
Abstract
Salt marsh habitat loss and conversion are well documented across the marine-coastal district of New York. Regionally, these losses are characterized by marsh edge erosion, ditch and creek widening, internal ponding, and conversion from irregularly flooded marsh to regularly flooded marsh and intertidal mudflats. These changes in horizontal extent and shifts in vegetation composition suggest that NY’s salt marshes may not be keeping pace with sea level rise. To evaluate elevation building processes, deep rod surface elevation tables, marker horizons, and shallow rod surface elevation tables (SET-MHs and shallow RSETs) were installed as a network across Long Island, NY. Contributions of surface, shallow subsurface, and deeper processes to overall elevation changes were observed from 2008 to 2022. Using a linear mixed model approach, surface accretion, shallow subsurface rootzone growth, and deeper below-ground processes were evaluated against regional sea level rise, nutrient loading, and marsh area trends. We found that marshes on Long Island are not keeping pace with sea level rise because they lack vertical elevation growth within the rootzone. Optimizing conditions for belowground growth of native salt marsh plants and preservation of organic matter within the peat matrix is key for restoring salt marshes to a positive elevation trajectory relative to sea level rise. Much like a retirement savings account, knowing whether our marshes are increasing in elevation is important, but understanding the full suite of deposits and withdrawals is critical for managing this valuable resource for the future.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Tidal salt marshes in the marine-coastal district of New York (NY, USA) are intrinsically valuable and deliver numerous ecosystem services to both people and adjacent nature. Like all marshes, they provide critical habitat for a variety of plant and animal species, fuel the coastal food web, sequester carbon, cycle nutrients, filter coastal waters, reduce risk from storms, and provide opportunities for recreation (Barbier et al. 2011). Marshes depend on both minerogenic and biogenic processes for elevation growth (Reed 1995; Nyman et al. 2006; Kirwan and Megonigal 2013; Kirwan et al. 2016; Elsey-Quirk et al. 2022). Like many other salt marshes in the Northeastern USA, NY salt marshes do not have large mobile sediment supplies from river systems. Therefore, NY salt marshes may be more reliant on their vegetative growth and storage of organic matter to keep pace with sea level rise, making them particularly vulnerable to environmental conditions that influence those processes.
Even though NY’s Tidal Wetland Act (1973) stopped direct tidal wetland loss by conversion through building and filling (Tidal Wetlands Act NY State Environmental Conservation Law § 25-0101 1973), wetland loss continues as these habitats shrink in aerial extent and become more frequently flooded eventually converting from emergent wetland to intertidal mudflat or shallow open water habitat (Hartig et al. 2002; Long Island Tidal Wetlands Trends Analysis 2015; Smith et al. 2021). Ongoing changes in horizontal extent and shifts in vegetation composition suggest that NY’s salt marshes may not be growing vertically to keep pace with sea level rise. For wetlands to maintain themselves within their vegetation growth range relative to tidal inundation as sea levels rise, they need to grow vertically at a pace that maintains their elevation relative to sea level rise. When tidal marshes have what they need to be productive (mobile sediment supply, full tidal signal including episodic storm events, and clean water), they can grow vertically to persist in place and even grow horizontally if migration potential exists (e.g., Turner et al. 2000; Morris et al. 2002; Nyman et al. 2006; Kirwan and Megonigal 2013; Kirwan et al. 2016; Blum et al. 2021). When these natural processes impacting vertical growth are disrupted, marsh elevation growth falls behind sea level rise generating an “elevation deficit” (Cahoon et al. 1999; Cahoon and Guntenspergen 2010), whereby over time, their function and long-term viability are increasingly diminished. To better manage these systems so they persist into the future, it is important to accurately measure and characterize changes in marsh elevation over time to inform strategic management and restoration.
The surface elevation table-marker horizon (SET-MH) method, specifically the revised Rod SET (RSET) method, is recognized as the international standard for measuring precise elevation changes in tidal marshes (Boumans and Day 1993; Cahoon et al. 1995, 2002; Webb et al. 2013; Lynch et al. 2015). This method is used by scientists at academic institutions, natural resource agencies, and non-profit organizations to understand how salt marshes are responding to sea level rise and to identify which processes are responsible for observed marsh elevation trajectories. Measurements at deep RSET benchmarks measure overall marsh elevation change above the depth to which the benchmark is anchored; measurements above the feldspar marker horizon (MH) identify surface sediment accretion and elevation growth from that sediment and fine roots on the marsh surface; and measurements at shallow RSET benchmarks identify shallow elevation change within the active rootzone of marsh peat (typically < 1 m; in this study 0.5 m). The combination of these measurements across different and overlapping depths through the marsh peat suggests which constellation of surface and subsurface marsh elevation processes is at play. This may include surface sediment accretion, organic matter accumulation, belowground biomass production, decomposition, and compaction in the active rootzone, subsurface consolidation or compaction, shrinking or swelling of peat with porewater storage, and any groundwater flux that is above the depth of the deep RSET benchmark (Cahoon et al. 2011; Lynch et al. 2015).
There is an extensive and expanding network of SET-MH stations utilized for multiple purposes including monitoring, model confirmation, and hypothesis testing across the mid-Atlantic coast of the USA including but not limited to the following: in New Hampshire (Payne et al. 2019); Rhode Island (Raposa et al. 2017); within Fire Island National Seashore (Roman et al. 2007, 2023); New York City (Cahoon et al. 2019, Wigand et al. 2014, Morris et al. 2016, Hartig et al. this issue); New Jersey (Artigas et al. 2021; Weis et al. 2021, Elsey-Quirk et al. 2022); Delaware, Pennsylvania (Haaf et al. 2019); Maryland (Beckett et al. 2016); Virginia (Blum et al. 2021); and across the eastern seaboard (Yeates et al. 2020). Wigand et al. (2014) used SETs as a component of their exploration comparing disappearing and apparently stable marsh islands in Jamaica Bay, NY. Cahoon et al. (2019) expanded on the SET-MH dataset described by Wigand et al. (2014) and used linear mixed models to characterize the biogeomorphic processes operating within marshes and the relationships among marsh elevation capital, marsh surface elevation change, sea level rise, and marsh integrity. Morris et al. (2020) used SETs at four National Parks affected by Hurricane Sandy to examine how responses measured at those marshes compared to the predictions made by the marsh equilibrium model (MEM) (Morris et al. 2002, 2016) based on relationships between marsh elevation, tidal range, vegetation productivity, and sediment availability. Yeates et al. (2020) used the extensive network of SET stations across the Hurricane Sandy storm track to analyze the patterns of sediment deposition and erosion relative to the position of storm landfall to better understand how coastal marshes respond to large episodic events that are becoming more frequent and powerful with climate change. Blum et al. (2021) established deep and shallow RSETs across an elevation gradient in a mid-Atlantic marsh to explore the vertical marsh dynamics across a transect from low to high marsh which highlighted the importance of root-zone processes for elevation change and marsh transgression.
Gap in Existing Knowledge
This study fills a geographic gap in marsh elevation studies and presents the first published analysis of marsh elevation data from the mainland of Long Island, NY, outside the jurisdiction of the National Park Service (NPS). Long-term changes in marsh elevation on the bay side of Fire Island, a barrier island to the south of Long Island, were first reported by Roman et al. (2007) and updated by Roman et al. (2023). Additional studies examined the condition of marsh islands within NY’s Jamaica Bay (Wigand et al. 2014; Cahoon et al. 2019). Our network of SET stations spans a gradient of coastal development and nutrient loads from west to east across Long Island (Alldred et al. 2017; Watson et al. 2018; Krause et al. 2019). This design allows us to explore the processes responsible for elevation changes. Seven of the nine marshes included in this study have paired replicate deep and shallow RSET benchmarks in addition to surface marker horizons (MHs). Measuring deep and shallow benchmarks simultaneously allows exploration of the relative importance of marsh-building processes. Marsh processes acting on the surface (~< 0.1 m depth) include erosion, accretion, and fine surface root growth. Processes, acting within the active rootzone (< 0.5 m depth), include live root and rhizome bulking, death and collapse of roots and rhizomes, and accumulation or decomposition of organic matter. Processes, acting within the deeper subsurface (> 0.5 m), include larger subsidence or expansion processes at greater depths. While knowing if marshes are keeping pace with sea level rise is important, understanding the different drivers of that growth is critical for understanding why there are elevation shortfalls and tailoring strategic restoration, management, and conservation actions to ensure the long-term health and sustainability of these ecosystems.
In this paper, we present the results of a long-term study on salt marsh elevation trends in the marine-coastal district of Long Island, NY. We analyze marsh elevation data collected over 14 years (2008–2022) using both deep and shallow RSET-MH benchmarks. In addition to examining overall trends in marsh elevation relative to local sea level rise, we discuss the marsh surface layer, shallow active rootzone, and deeper subsurface processes that contribute to elevation growth. We want to know whether these marshes are keeping up with sea level rise and in the cases where they are falling short, to identify the reasons for elevation shortfalls so that we can develop targeted conservation and restoration strategies.
Questions addressed in this paper include:
-
1.
Are Long Island marshes keeping pace with local sea level rise?
-
2.
Are elevation deficits the result of surface or subsurface processes?
-
3.
Can we identify environmental conditions contributing to observed elevation deficits?
-
4.
Do these data suggest conservation or restoration approaches to increase the long-term viability of Long Island salt marshes?
Answers to these questions are critical for developing effective management strategies to protect and restore the function and long-term viability of these valuable ecosystems in the face of sea level rise and other environmental challenges.
Methods
Study Design
This study was set up primarily as a long-term monitoring effort, with an intentionally diagnostic mindset to reveal the underlying causes of elevation shortfalls. Study sites represent salt marshes located in different embayments and across several environmental gradients including density of development, degrees of shoreline hardening, nutrient input, historical loss trends, and potential future vulnerabilities. These co-factors, some of which surely influence the observed trends, are important to understand but go beyond the scope of this paper. By selecting diverse study sites and installing at least three replicate stations within each site, we maintain the potential for hypothesis testing.
Study Sites
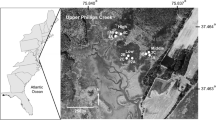
Study sites (Fig. 1) were established across a gradient of environmental conditions which can be generalized by higher wastewater-derived nutrient loading and denser development with a higher proportion of hardened shorelines to the west. Eastern sites may be exposed to lower nutrient loading overall, with a greater proportion of nitrogen coming from oceanic sources (Watson et al. 2018) versus anthropogenic (predominately wastewater). All marshes included in this study were likely intensely manipulated through historical agricultural practices (Adamowicz et al. 2020; Susan Adamowicz and Geoff Wilson pers. comms.), a condition not well understood at the time these study sites were established.

Distribution of SET-MH study sites across Long Island, NY. While the majority of study sites are in the eastern portion of LI, these sites are positioned across a number of environmental gradients
Sampling stations within each marsh site (Fig. 2) were selected randomly from a set of pre-selected candidate marsh segments, which were typically bound by grid-ditching, and selected to be representative of the marsh complex under study. Deep RSET stations were installed in triplicate during three separate efforts in 2008, 2010, and 2013 (Table 1). In all cases, the vegetation community was consistent across triplicate stations within each study site. A minimum of three feldspar marker horizon (MH) plots were paired with each deep RSET to measure surface accretion following the NPS SOP manual (Lynch et al. 2015). Shallow RSETs were later paired with deep RSET stations in 2013 at seven of nine sites (see Table 1 for site installation dates). All shallow rods were established to a consistent depth of 0.5 m to extend to the bottom of the expected active rootzone (C. Wigand pers. com). All deep RSETs were driven to 24.4 m depth or until refusal. Rod lengths for both the deep RSET and the shallow RSETs are included in Table S1 in the Supplementary Materials.

SET-MH station positions within each marsh study site
All deep and shallow RSET installations followed the guidance of Lynch et al. (2015) except for the placement of the shallow RSET relative to its paired deep RSET. Instead of setting the shallow RSETs ~1 m away from the deep RSETs as described by Lynch et al. (2015), our shallow RSETs were installed immediately adjacent to and nested within the deep RSET sampling footprint (see Fig. 3 for reference) to maximize the overlap of pin placement during data collection. The authors found this also minimized disturbance of the site and saved time and effort by reducing the need to reposition the plank for each reading. This may also capture the shallow processes within the same sampling area as the deep RSET resulting in a better accounting of the processes within a sampling station. The position of the two SET receivers and structures do not interfere with the placement of pins from either of the RSET arms. We cannot guarantee that there were no impacts of installing the shallow RSETs adjacent to the deep RSETs, but installations were done by hand from suspended benches and so no foot traffic trampled the stations. If there were any immediate impacts, we believe that they would have been acute and would not impact the long-term trajectory of the deep RSET any more than the initial installation of the deep RSET itself. Initial shallow RSET readings were performed a minimum of 1 month after installation to allow the marsh surface to recover from any acute impacts.

Shallow RSET installation nested within deep RSET station. Shown here before pipes were trimmed flush to receiver completing the installation. This placement differs from protocols (Lynch et al. 2015) because the authors wanted to maximize overlap between deep RSET and shallow RSET measurements, facilitate logistics of reading both stations, and minimize the chance of trampling if the benchmarks were more offset
RSET-Marker Horizon Measurements
Overall marsh surface elevation change was estimated through repeated measures following the SET-Marker Horizon (MH) method (Lynch et al. 2015) for each deep RSET station across all study sites. Shallow rootzone changes were measured from the shallow RSET to capture the total change within the shallow active rootzone. Detailed observations and measurements were recorded by individual RSET pin. When a RSET pin landed on an uncharacteristic marsh surface feature, such as a ribbed mussel or within a crab burrow, they were removed from the analysis before calculating trends.
Feldspar MH plots were established on the first reading of deep RSET stations with the original layer being repeatedly sampled throughout the study period. Surface accretion measures were taken by hand cutting small (40–80 mm) wedge-shaped marsh cores (three sides cut plumb, and the fourth side cut at a 45° angle to form a “bottom” and ease extraction and replacement); see Fig. 5.39 in Lynch et al. (2015) for an example, to a depth sufficient for measuring accretion above the marker horizon (typically < 15 cm). Triplicate measures were made on each marsh core from the marsh surface to the top of the marker horizon. Cores were replaced into the MH plot and gently tamped back into place using the ball of a foot.
All RSET (deep and shallow) and MH measures were made during the same site visit. Marker horizons were not sampled when the marsh surface was flooded. Measurement frequency was reduced from three readings annually (spring, summer, fall) at the start of the study to single end-of-growing-season readings in later years for seasonal consistency.
Marsh Elevations
Deep RSET receiver elevation estimates were collected via static occupation with a geodetic GPS/GNSS antenna (Leica GNSS GS14) at one centrally located RSET station within each study site to establish a benchmark elevation for the remaining RSET receivers. At each reference station, static occupations (average of 4.24 h ± 19 min; 1.2–6.7 h range) were collected for horizontal (latitudinal and longitudinal) and vertical (ortho and elliptical height (NAVD88 m)) position. Data were post-processed using OPUS Projects (US Department of Commerce 2023) and were averaged across all observations. The remaining station elevations were obtained by leveling (Leica Sprinter) from the site’s reference station.
RSET measures were corrected to marsh surface elevations by subtracting the total pin length from the sum of the receiver height, the distance from RSET receiver, as measured in meters NAVD88, to top of the RSET arm from where RSET measures are taken, and the measured pin height for each pin measure. This allows comparison across all sites to the common NAVD88 as well as an opportunity to assess the effect of starting elevation on the measured trend.
Estimating Marsh Elevation Components
Three outcomes, (1) total marsh elevation change (deep RSET), (2) shallow elevation change (shallow RSET), and (3) surface accretion rates (MH), were independently estimated for each study site using linear mixed model methods as discussed in Cahoon et al. (2019) and Russell et al. (2022) with time-adjusted pin height or depth of accretion as the response variable and time as the fixed-effect. Pins, which are nested within RSET arm directions and nested within stations, were treated as nested random effects, accounting for the lack of true independence of each measure, one of the strengths of the mixed model approach. The model allowed correlation between slopes and intercepts to accommodate any relationship between the starting elevation and the rate at which it changes over time. Each of these models (one model for each elevation component at each site) was then compared to a fixed intercept-only null model using AIC. If the reference model, which included the effect of time, was a stronger fit, the trend is considered significant. Regression coefficients with standard errors (Table S2) were then extracted for each site allocated across the vertical marsh processes. All statistical analysis was performed in R (version 4.2.1) mixed effects models where fit using the lme4 package (Bates et al. 2015).
From the three estimated outcomes of overall surface elevation change, shallow elevation change, and surface accretion, estimates of the relative contributions of each can be examined by deconstructing the overall observed vertical change in the marsh platform into its subcomponents. Repeated RSET measures yield the total overall change of the marsh platform (Lynch et al. 2015), which includes the contributions of the shallow rootzone and the surface accretion within. Removing the effect of the shallow rootzone processes from the RSET provides estimates of changes occurring below rootzone strata, such as deeper subsidence or expansion. Removal of the surface accretion portion of the shallow rootzone change provides insight into the biological processes occurring within the rootzone, such as root bulking through growth and expansion or shrinking through decomposition. Surface accretion is measured directly by the marker horizon (MH) plots and includes both biogenic and minerogenic sediments as well as increasing amounts of fine roots and rhizomes. See the column header of Table 2 for specific calculation methods of these component processes.
Correlations between all three estimated elevation components (total marsh elevation change, shallow elevation change, and surface accretion) and the derived subcomponents (change within rootzone, change below surface accretion, and change below rootzone) were assessed, recognizing that there is inherent dependence between these processes (e.g., surface accretion contributes to overall elevation change), and least squares linear regressions were fit to understand what mechanisms may be driving the changes observed across the different processes.
Inundation Rates and Elevation Capital
Water levels were collected at each study site for one growing season (June–Nov 2019) using HOBO U20-L water level recorders, corrected for barometric pressure. To capture the full tidal signal at each marsh complex, sensors were secured to stationary benchmarks along the seaward edge of the marsh platform in open water. Water elevations were corrected to NAVD88 datum using a Leica GNSS GS14-RTK system and measured several times throughout the monitoring period (n > 2). Water level recorders were lost at two sites: II and NG; thus, inundation at these sites is not presented. At each RSET station, tidal characteristics were calculated using the R package Tides (Cox and Schepers 2018), which produces estimates of high, low, and mean local sea levels and proportion of time flooded (for the monitoring period), and from that, normalized elevation capital (NEC) was determined, as defined by Morris et al. (2020) as follows:
was calculated for each RSET station. The proportion of time flooded and NEC were regressed against marsh elevation components to explore relationships between flooding regime and elevation processes.
Regional Sea Level Rise
Regional sea levels were estimated using linear regression of seasonally adjusted monthly mean sea level data acquired from NOAA tide gauges for stations closest to each study site (Montauk, NY — NOAA station # 8510560, 39.0 ± 6.2 km for eastern sites AH, BC, MP, PN, II, HC, CB; Sandy Hook, NJ — NOAA station # 8531680, 30.4 ± 1.8 for westernmost sites LW, NG). Sea level trends were estimated for each station from data collected across a full Metonic cycle or tidal epoch (19 years) which encompassed the full SET-MH monitoring period. Monthly mean water levels were extracted from NOAA-hosted (http://www.tidesandcurrents.gov) summary data using the VulnToolkit R package (Hill and Anisfeld 2021).
Nutrient Loads and Marsh Trends
To understand the potential impacts of nutrient loading on marsh elevation components and the potential relationships between marsh elevation components and vegetated marsh trends, we assessed if there were relationships between these measures and our SET-MH data. Site-level mean estimates (average across SET replicates) of each marsh elevation component (total marsh elevation change, shallow elevation change, and surface accretion) and derived subcomponents (total elevation change not attributed to accretion, change in root zone, and change below root zone) were regressed against % nitrogen in Spartina alterniflora grass blades collected at each marsh in the vicinity of the SET stations by Krause et al. (2019) as a measure of nitrogen exposure. Similarly, all marsh elevation components and subcomponents were regressed against the proportional change in overall vegetated marsh area (which includes both high and low marsh areas) for each corresponding SET study site marsh complex delineated and measured in the Long Island Tidal Wetlands Trends Analysis (2015). While the Tidal Wetlands Trends Analysis estimated change in high and low marsh separately, overall proportional change in marsh vegetation was used here to reduce mischaracterizations that could result from looking at these changes individually. For example, a high rate of conversion from high marsh to low marsh within a marsh complex will appear as a net positive change in low marsh area if the loss of high marsh is not considered. All marsh complex areas used in this analysis encompass the replicate study stations and extend beyond the immediate vicinity of the RSET replicates. Data were checked for normalcy and linearity through visual inspection, while recognizing that sample size (n = 9) will be limiting. All regressions were fit using least squares.
Results
All study sites had significant positive surface elevation trends through time as determined from comparison to each site’s respective null model. While positive trends were observed at all study sites (Fig. 4), the rates of overall elevation change, rootzone change, and accretion, although small, did vary across study sites and responses (Table 2) yielding unique combinations of contributions within each stratum to the overall vertical elevation change.

Overall elevation changes as measured on the deep RSET in black; shallow elevation changes as measured on the shallow RSET (when present) in blue; surface accretion as measured above the feldspar MH in yellow
Q1: Are Long Island Marshes Keeping Pace with Local Sea Level Rise?
All but one of the nine marsh study sites are growing in elevation at a rate slower than regional sea level rise as measured by the rate of water level rise at the closest NOAA tide gauge over the last 19 years (Metonic cycle). Sea levels for the most recent overlapping tidal epoch (2003–2022) rose 4.6 mm/year and 5.2 mm/year for Montauk, NY, and Sandy Hook, NJ, respectively (Fig. 5). These rates are similar to those reported by Cahoon et al. (2019) for Sandy Hook (2002–2016) and are significantly greater than the estimates calculated for long-term (> 80 years) sea level trends. A comparison of overall elevation change to recent sea level change at each site’s respective NOAA station (Fig. 6) (dark line with shaded 95% CI) indicates that while one site, Mashomack Point, may be maintaining vertical growth comparable to sea level, all other study sites are falling behind. Additionally, NEC indicates that all marshes are considered vulnerable as defined by Morris et al. (2020); further discussion on this is to follow.

Seasonally adjusted monthly mean sea levels for the most recent tidal epoch (2003–2022) at Sandy Hook, NJ (NOAA station 8,531,680) and Montauk, NY (NOAA station 8,510,560). Shading highlights deviation from the timeseries mean. A linear fit with a shaded 95% CI is provided along with the parameter coefficient of change over time (mm/year). Insets within each panel present long-term (Sandy Hook, NJ, 1933–2022; Montauk, NY, 1948–2022) mean sea levels for each tide station

Estimated overall marsh surface elevation change with error bars (± SE) compared to most recent tidal epoch (2003–2022) change estimates for Sandy Hook, NJ, and Montauk, NY (also Table 2, column VII), with shaded 95% CI. No site has surface elevation change estimates greater than regional sea level rates
Q2: Are Elevation Deficits the Result of Surface or Subsurface Processes?
Surface accretion accounts for much of the total elevation change at each study site. All sites other than Accabonac Harbor exhibit surface accretion rates that exceed overall elevation change. The components of elevation change in marsh peat are deconstructed into strata summarized in Table 2, and a profile view is visualized in Fig. 7. Except for Accabonac Harbor with positive elevation growth in the rootzone (V), and Lawrence Marsh with small negative growth in the rootzone (V), the mean values for the rootzone are within a standard error of zero, and therefore not significantly different from zero (Table 2). Accabonac Harbor is unique in that the lower rate of surface accretion (I) is offset by the relatively robust expansion within the rootzone (V) (1.75 mm/year ± 0.85) (Table 2; Fig. 8). Hubbard Creek displays a similar pattern of rootzone expansion (as visualized in the waterfall plot; Fig. 8), but it should be noted that the estimate for this site is within a standard error of 0, indicating that this may not be a significant contribution (Table 2). Except for Accabonac Harbor, rootzone processes do not contribute to positive elevation growth. Deeper processes below the rootzone (VI) appear to contribute to deficits in overall elevation change at 4 of 9 sites (Table 2; Fig. 8). The remaining sites are within one standard error of zero indicating no significant contribution. This indicates that deeper processes (> 0.5 m) do not contribute to positive elevation gains and at some sites offer a substantial reduction in overall elevation change.

Illustration of marker horizon (MH), deep RSET, and shallow RSET installations. Roman numerals in the drawing identify strata within the soil profile referred to in Table 2. Figure reorganized and redrawn from Fig. 2 in Blum et al. (2021) and elements of Figs. 6 and 7 in Lynch et al. (2015). Not to scale

Waterfall plot displaying the contributions (positive values in blue), reductions (negative values in red), and resulting overall change observed (in black) over the course of this sampling effort. Cedar Beach and Indian Island marshes do not have shallow RSETs installed. These plots are read from left to right. For example, at Accabonac Harbor, the average surface accretion above the marker horizon (I) was 2.29 mm/year; subsidence or expansion in rootzone strata (V) added another 1.75 mm/year; subsidence or expansion below the rootzone (VI) subtracted 1.17 mm/year resulting in an overall surface elevation change (II) as read from the deep RSET of 2.88 mm/year
Least squares regression analysis revealed some significant relationships between components of elevation change (Fig. S3). Positive relationships are found between surface accretion (I) and overall elevation change (II) (p = 0.032, R2 0.50) and between change below marker horizon (III) and change in the root zone (V) (p = 0.23, R2 0.68). Negative relationships were found between change in root zone (V) and change below root zone VI (p = 0.017, R2 = 0.71) and between surface accretion (I) and change in the root zone (V) (p = 0.020, R2 = 0.69). Care must be taken when interpreting these results due to the fact that these subcomponents are not independent measures and in some cases are components of one another (surface accretion contributes to overall elevation change so are inherently dependent on one another).
Q3: Can We Identify Environmental Conditions Contributing to Observed Elevation Deficits?
Tidal Inundations
Water level data from 2019 indicate that study sites sit within a gradient of semidiurnal tidal regimes which experience regular high tide inundations that range from 2 h 48 min to 4 h 44 min duration and which occur on 42–95% of high tides (Table 3). This range of tidal exposures results in some sites being subjected to more frequent and longer duration tidal flooding events, such as Cedar Beach and Hubbard, while other sites, Bass Creek, Lawrence Marsh, and Accabonac Harbor, are inundated less frequently. Accabonac Harbor has the lowest percentage of time inundated by the tides and is the only site with a mean marsh elevation at or greater than MHW (Table 3).
Least squares regression analysis revealed a significant relationship between a component of elevation change and % time flooded (Fig. S4). A negative relationship is found between % time flooded and change below marker horizon (III) (p = 0.002, R2 0.87).
Normalized elevation capital (NEC) estimates indicate that all study sites are vulnerable as described by Morris et al. (2020), with all sites falling below 0.5 NEC (Table 3), a point where it is predicted that accretion will decrease with increasing SLR based on the marsh equilibrium theory (Morris et al. 2020).
Least squares regression analysis revealed significant relationships between NEC and components of elevation change (Fig. S5). A negative relationship is found between NEC and Accretion (I) (p = 0.024, R2 0.67). A positive relationship is found between NEC and change below marker horizon (III) (p = 0.010, R2 0.77).
Nutrient Loads and Marsh Trends
Marsh elevation components were not significantly related to % nitrogen in Spartina alterniflora grass blades or change in overall vegetated area from 1974 to 2005.
Discussion
Q1 and Q2: Are Long Island Marshes Keeping Pace with Sea Level Rise? and Are Elevation Deficits from Surface or Subsurface Processes?
Tidal salt marsh systems across Long Island, NY, are growing vertically but experiencing an elevation deficit relative to local rates of sea level rise. These overall elevation growth results are consistent with what others are finding at marsh elevation stations in NY and southern New England (e.g., Raposa et al. 2017; Cahoon et al. 2019; Roman et al. 2023). Both minerogenic and biogenic processes are important components for peat building, but when large mineral sediment sources are not available, biogenic processes become more important (Cahoon et al. 2021; Carey et al. 2017; Nyman et al. 2006; Kirwan et al. 2010; Kirwan and Guntenspergen 2012). Even without large mobile sediment supplies like large river systems, most of the sites in this network are accreting sediments on their surface. Surface sediment accretion across all sites may be a sign of deterioration if these marshes have lost elevation relative to sea level rise, are being inundated more frequently, and there is available sediment from deteriorating marsh nearby (Wasson et al. 2019).
The deficient elevation component across this network of sites is peat formation within the rootzone (V) (below the surface marker horizon established up to 14 years ago but shallower than 0.5 m, the depth of the shallow RSET) that should be driving the elevation growth in these marshes. Apart from Accabonac Harbor, all sites have elevation changes in the rootzone that are either negative (Lawrence Marsh) or not significantly different from zero (Table 2). We interpret this to mean that none of these other sites is gaining any elevation from within this peat strata. Overall, this analysis indicates that vegetation growth and the preservation of organic matter within the active rootzone are critical for marsh persistence and are deficient in our region.
The impact of the rootzone elevation deficit in this network of marshes is illuminated compared to the findings of Blum et al. (2021) who found that in a mid-Atlantic tidal marsh where the low- and mid-elevation marsh habitats are growing at rates that exceed the local rate of sea level rise, the contribution of rootzone expansion contributed more than 37% of the overall elevation growth and always exceeded the overall marsh elevation change and the surface accretion. In contrast, except for Accabonac Harbor which is inundated the shortest amount of time, our data show no elevation growth within the critical rootzone, indicating that Long Island marshes are particularly vulnerable to sea level rise because they lack this critical component of elevation growth that is necessary for marshes without large mobile sediment supplies.
Although we found significant negative correlations between surface accretion and growth in the rootzone (V), we do not suspect that the deposition of new material is causing compaction and subsidence at this depth. Only one site (Lawrence Marsh) shows negative growth in the rootzone (V), and it has only intermediate surface accretion (I) (Table 2). Instead, we suspect that the significant negative relationship between inundation frequency and change below the marker horizon (III) (p = 0.002, R2 = 0.87) is a more important driver of these observed changes (see discussion below).
Q3: Can We Identify Environmental Conditions Contributing to Observed Elevation Deficits?
We were not able to identify the primary drivers or eliminate any specific processes or environmental conditions responsible for the lack of elevation growth in the rootzone in this study of seven marshes with paired deep and shallow SETs. Many factors are known to influence peat formation within the actively growing rootzone: deposition of sediment on the marsh surface that stimulates belowground growth (Walters and Kirwan 2016), elevation of the marsh platform relative to mean sea level and the resultant flooding frequencies (Morris et al. 2002; Kirwan and Guntenspergen 2012; Watson et al. 2014, 2017), disruption of natural hydrology (Adamowicz et al. 2020), elevated nutrient loads (Deegan et al. 2012; Alldred et al. 2017; Watson et al. 2017), and interactions among these factors.
We found a significant negative relationship (p = 0.002, R2 = 0.87) between measures of inundation frequency and the components below surface accretion (III), which includes both changes in the rootzone and deeper processes like subsidence (Fig. S4). This suggests that there may be some process related to inundation frequency that reduces root growth, increases decomposition, and/or increases compaction below the surface accretion layer. Our marshes are not only failing to keep pace, but they are also not well positioned within their tidal frames to maintain their current rates of vertical growth through rootzone processes and are reliant on the surface accretion processes for vertical marsh growth.
There are widely recognized inconsistencies in marsh responses to nutrient enrichment that influence marsh sustainability, and this puzzle is aptly characterized as the mythical Gordian Knot by Mozdzer et al. (2020). We did not find a significant relationship between % nitrogen in Spartina alterniflora leaves and any of our elevation growth components; however, this might be an artifact of the small sample size. In a related study using the same nitrogen tissue data across a much larger number of salt marsh sites, including all the marshes in this study, Krause et al. (2019) found that regularly flooded (“low”) marshes across Long Island exhibited greater rates of loss when exposed to higher nitrogen loads (1974–2005). Interestingly, in that same study, Krause et al. (2019) found that irregularly flooded (“high”) marshes across Long Island exposed to higher nutrient loads exhibited lower rates of loss over time. Inundation frequency and excess nitrogen may have a synergistic effect on low marshes (Watson et al. 2017; Krause et al. 2019). The inverse responses of high marsh and low marsh habitats to nitrogen loading and the fact that the vegetation trends represent entire marsh complex changes may explain why we did not find a significant relationship between our marsh elevation components and change in overall vegetated area across our marsh study sites.
Other studies conducted on Long Island marshes have found conflicting relationships between nitrogen loads and belowground biomass (Alldred et al. 2017; Crosby et al. 2021; Watson et al. 2022a). Crosby et al. (2021) found no effect of nitrogen loading on total belowground biomass in Spartina alterniflora creek banks in Long Island Sound, Alldred et al. (2017) found decreased total belowground biomass (live and dead), and Watson et al. (2022a) found increased belowground biomass (live and dead material in ingrowth bags initially filled with local sand). Both Crosby et al. (2021) and Watson et al. (2022a) found evidence of increased decomposition of organic matter in marshes exposed to higher nutrient loads, suggesting the role that nitrogen loading plays in decomposition of organic matter could influence accumulation and elevation growth within the rootzone. Watson et al. (2022a) suggest that, depending on the scale of each impact, increased decomposition rates could counteract elevation gains from increased belowground growth. Watson et al. (2022a) further suggests that a combination of these stressors of high nutrients, low sediment supply, and increasing flooding can cause greater marsh deterioration than would occur if only one of these stressors were present. Though beyond the scope of this effort, increasing temperatures may also be contributing to enhanced decomposition (Kirwan and Blum 2011; Carey et al. 2017).
Based on what others have reported in the literature, we suspect that all these potential factors, high nutrients, low sediment supply, and increasing flooding and/or water logging, are at play and perhaps interacting to influence peat formation in the rootzone across our network of marshes. Many Long Island marshes, including those in this study, are displaying signs of waterlogging and hydrological impairment (Long Island Tidal Wetlands Trends Analysis 2015; Smith et al. 2021; Watson et al. 2022a, b) that can impact belowground growth (Alldred et al. 2020). Marshes across Long Island and in southern New England are generalized to be at sub-optimal elevations for peak vegetation productivity (Watson et al. 2014) and increased inundation alone is shown to reduce belowground biomass (Watson et al. 2017; Payne et al. 2019).
The interdependence of these processes and our small number of sites with rootzone elevation growth estimates may be obscuring our ability to identify a single driver for observed elevation deficits. However, the NEC values that we calculate for marshes in this study suggest that they are all vulnerable according to marsh equilibrium theory (Morris et al. 2020). All sites, with the exception of Accabonac Harbor, which is positioned right at mean high water, are at elevations below mean high water (Table 3). Additionally, the elevations of all marshes in this study are below the empirically derived optima for vigorous S. alterniflora growth (0.89 m NAVD88) as calculated by Watson et al. (2014) for Long Island and southern New England salt marshes. Because these marshes are at elevations suboptimal for vigorous vegetation growth and are experiencing increased inundation times with elevated nutrient loads and low sediment loads, it is difficult to identify which of these potential threats is the most influential. Instead, all these factors may be interacting and creating a negative feedback loop compromising the long-term viability and function of these critical systems.
Q4: Do Data Suggest Conservation or Restoration Approaches to Increase Long-term Viability of Long Island Salt Marshes?
Our data suggest that conservation and restoration approaches should be designed to optimize peat formation through belowground vegetative growth and preservation of organic matter in the rootzone so these processes result in positive elevation growth and marshes can keep pace with sea level rise. Conservation goals and approaches should be crafted for each marsh system or starting condition. Conservation goals should also extend beyond the current marsh footprint into their marsh migration corridors. The extent of marshes with elevation deficits is at a scale that makes addressing individual marsh complexes in a timely manner impractical, and resource managers will need to leverage the natural process of migration to maximize the likelihood that tidal salt marsh habitats can sustain themselves in the future.
Our findings are consistent with what others are finding across the region: vegetation growing conditions and preservation of organic matter in the rootzone need to be optimized for marshes to build elevation, keep pace with sea level rise, and be resilient in the face of climate change both in their current locations and into their migration pathways (Turner et al. 2000, 2004; Nyman et al. 2006; Alldred et al. 2020; Cahoon et al. 2020; Elsey-Quirk et al. 2022). Three potential threats that should be explored to increase conservation success for these ecosystems include sub-optimal elevation for vegetation growth, impaired hydrology, and excess nutrient loading.
Sub-optimal Elevation for Vegetation Growth
In cases where too much elevation has been lost, thin-layer placement or beneficial use of sediment might be appropriate to boost the elevation of the marsh platform so that it sits higher within the tidal frame and is more optimal for vigorous vegetation growth to build elevation and keep pace with sea level rise (Cahoon et al. 2019; Raposa et al. 2022, 2023). Adding sediment and elevation, especially in a marsh without a large mobile sediment supply, can reverse a negative elevation trajectory (Cahoon et al. 2019). Artificially enhancing the elevation of a marsh with sediment would mimic sediment deposition from a storm layer that could position the marsh platform at an elevation where vigorous plant growth could trap sediment and add organic matter and root and rhizome volume to build elevation. The marsh should be considered part of a whole system within its embayment, and ideally, sediments should come from outside of that system so that they are truly additional (Ganju 2019).
Impaired Hydrology
Impaired hydrology can increase the percentage of time that a marsh is flooded regardless of starting elevation by disrupting the natural movement of the tide onto and off the marsh platform. Adamowicz et al. (2020) present a comprehensive approach to marsh restoration that starts with restoring hydrologic impairments to halt marsh subsidence, stimulate vegetation growth, and rebuild marsh elevation. Innovative techniques incorporated in this approach include application of runnels (Besterman et al. 2022; Perry et al. 2022; Watson et al. 2022b) to alleviate waterlogging and flooding stress and implementation of ditch remediation (Burdick et al. 2020) to restore single-channel hydrology disrupted by extensive historical agricultural and mosquito ditches. Restoring single channel hydrology will alleviate the oxidation and subsidence caused by extensive ditching and focus the tide’s energy and volume into and out of primary channels for each marsh tideshed (Adamowicz et al. 2020) instead of splitting it across multiple ditches. Burdick et al. (2020) pose the research question of whether single channel hydrology might also facilitate delivery of sediment to the marsh surface by hydraulic forcing through a primary ditch rather than deposition of sediment in mosquito ditches as the tide dissipates and slows as it moves through numerous ditches.
Excess Nutrient Loading
Elevated nutrient loading is associated with both positive and negative effects on belowground vegetation growth, but it consistently increases decomposition rates of organic matter. Nitrogen reduction solutions on Long Island will be site-specific but can include replacement of traditional on-site wastewater treatment infrastructure such as septic systems and cesspits with nitrogen reducing technology (NYS DEC Long Island Nitrogen Action Plan — LINAP), ensure the best treatment technology for reducing nitrogen at sewage treatment plants, ensure sewer laterals, pipes, and treatment facilities are properly maintained to reduce inflow and infiltration, and that treated effluent is sent away from sensitive water bodies (A Western Bays Resiliency Initiative 2023). Although better management of stormwater to remove nutrients is suggested for some geographies (Koch et al. 2014), a local model of nitrogen sources on Long Island, NY, suggests that surface nitrogen sources in this region are minor compared to the larger wastewater sources (Smith 2020; Kinney and Valiela 2011). Reduction of nutrient loading to coastal waters benefits people and nature in many ways by supporting ecological recovery that supports recreational and commercial opportunities beyond salt marsh resilience to climate change (e.g., Greening et al. 2014; A Western Bays Resiliency Initiative 2023).
Conclusions
Our findings show that New York’s Long Island marshes are not keeping pace with sea level rise because they lack vertical elevation growth within the rootzone. This analysis provides valuable insights into the current and future condition of these important coastal habitats and can inform management strategies to protect and preserve these ecosystems in the face of sea level rise and other environmental challenges.
Even though this analysis does not resolve the question of whether elevation relative to mean high water, increased saturation or inundation frequency because of hydrological impairments, or elevated nutrients is primarily responsible for the observed elevation deficit in the rootzone, it nevertheless informs our conservation, management, and restoration strategies. Optimizing conditions for belowground growth and preservation of organic matter in the rootzone will be key for getting salt marshes on a trajectory to keep pace with sea level rise and migrate landward, where possible.
Data Availability
The data sets and analysis generated during the current study are available from the corresponding author on reasonable request.
References
A Western Bays Resiliency Initiative. 2023. About the project. Bay park conveyance project. https://www.bayparkconveyance.org/about. Accessed 14 Feb 2023.
Adamowicz, S.C., G. Wilson, D.M. Burdick, W. Ferguson, and R. Hopping. 2020. Farmers in the marsh: Lessons from history and case studies for the future. Wetland Science and Practice 37: 182–195.
Alldred, M., J.J. Borrelli, T. Hoellein, D. Bruesewitz, and C. Zarnoch. 2020. Marsh plants enhance coastal marsh resilience by changing sediment oxygen and sulfide concentrations in an urban, eutrophic estuary. Estuaries and Coasts 43: 801–813. https://doi.org/10.1007/s12237-020-00700-9.
Alldred, M., A. Liberti, and S.B. Baines. 2017. Impact of salinity and nutrients on salt marsh stability. Ecosphere 8: e02010. https://doi.org/10.1002/ecs2.2010.
Artigas, F.J., J. Grzyb, and Y. Yao. 2021. Sea level rise and marsh surface elevation change in the Meadowlands of New Jersey. Wetlands Ecology and Management 29: 181–192. Springer Netherlands. https://doi.org/10.1007/s11273-020-09777-2.
Barbier, E.B., S.D. Hacker, C. Kennedy, E.W. Koch, A.C. Stier, and B.R. Silliman. 2011. The value of estuarine and coastal ecosystem services. Ecological Monographs 81: 169–193. https://doi.org/10.1890/10-1510.1.
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67: 1–48. https://doi.org/10.18637/jss.v067.i01.
Beckett, L.H., A.H. Baldwin, and M.S. Kearney. 2016. Tidal marshes across a Chesapeake Bay subestuary are not keeping up with sea-level rise. PLoS ONE 11: e0159753. Public Library of Science. https://doi.org/10.1371/journal.pone.0159753.
Besterman, A.F., R.W. Jakuba, W. Ferguson, D. Brennan, J.E. Costa, and L.A. Deegan. 2022. Buying time with runnels: A climate adaptation tool for salt marshes. Estuaries and Coasts 45: 1491–1501. https://doi.org/10.1007/s12237-021-01028-8.
Blum, L.K., R.R. Christian, D.R. Cahoon, and P.L. Wiberg. 2021. Processes influencing marsh elevation change in low- and high-elevation zones of a temperate salt marsh. Estuaries and Coasts 44: 818–833. https://doi.org/10.1007/s12237-020-00796-z.
Boumans, R.M.J., and J.W. Day. 1993. High precision measurements of sediment elevation in shallow coastal areas using a sedimentation-erosion table. Estuaries 16: 375–380. https://doi.org/10.2307/1352509.
Burdick, D.M., G.E. Moore, S.C. Adamowicz, G.M. Wilson, and C.R. Peter. 2020. Mitigating the legacy effects of ditching in a New England salt marsh. Estuaries and Coasts 43: 1672–1679. https://doi.org/10.1007/s12237-019-00656-5.
Cahoon, D.R., J. Day, and D. Reed. 1999. The influence of surface and shallow subsurface soil processes on wetland elevation: A synthesis. Current Topics in Wetland Biogeochemistry 3: 72–88.
Cahoon, D.R., and G.R. Guntenspergen. 2010. Climate change, sea-level rise, and coastal wetlands. In National Wetlands Newsletter, vol. 32, 1st ed. Washington, DC, USA: Environmental Law Institute®.
Cahoon, D.R., J. Lynch, B. Perez, B. Segura, R. Holland, C. Stelly, G. Stephenson, and P. Hensel. 2002. High-precision measurements of wetland sediment elevation: II. The rod surface elevation table. Journal of Sedimentary Research. 72: 734–739. https://doi.org/10.1306/020702720734.
Cahoon, D.R., J.C. Lynch, C.T. Roman, J.P. Schmit, and D.E. Skidds. 2019. Evaluating the relationship among wetland vertical development, elevation capital, sea-level rise, and tidal marsh sustainability. Estuaries and Coasts 42: 1–15. https://doi.org/10.1007/s12237-018-0448-x.
Cahoon, D.R., K.L. McKee, and J.T. Morris. 2021. How plants influence resilience of salt marsh and mangrove wetlands to sea-level rise. Estuaries and Coasts 44: 883–898. https://doi.org/10.1007/s12237-020-00834-w.
Cahoon, D.R., B.C. Perez, B.D. Segura, and J.C. Lynch. 2011. Elevation trends and shrink–swell response of wetland soils to flooding and drying. Estuarine, Coastal and Shelf Science 91: 463–474. https://doi.org/10.1016/j.ecss.2010.03.022.
Cahoon, D.R., D.J. Reed, and J.W. Day. 1995. Estimating shallow subsidence in microtidal salt marshes of the southeastern United States: Kaye and Barghoorn revisited. Marine Geology 128: 1–9. https://doi.org/10.1016/0025-3227(95)00087-F.
Cahoon, D.R., D.J. Reed, J.W. Day, J.C. Lynch, A. Swales, and R.R. Lane. 2020. Applications and utility of the surface elevation table–marker horizon method for measuring wetland elevation and shallow soil subsidence-expansion: Discussion/reply to: Byrnes M., Britsch L., Berlinghoff J., Johnson R., and Khalil S. 2019. Recent subsidence rates for Barataria Basin, Louisiana. Geo-Marine Letters 39:265–278. Geo-Marine Letters 40: 809–815. https://doi.org/10.1007/s00367-020-00656-6.
Carey, J.C., S.B. Moran, R.P. Kelly, et al. 2017. The declining role of organic matter in New England salt marshes. Estuaries and Coasts 40: 626–639.
Cox, T., and L. Schepers. 2018. Tides: Quasi-periodic time series characteristics (R package version 2.1). https://CRAN.R-project.org/package=Tides.
Crosby, S.C., N.C. Spiller, D.S. Healy, L. Brideau, L.M. Stewart, J.M. Vaudrey, K E. Tietz, and P.J. Fraboni. 2021. Assessing the resiliency of salt marshes under increasing nitrogen loading. Springer US.
Deegan, L., D.S. Johnson, R.S. Warren, B.J. Peterson, J.W. Fleeger, S. Fagherazzi, and W.M. Wollheim. 2012. Coastal eutrophication as a driver of salt marsh loss. Nature 490: 388–392. https://doi.org/10.1038/nature11533.
Elsey-Quirk, T., E.B. Watson, K. Raper, D. Kreeger, B. Paudel, L. Haaf, M. Maxwell-Doyle, A. Padeletti, E. Reilly, and D.J. Velinsky. 2022. Relationships between ecosystem properties and sea-level rise vulnerability of tidal wetlands of the U.S. Mid-Atlantic. Environmental Monitoring and Assessment 194: 292. https://doi.org/10.1007/s10661-022-09949-y.
Ganju, N.K. 2019. Marshes are the new beaches: Integrating sediment transport into restoration planning. Estuaries and Coasts 42: 917–926. https://doi.org/10.1007/s12237-019-00531-3.
Greening, H., A. Janicki, E.T. Sherwood, R. Pribble, and J.O.R. Johansson. 2014. Ecosystem responses to long-term nutrient management in an urban estuary: Tampa Bay, Florida, USA. Estuarine, Coastal and Shelf Science 151: A1–A16. https://doi.org/10.1016/j.ecss.2014.10.003.
Haaf, L., E.B. Watson, T. Elsey-Quirk, K. Raper, A. Padeletti, M. Maxwell-Doyle, D. Kreeger, and D. Velinsky. 2019. Sediment accumulation, elevation change, and the vulnerability of tidal marshes in the Delaware Estuary and Barnegat Bay to accelerated sea level rise. bioRxiv. https://doi.org/10.1101/821827.
Hartig, E.K., V. Gornitz, A. Kolker, F. Mushacke, and D. Fallon. 2002. Anthropogenic and climate-change impacts on salt marshes of Jamaica Bay, New York City. Wetlands 22: 71–89. https://doi.org/10.1672/0277-5212(2002)022[0071:AACCIO]2.0.CO;2.
Hill, T., and S. Anisfeld. 2021. VulnToolkit: Analysis of tidal datasets (R package version 1.1.4). https://doi.org/10.5281/zenodo.4598510. https://CRAN.R-project.org/package=VulnToolkit.
Holmquist, J.R., L.N. Brown, and G.M. MacDonald. 2021. Localized scenarios and latitudinal patterns of vertical and lateral resilience of tidal marshes to sea-level rise in the contiguous United States. Earth’s Future 9. https://doi.org/10.1029/2020EF001804.
Kinney, E.L., and I. Valiela. 2011. Nitrogen loading to Great South Bay: Land use, sources, retention, and transport from land to bay. Journal of Coastal Research 274: 672–686. https://doi.org/10.2112/JCOASTRES-D-09-00098.1.
Kirwan, M.L., and L.K. Blum. 2011. Enhanced decomposition offsets enhanced productivity and soil carbon accumulation in coastal wetlands responding to climate change. Biogeosciences 8: 987–993. https://doi.org/10.5194/bg-8-987-2011.
Kirwan, M.L., and G.R. Guntenspergen. 2012. Feedbacks between inundation, root production, and shoot growth in a rapidly submerging brackish marsh. Journal of Ecology 100: 764–770. https://doi.org/10.1111/j.1365-2745.2012.01957.x.
Kirwan, M.L., G.R. Guntenspergen, A. D’Alpaos, J.T. Morris, S.M. Mudd, and S. Temmerman. 2010. Limits on the adaptability of coastal marshes to rising sea level. Geophysical Research Letters 37. John Wiley & Sons, Ltd. https://doi.org/10.1029/2010GL045489.
Kirwan, M.L., and J.P. Megonigal. 2013. Tidal wetland stability in the face of human impacts and sea-level rise. Nature 504: 53–60. https://doi.org/10.1038/nature12856.
Kirwan, M.L., S. Temmerman, E.E. Skeehan, G.R. Guntenspergen, and S. Fagherazzi. 2016. Overestimation of marsh vulnerability to sea level rise. Nature Climate Change 6: 253–260. https://doi.org/10.1038/nclimate2909.
Koch, B.J., C.M. Febria, M. Gevrey, L.A. Wainger, and M.A. Palmer. 2014. Nitrogen removal by stormwater management structures: A data synthesis. JAWRA Journal of the American Water Resources Association 50: 1594–1607. https://doi.org/10.1111/jawr.12223.
Krause, J.R., E. Watson, C. Wigand, and N. Maher. 2019. Are tidal salt marshes exposed to nutrient pollution more vulnerable to sea level rise? Wetlands (Wilmington, N.C.) 40: 1–10. https://doi.org/10.1007/s13157-019-01254-8.
Long Island Tidal Wetlands Trends Analysis. 2015. Prepared for the New England Interstate Water Pollution Control Commission. Cameron Engineering & Associates, LLC.
Lynch, J., P. Hensel, and D.R. Cahoon. 2015. The surface elevation table and marker horizon technique: A protocol for monitoring wetland elevation dynamics. Report NPS/NCBN/NRR—2015/1078. Natural Resource Report. USGS Publications Warehouse.
Morris, J.T., D.C. Barber, J.C. Callaway, R. Chambers, S.C. Hagen, C.S. Hopkinson, B.J. Johnson, et al. 2016. Contributions of organic and inorganic matter to sediment volume and accretion in tidal wetlands at steady state. Earth’s Future 4: 110–121. https://doi.org/10.1002/2015EF000334.
Morris, J.T., J. Lynch, K.A. Renken, S. Stevens, M. Tyrrell, and H. Plaisted. 2020. Tidal and hurricane impacts on saltmarshes in the Northeastern Coastal and Barrier Network: Theory and empirical results. Estuaries and Coasts 43: 1658–1671. https://doi.org/10.1007/s12237-020-00790-5.
Morris, J.T., P.V. Sundareshwar, C.T. Nietch, B.R. Kjerfve, and D.R. Cahoon. 2002. Responses of coastal wetlands to rising sea level. Ecology 83: 2869–2877. https://doi.org/10.1890/0012-9658(2002)083[2869:ROCWTR]2.0.CO;2.
Mozdzer, T.J., E.B. Watson, W.H. Orem, C.M. Swarzenski, and R.E. Turner. 2020. Unraveling the Gordian Knot: Eight testable hypotheses on the effects of nutrient enrichment on tidal wetland sustainability. Science of the Total Environment 743: 140420. https://doi.org/10.1016/j.scitotenv.2020.140420.
Nyman, J.A., R.J. Walters, R.D. Delaune, and W.H. Patrick. 2006. Marsh vertical accretion via vegetative growth. Estuarine, Coastal and Shelf Science 69: 370–380. https://doi.org/10.1016/j.ecss.2006.05.041.
Payne, A.R., D.M. Burdick, and G.E. Moore. 2019. Potential effects of sea-level rise on salt marsh elevation dynamics in a New Hampshire estuary. Estuaries and Coasts 42: 1405–1418. Springer.
Perry, D.C., W. Ferguson, and C.S. Thornber. 2022. Salt marsh climate change adaptation: Using runnels to adapt to accelerating sea level rise within a drowning New England salt marsh. Restoration Ecology 30: e13466. https://doi.org/10.1111/rec.13466.
Raposa, K.B., M. Bradley, C. Chaffee, N. Ernst, W. Ferguson, T.E. Kutcher, R.A. McKinney, et al. 2022. Laying it on thick: Ecosystem effects of sediment placement on a microtidal Rhode Island salt marsh. Frontiers in Environmental Science 10. https://doi.org/10.3389/fenvs.2022.939870.
Raposa, K.B., M.L. Cole Ekberg, D.M. Burdick, N.T. Ernst, and S.C. Adamowicz. 2017. Elevation change and the vulnerability of Rhode Island (USA) salt marshes to sea-level rise. Regional Environmental Change 17: 389–397. https://doi.org/10.1007/s10113-016-1020-5.
Raposa, K.B., A. Woolfolk, C.A. Endris, M.C. Fountain, G. Moore, M. Tyrrell, R. Swerida, et al. 2023. Evaluating thin-layer sediment placement as a tool for enhancing tidal marsh resilience: A coordinated experiment across eight US National Estuarine Research Reserves. Estuaries and Coasts. https://doi.org/10.1007/s12237-022-01161-y.
Reed, D.J. 1995. The response of coastal marshes to sea-level rise: Survival or submergence? Earth Surface Processes and Landforms 20: 39–48. https://doi.org/10.1002/esp.3290200105.
Roman, C.T., D.R. King, D.R. Cahoon, J.C. Lynch, and P.G. Appleby. 2007. Evaluation of marsh development processes at Fire Island National Seashore: Recent and historic perspectives. Report NPS/NER/NRTR-2007/089. Boston, MA. USGS Publications Warehouse.
Roman, C.T., J.C. Lynch, and D.R. Cahoon. 2023. Twenty-year record of salt marsh elevation dynamics in response to sea-level rise and storm-driven barrier island geomorphic processes: Fire Island, NY, USA. Estuaries and Coasts. https://doi.org/10.1007/s12237-023-01234-6.
Russell, B.T., K.A. Cressman, J.P. Schmit, S. Shull, J.M. Rybczyk, and D.L. Frost. 2022. How should surface elevation table data be analyzed? A comparison of several commonly used analysis methods and one newly proposed approach. Environmental and Ecological Statistics. https://doi.org/10.1007/s10651-021-00524-1.
Smith, C.D.M. 2020. Suffolk County subwatersheds wastewater plan. Suffolk County Department of Health Services. https://www.suffolkcountyny.gov/swp.html.
Smith, J.A.M., S.C. Adamowicz, G.M. Wilson, and I. Rochlin. 2021. “Waffle” pools in ditched salt marshes: Assessment, potential causes, and management. Wetlands Ecology and Management. https://doi.org/10.1007/s11273-021-09835-3.
Tidal Wetlands Act NY State Environmental Conservation Law § 25–0101. 1973.
Turner, R.E., E.M. Swenson, C.S. Milan, J.M. Lee, and T.A. Oswald. 2004. Below-ground biomass in healthy and impaired salt marshes. Ecological Research 19: 29–35. https://doi.org/10.1111/j.1440-1703.2003.00610.x.
Turner, R.E., E.M. Swenson, and C.S. Milan. 2000. Organic and inorganic contributions to vertical accretion in salt marsh sediments. In Concepts and controversies in tidal marsh ecology, ed. M.P. Weinstein and D.A. Kreeger, 583–595. Dordrecht: Springer Netherlands. https://doi.org/10.1007/0-306-47534-0_27.
US Department of Commerce, N. 2023. OPUS Projects. https://geodesy.noaa.gov/OPUS-Projects/OpusProjects.shtml. Accessed 10 Feb.
Walters, D.C., and M.L. Kirwan. 2016. Optimal hurricane overwash thickness for maximizing marsh resilience to sea level rise. Ecology and Evolution 6: 2948–2956. https://doi.org/10.1002/ece3.2024.
Wasson, K.,N. K. Ganju, Z. Defne, C. Endris, T. Elsey-Quirk, K.M. Thorne, C.M. Freeman, G. Guntenspergen, D.J. Nowacki, and K.B. Raposa. 2019. Understanding tidal marsh trajectories: Evaluation of multiple indicators of marsh persistence. Environmental Research Letters 14: 124073. IOP Publishing. https://doi.org/10.1088/1748-9326/ab5a94.
Watson, E.B., A.J. Oczkowski, C. Wigand, A.R. Hanson, E.W. Davey, S. Crosby, R.L. Johnson, and H. Andrews. 2014. utrient enrichment and precipitation changes do not enhance resiliency of salt marshes to sea level rise in the Northeastern U.S. Climatic Change 125: 501–509. Springer, New York, NY.
Watson, E.B., C. Wigand, E.W. Davey, H.M. Andrews, J. Bishop, and K.B. Raposa. 2017. Wetland loss patterns and inundation-productivity relationships prognosticate widespread salt marsh loss for southern New England. Estuaries and Coasts 40: 662–681. https://doi.org/10.1007/s12237-016-0069-1.
Watson, E.B., Elisabeth Powell, Nicole P. Maher, Autumn J. Oczkowski, Bhanu Paudel, Adam Starke, Katelyn Szura, and Cathleen Wigand. 2018. Indicators of nutrient pollution in Long Island, New York, estuarine environments. Marine Environmental Research 134: 109–120. https://doi.org/10.1016/j.marenvres.2018.01.003.
Watson, E.B., F.I. Rahman, A. Woolfolk, R. Meyer, N. Maher, C. Wigand, and A.B. Gray. 2022a. High nutrient loads amplify carbon cycling across California and New York coastal wetlands but with ambiguous effects on marsh integrity and sustainability. PLoS ONE 17: e0273260. Public Library of Science. https://doi.org/10.1371/journal.pone.0273260.
Watson, E.B., W. Ferguson, L.K. Champlin, J.D. White, N. Ernst, H.A. Sylla, B.P. Wilburn, and C. Wigand. 2022b. Runnels mitigate marsh drowning in microtidal salt marshes. Frontiers in Environmental Science 10. https://doi.org/10.3389/fenvs.2022.987246.
Webb, E.L., D.A. Friess, K.W. Krauss, D.R. Cahoon, G.R. Guntenspergen, and J. Phelps. 2013. A global standard for monitoring coastal wetland vulnerability to accelerated sea-level rise. Nature Climate Change 3: 458–465. Springer Science and Business Media LLC. https://doi.org/10.1038/nclimate1756.
Weis, J.S., E.B. Watson, B. Ravit, C. Harman, and M. Yepsen. 2021. The status and future of tidal marshes in New Jersey faced with sea level rise. Anthropocene Coasts 4: 168–192. NRC Research Press. https://doi.org/10.1139/anc-2020-0020.
Wigand, C., C.T. Roman, E. Davey, M. Stolt, R. Johnson, A. Hanson, E.B. Watson, et al. 2014. Below the disappearing marshes of an urban estuary: Historic nitrogen trends and soil structure. Ecological Applications 24: 633–649. https://doi.org/10.1890/13-0594.1.
Yeates, A.G., J.B. Grace, J.H. Olker, G.R. Guntenspergen, D.R. Cahoon, S. Adamowicz, S.C. Anisfeld, et al. 2020. Hurricane Sandy effects on coastal marsh elevation change. Estuaries and Coasts 43: 1640–1657. https://doi.org/10.1007/s12237-020-00758-5.
Acknowledgements
We have many people to thank for their contributions to this work including: Kirk Raper for water level and static elevation data collection and processing. Jeff Wagenhauser, Rob Shao, Matt Grasso, Alison Branco, Sarah Schaefer-Brown, Elizabeth Hornstein plus the many TNC volunteers over the years, who have been reliable assistants in the field. Thank you to the Peconic Estuary Partnership for including this effort in the good work they do. We would like to thank TNC Coastal, Marine, Climate Adaptation, and Oceans Team Leads, Sarah Newkirk, Wayne Grothe, Alison Branco, and Carl Lobue who supported this work over the years because they saw the value in building long-term datasets. We would like to thank Elizabeth Watson and Adrianna Zito-Livingston for reviewing early drafts of this manuscript; any omissions that remain are our own. We would like to thank Anna Bartlett for editing the manuscript for clarity and impact. Lastly, we are grateful to Suffolk County Parks for providing access to their lands and for partnering with us on this long-term data collection effort.
Funding
Funding was received from David and Lisa Walker, the William E. & Maude S. Pritchard Charitable Trust, the Zegar Family Foundation, the Knapp Swezey Foundation, Inc., and the Garden Club of Lawrence.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Mead Allison
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Maher, N., Starke, A. Suboptimal Rootzone Growth Prevents Long Island (NY) Salt Marshes from Keeping Pace with Sea Level Rise. Estuaries and Coasts (2023). https://doi.org/10.1007/s12237-023-01295-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12237-023-01295-7