
Inagaki N, Yamatodani A, Ando-Yamamoto M, Tohyama M, Watanabe T, Wada H: Organization of histaminergic fibers in the rat brain. J Comp Neurol 1988, 273: 283–300. 10.1002/cne.902730302
Airaksinen MS, Panula P: The histaminergic system in the guinea pig central nervous system: an immunocytochemical mapping study using an antiserum against histamine. J Comp Neurol 1988, 273: 163–186. 10.1002/cne.902730204
Airaksinen MS, Flugge G, Fuchs E, Panula P: Histaminergic system in the tree shrew brain. J Comp Neurol 1989, 286: 289–310. 10.1002/cne.902860302
Watanabe T, Taguchi Y, Shiosaka S, Tanaka J, Kubota H, Terano Y, Tohyama M, Wada H: Distribution of the histaminergic neuron system in the central nervous system of rats; a fluorescent immunohistochemical analysis with histidine decarboxylase as a marker. Brain Res 1984, 295: 13–25. 10.1016/0006-8993(84)90811-4
Panula P, Pirvola U, Auvinen S, Airaksinen MS: Histamine-immunoreactive nerve fibers in the rat brain. Neuroscience 1989, 28: 585–610. 10.1016/0306-4522(89)90007-9
Takemura M, Kitanaka N, Kitanaka J: Signal transduction by histamine in the cerebellum and its modulation by N-methyltransferase. Cerebellum 2003, 2: 39–43. 10.1080/14734220310015601
Connelly WM, Shenton FC, Lethbridge N, Leurs R, Waldvogel HJ, Faull RL, Lees G, Chazot PL: The histamine H4 receptor is functionally expressed on neurons in the mammalian CNS. Br J Pharmacol 2009, 157: 55–63. 10.1111/j.1476-5381.2009.00227.x
Strakhova MI, Nikkel AL, Manelli AM, Hsieh GC, Esbenshade TA, Brioni JD, Bitner RS: Localization of histamine H4 receptors in the central nervous system of human and rat. Brain Res 2009, 1250: 41–48.
Lethbridge NL, Chazot PL: Immunological identification of the mouse H4 histamine receptor on spinal cord motor neurons using a novel anti-mouse H4R antibody. Inflamm Res 2010,59(Suppl 2):S197-S198.
Horio Y, Mori Y, Higuchi I, Fujimoto K, Ito S, Fukui H: Molecular cloning of the guinea-pig histamine H1 receptor gene. J Biochem 1993, 114: 408–414.
Traiffort E, Leurs R, Arrang JM, Tardivel-Lacombe J, Diaz J, Schwartz JC, Ruat M: Guinea pig histamine H1 receptor. I. Gene cloning, characterization, and tissue expression revealed by in situ hybridization. J Neurochem 1994, 62: 507–518.
Palacios JM, Young WS, Kuhar MJ: Autoradiographic localization of histamine-H-1 receptors in brain using mepyramineh-3 - preliminary studies. Eur J Pharmacol 1979, 58: 295–304. 10.1016/0014-2999(79)90478-3
Korner M, Bouthenet ML, Ganellin CR, Garbarg M, Gros C, Ife RJ, Sales N, Schwartz JC: [I-125] iodobolpyramine, a highly sensitive probe for histamine H1-receptors in Guinea-Pig brain. Eur J Pharmacol 1986, 120: 151–160. 10.1016/0014-2999(86)90535-2
Rotter A, Frostholm A: Cerebellar histamine-H1 receptor distribution: an autoradiographic study of Purkinje cell degeneration, staggerer, weaver and reeler mutant mouse strains. Brain Res Bull 1986, 16: 205–214. 10.1016/0361-9230(86)90034-1
Taylor JE, Yaksh TL, Richelson E: Histamine H1 receptors in the brain and spinal cord of the cat. Brain Res 1982, 243: 391–394. 10.1016/0006-8993(82)90268-2
Hill SJ, Young JM: Histamine H1-receptors in the brain of the guinea-Pig and the Rat - differences in ligand-binding properties and regional distribution. Br J Pharmacol 1980, 68: 687–696. 10.1111/j.1476-5381.1980.tb10861.x
Vizuete ML, Traiffort E, Bouthenet ML, Ruat M, Souil E, Tardivel-Lacombe J, Schwartz JC: Detailed mapping of the histamine H2 receptor and its gene transcripts in guinea-pig brain. Neuroscience 1997, 80: 321–343. 10.1016/S0306-4522(97)00010-9
Karlstedt K, Senkas A, Ahman M, Panula P: Regional expression of the histamine H(2) receptor in adult and developing rat brain. Neuroscience 2001, 102: 201–208. 10.1016/S0306-4522(00)00464-4
Terao A, Steininger TL, Morairty SR, Kilduff TS: Age-related changes in histamine receptor mRNA levels in the mouse brain. Neurosci Lett 2004, 355: 81–84. 10.1016/j.neulet.2003.10.061
Honrubia MA, Vilaro MT, Palacios JM, Mengod G: Distribution of the histamine H(2) receptor in monkey brain and its mRNA localization in monkey and human brain. Synapse 2000, 38: 343–354. 10.1002/1098-2396(20001201)38:3<343::AID-SYN14>3.0.CO;2-M
Arrang JM, Garbarg M, Schwartz JC: Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 1983, 302: 832–837. 10.1038/302832a0
Leurs R, Hoffmann M, Wieland K, Timmerman H: H3 receptor gene is cloned at last. Trends Pharmacol Sci 2000, 21: 11–12. 10.1016/S0165-6147(99)01411-X
Chazot PL, Hann V, Wilson C, Lees G, Thompson CL: Immunological identification of the mammalian H3 histamine receptor in the mouse brain. Neuroreport 2001, 12: 259–262. 10.1097/00001756-200102120-00016
Pillot C, Heron A, Cochois V, Tardivel-Lacombe J, Ligneau X, Schwartz JC, Arrang JM: A detailed mapping of the histamine H(3) receptor and its gene transcripts in rat brain. Neuroscience 2002, 114: 173–193. 10.1016/S0306-4522(02)00135-5
Pollard H, Moreau J, Arrang JM, Schwartz JC: A detailed autoradiographic mapping of histamine H3 receptors in rat brain areas. Neuroscience 1993, 52: 169–189. 10.1016/0306-4522(93)90191-H
Drutel G, Peitsaro N, Karlstedt K, Wieland K, Smit MJ, Timmerman H, Panula P, Leurs R: Identification of rat H3 receptor isoforms with different brain expression and signaling properties. Mol Pharmacol 2001, 59: 1–8.
Tardivel-Lacombe J, Rouleau A, Heron A, Morisset S, Pillot C, Cochois V, Schwartz JC, Arrang JM: Cloning and cerebral expression of the guinea pig histamine H3 receptor: evidence for two isoforms. Neuroreport 2000, 11: 755–759. 10.1097/00001756-200003200-00020
Plisson C, Gunn RN, Cunningham VJ, Bender D, Salinas CA, Medhurst AD, Roberts JC, Laruelle M, Gee AD: 11C-GSK189254: a selective radioligand for in vivo central nervous system imaging of histamine H3 receptors by PET. J Nucl Med 2009, 50: 2064–2072. 10.2967/jnumed.109.062919
Ashworth S, Rabiner EA, Gunn RN, Plisson C, Wilson AA, Comley RA, Lai RYK, Gee AD, Laruelle M, Cunningham VJ: Evaluation of C-11-GSK189254 as a Novel Radioligand for the H-3 Receptor in Humans Using PET. J Nucl Med 2010, 51: 1021–1029. 10.2967/jnumed.109.071753
Nguyen T, Shapiro DA, George SR, Setola V, Lee DK, Cheng R, Rauser L, Lee SP, Lynch KR, Roth BL, O’Dowd BF: Discovery of a novel member of the histamine receptor family. Mol Pharmacol 2001, 59: 427–433.
Nakamura T, Itadani H, Hidaka Y, Ohta M, Tanaka K: Molecular cloning and characterization of a new human histamine receptor, HH4R. Biochem Biophys Res Commun 2000, 279: 615–620. 10.1006/bbrc.2000.4008
Coge F, Guenin SP, Rique H, Boutin JA, Galizzi JP: Structure and expression of the human histamine H4-receptor gene. Biochem Biophys Res Commun 2001, 284: 301–309. 10.1006/bbrc.2001.4976
Li WC, Tang XH, Li HZ, Wang JJ: Histamine excites rat cerebellar granule cells in vitro through H-1 and H-2 receptors. J Physiology Paris 1999, 93: 239–244. 10.1016/S0928-4257(99)80157-0
He YC, Wu GY, Li D, Tang B, Li B, Ding Y, Zhu JN, Wang JJ: Histamine promotes rat motor performances by activation of H(2) receptors in the cerebellar fastigial nucleus. Behav Brain Res 2012, 228: 44–52. 10.1016/j.bbr.2011.11.029
Shen B, Li HZ, Wang JJ: Excitatory effects of histamine on cerebellar interpositus nuclear cells of rats through H-2 receptors in vitro. Brain Res 2002, 948: 64–71. 10.1016/S0006-8993(02)02950-5
Tang B, Zhang J, Yu L, Li HZ, Zhu JN, Wang JJ: Excitation of histamine on neuronal activity of cerebellar fastigial nucleus in rat. Inflamm Res 2008, 57: S41-S42. 10.1007/s00011-007-0637-8
Song YN, Li HZ, Zhu JN, Guo CL, Wang JJ: Histamine improves rat rota-rod and balance beam performances through H(2) receptors in the cerebellar interpositus nucleus. Neuroscience 2006, 140: 33–43. 10.1016/j.neuroscience.2006.01.045
Qin YT, Ma SH, Zhuang QX, Qiu YH, Li B, Peng YP, Wang JJ: Histamine evokes excitatory response of neurons in the cerebellar dentate nucleus via H2 receptors. Neurosci Lett 2011, 502: 133–137. 10.1016/j.neulet.2011.05.241
Tian L, Wen YQ, Li HZ, Zuo CC, Wang JJ: Histamine excites rat cerebellar Purkinje cells via H2 receptors in vitro. Neurosci Res 2000, 36: 61–66. 10.1016/S0168-0102(99)00109-1
Kirischuk S, Verkhratsky A: [Ca2+](i) recordings from neural cells in acutely isolated cerebellar slices employing differential loading of the membrane permeant form of the calcium indicator fura-2. Pflug Arch Eur J Phy 1996, 431: 977–983. 10.1007/s004240050094
Timm J, Marr I, Werthwein S, Elz S, Schunack W, Schlicker E: H2 receptor-mediated facilitation and H3 receptor-mediated inhibition of noradrenaline release in the guinea-pig brain. Naunyn Schmiedeberg’s Arch Pharmacol 1998, 357: 232–239. 10.1007/PL00005162
Hosli E, Hosli L: Autoradiographic localization of binding sites for [3H]histamine and H1- and H2-antagonists on cultured neurones and glial cells. Neuroscience 1984, 13: 863–870. 10.1016/0306-4522(84)90101-5
Mele T, Juric DM: Identification and pharmacological characterization of the histamine H3 receptor in cultured rat astrocytes. Eur J Pharmacol 2013, 720: 198–204. 10.1016/j.ejphar.2013.10.028
Kirischuk S, Tuschick S, Verkhratsky A, Kettenmann H: Calcium signalling in mouse Bergmann glial cells mediated by alpha1-adrenoreceptors and H1 histamine receptors. Eur J Neurosci 1996, 8: 1198–1208. 10.1111/j.1460-9568.1996.tb01288.x
Rodriguez J, Moran J, Blanco I, Patel AJ: Effect of histamine on the development of astroglial cells in culture. Neurochem Res 1989, 14: 693–700. 10.1007/BF00964881
Jung S, Pfeiffer F, Deitmer JW: Histamine-induced calcium entry in rat cerebellar astrocytes: evidence for capacitative and non-capacitative mechanisms. J Physiol 2000,527(Pt 3):549–561.
Barmack NH: Central vestibular system: vestibular nuclei and posterior cerebellum. Brain Res Bull 2003, 60: 511–541. 10.1016/S0361-9230(03)00055-8
Straka H, Vibert N, Vidal PP, Moore LE, Dutia MB: Intrinsic membrane properties of vertebrate vestibular neurons: function, development and plasticity. Prog Neurobiol 2005, 76: 349–392. 10.1016/j.pneurobio.2005.10.002
Schwartz JC, Arrang JM, Garbarg M, Pollard H, Ruat M: Histaminergic transmission in the mammalian brain. Physiol Rev 1991, 71: 1–51.
Steinbusch HW: Distribution of histaminergic neurons and fibers in rat brain. Comparison with noradrenergic and serotonergic innervation of the vestibular system. Acta Otolaryngol Suppl 1991, 479: 12–23.
Tighilet B, Lacour M: Distribution of histaminergic axonal fibres in the vestibular nuclei of the cat. Neuroreport 1996, 7: 873–878. 10.1097/00001756-199603220-00008
Zhou L, Zhou W, Zhang S, Liu B, Leng Y, Zhou R, Kong W: Changes in histamine receptors (H1, H2, and H3) expression in Rat medial vestibular nucleus and flocculus after unilateral labyrinthectomy: histamine receptors in vestibular compensation. PLoS One 2013, 8: e66684. 10.1371/journal.pone.0066684
Palacios JM, Wamsley JK, Kuhar MJ: The distribution of histamine H1-receptors in the rat brain: an autoradiographic study. Neuroscience 1981, 6: 15–37. 10.1016/0306-4522(81)90240-2
Peng SY, Zhuang QX, He YC, Zhu JN, Wang JJ: Histamine excites neurons of the inferior vestibular nucleus in rats by activation of H1 and H2 receptors. Neurosci Lett 2013, 541: 87–92.
Zhuang QX, Wu YH, Wu GY, Zhu JN, Wang JJ: Histamine excites rat superior vestibular nuclear neurons via postsynaptic H1 and H2 receptors in vitro. Neuro Signals 2013, 21: 174–183. 10.1159/000341980
Wang JJ, Dutia MB: Effects of histamine and betahistine on rat medial vestibular nucleus neurones: possible mechanism of action of anti-histaminergic drugs in vertigo and motion sickness. Exp Brain Res 1995, 105: 18–24.
Zhang J, Han XH, Li HZ, Zhu JN, Wang JJ: Histamine excites rat lateral vestibular nuclear neurons through activation of post-synaptic H2 receptors. Neurosci Lett 2008, 448: 15–19. 10.1016/j.neulet.2008.10.027
Zhang XY, Yu L, Zhuang QX, Peng SY, Zhu JN, Wang JJ: Postsynaptic mechanisms underlying the excitatory action of histamine on medial vestibular nucleus neurons in rats. Br J Pharmacol 2013, 170: 156–169. 10.1111/bph.12256
Bergquist F, Dutia MB: Central histaminergic modulation of vestibular function - a review. Sheng Li Xue Bao 2006, 58: 293–304.
Bergquist F, Ruthven A, Ludwig M, Dutia MB: Histaminergic and glycinergic modulation of GABA release in the vestibular nuclei of normal and labyrinthectomised rats. J Physiol 2006, 577: 857–868. 10.1113/jphysiol.2006.120493
Tighilet B, Trottier S, Mourre C, Lacour M: Changes in the histaminergic system during vestibular compensation in the cat. J Physiol 2006, 573: 723–739. 10.1113/jphysiol.2006.107805
Bristow LJ, Bennett GW: Biphasic effects of intra-accumbens histamine administration on spontaneous motor-activity in the Rat - a role for central histamine-receptors. Br J Pharmacol 1988, 95: 1292–1302. 10.1111/j.1476-5381.1988.tb11767.x
Chiavegatto S, Nasello AG, Bernardi MM: Histamine and spontaneous motor activity: biphasic changes, receptors involved and participation of the striatal dopamine system. Life Sci 1998, 62: 1875–1888. 10.1016/S0024-3205(98)00154-4
Onodera K, Yamatodani A, Watanabe T, Wada H: Neuropharmacology of the histaminergic neuron system in the brain and its relationship with behavioral-disorders. Prog Neurobiol 1994, 42: 685–702. 10.1016/0301-0082(94)90017-5
Inoue I, Yanai K, Kitamura D, Taniuchi I, Kobayashi T, Niimura K, Watanabe T, Watanabe T: Impaired locomotor activity and exploratory behavior in mice lacking histamine H-1 receptors. Proc Natl Acad Sci U S A 1996, 93: 13316–13320. 10.1073/pnas.93.23.13316
Toyota H, Dugovic C, Koehl M, Laposky AD, Weber C, Ngo K, Wu Y, Lee DH, Yanai K, Sakurai E, Watanabe T, Liu C, Chen J, Barbier AJ, Turek FW, Fung-Leung WP, Lovenberg TW: Behavioral characterization of mice lacking histamine H(3) receptors. Mol Pharmacol 2002, 62: 389–397. 10.1124/mol.62.2.389
Abe H, Honma S, Ohtsu H, Honma K: Circadian rhythms in behavior and clock gene expressions in the brain of mice lacking histidine decarboxylase. Brain Res Mol Brain Res 2004, 124: 178–187. 10.1016/j.molbrainres.2004.02.015
Zhu JN, Wang JJ: The cerebellum in feeding control: possible function and mechanism. Cell Mol Neurobiol 2008, 28: 469–478. 10.1007/s10571-007-9236-z
Gianlorenco ACL, Canto-de-Souza A, Mattioli R: Microinjection of histamine into the cerebellar vermis impairs emotional memory consolidation in mice. Brain Res Bull 2011, 86: 134–138. 10.1016/j.brainresbull.2011.05.014
Gianlorenco ACL, Serafim KR, Canto-de-Souza A, Mattioli R: Emotional memory consolidation impairment induced by histamine is mediated by H-1 but not H-2 receptors. Brain Res Bull 2012, 89: 197–202. 10.1016/j.brainresbull.2012.09.003
Gianlorenco AC, Canto-de-Souza A, Mattioli R: Intra-cerebellar microinjection of histamine enhances memory consolidation of inhibitory avoidance learning in mice via H2 receptors. Neurosci Lett 2013,557(Pt B):159–164.
Ferrarin M, Gironi M, Mendozzi L, Nemni R, Mazzoleni P, Rabuffetti M: Procedure for the quantitative evaluation of motor disturbances in cerebellar ataxic patients. Med Biol Eng Comput 2005, 43: 349–356. 10.1007/BF02345812
Schmahmann JD: Disorders of the cerebellum: ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J Neuropsych Clin N 2004, 16: 367–378. 10.1176/appi.neuropsych.16.3.367
Prokopenko SV, Rudnev VA, Afanasieva EV, Abramov VG: A use of betaserc in ataxic syndromes. Zh Nevropatol Psikh 2004, 104: 41–45.
Bardgett ME, Points M, Kleier J, Blankenship M, Griffith MS: The H3 antagonist, ciproxifan, alleviates the memory impairment but enhances the motor effects of MK-801 (dizocilpine) in rats. Neuropharmacology 2010, 59: 492–502. 10.1016/j.neuropharm.2010.07.004
Mandhane SN, Shah JH, Bahekar PC, Mehetre SV, Pawar CA, Bagad AS, Chidrewar GU, Rao CT, Rajamannar T: Characterization of anti-inflammatory properties and evidence for no sedation liability for the novel antihistamine SUN-1334H. Int Arch Allergy Immunol 2010, 151: 56–69. 10.1159/000232571
Acknowledgement
Researches from our group were supported by grants 31070959, 31071021, 31171050, 31330033, and 91332124 from the National Natural Science Foundation of China; RFDP grant 20100091110016, SRFDP/RGC ERG Joint Research Scheme grant 20130091140003 and NCET Program from the State Educational Ministry of China; grant BK2011014 from the Natural Science Foundation of Jiangsu Province, China; and grant 2013 T60520 from China Postdoctoral Science Foundation.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interest
The authors declare that they have no competing interests.
Authors’ contributions
BL drafted the manuscript and figures. JNZ designed the review, conceived the figures, and drafted the manuscript. JJW gave comments and suggestions based on the area reviewed and helped to draft the manuscript. JNZ and JJW developed conclusions. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Li, B., Zhu, JN. & Wang, JJ. Histaminergic afferent system in the cerebellum: structure and function. cerebellum ataxias 1, 5 (2014). https://doi.org/10.1186/2053-8871-1-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/2053-8871-1-5