
Abstract
Nonassociative learning is a basic neuroadaptive behavior exhibited across animal phyla and sensory modalities but its role in brain intelligence is unclear. Current literature on habituation and sensitization, the classic "dual process" of nonassociative learning, gives highly incongruous accounts between varying experimental paradigms. Here we propose a general theory of nonassociative learning featuring four base modes: habituation/primary sensitization in primary stimulus-response pathways, and desensitization/secondary sensitization in secondary stimulus-response pathways. Primary and secondary modes of nonassociative learning are distinguished by corresponding activity-dependent recall, or nonassociative gating, of neurotransmission memory. From the perspective of brain computation, nonassociative learning is a form of integral-differential calculus whereas nonassociative gating is a form of Boolean logic operator – both dynamically transforming the stimulus-response relationship. From the perspective of sensory integration, nonassociative gating provides temporal filtering whereas nonassociative learning affords low-pass, high-pass or band-pass/band-stop frequency filtering – effectively creating an intelligent sensory firewall that screens all stimuli for attention and resultant internal model adaptation and reaction. This unified framework ties together many salient characteristics of nonassociative learning and nonassociative gating and suggests a common kernel that correlates with a wide variety of sensorimotor integration behaviors such as central resetting and self-organization of sensory inputs, fail-safe sensorimotor compensation, integral-differential and gated modulation of sensorimotor feedbacks, alarm reaction, novelty detection and selective attention, as well as a variety of mental and neurological disorders such as sensorimotor instability, attention deficit hyperactivity, sensory defensiveness, autism, nonassociative fear and anxiety, schizophrenia, addiction and craving, pain sensitization and phantom sensations, etc.
Similar content being viewed by others

1. Background
Brain calculus – or integral-differential neural dynamics – is an emerging paradigm in computational neuroscience [1, 2]. In behavioral neuroscience, the dynamics of sensorimotor integration are often ascribed to learning and memory. We hereby propose a general framework of nonassociative learning and nonassociative gating that demonstrate brain calculus and Boolean logic computations. The resultant neural network theory proves to illuminate a variety of behavioral and brain functions and disorders.
This article is written with a broad readership in mind. Beginning with a thorough review of the oft-conflicting literature on habituation and sensitization, the so-called "dual process" of nonassociative learning, Section 2 develops a unified framework of primary and secondary sensitization in analogy to pain sensitization. In Section 3, we introduce the notion of response desensitization [3] and show that this novel nonassociative learning mechanism provides a common kernel which may explain a variety of sensory remapping phenomena. Section 4 presents a novel behavioral paradigm called nonassociative gating which affords activity-dependent temporal filtering or Boolean logic-gating of the stimulus-response relationship. These emergent concepts cumulate in a general theoretical framework elaborated in Section 5, which expounds the computational roles of nonassociative learning as gated neural integrator and differentiator (low-pass and high-pass filter) in neural pathways. Section 6 discusses the functional roles of the various modes of nonassociative learning in brain intelligence as the building blocks of a "sensory firewall" for Cartesian mind-body internal model adaptation. Section 7 concludes the discourse.
2. Dual-process theory revisited
Although a universally agreed model of nonassociative learning is presently lacking (for reviews see [4–8]), a useful starting point is the classic dual-process theory of response habituation and sensitization [9–11]. In the following, we present a unified framework that extends and reconciles the dual-process theory and other models of nonassociative learning.
2.1. Habituation and sensitization: the 'dual-process theory'
2.1.1. Dual-process theory
According to this classic theory, an animal's behavioral response to a repetitive stimulus may wane or wax through two complementary learning processes called habituation and sensitization. At the system level these processes are thought to correspond, respectively, to short-term depression (STD) of neurotransmission in a primary stimulus-response pathway and short-term potentiation (STP) or facilitation of neurotransmission in a secondary, collateral pathway or "state system" that presumably determines the animal's general level of excitation, arousal or motivation to respond. Here, "short-term" plasticity (potentiation or depression) refers to the short-term modifiability and short-term memory commonly seen in nonassociative learning although long-term memory (> 24 hr) is also possible [12–16]. In some model systems habituation is induced by an innocuous stimulus (such as gentle touch) and sensitization is induced by noxious stimulus (forceful touch or electrical shock). In other systems habituation and sensitization could be induced by the same stimulus (e.g., startle response to repetitive loud noise).
Certain conjectures of the dual-process theory have subsequently been verified in a variety of invertebrate and mammalian brain systems [17–30]. Circumstantial evidence for dual-process learning could also be inferred, albeit unwittingly, from other animal models of nonassociative learning reported in the literature (reviewed in [31]).
2.1.2. Properties of response habituation
Typically, habituation may be induced by a stimulus that is presented continuously or intermittently with a variable interstimulus interval (ISI). The dual-process theory defined habituation by the following stimulus-response criteria [9, 10, 32]: 1) exponential development with repeated stimulus applications, causing exponential decrease of response to the stimulus; 2) spontaneous recovery with a short-term memory upon cessation of stimulus; 3) successive potentiation or accumulation with repeated training sessions; 4) dependence on stimulus frequency with rate and magnitude of habituation being directly related to frequency of stimulus bouts (and inversely related to ISI); 5) dependence on stimulus intensity with rate and magnitude of habituation being inversely related to stimulus intensity; 6) dependence on stimulus quantity with spontaneous recovery of habituation becoming much slower after an excessive number of stimulus bouts; 7) cross-modal generalization or transfer of habituation to other stimuli that share common habituating elements with the primary stimulus; 8) dishabituation or trumping of habituation by a novel stimulus; and 9) habituation of dishabituation upon repeated applications of the dishabituating stimulus.
These postulated properties of habituation have been borne out for the most part in many animal models from nematodes [33] to mammals – down to the level of a monosynaptic junction in the hippocampus [34]. It is generally assumed that response habituation is mediated primarily by homosynaptic depression in the stimulus-response pathway [32] although other mechanisms such as increased inhibition (see [5, 35]) or decreased neuronal excitability [36] are also possible.
2.2. A unified framework for response sensitization
Although the above characterizations of habituation appear to prevail across animal phyla and sensory modalities, those of response sensitization are less clear. The lack of a consistent taxonomy for sensitization has made it difficult to decipher and relate the vast amounts of pertinent (and oft-conflicting) data from diverse animal models, sparking considerable confusion and controversy in the literature. Here we review two conventional characterizations, intrinsic and extrinsic sensitization, which have occasioned renewed interests (reviewed in [31]). We then propose a unified framework that reconciles the discrepancies between these characterizations of sensitization and the dual-process theory.
2.2.1. Ambiguities of intrinsic and extrinsic sensitization
According to the dual-process theory, sensitization may be induced by repeated applications of a primary, or intrinsic, stimulus. Such "intrinsic sensitization" has been implicated in the increment phase of the rat acoustic startle response [37] and the monosynaptic ventral root reflex of the frog spinal cord [38]. It may also account for certain forms of sensitization such as the reported incremental sensitization of defensive striking in larval Manduca Sexta [39], iterative enhancement of the sea slug Tritonia swim response [40, 41], warm-up phase in the local bending reflex of the medicinal leech Hirudo medicinalis [42], the "windup" or central sensitization of mammalian pain pathways [43–46] and the progressive intensification of the evoked irritant sensation upon repeated applications of the pungent chemical capsaicin to the tongue [47].
More commonly, sensitization is characterized as an increase in the response to a primary stimulus after priming by an extrinsic, often strong and noxious stimulus. This form of sensitization has been variously referred to as "conventional" or "nociceptive" sensitization [39] or "extrinsic" sensitization [10, 27, 37, 42, 48, 49]. Its underlying mechanisms (as demonstrated in the Aplysia gill withdrawal reflex) may include short-term presynaptic or heterosynaptic facilitation of convergent pathways [50, 51] or long-term cellular changes [52, 53].
Although such an intrinsic-extrinsic classification of sensitization is useful, their distinction is not always clear-cut. Thus, a stimulus could sometimes induce both forms of sensitization simultaneously rather than one or the other exclusively. In the snail Helix aspersa tentacle withdrawal reflex, for instance, mixed intrinsic-extrinsic sensitization may be induced by a strong intrinsic stimulus when combined with inputs from the CNS [27, 49].
Another anomaly to the above classification scheme is exemplified by the whole-body shortening reflex of the medicinal leech Hirudo medicinalis [48, 54], in which a stimulus at one site of the leech body wall may sensitize the response to the same stimulus at a proximal but distinct body wall site. This phenomenon is analogous to extrinsic sensitization even though it involves only "intrinsic" stimuli and does not generalize to other loci or other sensory modalities. Similarly, in the "intrinsic sensitization" of Aplysia tail withdrawal reflex, activity of one tail sensory neuron during habituation training may heterosynaptically facilitate the response to a proximal but untrained tail sensory neuron [55]. Apart from the similarity of the sensitizing and test stimuli, however, such "intrinsic sensitization" is mechanistically analogous to "extrinsic sensitization" in Aplysia gill withdrawal reflex [51]. These seeming anomalies call for a revamping of the taxonomy for sensitization.
2.2.2. Primary and secondary sensitization
The above intrinsic-extrinsic classification of sensitization is based solely on the induction process. As pointed out by Prescott [31], it is important to distinguish the induction and expression phases of sensitization. Here we propose a unified framework that rectifies the ambiguities of intrinsic and extrinsic sensitization.
In keeping with the dual-process theory we refer to the stimulus that induces learning as the primary stimulus and the corresponding stimulus-response pathway the primary pathway. Further, any collateral pathway that is indirectly influenced (e.g., through heterosynaptic or presynaptic modulation) by the primary pathway is termed secondary pathway and the corresponding driving stimulus a secondary stimulus. Under this nomenclature, we define primary and secondary sensitization as sensitization expressed in the primary or secondary pathway, respectively, regardless of any extrinsic influences on the corresponding induction process (Fig. 1A).

Primary and secondary sensitization and input-gating effect and their correspondence to hyperalgesia and allodynia forms of pain sensitization. A. Schematic illustration of habituation (H) and secondary sensitization (S) mediated by homosynaptic STD (filled inner triangle, blue) in primary pathway and heterosynaptic STP (open inner triangle, red) in secondary pathway. Open triangles denote non-adaptive excitatory synapses. Primary sensitization could occur independently of habituation through homosynaptic STP (open inner triangle, red) in primary pathway. Other primary-secondary pathway configurations of nonassociative learning are also possible [140]. B. Input-gating effect: upon cessation of primary input, all memory components in primary pathway are gated off abruptly and become latent. C. Schematic illustration of hyperalgesia and allodynia respectively as primary and secondary sensitization at peripheral (P) or central (C) sites. The pain sensation and sensitization are relieved once the stimulus ceases – a behavior that epitomizes the input-gating effect.
This emphasis on expression instead of induction of sensitization circumvents the ambiguities in previous studies. On one hand, primary sensitization is analogous to intrinsic sensitization in that both are induced and expressed directly in the primary pathway. On the other hand primary sensitization does not exclude possible extrinsic influences as does intrinsic sensitization. Similarly, secondary sensitization is analogous to extrinsic sensitization in that both are expressed for a stimulus different than the sensitizing stimulus, but secondary sensitization is distinguished from primary sensitization by its indirect expression. Thus, secondary sensitization satisfactorily accounts for extrinsic sensitization and reconciles the seeming discrepancy with the above-mentioned anomalies [48, 54, 55].
2.2.3. Sensitization of pain: hyperalgesia and allodynia
The above definitions of primary and secondary sensitization of sensory inputs may also shed light on the problem of peripheral or central sensitization of pain pathways following physical insults. Peripheral sensitization is mediated by noxious input-dependent release of inflammatory neuromodulators which (by activating protein kinases) increase the transduction sensitivity and excitability of the nociceptor terminal. "Classical" central sensitization is mediated by activity-dependent increases in excitability or expansion in receptive fields of nociceptive relay neurons at the superficial (lamina I) or deep (lamina V) dorsal horn of the spinal cord [56] or higher-order central sites. Inflammatory modulators increase the excitability of these relay neurons by activating protein kinases, blocking specific glycine receptor subtype [57] or upregulating specific sodium channels [58]. In recent years, other forms of central sensitization have been found which involve activity-dependent increases in synaptic efficacy of these relay neurons, with varying onset latencies and memory durations reflecting distinct transcription-dependent or -independent cellular events [59, 60]. These modern forms of central sensitization have been likened to synaptic plasticity-related learning and memory [59–63]. Both peripheral sensitization and the classical or modern forms of central sensitization contribute to hyperalgesia (increased responsiveness to noxious stimuli) although their mechanisms and loci in the pain pathway may vary. In the present framework, such nonassociative and input-dependent hypersensitivity mechanisms of hyperalgesia are in perfect agreement with the notion of primary sensitization as defined in Section 2.2.2.
In contrast to hyperalgesia, which pertains to the same nociceptive input perpetuating the pain sensation, allodynia is hypersensitivity to normally innocuous inputs (such as gentle touch) secondary to a nociceptive input. A prevailing explanation of tactile allodynia is that low-threshold mechanosensitive Aβ afferents with weak synaptic connection at nociceptive relay neurons may be presynaptically or heterosynaptically sensitized by the primary nociceptive input, thus facilitating this normally silent tactile pathway (Fig. 1C) [59, 64]. A similar explanation may also apply to spontaneous pain if the sensitized convergent pathway has tonic activity. Such nonassociative and activity-dependent sensitization of convergent pathway mediating allodynia lends further support for our definition of secondary sensitization as a generic mode of nonassociative learning in neural pathways (Sect. 2.2.2).
In some instances, pain sensations (especially milder types of pain) may habituate upon repeated presentation of the stimuli [65]. The pain habituation, hyperalgesia and allodynia effects of pain sensation are analogous to the habituation, primary sensitization and secondary sensitization forms of nonassociative learning.
2.3. Dual-process theory reconciled
2.3.1. Relations to intrinsic and extrinsic sensitization
Our definitions of primary and secondary sensitization clarify the ambiguities of the dual-process theory. The original theory pertaining to an acute spinal cat preparation was predicated on a sensitization-habituation complex observed in the cat's hindlimb flexion reflex response to a repetitive electrical stimulus. The sensitization was attributed to certain interneurons presumably located in an extrinsic "state" system that was directly activated by the primary stimulus [10, 11]. As such, the sensitization process on which the theory was based is neither intrinsic nor extrinsic sensitization.
By contrast, the sensitization-habituation complex of the cat hindlimb flexion reflex fits well with the notion of primary sensitization and habituation. Rather than mediated by an extrinsic state system as originally proposed, sensitization induced by a repetitive primary stimulus could be expressed in the primary pathway(s) as with habituation, thus evidencing primary sensitization. Indeed, as demonstrated in the frog spinal reflex, primary sensitization and habituation could occur even across the same synaptic junction [38].
Another instance of sensitization in the cat hindlimb flexion reflex was observed when a strong stimulus was delivered at skin sites near the primary stimulus that induced the dual-process response sensitization-habituation. The resulting response sensitization differed from dishabituation in that it decayed spontaneously regardless of the continuance/discontinuance of the primary stimulus [10]. However, this form of sensitization is clearly distinguishable from the first, which was elicited by the primary stimulus itself. Rather, it resembles secondary sensitization as defined in Section 2.2.2 (but with the primary and secondary pathways reversed) in that it was induced by a separate stimulus, perhaps via heterosynaptic or presynaptic facilitation. Thus, primary and secondary sensitization effectively account for all experimental data that formed the cornerstone of the dual-process theory.
Another experimental paradigm that motivated the dual-process theory was the rat acoustic startle response [10, 66]. As with the cat hindlimb flexion reflex, primary and secondary sensitization are evident in this reflex in the form of complex response sensitization-habituation to a repetitive primary (auditory) stimulus and a subsequent, spontaneously-decaying sensitization triggered by a secondary (visual) stimulus. Thus, the present definitions of habituation and primary/secondary sensitization provide a unified theoretical framework that reconciles the dual-process theory and varying definitions of sensitization.
2.3.2. Secondary sensitization as "generalization of sensitization"
The notion of secondary sensitization also rectifies another archaic conjecture of the dual-process theory, namely, the so-called "generalization of sensitization" where sensitization to one input may supposedly spread to other inputs [9, 10]. This conjecture is in actuality an oxymoron as the generalization of intrinsic sensitization simply amounts to extrinsic sensitization, both being instances of secondary sensitization.
2.3.3. Dishabituation as primary or secondary sensitization
According to the dual-process theory, the so-called "dishabituation" effect was neither a disruption of habituation nor an independent process in itself, but rather, an instance of sensitization superimposed on habituation – such that the dual process of habituation and sensitization would adequately account for all incrementing and decrementing behavioral responses. Although this view was later challenged by studies of Aplysia gill- and siphon-withdrawals, which revealed certain subtle differences between dishabituation and sensitization at the behavioral and cellular levels [67, 68], such discrepancies were subsequently found to be attributable to an interaction between habituation and inhibition in some modulatory pathways [69, 70]. Thus, dishabituation may represent a form of sensitization that is gated by habituation (see Sect. 4.3.2).
Most previous studies of dishabituation used a secondary stimulus to reverse habituation. In a recent model of tail-elicited siphon withdrawal in Aplysia [69, 71], however, dishabituation is expressed in reflex pathways both ipsilateral or contralateral to the primary stimulus even though habituation and sensitization are expressed only in the pathway ipsilateral to the primary stimulus. This finding is consistent with the notion of primary and secondary sensitization, in that dishabituation could be expressed in both primary and secondary pathways rather than confined to the primary pathway as suggested by the dual-process theory.
Thus, the present framework brings into harmony a body of confounding observations relating to intrinsic, extrinsic and anomalous sensitization, generalization of sensitization and dishabituation, which are otherwise incongruous with the dual-process theory.
3. Desensitization: a novel form of nonassociative learning
The above framework of habituation and primary/secondary sensitization is complementary to a new mode of nonassociative learning called response desensitization. In the following, we review the experimental evidence of response desensitization and show how this novel concept may yield new insights to some sensory remapping behaviors such as phantom sensation and drug addiction.
3.1. Desensitization as nonassociative learning
3.1.1. Desensitization as secondary habituation
A corollary to the above definitions of primary and secondary sensitization is the notion of primary and secondary habituation. For simplicity, we abbreviate primary and secondary habituation as habituation and desensitization, respectively (Fig. 2A).

Response desensitization and phantom sensation. A. Schematic illustration of desensitization (D) in relation to habituation (H) and primary sensitization (S). The secondary pathway is desensitized by heterosynaptic STD (filled inner triangle, blue) secondary to the primary stimulus, leaving it dormant (broken line). B. Phantom sensation: following deafferentation, the primary stimulus ceases and the secondary pathway is re-sensitized, producing a phantom sensation.
3.1.2. Desensitization of descending pathways
Instances of response desensitization have often gone unnoticed because it is easily mistaken for "habituation". An example is the crayfish tail-flip escape response to repetitive primary afferent activations, which exhibited tonic GABAergic inhibition via a descending pathway to the motor circuitry [72]. Desensitization as a new mode of nonassociative learning was first formalized in studies of the rat Hering-Breuer reflex, which evidenced a decrementing response adaptation secondary to habituation upon sustained application of a primary (vagal) stimulus and a short-term memory of the adaptation upon termination of the primary stimulus [3]. The secondary adaptation component was selectively abolished by pontine lesion or pharmacological blockade of NMDA receptor-gated channels, suggesting STD of tonic excitation (or STP of tonic inhibition) of some descending ponto-medullary pathway [73, 74]. The habituation-desensitization paradigm exemplified by these animal models is in contrast to the purported habituation-sensitization dual process that has permeated previous studies of nonassociative learning.
3.1.3. Desensitization as generalization/transfer of habituation
Indeed, response desensitization has long lurked under the dual-process theory as the putative "generalization of habituation" [75] or "transfer of habituation" [76, 77] (see Sect. 2.1.2), sometimes also called "extrinsic habituation" [37]. Generalization of habituation between sensory modalities is best illustrated by the Aplysia siphon and gill withdrawal reflex, in which habituation training at one sensory site (gill) may transfer to an untrained site (siphon) through heterosynaptic modulation (e.g., heterosynaptic depression or inhibition [78]) via the peripheral nervous system [77] or some perceptron-like parallel processing [79]. Generalization of habituation between sensory sites is seen in the escape swim of the marine mollusk Tritonia diomedea [80] and the shortening reflex of the medicinal leech Hirudo medicinalis [48, 54], where response habituation elicited at one body site may transfer to an untrained site.
The putative generalization/transfer of habituation is converse to the secondary sensitization in similar animal models (albeit with differing stimulus intensity or type). In particular, the heterosynaptic inhibition in Aplysia is mechanistically opposite to the heterosynaptic facilitation that is thought to contribute to the extrinsic sensitization of its gill- and siphon-withdrawal reflex [51]. Therefore, generalization/transfer of habituation is operationally and mechanistically analogous to secondary sensitization but with differing response polarity and activation threshold. As such, it represents a distinct form of nonassociative learning in its own right. These observations lend further support for response desensitization as a bona fide mode of nonassociative learning rather than an extension of habituation.
3.2. Desensitization and referred pain sensations
3.2.1. Somatosensory remapping
The present notion of response desensitization as a new mode of nonassociative learning may shed light on the enigmatic "referred phantom sensation" (such as phantom pain) experienced by some amputees [46, 81, 82]. Recent findings have linked such phantom sensations to remapping at cortical [83] and thalamic or sub-thalamic levels [84] such that the deprived primary pathway is referred to a separate pathway with distinct receptive field and an expanded central representation that invades the original primary representation. A possible mechanism of such remapping is collateral sprouting (a rather slow process); another prevailing hypothesis is that such referred pathway may be preexisting but latent, and are unmasked after deafferentation [85, 86] presumably by disinhibition [86–89].
However, disinhibition is a fast neurotransmission process that may take effect rapidly. Although rapid somatosensory reorganization post-deafferentation (within minutes) has been reported [90, 91], amputated subjects generally do not experience phantom sensations until much later. As pointed out by Chen, Cohen and Hallet [92], the mechanisms of nervous system reorganization following injury may differ depending on the timeframe. The timeframe of phantom sensations (reportedly in hours or days) [93, 94] does not appear to match those of sprouting (in weeks or months) or disinhibition (in seconds and minutes), suggesting that other mechanisms of remapping might be involved.
3.2.2. Two-tier learning model of phantom sensation
As an alternative hypothesis, we suggest that phantom sensation might result from unmasking of latent somatosensory pathways through learning and memory instead of (or in addition to) disinhibition, perhaps by means of synaptic plasticity such as long-term potentiation (LTP) (which has been implicated in the reshaping of cortical motor maps [95]). In support of this hypothesis, recent evidence indicates that somatosensory reorganization associated with perceptual learning in human subjects may occur within a timeframe of hours of training and may be controlled by similar basic mechanisms that underlie NMDA receptor-dependent synaptic plasticity such as LTP [96]. Furthermore, similar molecular mechanisms that underlie NMDA receptor-dependent homosynaptic LTD in the rat visual cortex have been linked to the characteristic visual impairment resulting from hours of monocular deprivation during early postnatal life [97, 98]. These recent findings point to a possible role for synaptic plasticity such as LTD/LTP in the masking/unmasking of somatosensory pathways before and after deafferentation.
In keeping with the notion of unmasking of preexisting pathways, we propose a two-tier nonassociative learning model of somatosensory organization, with the primary and referred sensations being mediated by a primary pathway and a latent surrogate pathway, respectively (Fig. 2B). The primary and surrogate (secondary) pathways are functionally equivalent to the primary and secondary pathways of nonassociative learning (Fig. 1A). In contrast to the inhibition/disinhibition hypothesis of unmasking, the present theory postulates that the surrogate pathway may be normally desensitized and, hence, rendered ineffective by the primary pathway. Deafferentation abolishes ("gates off", see Sect. 4.1) the primary pathway and its sensory dominance, allowing the intact surrogate pathway to strengthen over time through synaptic plasticity processes such as LTP. The resultant sensitization effect unmasks the surrogate pathway, giving a phantom sensation. In particular, if the primary pathway is part of a pain pathway then the referred phantom sensation may give rise to phantom pain if the surrogate pathway or its referred central representation in the pain pathway is hypersensitized.
The above model suggests a new perspective to the neural reorganization that reportedly underlies phantom sensations. In those patients, recordings in the thalamic region that normally respond to the missing limb revealed new receptive fields on its stump; microstimulation of this remapped thalamic region evoked phantom sensations of the missing limb, including phantom pain [84]. These findings suggest that the thalamic representation of the amputated limb was remapped to a surrogate pathway from the stump of the missing limb, presumably via learning. This two-tier learning model of somatosensory remapping based on the general theory of nonassociative learning provides a coherent explanation of referred phantom sensation and phantom pain and related experimental observations in relation to a general class of synaptic plasticity and nonassociative learning processes widely reported across animal phyla from invertebrates to humans.
3.2.3. Capsaicin sensitization and "desensitization"
The response desensitization as defined above is distinct from the desensitization of nociception associated with certain irritants such as capsaicin, the pungent chemical in red chili pepper. In human subjects, the burning/pricking sensation elicited by oral capsaicin typically intensifies with its repeated applications at an ISI of ~1 min, a hyperalgesic effect that is akin to response sensitization. However, following a hiatus of several minutes reapplication of capsaicin elicits a much weaker sensation. This latent refractory process has been called "desensitization" by some authors [47, 99] in analogy to desensitization of nociceptive vanilloid receptors, which are generally thought to mediate the pungency of capsaicin [100].
The waxing and waning of the pungency following repeated capsaicin application is reminiscent of the sensitization-habituation dual process of nonassociative learning [10, 37]. As such, the refractory response to capsaicin following sensitization training should be a classic case of habituation in the primary nociceptive pathway instead of "desensitization". The successive increase and decrease of capsaicin pungency on varying timescales indicates that the habituation component develops more slowly but lasts longer than sensitization. If so, a weaker sensitization effect should unmask the progressive development of habituation during behavioral training. Indeed, repetitive oral application of other irritants such as nicotine, menthol, zingerone or mustard oil elicits sensations that decline successively across trials [101–104], evidencing response habituation with weak or no sensitization.
It has been suggested that capsaicin sensitization may be mediated by an increase in excitability of peripheral nociceptors or central relay neurons, or spatial recruitment of vanilloid receptors in nociceptor endings [99]. These hypothesized cellular mechanisms are consistent with primary sensitization (as defined in Sect. 2.2.2). On the other hand, capsaicin sensitization has been shown to promote hypersensitivity to and aftersensations of other pain stimuli applied to the affected site [105]. This secondary hyperalgesic effect is indicative of secondary sensitization or central sensitization involving secondary nociceptive or allodynic pathways, perhaps via wide dynamic range neurons in spinal dorsal horn [56].
3.3. Desensitization and drug addiction
It is well-known that repeated drug administrations may result in drug tolerance and/or sensitization [106], which are attributable to the dual process of response habituation and sensitization [107–109]. The notion of response desensitization presently proposed adds a new dimension to the understanding of the behavioral mechanisms of drug tolerance and, indeed, of drug craving and addiction itself.
3.3.1. Sensitization models of craving
Craving plays an important role in the pathogenesis of many addictive disorders (such as alcohol, nicotine, narcotic or other psychostimulant drug dependencies) but its mechanism has remained unclear [110, 111]. Current models of craving (for overviews, see [112, 113]) ascribe this psychophysical drive to certain cognitive or neuroadaptive processes such as behavioral sensitization – a phenomenon characterized by enhanced psychomotor and motivational effects of an addictive drug along with increased midbrain dopamine neurons reactivity upon repeated drug applications [114, 115]. Recent evidence suggests a possible link between behavioral sensitization and LTD of AMPA receptor-mediated synaptic transmission in the nucleus accumbens [116]. Although behavioral sensitization is not tantamount to craving, it is often thought to induce compensatory homeostatic or incentive-motivational adaptations which, in turn, could incubate craving or "pathological wanting" that may be rekindled by stress or drug cues during prolonged abstinence [117–121].
3.3.2. Desensitization model of craving
In contrast to previous sensitization models of craving during relapse, we propose a desensitization model of craving during the onset of addiction, as follows (Fig. 3). Central to our model is the notion that craving of any kind may represent an innate (rather than acquired) instinct, not fundamentally different than basic instincts such as thirst, hunger, sex, and yearning for love or happiness, etc. However, unlike ordinary psychophysical drives that are critical for animal survival or procreation and are expressed at birth or during puberty, craving for substance of abuse is functionally deleterious (hence "pathological") and hence its expression is likely to be repressed through evolution. In a naïve (or "innocent") state, pathological craving may be inhibited intrinsically by certain tonic central inputs that promote self-restraint (or self-reward) – presumably via some midbrain dopaminergic or glutamatergic pathways – hence keeping craving and addiction in check. Exposure to an addictive drug may disrupt this equilibrium state and arouse addiction in multiple possible ways. Firstly, it activates (i.e., "gates on", see Sect. 4.1) a normally-latent primary sensory pathway that serves to relieve craving and evoke gratification in addition to the central (secondary) craving-inhibiting pathways, thus producing an immediate euphoric effect. Secondly, activity in the primary pathway may in turn desensitize the secondary craving-inhibiting pathways, thus debilitating the brain's natural defensive against addiction. This critical step may correspond to the "loss of inhibitory control in decision making" against addiction suggested by some investigators [118]. Thirdly, desensitization in the secondary pathways (together with possible habituation in the primary pathway) may cause increasing tolerance to the drug, hence drawing higher-and-higher dosages in order to relieve craving or regain euphoria, further deepening the addiction. Finally, abrupt abstinence in a desensitized state may unleash the craving and precipitate any accompanying withdrawal symptoms, which may subside over time as the reward system gradually re-sensitizes.

Graphical depiction of a theoretical model of addiction and recovery. (A) In the naïve state, an intrinsic secondary input suppresses craving via an inhibitory pathway. (B) Upon initial exposure to an addictive stimulus, the craving center is further inhibited by the primary pathway resulting in a feeling of euphoria. (C) With continued exposure to the addictive stimulus, the secondary pathway is desensitized by the primary pathway (with possible habituation in the primary pathway not shown), resulting in craving and the onset of addictive behavior. The latter can lead to more craving via a positive feedback vicious cycle as well as mobilization of a tertiary process that can independently perpetuate the addictive behavior. (D) Sudden withdrawal of the addictive stimulus precipitates a state of "cold turkey" characterized by enhanced craving due to the loss of the primary input, continued desensitization of the secondary pathway and continued positive feedback from the addictive behavior. (E) Sustained abstinence will allow resensitization of the secondary pathway and temporary relief of craving. The addictive behavior subsides, but the tertiary process is still lurking and intensifying. Complete rehabilitation to the naïve state (A) calls for extirpation of the tertiary process. (F) Otherwise, reactivation of the tertiary pathway by contextual cues, memory or stress could once again desensitize the secondary pathway, triggering a relapse.
The above model predictions depict the early drug-induced degeneration of the brain's reward system from a naïve state to a desensitized state and the subsequent recovery during abstinence. However, once exposed to an addictive stimulus the brain may not be totally "innocent" anymore in that the reward system may begin to give way to other, non-reward related hysteretic mechanisms which may ensue even after recovery. For example, repeated drug exposures may mobilize other neuroadaptive processes, such as behavioral sensitization, which promote relapses (see above). Also, a craving-driven addiction could later turn into a completely craving-free habit or even compulsion [122–124], perhaps via the dynamic modulation of some cortical-basal ganglia circuits [125]. These tertiary hysteretic processes could perpetuate the drug-seeking behavior independent of dopamine-mediated reward [126, 127], with or without provoking craving [128, 129]. On the basis of these observations, we suggest that desensitization of secondary craving-inhibiting pathways and sensitization of tertiary hysteretic pathways may underlie the acquisition and maintenance of addiction behavior, respectively, such that their sequential inductions upon the first encounter with an addictive drug create a watershed effect that irreversibly usurps the brain's built-in self-restraint mechanism to stave off addiction (Fig. 3).
3.3.3. Context-dependent habituation and desensitization
In contrast to habituation and primary sensitization (the quintessential dual process of nonassociative learning), desensitization and secondary sensitization could also involve associative training by primary and secondary stimuli. Some classic models of nonassociative learning such as Aplysia siphon and gill-withdrawal reflex are known to demonstrate classical conditioning, which shares similar cellular and molecular mechanisms with nonassociative learning [130–132]. A variant of conventional habituation called context-dependent habituation has been shown in some animal models such as the nematode C. elegans [133] and the crab Chasmagnathus [134], where the retention of habituation after training is also influenced by certain environmental cues. The dependence on environmental cues indicates that the primary stimulus-response pathway is not merely habituated by the primary input but likely also desensitized by certain context-dependent sensory inputs in an associative manner.
Such context-dependent learning effect has important implications in certain addictive disorders, where environmental cues are known to promote drug-dependent behavioral sensitization and relapses [121, 135]. Hence, environmental cues may serve as a conditioned stimulus to certain tertiary pathways which, when activated, may independently desensitize the secondary craving-inhibiting pathways during abstinence (Fig. 3) in a manner analogous to the conditioned drug response in associative learning [136, 137]. Further studies are needed to elucidate the possible role of associative learning in the induction of secondary sensitization and desensitization in these experimental models.
4. Temporal filtering by nonassociative gating
The notion of primary and secondary sensitization introduced in Section 2 underscores a novel behavioral paradigm we call nonassociative gating. Several forms of nonassociative gating have been identified that provide computational capabilities complementary to nonassociative learning.
4.1. Input gating
4.1.1. Input gating and memory recall
An important property of neurotransmission memory is that it is discernible only during recall, i.e., when the pathway is activated by a stimulus eliciting combined reflex and memory responses. Once the activation ceases, the corresponding memory becomes latent and unobservable. We call this an input gating effect, namely an on-off switching of neurotransmission memory by the stimulus itself (Fig. 1B). Thus, a primary stimulus that induces learning and memory in the primary pathway may simultaneously recall the memory by "gating" it on. Conversely, memory in the primary pathway is automatically gated off once the primary stimulus disappears and thus any residual response must reflect persisting activity in the secondary pathway [2, 138].
For instance, deafferentation effectively gates off the primary pathway (Sect. 3.2.2). Another example is the phenomenon of pain sensitization (Fig. 1C). In hyperalgesia, increased pain sensation due to peripheral or central sensitization is elicited when a noxious stimulus is applied but these effects are promptly relieved (gated off) once the stimulus is removed. In the case of addiction, initial exposure to an addictive drug gates on a normally latent craving-suppressing primary pathway thereby setting off the addiction vicious cycle (Fig. 3).
4.1.2. Maximum interstimulus interval
The notion of input gating has important implications in determining the maximum ISI for nonassociative learning experiments. With a repetitive stimulus the primary memory is simultaneously induced and recalled at successive stimulus episodes but is gated off in between. Therefore, for optimal memory recall an ISI should be no longer than the decay time of the primary memory. This maximum ISI condition for training is tacit in studies of nonassociative learning reported in the literature.
4.1.3. Primary and secondary memory
The activity-dependent and pathway-specific properties of input gating make it a useful behavioral marker for memory in the primary pathway vs. those via the secondary pathway, hereinafter referred to as primary and secondary memory, respectively. This marker readily distinguishes nonassociative learning modes mediated by the primary and secondary pathways. Thus, habituation and desensitization are readily distinguished by the absence/presence of a STD memory trace in the resultant behavioral response, whereas primary and secondary sensitization are distinguished by corresponding absence or presence of a STP memory trace. These criteria have been successfully applied to the experimental classifications of primary and secondary memory in the rat respiratory chemoreflex and mechanoreflex [2, 3, 74, 138–140].
4.2. Output gating
4.2.1. Output gating and refractory period
Memory recall requires not only an enabling input but, also, an observable output. In some sensory modalities such as olfaction and vision, the output of nonassociative learning is registered continuously as a sensory percept in the brain without fail and thus the memory trace is gated only by the input. In other sensory modalities, however, the behavioral output may be registered as discrete motor response (or other effector response) with a definite refractory period. If so, the memory trace may be gated off during the refractory of the output as well. We call this an output gating effect in contradistinction to input gating.
4.2.2. Minimum interstimulus interval
The notion of output gating has important implications in determining the minimum ISI for nonassociative learning experiments. For example, in the classic Aplysia gill-withdrawal reflex a strong tactile stimulus to the siphon may produce a strong and long-lasting gill response which, if unabated, may mask the responses to subsequent stimuli [141]. In this case, the minimum ISI for producing a demonstrable habituation effect is limited by the refractory period of the gill response. Conversely, a behavioral system with negligible refractory in the effector would require little or no ISI. This minimum ISI condition is tacit in studies of nonassociative learning reported in the literature.
4.3. Extrinsic gating
4.3.1. Phase-dependent gating
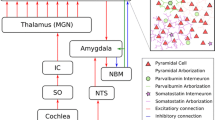
In contrast to input and output gating, which are intrinsic to any stimulus-response pathway, neurotransmission gating may also arise from extrinsic factors. In particular, we define phase-dependent gating as the on-off switching of a stimulus-response pathway by a phasic command signal independent of the pathway's input and output. This type of extrinsic gating is exemplified by the mammalian carotid chemoreflex modulation of the respiratory rhythm in which separate STP and STD chemoreflex afferent pathways are temporally gated to either the inspiratory or expiratory phase of the respiratory pattern generator (Fig. 4). Such phase-dependent gating allows the chemoreceptor input to selectively modulate each respiratory phase in an orderly manner via separate STP or STD pathways, much like the on-off switching of two-way traffic lights at an intersection [74, 138].

Phase-dependent gating and primary/secondary (monophasic/biphasic) integration and differentiation mechanisms as exemplified by the carotid chemoreflex modulation of respiratory rhythm in rats. The chemoafferent input from the carotid sinus nerve (CSN) is habituated by a monophasic (input gated) differentiator in the nucleus tractus solitarius (NTS). The output from NTS is relayed by parallel pathways to I (inspiratory) or E (expiratory) neurons of the respiratory pattern generator or to I-PMN (inspiratory premotor neuron). Switches denote gating to either I or E phase. Each pathway is modulated by two biphasic integrators (∫L with fast or slow time constant) which either add or subtract to produce net short-term potentiation (STP) or depression (STD) effects. Adapted from [138].
4.3.2. Learning-dependent gating
Gating may also be triggered by activity-dependent plasticity rather than a phasic command. This type of extrinsic gating is exemplified by Aplysia tail-elicited siphon withdrawal reflex where a modulatory network that normally inhibits the sensitization of contralateral siphon response is relieved after habituation (see Sect. 2.3.3). Such learning-dependent gating has been suggested to account for the bilateral expression of dishabituation vis-à-vis ipsilateral expression of sensitization in this experimental preparation [69, 71].
4.4. Nonassociative gating as Boolean 'toggle switch'
4.4.1. Nonassociative gating: a new behavioral paradigm
Input/output gating and extrinsic gating are instances of nonassociative gating. As with nonassociative learning, the induction of such gating effects is activity-dependent and nonassociative, and their expressions may be intrinsic or extrinsic to the neurotransmission pathway. Furthermore, nonassociative gating displays certain computational characteristics that are complementary to nonassociative learning.
4.4.2. Boolean on-off switching and temporal filtering
In input gating, the stimulus itself provides a logic 'on' signal that enables memory recall whereas in output gating, the effector response serves as a logic 'off' signal that momentarily disables or attenuates memory recall. Similarly, in phase-dependent or learning-dependent gating an extrinsic signal independent of the current input or output provides the on-off command for the memory trace. Thus, nonassociative gating operates like a Boolean toggle switch that may turn the memory trace on or off depending on the logic value of the command signal.
Alternatively, nonassociative gating may be viewed as a temporal filter that selectively passes or stops memory recall within specific time windows during nonassociative learning. Such signal filtering in the time domain contrasts with the signal filtering in the frequency domain by nonassociative learning.
5. Frequency filtering by nonassociative learning
Based on the above, we propose a theory of gated integral-differential neural computation (or low-pass and high-pass frequency filtering) by nonassociative learning. Prescott [31] has proposed a mathematical model that mimics the kinetics of habituation and intrinsic sensitization development and their interaction using linear first-order differential equations. Dragoi [142] has proposed a similar model of suppressive and facilitatory interactions during nonassociative learning but with nonlinear first-order differential equations in order to simulate the rate sensitivity property of habituation. Staddon and Higa [143] have proposed a feedback/feedforward integrator model of habituation. Shen [144] has proposed a STP model of neural integrator. The present theory differs from the previous models in that it is structurally-based and includes all four modes of nonassociative learning (Sect. 2.4.1) as well as nonassociative gating (Sect. 3.1.3), which provide a complete mathematical basis for gated integral-differential computation.
5.1. Definitions of neural integrator and differentiator
5.1.1. Leaky integrator and differentiator
Numeric integration and differentiation are elemental calculus operations. They also underlie all temporal dynamics and kinematics phenomena in Nature. An analog integrator/differentiator is a physical process that demonstrates integral/differential input-output transformation in real time. Analog integrator and differentiator are subject to response-limiting leakages. Leaky integrators are commonly used in electrophysiology experiments to obtain a moving-average estimate of neuronal firing frequency called 'neurogram.'
5.1.2. Integrator and differentiator response characteristics
In the time domain, a leaky integrator's response to a constant-step input exhibits exponential saturation during on-transient and exponential decay during off-transient, whereas the corresponding response of a leaky differentiator demonstrates exponential decay from an initial overshoot and exponential recovery from rebound undershoot (Fig. 5A; Eq. 2 in Appendix I). An integrator or differentiator that sustains an off-transient response is said to be biphasic (or else, monophasic); it is said to be inverted if the gain is negative (Eq. 3 in Appendix I). The dynamical order of a compound integrator/differentiator is the number of integrators/differentiators it is composed of, and its memory order is the number of component integrators/differentiators that are biphasic. Under these broad definitions, a neural system that displays such integral/differential neurotransmission characteristics is called a neural integrator/differentiator.

Time and frequency response characteristics of integrator (left panels) and differentiator (right panels). A. The temporal response of a leaky integrator to a constant-step stimulus (horizontal bar) consists of abrupt reflex increase/decrease of the response at stimulus onset/cessation followed by exponentially increasing/decaying (potentiation/afterdischarge) on/off transients. A leaky differentiator has similar reflex components but with exponentially decaying (accommodation) on-transients and rebound off-transients, which are opposite to those of an integrator (overlaying dotted lines). In both cases the off-transients may be rectified, with the response becoming monophasic (not shown) instead of biphasic. The time scales chosen are typical of oculomotor integrator and respiratory integrator. B. In the frequency domain, an integrator/differentiator behaves like a low-pass/high-pass filter. The pass-band in both cases is the frequency range where the neurotransmission gain (normalized to unity) is highest and relatively constant. The high and low cut-off frequencies (vertical dotted lines) of these filters are inversely proportional to the time constants of the corresponding integrator and differentiator shown in A. C. Examples of RC integrator (C = 3.77, R1 = 1.60, R2 = 24.0, R3 = 0.73) and differentiator (C = 1.0, R1 = 1.75, R2 = 24.0, R3 = 10.2) circuits with a current source (IS) input and voltage (V0) output. Units are arbitrary and RC values correspond to parameter values as defined in Eq. 1 (Appendix I) for integrator (a = 0.125; b = 0.125; c = 1; d = 0.5) and differentiator (a = 0.125; b = 0.125; c = -1; d = 1.5).
5.1.3. Low-pass and high-pass frequency filter characteristics
From linear systems theory [145], integrator/differentiator response characteristics in the time domain correspond to low-pass/high-pass filter characteristics in the frequency domain (Eq. 4 in Appendix I). The time constant of a leaky integrator or differentiator is inversely proportional to the cut-off frequency of the equivalent low-pass or high-pass filter, respectively (Fig. 5B). In addition to frequency filtering, a leaky integrator/differentiator also introduces phase shift (phase-lag/phase-lead) in the input-output relationship.
5.1.4. Complementarities of neural integrator and differentiator
Mathematically, a neural differentiator is an additive complement of an integrator, i.e., the response of a differentiator is complementary to that of an integrator with similar time constants (Fig. 5A). The frequency characteristics of a low-pass and high-pass filter with matched cut-off frequencies are also complementary to one another (Fig. 5B). Indeed, combination of such complementary filters in parallel approximates an all-pass filter with constant throughput gain at all frequencies.
5.2. Reverberation models of neural integrator
5.2.1. Reverberating neural network hypothesis
A widespread hypothesis of neural integrator is that of reverberation in a recurrent neural network. This hypothesis has been studied most extensively in two experimental models: the "afterdischarge" phenomenon in the chemoreflex control of breathing [146] and the oculomotor integrator [147–149]. The proposed mechanism involves two steps: intrinsic membrane properties of a neuron provide a trace capacitance that acts as a seed for the integrator, and positive feedbacks via a recurrent network allow continual refreshment of the seed. The network reverberation hypothesis is bolstered by the finding that the goldfish oculomotor integrator during normal saccadic movements could not be reproduced by saccade-like changes in neuronal firing induced by intracellular current injection, suggesting that the integrator effect is dependent on persistent changes in synaptic inputs [150]. It has been proposed that such a recurrent network can be made robust by certain bistable neuronal processes [151, 152] or recurrent synaptic excitation with asynchronous transmitter release [153], or by external sensory error feedback [154].
5.2.2. Reverberating neuronal ion-channels hypothesis
Alternatively, reverberation of excitatory activity could also occur at the single-neuron level via a cascade of membrane ion channels [155, 156]. The persistent activity can be elicited bi-directionally by excitatory and inhibitory inputs in a graded fashion, similar to a biphasic integrator.
5.2.3. Dendritic calcium self-amplification hypothesis
A recent mathematical model [157] posits that temporal integration in a single neuron may result from self-amplifying calcium dynamics through a cellular process called "calcium induced calcium release." According to this model, synaptic inputs modulate the regenerative propagation of calcium waves along dendritic processes, resulting in calcium-dependent currents that vary directly with the temporal sum of prior synaptic inputs. This mechanism of neural integrator is thought to be robust by virtue of the intrinsic nonlinear spatiotemporal summation of the calcium waves.
5.3. Nonassociative learning models of neural integrator and differentiator
5.3.1. Nonassociative learning hypothesis of neural integrator and differentiator
Although the above models of neural integrator are all plausible, none of them can explain neural differentiator. In contrast, nonassociative learning is based on activity-dependent changes in synaptic efficacy [51, 139, 140, 158] or neuronal excitability [10, 159] that are well documented in single neurons. These neural mechanisms are highly stable and robust with short- or long-term memories ranging from seconds to days or months, and provide a plausible explanation of both neural integrator [2, 139, 144] and differentiator [2, 140]. For example, synaptic STD or LTD in brainstem NTS [160–162] may provide the habituation or monophasic differentiator effects in various cardiorespiratory reflexes [3, 138] (see Fig. 4).
5.3.2. Primary and secondary integrator/differentiator
To a first approximation, the integrator and differentiator characteristics defined in Figure 5 are mimicked by the augmenting characteristics of primary and secondary sensitization and decrementing characteristics of habituation and desensitization, respectively. In particular, primary sensitization/habituation represents a primary integrator/differentiator with monophasic characteristics whereas secondary sensitization/desensitization represents a secondary integrator/differentiator with biphasic characteristics (see Appendix II for mathematical details).
The monophasic characteristics of primary sensitization and habituation are due to input gating (Sect. 3.1). The biphasic characteristic of desensitization [3, 72] may be ascribed to tonic activity in the secondary pathway, which provides a continual recall of the secondary memory (Sect. 2.5). In contrast, in secondary sensitization the roles of the primary and secondary pathways are often reversed (sensitization in the primary pathway induced by a strong secondary stimulus), and thus a biphasic integrator response would be sustained by continued repetitive application of the primary stimulus itself. Figure 4 shows examples of primary and secondary integrator and differentiator demonstrated in the mammalian carotid chemoreflex pathways [138].
Although monophasic integrator and differentiator are driven directly by the primary input, biphasic integrator and differentiator require a secondary input with persistent activity. The latter could come from reverberations at the neuronal or neural network levels in central neurons [163]. Alternatively, it may come from tonic central or peripheral inputs. For example, tonic secondary inputs to the biphasic integrators in the carotid chemoreflex pathways (Fig. 4) or vagal Hering-Breuer reflex may derive from central chemoreceptors, which provide persistent activation of the respiratory pattern generator and its afferent pathways [74].
According to the complementary relationships of neural integrator and differentiator (Sect. 5.1.4), a secondary differentiator is the combination of two separate processes: a primary reflex and an inverted secondary integrator (Fig. 6C, 6D). Similarly, a primary differentiator is comprised of a primary reflex and an inverted primary integrator (Figs. 6A, 6C). Thus, a primary or secondary neural differentiator is realized by nonassociative learning as the difference (antagonistic excitation-inhibition combination) between a primary reflex and a primary or secondary neural integrator.

Compound neural integrator and differentiator models. Boxes show model simulations in arbitrary units and parameter values. Conventions are same as Fig. 1. A. Primary differentiator-secondary integrator is realized by activity-dependent habituation-sensitization in corresponding primary-secondary pathways. Step application of primary input at a constant firing rate (inset) induces synaptic STD and STP in primary and secondary pathway, respectively, resulting in temporal differentiation and integration of the transmitted signals (lower and upper boxes). Resultant response (last box) at output neuron shows a compound differentiator-integrator characteristic. B. A double integrator may arise from STP in both primary and secondary pathways. C. A double differentiator is similar to a double integrator but with STD instead of STP in both pathways. D. A primary integrator-secondary differentiator can be realized in a similar fashion with a STP-STD combination. Structurally, primary differentiator (A, C) and secondary differentiator (B, D) are comprised of the primary reflex in conjunction with an inverted primary and secondary integrator, respectively, demonstrating the complementarities of integrator and differentiator. See text and Appendix.
5.3.3. Second-order integrators/differentiators
The four basic modes of nonassociative learning – habituation, desensitization, primary sensitization, secondary sensitization – constitute a complete orthogonal (non-redundant) mathematical basis that empowers the brain to perform basic integral-differential calculus of any dynamical and memory orders. In particular, an integrator and differentiator may combine to form an integrator-differentiator pair with second-order dynamics and frequency characteristics. The simplest example is a primary integrator-differentiator pair in the form of a sensitization-habituation complex produced by repetitive application of a primary stimulus, as demonstrated in the classic hindlimb flexion reflex of the spinal cat or the rat acoustic startle reflex shown in the dual-process theory [9–11]. The combined primary integrator-differentiator pair acts like a band-pass or band-stop filter, which selectively admits or rejects afferent inputs that are fluctuating around certain mid-frequencies.
Nonassociative learning in the primary and secondary pathways may also work in tandem to form second-order integrator-differentiator pairs. Four different combinations are possible. Habituation in conjunction with secondary sensitization gives a primary differentiator – secondary integrator pair (Fig. 6A). This is similar to the primary integrator-differentiator pair in the acoustic startle reflex but with a secondary memory. Similarly, concurrent primary and secondary sensitization results in a second-order low-pass filter in the form of a primary integrator and secondary integrator (Fig. 6B). A second-order differentiator/high-pass filter is formed by habituation in the primary pathway and desensitization in the secondary pathway (Fig. 6C), as demonstrated in the Hering-Breuer reflex or carotid chemoreflex modulation of expiratory duration in the rat [2, 3, 74, 140]. Finally, it is conceivable that sensitization-desensitization in the primary-secondary pathway may give rise to a second-order integrator-differentiator pair (Fig. 6D) with band-pass or band-stop filter characteristics similar to those in Figure 6A. Examples of second-order integrator and differentiator are shown in Figure 4 for the mammalian carotid chemoreflex pathways [138].
6. Nonassociative learning and brain intelligence
In addition to performing kinematic transformations, nonassociative learning may also contribute to the integral-differential calculus and Boolean logic computations that are basic to brain decision processes. These neural integrators, differentiators and logic operators provide some of the basic building blocks of brain intelligence.
6. 1. Intelligent roles of neural integrator and differentiator
6.1.1. Neural integrator: possible roles in sensory defensiveness, alarm reaction and sensorimotor instability
Behaviorally, a neural integrator can boost an animal's responsiveness to a recurrent noxious stimulus and (by cross-modal transfer) to other inputs even after the primary stimulus has ceased. The resultant heightening and widening of vigilance put the animal on the alert once this self-defense mechanism is triggered. This 'alarm reaction' instinct sets one free to economize and relax (by staying idle and calm) most of the time until fear-arousing episodes (e.g., terrorist attacks) set in. On the other hand, inordinate sensitization of the primary or secondary pathways could result in hypersensitivity to innocuous sensory stimuli. This mechanism is compatible with certain forms of sensory integration dysfunction such as sensory defensiveness [164] or nonassociative fear or anxiety toward impending adverse stimuli [165].
In sensorimotor control, temporal integration of error feedback may help to minimize the resultant steady-state error. On the other hand, low-pass filtering of the feedback signal may introduce excessive phase lags (phase delays) that tend to destabilize closed-loop control [145]. It has been suggested that spontaneous oscillations of sensorimotor regulation may develop with increased delays in sensory feedback [166–168].
6.1.2. Neural differentiator: possible roles in selective attention, central resetting, sensory self-organization and fail-safe compensation
Functionally, a neural differentiator is a high-pass filter that preferentially admits time-varying signals, rejecting any DC biases that tend to saturate or suppress neurotransmission. This high-pass filtering effect allows the animal to automatically recalibrate the sensitivities of the primary and secondary pathways against varying background activities thereby extending the dynamic range of the stimulus-response relationship. Thus, a sustained primary input (e.g., hypertension or bronchopulmonary afferent hyperactivity) may induce compensatory habituation-desensitization or "central resetting" of primary and secondary pathways [160, 169, 170], whereas abolition of the primary input (e.g., due to impairment of sensory receptors or afferent pathways) may elicit compensatory dishabituation of the primary pathway and re-sensitization of the secondary pathway. As such, the secondary pathway provides a reserve surrogate or backup for the primary pathway should it ever fail. Such sensory self-organization provides a fail-safe mechanism for optimal compensation against hyper- or hypo-activity of afferent feedback in sensorimotor systems [2, 171, 172].
Another useful function of habituation and desensitization is to tune out repetitive inputs that prove to be innocuous, thus allowing selective attention to potentially important inputs [173–175]. Failure to do so may lead to sensory defensiveness in some individuals [164, 176] and in patients with autism [177], as well as nonassociative fear and anxiety [165] or other forms of hyper-reactivity. On the other hand, because habituation and desensitization tend to suppress persistent afferent inputs, their overexpression may have deleterious effects in certain sensorimotor reflexes. For example, abnormal expression of LTD in the NTS of newborn mice devoid of functional NMDA receptors is implicated in the progressive respiratory failure and early death in these mutant animals [162].
6.1.3. Compound neural integrator-differentiator: possible roles in novelty detection and selective attention
A compound integrator-differentiator/differentiator-integrator is functionally equivalent to a band-pass/band-stop filter that preferentially admits/rejects inputs whose temporal variability falls within some intermediate frequency band or time scale. For example, the acoustic startle reflex is sensitized by continuous background noise or novel inputs but may habituate on discrete repetitive tones [66]. Such a combined sensitization-habituation or sensitization-desensitization response pattern allows maximal vigilance to unexpected (and potentially alarming) inputs over mundane and insipid ones, thus sharpening novelty detection and selective attention [178]. On the other hand, excessive band-selective filtering may lead to paranoia; indeed, increased sensitization and reduced habituation are trait markers of patients with schizophrenia [179]. The possible role of this band-selective filtering mechanism in other sensory novelty-detection tasks such as dynamic predictive coding of unexpected visual information by the retina [180], or more complex selective attention tasks such as selective visual attention [181][182], deserves further study.
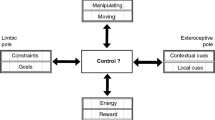
6.2 A sensory firewall for Cartesian mind-body internal model adaptation
The array of low-pass, high-pass and band-pass/band-stop frequency filtering effects of nonassociative learning, together with the associated Boolean logic temporal filtering effects of nonassociative gating, provide a finely-tuned intelligent "firewall" that continuously screens all incoming signals into actionable and non-actionable categories in order to prioritize (Fig. 7). This firewall mechanism shields the mind from the vast amounts of inundating sensory information that constantly compete with one another for attention, and spares it the trouble of having to respond to every tingling except the most salient ones. The triage process not only helps to preserve mental sanity but also conserve physical energy, both of which are important for survival. On the other hand, breakdown of the nonassociative learning processes may result in mistuning of the firewall and hence, distortions in the sensory percept (much like the frequency distortions heard in an audio system with unequal tone control). Nonassociative learning therefore plays an important role in balancing the sensory inputs. This could be the first step in the brain's putative ability of creating explicit internal models of the environment [183][184], as implicit in René Descartes' mind-body interactionism [185].

Nonassociative learning as a sensory firewall that constantly screens all environmental inputs and decides their saliency and priority for internal model adaptation in mind-body interaction. This high-pass, low-pass or band-pass/band-stop signal filtering action is analogous to tone control in an audio system.
7. Conclusion
A general theory of nonassociative learning comprised of habituation, sensitization and desensitization in primary or secondary pathways has been presented. The defining phenotypes of these varying modes of nonassociative learning are their distinct integral-differential computation capabilities, which are shown to correlate with many intelligent or maladaptive brain behaviors. In addition, the notion of nonassociative gating with intrinsic Boolean logic computation capability has been introduced as a basic behavioral paradigm that may act independently or in tandem with nonassociative learning. Together, nonassociative learning and nonassociative gating constitute an intelligent firewall that constantly triages vast amounts of sensory information into actionable and non-actionable categories in order to prioritize. This unified framework of nonassociative learning and nonassociative gating sheds new lights on the ultra secrets of brain intelligence and brain disorders. The underlying functional and structural organization principles [186] are shown to be generally applicable to a wide variety of brain systems across animal phyla and sensory modalities in health and in disease states. These system-level principles are fundamental to a systems medicine approach [187][188] to the management of human health and disease at the organ, organism and community level.
APPENDIX I. Integrator and differentiator equations
A first-order leaky integrator or differentiator (Fig. 5) is described by the following equations:
= -ax + bu (1a)
y = cx + du (1b)
where y, u are the output and input of the integrator or differentiator, respectively; x and are state variable and its rate of change in time; a, b, c, d are parameters and a > 0. The terms cx and du indicate respectively the indirect (adaptive/dynamic) effect and direct (feedforward reflex) effects of the input on the output. For a constant-step input, u ≡ constant for 0 <t <T where T is the end of input, the solution for Eq. 1 under zero initial condition for x is:
y(t) = du + (cbu/a)(1 - e-at)for 0 <t ≤ T (2)
This model represents an integrator (Fig. 5A, left panel) if both terms on the right hand side of Eq. 2 have the same sign, or a differentiator (Fig. 5A, right panel) if they have opposite signs. An integrator or differentiator is said to be inverted (with negative gain) if the input exerts an opposite direct effect on the output, i.e., d < 0. It is called biphasic or monophasic depending on the presence or absence of a post-stimulus response (for t >T):
Biphasic: y(t) = [y(T) - y(0)]e-a(t-T)for t >T (3a)
Monophasic: y = 0 for t >T (3b)
where y(0), y(T) are respectively the outputs at the beginning and end of the step input.
From linear systems theory [145], the equivalent transfer function for the model of Eq. 1 is (Fig. 5B):
where s is the complex frequency and Y, U are the Laplace transforms of y, u, respectively.
APPENDIX II. Nonassociative-learning integrator/differentiator models
From Eq. 2 the response of a primary integrator or differentiator (Fig. 6) to a step input u1 applied to the primary pathway is given by:
where the subscript '1' indicates attribute to the primary pathway. Because of the input-gating effect (Sect. 4.4.1) the primary pathway is silenced once the primary input is off (for t >T).
A secondary integrator or differentiator is described by the following model equation:
2 = -a2x2 + b2u2 + b3u1
y2 = c2x2 + d2u2 (6)
where the subscript '2' indicates attribute to the secondary pathway and b3 represents the influence of the primary stimulus on the secondary pathway. This model represents an integrator or differentiator if b2 and b3 have the same or opposite signs, respectively. Assuming a tonic bias input u2 in the secondary pathway and a step input u1 in the primary pathway, the response of the above secondary integrator or differentiator is:
and
Finally, the resultant response of the dual-process integrator and/or differentiator is the sum of y1 and y2:
y = y1 + y2 (8)
References
McCormick DA: Brain calculus: neural integration and persistent activity. Nat Neurosci. 2001, 4 (2): 113-114. 10.1038/83917.
Poon C-S, Siniaia MS: Plasticity of cardiorespiratory neural processing: Classification and computational functions. Respir Physiol. 2000, 122 (2-3): 83-109. 10.1016/S0034-5687(00)00152-3.
Siniaia MS, Young DL, Poon C-S: Habituation and desensitization of the Hering-Breuer reflex in rat. J Physiol (Lond). 2000, 523 (2): 479-491. 10.1111/j.1469-7793.2000.t01-1-00479.x.
Chernigovsky VN, Mysiashikova SS, Siniaia MS, Mokrushin AA: Habituation in Visceral Systems. 1980, Leningrad, Russia: Nayka
Christoffersen GR: Habituation: events in the history of its characterization and linkage to synaptic depression. A new proposed kinetic criterion for its identification. Prog Neurobiol. 1997, 53 (1): 45-66. 10.1016/S0301-0082(97)00031-2.
Kandel ER: A Cell-Biological Approach to Learning. 1978, Bethesda, MD: Society for Neuroscience
Siniaia MS, Silakov VL: Plasticity of the Visceral Analyzer. 1990, Leningrad, Russia: Nayka;
Peeke HVS, Petrinovich L, (Eds): Habituation, Sensitization, and Behavior. 1984, Orlando, Fl: Academic Press
Thompson RF, Groves PM, Teyler TJ, Roemer RA: A dual-process theory of habituation: theory and behavior. Habituation. Edited by: Peeke HVS, Herz MJ. 1973, New York: Academic Press, 1: 239-271.
Groves PM, Thompson RF: Habituation: a dual-process theory. Psychol Rev. 1970, 77 (5): 419-450. 10.1037/h0029810.
Groves PM, Thompson RF: A dual-process theory of habituation: neural mechanisms. Habituation. Edited by: Peeke HVS, Herz MJ. 1973, New York: Academic Press, 2: 175-205.
Scholz KP, Byrne JH: Long-term sensitization in Aplysia: biophysical correlates in tail sensory neurons. Science. 1987, 235 (4789): 685-687.
Castellucci VF, Carew TJ, Kandel ER: Cellular analysis of long-term habituation of the gill-withdrawal reflex of Aplysia californica. Science. 1978, 202 (4374): 1306-1308.
Bailey CH, Chen M: Morphological basis of long-term habituation and sensitization in Aplysia. Science. 1983, 220 (4592): 91-93.
Leaton RN, Supple WF: Cerebellar vermis: essential for long-term habituation of the acoustic startle response. Science. 1986, 232 (4749): 513-515.
Stopfer M, Chen X, Tai U-T, Huang GS, Carew TJ: Site specificity of short-term and long-term habituation in the tail-elicited siphon withdrawal reflex in Aplysia. J Neurosci. 1996, 16: 4923-4932.
Hine B, Paolino RM: Increases in heart rate accompanying decreases in activity and defecation: support for a dual process theory of habituation. Behav Biol. 1972, 7 (3): 427-433. 10.1016/S0091-6773(72)80115-9.
Edwards JA, Siddle DA: Dishabituation of the electrodermal orienting response following decay of sensitization. Biol Psychol. 1976, 4 (1): 19-28. 10.1016/0301-0511(76)90027-2.
Harding GB, Hagman JD: Stimulus intensity effects in electrodermal habituation. J Exp Psychol [Hum Learn]. 1977, 3 (5): 600-607. 10.1037/0278-7393.3.5.600.
Wester K, Sagvolden T: Habituation of the orienting response: experimental evidence in favor of a "dual-process" theory. Behav Neural Biol. 1979, 26 (4): 479-484. 10.1016/S0163-1047(79)91518-8.
Megela AL, Teyler TJ: Habituation and the human evoked potential. J Comp Physiol Psychol. 1979, 93 (6): 1154-1170. 10.1037/h0077630.
Peeke HV: Habituation, sensitization, and redirection of aggression and feeding behavior in the three-spined stickleback (Gasterosteus aculeatus L.). J Comp Psychol. 1983, 97 (1): 43-51. 10.1037/0735-7036.97.1.43.
Kaplan PS, Werner JS: Habituation, response to novelty, and dishabituation in human infants: tests of a dual-process theory of visual attention. J Exp Child Psychol. 1986, 42 (2): 199-217. 10.1016/0022-0965(86)90023-8.
Pitman DL, Ottenweller JE, Natelson BH: Effect of stressor intensity on habituation and sensitization of glucocorticoid responses in rats. Behav Neurosci. 1990, 104 (1): 28-36. 10.1037/0735-7044.104.1.28.
Plappert CF, Pilz PK, Schnitzler HU: Interaction between acoustic and electric sensitization of the acoustic startle response in rats. Behav Brain Res. 1999, 103 (2): 195-201. 10.1016/S0166-4328(99)00034-0.
Plappert CF, Pilz PK, Becker K, Becker CM, Schnitzler HU: Increased sensitization of acoustic startle response in spasmodic mice with a mutation of the glycine receptor alpha1-subunit gene. Behav Brain Res. 2001, 121 (1–2): 57-67. 10.1016/S0166-4328(00)00385-5.
Prescott SA, Chase R: Sites of plasticity in the neural circuit mediating tentacle withdrawal in the snail Helix aspersa: implications for behavioral change and learning kinetics. Learn Mem. 1999, 6 (4): 363-380.
Pilz PK, Schnitzler HU: Habituation and sensitization of the acoustic startle response in rats: amplitude, threshold, and latency measures. Neurobiol Learn Mem. 1996, 66 (1): 67-79. 10.1006/nlme.1996.0044.
Bee MA: Habituation and sensitization of aggression in bullfrogs (Rana catesbeiana): testing the dual-process theory of habituation. J Comp Psychol. 2001, 115 (3): 307-316. 10.1037/0735-7036.115.3.307.
Bashinski HS, Werner JS, Rudy JW: Determinants of infant visual fixation: evidence for a two-process theory. J Exp Child Psychol. 1985, 39 (3): 580-598. 10.1016/0022-0965(85)90058-X.
Prescott SA: Interactions between depression and facilitation within neural networks: updating the dual-process theory of plasticity. Learn Mem. 1998, 5 (6): 446-466.
Thompson RF, Spencer WA: Habituation: a model phenomenon for the study of neuronal substrates of behavior. Psychol Rev. 1966, 73 (1): 16-43. 10.1037/h0022681.
Rose JK, Rankin CH: Analyses of habituation in Caenorhabditis elegans. Learn Mem. 2001, 8 (2): 63-69. 10.1101/lm.37801.
Teyler TJ, Alger BE: Monosynaptic habituation in the vertebrate forebrain: the dentate gyrus examined in vitro. Brain Res. 1976, 115 (3): 413-425. 10.1016/0006-8993(76)90358-9.
Wiel DE, Wood ER, Weeks JC: Habituation of the prolegwithdrawal reflex in Manduca sexta does not involve changes in motoneuron properties or depression at the sensorimotor synapse. Neurobiol Learn Mem. 2001, 76 (1): 57-80. 10.1006/nlme.2000.3982.
Burrell BD, Sahley CL, Muller KJ: Non-associative learning and serotonin induce similar bi-directional changes in excitability of a neuron critical for learning in the medicinal leech. J Neurosci. 2001, 21 (4): 1401-1412.
Davis M, File SE: Intrinsic and extrinsic mechanisms of habituation and sensitization: implications for the design and analysis of experiments. Habituation, Sensitization, and Behavior. Edited by: Peeke HVS, Petrinovich LF. 1984, New York: Academic Press, 287-323.
Farel PB: Dual processes control response habituation across a single synapse. Brain Res. 1974, 72 (2): 323-327. 10.1016/0006-8993(74)90874-9.
Walters ET, Illich PA, Weeks JC, Lewin MR: Defensive responses of larval Manduca sexta and their sensitization by noxious stimuli in the laboratory and field. J Exp Biol. 2001, 204 (Pt 3): 457-469.
Brown GD: Nonassociative learning processes affecting swimming probability in the seaslug Tritonia diomedea: habituation, sensitization and inhibition. Behav Brain Res. 1998, 95 (2): 151-165. 10.1016/S0166-4328(98)00072-2.
Brown GD, Frost WN, Getting PA: Habituation and iterative enhancement of multiple components of the Tritonia swim response. Behav Neurosci. 1996, 110 (3): 478-485. 10.1037/0735-7044.110.3.478.
Lockery SR, Kristan WB: Two forms of sensitization of the local bending reflex of the medicinal leech. J Comp Physiol [A]. 1991, 168 (2): 165-177.
Baranauskas G, Nistri A: Sensitization of pain pathways in the spinal cord: cellular mechanisms. Prog Neurobiol. 1998, 54: 349-365. 10.1016/S0301-0082(97)00067-1.
Woolf CJ: Windup and central sensitization are not equivalent. Pain. 1996, 66: 105-108. 10.1016/0304-3959(96)03114-4.
Willis WD: Role of neurotransmitters in sensitization of pain responses. Ann NY Acad Sci. 2001, 933: 142-156.
Melzack R, Coderre TJ, Katz J, Vaccarino AL: Central neuroplasticity and pathological pain. Ann N Y Acad Sci. 2001, 933: 157-174.
Green BG: Capsaicin sensitization and desensitization on the tongue produced by brief exposures to a low concentration. Neurosci Lett. 1989, 107 (1–3): 173-178. 10.1016/0304-3940(89)90812-4.
Burrell BD, Sahley CL: Generalization of habituation and intrinsic sensitization in the leech. Learn Mem. 1998, 5 (6): 405-419.
Prescott SA, Chase R: Two types of plasticity in the tentacle withdrawal reflex of Helix aspersa are dissociated by tissue location and response measure. J Comp Physiol A. 1996, 179: 407-414. 10.1007/BF00194994.
Castellucci VF, Kandel ER: Presynaptic facilitation as a mechanism for behavioral sensitization in Aplysia. Science. 1976, 194: 1176-1178.
Cohen TE, Kaplan SW, Kandel ER, Hawkins RD: A simplified preparation for relating cellular events to behavior: mechanisms contributing to habituation, dishabituation, and sensitization of the Aplysia gill-withdrawal reflex. J Neurosci. 1997, 17: 2886-2899.
Kennedy TE, Kuhl D, Barzilai A, Sweatt JD, Kandel ER: Long-term sensitization training in Aplysia leads to an increase in calreticulin, a major presynaptic calcium-binding protein. Neuron. 1992, 9 (6): 1013-1024. 10.1016/0896-6273(92)90062-I.
Bailey CH, Chen M: Long-term sensitization in Aplysia increases the number of presynaptic contacts onto the identified gill motor neuron L7. Proc Natl Acad Sci USA. 1988, 85 (23): 9356-9359. 10.1073/pnas.85.23.9356.
Burrell BD, Sahley CL: Serotonin depletion does not prevent intrinsic sensitization in the leech. Learn Mem. 1999, 6 (5): 509-520. 10.1101/lm.6.5.509.
Stopfer M, Carew TJ: Heterosynaptic facilitation of tail sensory neuron synaptic transmission during habituation in tail-induced tail and siphon withdrawal reflexes of Aplysia. J Neurosci. 1996, 16 (16): 4933-4948.
Craig AD: Pain mechanisms: labeled lines versus convergence in central processing. Annu Rev Neurosci. 2003, 26: 1-30. 10.1146/annurev.neuro.26.041002.131022.
Harvey RJ, Depner UB, Wassle H, Ahmadi S, Heindl C, Reinold H, Smart TG, Harvey K, Schutz B, Abo-Salem OM: GlyR alpha3: an essential target for spinal PGE2-mediated inflammatory pain sensitization. Science. 2004, 304 (5672): 884-887. 10.1126/science.1094925.
Hains BC, Klein JP, Saab CY, Craner MJ, Black JA, Waxman SG: Upregulation of sodium channel Nav1.3 and functional involvement in neuronal hyperexcitability associated with central neuropathic pain after spinal cord injury. J Neurosci. 2003, 23 (26): 8881-8892.
Ji RR, Kohno T, Moore KA, Woolf CJ: Central sensitization and LTP: do pain and memory share similar mechanisms?. Trends Neurosci. 2003, 26 (12): 696-705. 10.1016/j.tins.2003.09.017.
Sandkuhler J: Learning and memory in pain pathways. Pain. 2000, 88 (2): 113-118. 10.1016/S0304-3959(00)00424-3.
Coderre TJ, Katz J, Vaccarino AL, Melzack R: Contribution of central neuroplasticity to pathological pain: review of clinical and experimental evidence. Pain. 1993, 52 (3): 259-285. 10.1016/0304-3959(93)90161-H.
Ito S, Okuda-Ashitaka E, Imanishi T, Minami T: Central roles of nociceptin/orphanin FQ and nocistatin: allodynia as a model of neural plasticity. Prog Brain Res. 2000, 129: 205-218.
Woolf CJ, Salter MW: Neuronal plasticity: increasing the gain in pain. Science. 2000, 288 (5472): 1765-1769. 10.1126/science.288.5472.1765.
Cervero F, Laird JM, Garcia-Nicas E: Secondary hyperalgesia and presynaptic inhibition: an update. Eur J Pain. 2003, 7 (4): 345-351. 10.1016/S1090-3801(03)00047-8.
Milne RJ, Kay NE, Irwin RJ: Habituation to repeated painful and non-painful cutaneous stimuli: a quantitative psychophysical study. Exp Brain Res. 1991, 87 (2): 438-444. 10.1007/BF00231861.
Davis M, Parisi T, Gendelman DS, Tischler M, Kehne JH: Habituation and sensitization of startle reflexes elicited electrically from the brainstem. Science. 1982, 218 (4573): 688-690.
Marcus EA, Nolen TG, Rankin CH, Carew TJ: Behavioral dissociation of dishabituation, sensitization, and inhibition in Aplysia. Science. 1988, 241 (4862): 210-213.
Hochner B, Klein M, Schacher S, Kandel ER: Additional component in the cellular mechanism of presynaptic facilitation contributes to behavioral dishabituation in Aplysia. Proc Natl Acad Sci USA. 1986, 83 (22): 8794-8798. 10.1073/pnas.83.22.8794.
Bristol AS, Marinesco S, Carew TJ: Neural circuit of tail-elicited siphon withdrawal in Aplysia. II. Role of gated inhibition in differential lateralization of sensitization and dishabituation. J Neurophysiol. 2004, 91 (2): 678-692. 10.1152/jn.00667.2003.
Hawkins RD, Cohen TE, Greene W, Kandel ER: Relationships between dishabituation, sensitization, and inhibition of the gill-and siphon-withdrawal reflex in Aplysia californica: effects of response measure, test time, and training stimulus. Behav Neurosci. 1998, 112 (1): 24-38. 10.1037/0735-7044.112.1.24.
Bristol AS, Sutton MA, Carew TJ: Neural circuit of tail-elicited siphon withdrawal in Aplysia. I. Differential lateralization of sensitization and dishabituation. J Neurophysiol. 2004, 91 (2): 666-677. 10.1152/jn.00666.2003.
Krasne FB, Teshiba TM: Habituation of an invertebrate escape reflex due to modulation by higher centers rather than local events. Proc Natl Acad Sci USA. 1995, 92: 3362-3366. 10.1073/pnas.92.8.3362.
Song G, Poon C-S: Functional and structural models of pontine modulation of mechanoreceptor and chemoreceptor reflexes. Respir Physiol Neurobiol. 2004, 143 (2–3): 281-292. 10.1016/j.resp.2004.05.009.
Poon C-S: Organization of central pathways mediating the Hering-Breuer reflex and carotid chemoreflex. Adv Exp Med Biol. 2004, 551: 95-100.
Glanzman DL, Groves PM, Thompson RF: Stimulus generalization of habituation in spinal interneurons. Physiol Behav. 1972, 8: 155-158. 10.1016/0031-9384(72)90145-X.
Goldberg JI, Lukowiak K: Transfer of habituation between stimulation sites of the siphon withdrawal refelx in Aplysia californica. Can J Physiol Pharmacol. 1983, 61: 749-755.
Goldberg JI, Lukowiak K: Transfer of habituation in Aplysia: contribution of heterosynaptic pathways in habituation of the gill-withdrwal reflex. J Neurobiol. 1984, 15: 395-411. 10.1002/neu.480150602.
Montarolo PG, Kandel ER, Schacher S: Long-term heterosynaptic inhibition in Aplysia. Nature. 1988, 333: 171-174. 10.1038/333171a0.
Leonard JL, Edstrom JP: Parallel processing in an identified neural circuit: the Aplysia californica gill-withdrawal response model system. Biol Rev Camb Philos Soc. 2004, 79 (1): 1-59. 10.1017/S1464793103006183.
Frost WN, Brown GD, Getting PA: Parametric features of habituation of swim cycle number in the marine mollusc tritonia diomedea. Neurobiol Learn Mem. 1996, 65 (2): 125-134. 10.1006/nlme.1996.0015.
Ramachandran VS, Rogers-Ramachandran D: Phantom limbs and neural plasticity. Arch Neurol. 2000, 57 (3): 317-320. 10.1001/archneur.57.3.317.
Flor H: Phantom-limb pain: characteristics, causes, and treatment. Lancet Neurol. 2002, 1: 182-189. 10.1016/S1474-4422(02)00074-1.
Moore CI, Stern CE, Dunbar C, Kostyk SK, Gehi A, Corkin S: Referred phantom sensations and cortical reorganization after spinal cord injury in humans. Proc Natl Acad Sci USA. 2000, 97: 14703-14708. 10.1073/pnas.250348997.
Davis KD, Kiss ZHT, Luo L, Tasker RR, Lozano AM, Dostrovsky JO: Phantom sensations generated by thalamic microstimulation. Nature. 1998, 391: 385-387. 10.1038/34905.
Wall PD: The presence of ineffective synapses and the circumstances which unmask them. Philos Trans R Soc Lond B Biol Sci. 1977, 278: 361-372.
Ramachandran VS: Behavioral and magnetoencephalographic correlates of plasticity in the adult human brain. Proc Natl Acad Sci USA. 1993, 90: 10413-10420. 10.1073/pnas.90.22.10413.
Halligan PW, Marshall JC, Wade DT, Davey J, Morrison D: Thumb in cheek? Sensory reorganization and perceptual plasticity after limb amputation. NeuroReport. 1993, 4 (3): 233-236.
Kew JJM, Ridding MC, Rothwell JC, Passingham RE, Leigh PN, Sooriakumaran S, Frackowiak RSJ, Brooks DJ: Reorganization of cortical blood flow and transcranial magnetic stimulation maps in human subjects after upper limb amputation. J Neurophysiol. 1994, 72: 2517-2524.
Kaas JH: Plasticity of sensory and motor maps in adult mammals. Annu Rev Neurosci. 1991, 14: 137-167. 10.1146/annurev.ne.14.030191.001033.
Faggin BM, Nguyen KT, Nicolelis MA: Immediate and simultaneous sensory reorganization at cortical and subcortical levels of the somatosensory system. Proc Natl Acad Sci USA. 1997, 94: 9428-9433. 10.1073/pnas.94.17.9428.
Pettit MJ, Schwark HD: Receptive field reorganization in dorsal column nuclei during temporary denervation. Science. 1993, 262: 2054-2056.
Chen R, Cohen LG, Hallett M: Nervous system reorganization following injury. Neuroscience. 2002, 111 (4): 761-773. 10.1016/S0306-4522(02)00025-8.
Borsook D, Becerra L, Fishman S, Edwards A, Jennings CL, Stojanovic M, Papinicolas L, Ramachandran VS, Gonzalex RG, Breiter H: Acute plasticity in the human somatosensory cortex following amputation. NeuroReport. 1998, 9: 1013-1017.
Aglioti S, Cortese F, Franchini C: Rapid sensory remapping in the adult human brain as inferred from phantom breast perception. NeuroReport. 1994, 5: 473-476.
Hess G, Donoghue JP: Long-term potentiation of horizontal connections provides a mechanism to reorganize cortical motor maps. J Neurophysiol. 1994, 71: 2543-2547.
Dinse HR, Ragert P, Pleger B, Schwenkreis P, Tegenthoff M: Pharmacological modulation of perceptual learning and associated cortical reorganization. Science. 2003, 301: 91-94. 10.1126/science.1085423.
Heynen AJ, Yoon B-Y, Liu C-H, Chung HJ, Huganir RL, Bear MF: Molecular mechanism for loss of visual cortical responsiveness following brief monocular deprivation. Nat Neurosci. 2003, 6: 854-862. 10.1038/nn1100.
Rittenhouse CD, Shouval HZ, Paradiso MA, Bear MF: Monocular deprivation induces homosynaptic long-term depression in visual cortex. Nature. 1999, 397: 347-350. 10.1038/16922.
Dessirier JM, Simons CT, Sudo M, Sudo S, Carstens E: Sensitization, desensitization and stimulus-induced recovery of trigeminal neuronal responses to oral capsaicin and nicotine. J Neurophysiol. 2000, 84 (4): 1851-1862.
Szallasi A, Blumberg PM: Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol Rev. 1999, 51 (2): 159-212.
Cliff MA, Green BG: Sensory irritation and coolness produced by menthol: evidence for selective desensitization of irritation. Physiol Behav. 1994, 56 (5): 1021-1029. 10.1016/0031-9384(94)90338-7.
Prescott J, Stevenson RJ: Desensitization to oral zingerone irritation: effects of stimulus parameters. Physiol Behav. 1996, 60 (6): 1473-1480. 10.1016/S0031-9384(96)00248-X.
Dessirier JM, O'Mahony M, Carstens E: Oral irritant properties of menthol: sensitizing and desensitizing effects of repeated application and cross-desensitization to nicotine. Physiol Behav. 2001, 73 (1–2): 25-36. 10.1016/S0031-9384(01)00431-0.
Simons CT, Carstens MI, Carstens E: Oral irritation by mustard oil: self-desensitization and cross-desensitization with capsaicin. Chem Senses. 2003, 28 (6): 459-465. 10.1093/chemse/28.6.459.
Gottrup H, Kristensen AD, Flemming WB, Troels SJ: Aftersensations in experimental and clinical hypersensitivity. Pain. 2003, 103: 57-64. 10.1016/S0304-3959(02)00415-3.
Stewart J, Badiani A: Tolerance and sensitization to the behavioral effects of drugs. Behav Pharmacol. 1993, 4 (4): 289-312.
Baker TB, Tiffany ST: Morphine tolerance as habituation. Psychol Rev. 1985, 92 (1): 78-108. 10.1037/0033-295X.92.1.78.
McSweeney FK, Murphy ES, Kowal BP: Regulation of drug taking by sensitization and habituation. Exp Clin Psychopharmacol. 2005, 13 (3): 163-184. 10.1037/1064-1297.13.3.163.
Paletta MS, Wagner AR: Development of context-specific tolerance to morphine: support for a dual-process interpretation. Behav Neurosci. 1986, 100 (5): 611-623. 10.1037/0735-7044.100.5.611.
Drummond DC, Litten RZ, Lowman C, Hunt WA: Craving research: future directions. Addiction. 2000, 95 (Suppl 2): S247-255. 10.1080/09652140050111816.
Addolorato G, Leggio L, Abenavoli L, Gasbarrini G: Neurobiochemical and clinical aspects of craving in alcohol addiction: a review. Addict Behav. 2005, 30 (6): 1209-1224. 10.1016/j.addbeh.2004.12.011.
Anton RF: What is craving? Models and implications for treatment. Alcohol Res Health. 1999, 23 (3): 165-173.
Lowman C, Hunt WA, Litten RZ, Drummond DC: Research perspectives on alcohol craving: an overview. Addiction. 2000, 95 (Suppl 2): S45-54. 10.1080/09652140050111636.
Vanderschuren LJ, Kalivas PW: Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: a critical review of preclinical studies. Psychopharmacology (Berl). 2000, 151 (2–3): 99-120. 10.1007/s002130000493.
Vezina P: Sensitization of midbrain dopamine neuron reactivity and the self-administration of psychomotor stimulant drugs. Neurosci Biobehav Rev. 2004, 27 (8): 827-839. 10.1016/j.neubiorev.2003.11.001.
Brebner K, Wong TP, Liu L, Liu Y, Campsall P, Gray S, Phelps L, Phillips AG, Wang YT: Nucleus accumbens long-term depression and the expression of behavioral sensitization. Science. 2005, 310 (5752): 1340-1343. 10.1126/science.1116894.
DiFranza JR, Wellman RJ: A sensitization-homeostasis model of nicotine craving, withdrawal, and tolerance: integrating the clinical and basic science literature. Nicotine Tob Res. 2005, 7 (1): 9-26. 10.1080/14622200412331328538.
Robinson TE, Berridge KC: Addiction. Annu Rev Psychol. 2003, 54: 25-53. 10.1146/annurev.psych.54.101601.145237.
Grace AA: The tonic/phasic model of dopamine system regulation and its implications for understanding alcohol and psychostimulant craving. Addiction. 2000, 95 (Suppl 2): S119-128. 10.1080/09652140050111690.
Shaham Y, Hope BT: The role of neuroadaptations in relapse to drug seeking. Nat Neurosci. 2005, 8 (11): 1437-1439. 10.1038/nn1105-1437.
Bossert JM, Ghitza UE, Lu L, Epstein DH, Shaham Y: Neurobiology of relapse to heroin and cocaine seeking: An update and clinical implications. Eur J Pharmacol. 2005, 526 (1–3): 36-50. 10.1016/j.ejphar.2005.09.030.
Bancroft A, Wiltshire S, Parry O, Amos A: "It's like an addiction first thing...afterwards it's like a habit": daily smoking behaviour among people living in areas of deprivation. Soc Sci Med. 2003, 56 (6): 1261-1267. 10.1016/S0277-9536(02)00124-7.
Gerdeman GL, Partridge JG, Lupica CR, Lovinger DM: It could be habit forming: drugs of abuse and striatal synaptic plasticity. Trends Neurosci. 2003, 26 (4): 184-192. 10.1016/S0166-2236(03)00065-1.
Everitt BJ, Robbins TW: Neural systems of reinforcement for drug addiction: from actions to habits to compulsion. Nat Neurosci. 2005, 8 (11): 1481-1489. 10.1038/nn1579.
Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM: Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature. 2005, 437 (7062): 1158-1161. 10.1038/nature04053.
Hnasko TS, Sotak BN, Palmiter RD: Morphine reward in dopamine-deficient mice. Nature. 2005, 438 (7069): 854-857. 10.1038/nature04172.
Sora I, Wichems C, Takahashi N, Li XF, Zeng Z, Revay R, Lesch KP, Murphy DL, Uhl GR: Cocaine reward models: conditioned place preference can be established in dopamine-and in serotonin-transporter knockout mice. Proc Natl Acad Sci USA. 1998, 95 (13): 7699-7704. 10.1073/pnas.95.13.7699.
Hyman SE: Addiction: a disease of learning and memory. Am J Psychiatry. 2005, 162 (8): 1414-1422. 10.1176/appi.ajp.162.8.1414.
Wilson SJ, Sayette MA, Fiez JA: Prefrontal responses to drug cues: a neurocognitive analysis. Nat Neurosci. 2004, 7 (3): 211-214. 10.1038/nn1200.
Kandel ER, Abrams T, Bernier L, Carew TJ, Hawkins RD, Schwartz JH: Classical conditioning and sensitization share aspects of the same molecular cascade in Aplysia. Cold Spring Harb Symp Quant Biol. 1983, 48 (Pt 2): 821-830.
Hawkins RD: A cellular mechanism of classical conditioning in Aplysia. J Exp Biol. 1984, 112: 113-128.
Hawkins RD, Greene W, Kandel ER: Classical conditioning, differential conditioning, and second-order conditioning of the Aplysia gill-withdrawal reflex in a simplified mantle organ preparation. Behav Neurosci. 1998, 112 (3): 636-645. 10.1037/0735-7044.112.3.636.
Rankin CH: Context conditioning in habituation in the nematode Caenorhabditis elegans. Behav Neurosci. 2000, 114 (3): 496-505. 10.1037/0735-7044.114.3.496.
Hermitte G, Pedreira ME, Tomsic D, Maldonado H: Context shift and protein synthesis inhibition disrupt long-term habituation after spaced,but not massed, training in the crab Chasmagnathus. Neurobiol Learn Mem. 1999, 71 (1): 34-49. 10.1006/nlme.1998.3858.
Tirelli E, Laviola G, Adriani W: Ontogenesis of behavioral sensitization and conditioned place preference induced by psychostimulants in laboratory rodents. Neurosci Biobehav Rev. 2003, 27 (1–2): 163-178. 10.1016/S0149-7634(03)00018-6.
Dworkin BR: Learning and Physiological Regulation. 1993, Chicago: University of Chicago Press
Sokolowska M, Siegel S, Kim JA: Intraadministration associations: conditional hyperalgesia elicited by morphine onset cues. J Exp Psychol Anim Behav Process. 2002, 28 (3): 309-320. 10.1037/0097-7403.28.3.309.
Young DL, Eldridge FL, Poon CS: Integration-differentiation and gating of carotid afferent traffic that shapes the respiratory pattern. J Appl Physiol. 2003, 94 (3): 1213-1229.
Poon C-S, Siniaia MS, Young DL, Eldridge FL: Short-term potentiation of carotid chemoreflex: An NMDAR-dependent neural integrator. NeuroReport. 1999, 10: 2261-2265.
Poon C-S, Young DL, Siniaia MS: High-pass filtering of carotid-vagal influences on expiration in rat: role of N-methyl-D-aspartate receptors. Neurosci Lett. 2000, 284: 5-8. 10.1016/S0304-3940(00)00993-9.
Pinsker H, Kupfermann I, Castellucci V, Kandel E: Habituation and dishabituation of the gill-withdrawal reflex in Aplysia. Science. 1970, 167 (926): 1740-1742.
Dragoi V: A feedforward model of suppressive and facilitatory habituation effects. Biol Cybern. 2002, 86 (6): 419-426. 10.1007/s00422-001-0306-x.
Staddon JE, Higa JJ: Multiple time scales in simple habituation. Psychol Rev. 1996, 103 (4): 720-733. 10.1037/0033-295X.103.4.720.
Shen L: Neural integration by short term potentiation. Biol Cybern. 1989, 61 (4): 319-325. 10.1007/BF00203180.
Ogata K: Modern Control Engineering. 1997, Upper Saddle River, N.J.: Prentice Hall, 3
Eldridge FL, Millhorn DE: Oscillation, gating, and memory in the respiratory control system. Handbook of Physiology, Section 3: The Respiratory System, Vol II: Control of Breathing, part 1. Edited by: Fishman AP, Cherniack NS, Widdicombe JG. 1986, Bethesda, MD: American Physiological Society, 93-114.
Arnold DB, Robinson DA: The oculomotor integrator: testing of a neural network model. Exp Brain Res. 1997, 113 (1): 57-74. 10.1007/BF02454142.
Seung HS, Lee DD, Reis BY, Tank DW: Stability of the memory of eye position in a recurrent network of conductance-based model neurons. Neuron. 2000, 26: 259-271. 10.1016/S0896-6273(00)81155-1.
Rosen MJ: A theoretical neural integrator. IEEE Trans Biomed Eng. 1972, 19 (5): 362-367.
Aksay E, Gamkrelidze G, Seung HS, Baker R, Tank DW: In vivo intracellular recording and perturbation of persistent activity in a neural integrator. Nat Neurosci. 2001, 4 (2): 184-193. 10.1038/84023.
Koulakov AA, Raghavachari S, Kepecs A, Lisman JE: Model for a robust neural integrator. Nat Neurosci. 2002, 5 (8): 775-782. 10.1038/nn893.
Goldman MS, Levine JH, Major G, Tank DW, Seung HS: Robust persistent neural activity in a model integrator with multiple hysteretic dendrites per neuron. Cereb Cortex. 2003, 13 (11): 1185-1195. 10.1093/cercor/bhg095.
Lau PM, Bi GQ: Synaptic mechanisms of persistent reverberatory activity in neuronal networks. Proc Natl Acad Sci USA. 2005, 102 (29): 10333-10338. 10.1073/pnas.0500717102.
Major G, Baker R, Aksay E, Mensh B, Seung HS, Tank DW: Plasticity and tuning by visual feedback of the stability of a neural integrator. Proc Natl Acad Sci USA. 2004, 101 (20): 7739-7744. 10.1073/pnas.0401970101.
Egorov AV, Hamam BN, Fransen E, Hasselmo ME, Alonso AA: Graded persistent activity in entorhinal cortex neurons. Nature. 2002, 420 (6912): 173-178. 10.1038/nature01171.
Loewenstein Y, Mahon S, Chadderton P, Kitamura K, Sompolinsky H, Yarom Y, Hausser M: Bistability of cerebellar Purkinje cells modulated by sensory stimulation. Nat Neurosci. 2005, 8 (2): 202-211. 10.1038/nn1393.
Loewenstein Y, Sompolinsky H: Temporal integration by calcium dynamics in a model neuron. Nat Neurosci. 2003, 6 (9): 961-967. 10.1038/nn1109.
Gingrich KJ, Baxter DA, Byrne JH: Mathematical model of cellular mechanisms contributing to presynaptic facilitation. Brain Res Bull. 1988, 21 (3): 513-520. 10.1016/0361-9230(88)90167-0.
Wang L, Ross J: Variable threshold as a model for selective attention, (de)sensitization, and anesthesia in associative neural networks. Biol Cybern. 1991, 64 (3): 231-241. 10.1007/BF00201984.
Zhou Z, Champagnat J, Poon C-S: Phasic and long-term depression in brainstem nucleus tractus solitarius neurons: differing roles of AMPA receptor desensitization. J Neurosci. 1997, 17 (14): 5349-5356.
Zhou Z, Poon C-S: Field potential analysis of synaptic transmission in spiking neurons in a sparse and irregular neuronal structure in vitro. J Neurosci Methods. 2000, 94 (2): 193-203. 10.1016/S0165-0270(99)00144-2.
Poon C-S, Zhou Z, Champagnat J: NMDA receptor activity in utero averts respiratory depression and anomalous LTD in newborn mice. J Neurosci. 2000, 20: RC73 (71–76)
Wang XJ: Persistent neural activity: experiments and theory. Cereb Cortex. 2003, 13 (11): 1123-10.1093/cercor/bhg112.
Heller S: Too loud, too bright, too fast, too tight. What to do if you are sensory defensive in an overstimulating world. 2002, New York, NY: HarperCollins
Poulton R, Menzies RG: Non-associative fear acquisition: a review of the evidence from retrospective and longitudinal research. Behav Res Ther. 2002, 40 (2): 127-149. 10.1016/S0005-7967(01)00045-6.
Goldreich D, Krauzlis RJ, Lisberger SG: Effect of changing feedback delay on spontaneous oscillations in smooth pursuit eye movements of monkeys. J Neurophysiol. 1992, 67 (3): 625-638.
Mackey MC, Glass L: Oscillation and chaos in physiological control systems. Science. 1977, 197 (4300): 287-289.
Poon C-S: Synaptic plasticity and respiratory control. Bioengineering Approaches to Pulmonary Physiology and Medicine. Edited by: Khoo MCK. 1996, Los Angles: Plenum, 93-113.
Gonzalez ER, Krieger AJ, Sapru HN: Central resetting of baroreflex in the spontaneously hypertensive rat. Hypertension. 1983, 5 (3): 346-352.
Heesch C, Barron K: Is there a central nervous system component to acute baroreflex resetting in rats?. Am J Physiol. 1992, 262 (2 Pt 2): H503-510.
Young DL, Siniaia MS, Poon C-S: Model of nonassociative learning in vagal-pontine modulation of the respiratory rhythm. Adv Exp Med Biol. 2001, 499: 255-260.
Young DL, Poon C-S: Models of adaptive respiratory control and their clinical implications. BME. 2002, 16 (8): 12-19.
Ohman A, Lader M: Selective attention and "habituation" of the auditory averaged evoked response in humans. Physiol Behav. 1972, 8 (1): 79-85. 10.1016/0031-9384(72)90132-1.
Waters WF, McDonald DG, Koresko RL: Habituation of the orienting response: a gating mechanism subserving selective attention. Psychophysiology. 1977, 14 (3): 228-236.
Lorch EP, Horn DG: Habituation of attention to irrelevant stimuli in elementary school children. J Exp Child Psychol. 1986, 41 (1): 184-197. 10.1016/0022-0965(86)90057-3.
Baranek GT, Berkson G: Tactile defensiveness in children with developmental disabilities: responsiveness and habituation. J Autism Dev Disord. 1994, 24 (4): 457-471. 10.1007/BF02172128.
Perry W, Minassian A, Lopez B, Maron L, Lincoln A: Sensorimotor Gating Deficits in Adults with Autism. Biol Psychiatry. 2006
Kaplan PS, Werner JS: Habituation, response to novelty, and dishabituation in human infants: tests of a dual-process theory of visual attention. J Exp Child Psychol. 1986, 42 (2): 199-217. 10.1016/0022-0965(86)90023-8.
Meincke U, Light GA, Geyer MA, Braff DL, Gouzoulis-Mayfrank E: Sensitization and habituation of the acoustic startle reflex in patients with schizophrenia. Psychiatry Res. 2004, 126 (1): 51-61. 10.1016/j.psychres.2004.01.003.
Hosoya T, Baccus SA, Meister M: Dynamic predictive coding by the retina. Nature. 2005, 436 (7047): 71-77. 10.1038/nature03689.
Desimone R, Duncan J: Neural mechanisms of selective visual attention. Annu Rev Neurosci. 1995, 18: 193-222. 10.1146/annurev.ne.18.030195.001205.
Moore T: The neurobiology of visual attention: finding sources. Curr Opin Neurobiol. 2006, 16 (2): 159-165. 10.1016/j.conb.2006.03.009.
Ito M: Bases and implications of learning in the cerebellum – adaptive control and internal model mechanism. Prog Brain Res. 2005, 148: 95-109. 10.1016/S0079-6123(04)48009-1.
Tin C, Poon CS: Internal models in sensorimotor integration: perspectives from adaptive control theory. J Neural Eng. 2005, 2 (3): S147-163. 10.1088/1741-2560/2/3/S01.
Poon C-S, Merfeld D: Internal models: the state of the art (Editorial for special issue: Sensory Integration, State Estimation, and Motor Control in the Brain: Role of Internal Models). J Neural Eng. 2005, 2 (3): doi:10.1088/1741-2552/2/3/E1001
Young DL, Poon C-S: Soul searching and heart throbbing for biological modeling. Behav Brain Sci. 2001, 24 (6): 1080-1081.
Ahn AC, Tewari M, Poon CS, Phillips RS: The Limits of Reductionism in Medicine: Could Systems Biology Offer an Alternative?. PLoS Med. 2006, 3 (6): e208-10.1371/journal.pmed.0030208.
Ahn AC, Tewari M, Poon CS, Phillips RS: The Clinical Applications of a Systems Approach. PLoS Med. 2006, 3 (7): e209-10.1371/journal.pmed.0030209.
Acknowledgements
We thank Drs. K. Lukowiak, B.R. Dworkin, J.R. DiFranza and members of the Poon Lab for helpful discussions. This work was supported in part by National Institutes of Health grants HL067966 and HL072849. DLY was recipient of a National Institute of Mental Health predoctoral fellowship MH012697.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
CSP conceived the theory, reviewed and integrated the literature and wrote the manuscript. DLY performed the mathematical modeling and prepared the illustrations. Both read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Poon, CS., Young, D.L. Nonassociative learning as gated neural integrator and differentiator in stimulus-response pathways. Behav Brain Funct 2, 29 (2006). https://doi.org/10.1186/1744-9081-2-29
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1744-9081-2-29