
Abstract
Plant cell walls are highly dynamic and chemically complex structures surrounding all plant cells. They provide structural support, protection from both abiotic and biotic stress as well as ensure containment of turgor. Recently evidence has accumulated that a dedicated mechanism exists in plants, which is monitoring the functional integrity of cell walls and initiates adaptive responses to maintain integrity in case it is impaired during growth, development or exposure to biotic and abiotic stress. The available evidence indicates that detection of impairment involves mechano-perception, while reactive oxygen species and phytohormone-based signaling processes play key roles in translating signals generated and regulating adaptive responses. More recently it has also become obvious that the mechanisms mediating cell wall integrity maintenance and pattern triggered immunity are interacting with each other to modulate the adaptive responses to biotic stress and cell wall integrity impairment. Here we will review initially our current knowledge regarding the mode of action of the maintenance mechanism, discuss mechanisms mediating responses to biotic stresses and highlight how both mechanisms may modulate adaptive responses. This first part will be focused on Arabidopsis thaliana since most of the relevant knowledge derives from this model organism. We will then proceed to provide perspective to what extent the relevant molecular mechanisms are conserved in other plant species and close by discussing current knowledge of the transcriptional machinery responsible for controlling the adaptive responses using selected examples.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cell walls are essential for plant development and tightly involved in many signaling processes. They provide mechanical support required for land plants growing on soil and are normally the first barriers involved in perception of stimuli and activation of signaling pathways inducing responses in the plasma membrane, cytoskeleton, cytoplasm, and organelles. Relevant stimuli during interactions between plants and their environment can have either a physical or chemical nature (Lamers et al. 2020; Le Gall et al. 2015). Physical stimuli include light, temperature, turgor pressure and mechanical forces, which affect the position of the cell wall with respect to the plasma membrane and could also include pressure in cytoskeleton structures (Bacete and Hamann 2020; Vaahtera et al. 2019). Chemical stimuli include non-plant derived molecules such as pathogen-associated molecular patterns (PAMPs) or plant-derived ones like damage-associated molecular patterns (DAMPs), volatiles, REACTIVE OXYGEN SPECIES (ROS) (including nitric oxide) and phytohormones (Chae et al. 2021; DeFalco and Zipfel 2021; Engelsdorf et al. 2018; Widhalm et al. 2015; Wong et al. 2021). Whether mechanical forces can also generate chemical signals by releasing components of the cell wall needs to be further explored.
Plants cells monitor the integrity of their cell walls and integrity impairment leads to responses mediated by the cell wall integrity (CWI) maintenance mechanism (Rui and Dinneny 2020; Somerville 2004; Vaahtera et al. 2019). CWI impairment is caused by cell wall damage (CWD) occurring during exposure to abiotic and biotic stress as well as growth and development. Receptor-like kinases and ion channels are typically involved in sensing CWI impairment and activating responses, including hormone biosynthesis and signaling, ROS, calcium transport and transcriptional regulation (Basu and Haswell 2020; Lamers et al. 2020; Chao Li et al. 2016a, b, c). Crosstalk between different pathways involving the same molecular components has made it difficult to fully comprehend the specific responses activated after exposure to a particular environmental stimulus or combination thereof. Nevertheless, CWI maintenance seems to be an attractive model system to study integration of signals induced by physical and chemical stimuli since previous work has shown that perception of both is relevant for integrity maintenance (Engelsdorf et al. 2018; Basu and Haswell 2020). Combining this model system with novel plant genome editing techniques that target specific genes and pathways should help us characterize gene hubs and genes with specific functions (Zhang et al. 2019; Zhu et al. 2020). The resulting knowledge will enable us to understand the mode of action of the mechanisms responsible for signal integration.
Typical methods to investigate CWI maintenance mechanisms involve the use of chemical agents or active enzymes to cause CWD (Engelsdorf et al. 2018). Cellulose is the main load-bearing component of cell walls and has been targeted by cellulose biosynthesis inhibitors (CBIs), like isoxaben (Desprez et al. 2002; Scheible et al. 2001). This pesticide affects the function of CELLULOSE SYNTHASE (CESA) proteins required for primary cell wall formation, resulting in reduced number of cellulose microfibrils leading to cell wall failure (Tateno et al. 2016). Interestingly, dicot plants are more sensitive to CBIs than monocots but the reason for this remains unknown (García-Angulo et al. 2012; Tateno et al. 2016). Techniques used to investigate cell wall composition and structure such as monoclonal antibodies, Fourier-transform infrared spectroscopy, Brillouin microscopy and labeling dyes are described in other reviews and will therefore not be reviewed here (Alonso-Simón et al. 2011; Prevedel et al. 2019; Rydahl et al. 2018; Ursache et al. 2021).
Most of the knowledge we currently have regarding the CWI maintenance mechanism and how it contributes to plant adaptation to environmental challenges comes from studies in Arabidopsis thaliana (hereafter Arabidopsis). Nevertheless, an increasing number of studies has started to shed light on the similarities and differences of CWI maintenance components in other plant species, including ferns, crops, and trees. Understanding how CWI functions in different plants species could have big economic impacts on plant-derived food and biofuel production (Ezquer et al. 2020; Ha et al. 2021). Climate change is causing extreme weather events that have profound negative impact on agricultural production (Intergovernmental Panel on Climate Change 2019). Therefore, understanding how plant cells react to abiotic and biotic stress will be beneficial for food crop productivity. In addition, plant biomass has the potential to be used as a sustainable resource to replace raw materials currently used for energy production or act as source for fine chemicals (Yoo et al. 2020). The latter is illustrated by lignin, which consists of monomers of interest to the chemical industry as precursors for plastics, and whose production is at least partially regulated by the CWI maintenance mechanism (Denness et al. 2011; Miedes et al. 2014). Therefore it is of particular interest to understand the CWI maintenance mechanism process as it can potentially enable us to become a more carbon–neutral society (European Commission. Directorate-General for Research and Innovation 2018).
Recent publications have focused on how mechanoperception affects CWI (Bacete and Hamann 2020), the relevance of differences in cell wall composition between monocots and dicots for CWI (Gigli-Bisceglia et al. 2020), and how these differences affect defense responses (Molina et al. 2021). Therefore, these topics will not be discussed here. In this review we will focus on chemical signals contributing to CWI maintenance and on the transcriptional regulation of relevant cell wall biosynthesis genes. We start by discussing the currently available information in Arabidopsis, proceed to summarize the knowledge available in other plant species and conclude by comparing and contrasting similarities and differences between molecular components and their interactions.
Cell wall integrity maintenance: knowledge obtained from using Arabidopsis as model system
The role of cell-surface receptors: CrRLK1L, WAK and their interactions
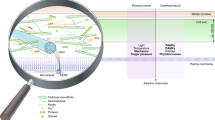
Environmental signals are mainly sensed in the cell wall-plasma membrane continuum (Fig. 1). Displacement between the two caused by mechanical forces can activate membrane-bound receptors and ion channels (Bacete and Hamann 2020; Basu and Haswell 2020; Codjoe et al. 2021; Yoshimura et al. 2021). Almost 20 different kinase protein families have been discovered in plants and several of them have been implicated in plasma membrane-located signaling processes (Dievart et al. 2020). Typically, the kinases contain a glycosylated extracellular ligand-binding domain, a hydrophobic transmembrane domain and a cytoplasmic kinase catalytic domain. Examples of such kinases are the members of the Catharanthus roseus receptor-like kinase1-like (CrRLK1L) and Wall-associated kinase (WAK) subfamilies, which seem to interact with pectin via their extracellular domains (Decreux and Messiaen 2005; Feng et al. 2018; Gonneau et al. 2018; Kohorn 2016). This also supports the notion that they may function in sensing cell wall matrix disturbance. Different aspects of development, CWI maintenance, reproduction and response to stress are regulated by CrRLK1L and WAK proteins but a large number of family members, their broad expression domains and the resulting redundancy has made it difficult to discern specific biological functions of individual family members (Franck et al. 2018).

Receptors and their interactors participating in CWI maintenance in Arabidopsis. Membrane-bound receptor-like kinases can activate signaling pathways in response to environmental or chemical cues. Different interaction partners (co-receptors) and ligands (RALF peptides) seem to determine the specific activities of signaling process receptors, such as FER and THE1. Interaction with cell wall components (i.e. WAK or FER interactions with pectin) could trigger downstream signaling events
The most studied member of this family, FERONIA (FER), illustrates this difficulty. FER has been implicated in many different biological processes including female fertility, cell elongation, immunity responses and mechanosensing (Höfte, 2015; Ortiz-Morea et al. 2021; Zhong et al. 2022). The role of FER in so many diverse processes has been explained by its capacity to bind to different interactors. Rapid alkalinization factor (RALF) peptides are growth regulators found in different plant species (Zhang et al. 2020b). FER seems to work as a scaffold, because it can interact with several different RALF peptides and other molecular interaction partners (Stegmann et al. 2017). FER functions as receptor for several RALFs including RALF1 (Abarca et al. 2021). The FER-RALF1 interaction enhances FER phosphorylation capacity and inhibits proton transport mediated by H+-ATPase pumps across the plasma membrane, which in turn probably affects modification of cell walls according to the acid growth hypothesis (Haruta et al. 2014). FER also regulates plant immunity by interacting with RALF23 to modulate elf18-induced ROS production (Xiao et al. 2019). Additionally, FER enhances flg22-induced FLS2-BAK1 complex formation, making FER an interactor/context-dependent positive or a negative regulator of plant immunity. This is illustrated by the phenotypes of fer loss of function mutants, which are hyposensitive to flg22, elf18 and chitin but more susceptible to Pseudomonas syringae pv. tomato DC3000 coronatine-minus strain (Stegmann et al. 2017). The FER-induced resistance is due to the activation of ethylene/jasmonic acid (JA) pathways (Kessler et al. 2010; Stegmann et al. 2017). FER also interacts with RHO OF PLANTS GUANINE EXCHANGE FACTOR 1 (ROPGEF1) and functions in the RAC/ROC (after the small family of GTPases Rho, Rac and Cdc42) signaling pathway to regulate ROS-mediated root hair growth (Duan et al. 2010) and to suppress abscisic acid (ABA) signaling by activating the phosphatase ABA INSENITIVE2 (ABI2) (Yu et al. 2012). FER also contributes to cell expansion by interaction with extracellular Leucine-rich repeat extensins (LRXs) to promote vacuole expansion (Dünser et al. 2019). The connection between the extracellular signal and vacuolar expansion has not been fully elucidated, but the current hypothesis suggests that actin dynamics, which are regulated by the FER-interacting RAC/ROP GTPases, could be modulating vacuole size. Actin dynamics have been recently shown to respond to osmotic stress (NaCl and mannitol) by changing the orientation of their filaments by 90 degrees in Arabidopsis and Chlamydomonas (Vilarrasa-Blasi et al. 2021). The FER-LRX-RALF module regulates the activity of salicylic acid (SA), JA, ABA and ROS pathways during plant growth under abiotic stress (Zhao et al. 2021). FER has also been implicated on the growth recovery phase after salt stress, where it is necessary to increase cytosolic calcium concentrations and to prevent cell bursting (Feng et al. 2018). This evidence suggests that FER could be implicated in vacuolar-driven cellular expansion upon exposure to abiotic stress, however the molecular mechanism remains to be determined.
THESEUS1 (THE1), the founder member of the CrRLK1L gene subfamily, was identified as a suppressor of the short-hypocotyl phenotype observed in the cellulose-deficient procuste1-1 (pcr1-1) mutant (Hématy et al. 2007). Knockout mutants (the1-1, the1-2, the1-3 and the1-6) show no phenotypic defects in non-stressed conditions, suggesting that THE1 becomes active only if CWI has been impaired, supporting the notion that it functions as CWI sensor. The the1-4 allele contains a T-DNA insertion in the region of the protein connecting the transmembrane and the kinase domains (Merz et al. 2017). Interestingly, in this allele, a truncated protein is produced that enhances the prc1-1 phenotype, indicating that the kinase domain is not necessary for THE1 activation in cellulose deficient plants but for reducing its activity (Merz et al. 2017). Intriguingly the the1-3 allele contains a T-DNA insertion 120 bases downstream from the the1-4 insertion site and results in a loss-of-function allele (Merz et al. 2017). To explain the opposite effects, it was proposed that antisense transcripts (detected for both alleles but tenfold higher in the1-3) prevent gene expression in the1-3 but not the1-4, thus making the1-4 a gain-of-function allele. Furthermore, THE1 is required for cellulose-deficiency-induced lignin accumulation (Denness et al. 2011; Hématy et al. 2007). Importantly, THE1 does not influence cellulose deposition but rather senses cell wall perturbations (Hématy et al. 2007; Lindner et al. 2012). By combining a genetic analysis with measurements of JA, SA and lignin, it was shown that THE1 works in the same pathway as the mechanosensitive Ca+ channel MATING PHEROMONE INDUCED DEATH1 (MID1)—COMPLEMENTING ACTIVITY 1 (MCA1) and the RLK FEI2 regulating CWI (Engelsdorf et al. 2018). THE1 also interacts with a ROPGEF family member (ROPGEF4), similar to FER (Qu et al. 2017). This interaction seems to mediate plant defense responses against the necrotrophic fungi Bortrytis cinerea. THE1 functions as a receptor for RALF34 in a pH-dependent manner (Gonneau et al. 2018) to regulate lateral root initiation, with its specific mode of action to be determined.
Even though FER and THE1 have similar expression patterns in vegetative tissues and interaction partners from the same protein families (RALF and ROPGEF), their knockout phenotypes are drastically different. the1 seedlings and plants only exhibit a visible phenotype when CWI is impaired (by treating them with a cellulose biosynthesis inhibitor or by combining them with cellulose-deficient mutants; Engelsdorf et al. 2018; Hématy et al. 2007). In contrast, fer plants exhibit very pronounced phenotypes, even in non-stressed conditions, exemplified by dwarf growth and reduced leave size due to reduced cell elongation (Deslauriers and Larsen 2010; Guo et al. 2009a, b). Mutations in other members of the CrRLK1L family exhibit only minor effects on cell elongation (HERCULES RECEPTOR KINASE1; Guo et al. 2009a) and polar growth of root hairs (ANXUR1 and ANXUR2;Boisson-Dernier et al. 2009; Miyazaki et al. 2009). This raises the intriguing question why mutations in only one family member (FER) result in diverse and often strong mutant phenotypes. This is particularly surprising since the majority of the family members have broad expression domains and their expression is activated by specific abiotic and biotic stresses (Franck et al. 2018; Lindner et al. 2012), implying that they could participate in responding to different environmental challenges. One possible explanation for the differential effects observed being that redundancy exists within the family, which does not include FER, leading then to the obvious question why FER is special.
As previously alluded to, FER plays essential roles in cell wall synthesis, stress response, cellular growth, morphogenesis and fertilization (Escobar-Restrepo et al. 2007; Franck et al. 2018; Guo et al. 2018; Shih et al. 2014). FER can bind to pectin fragments in vitro, to RALF peptides and also interact with LRXs. These characteristics have supported the notion that FER could act as a CWI sensor and core component of CWI maintenance (Feng et al. 2018; Shih et al. 2014). However, it is important to note that a FER knock down allele leads to enhanced production of JA, SA and lignin after exposure of fer-5 seedlings to CWD, similar to the effects of the the1-4 gain of function allele (Engelsdorf et al. 2018). These effects indicate that Arabidopsis plants can still perceive CWD (induced by cellulose deficiency) in the absence of FER. This suggests in turn that FER is not essentially required for detection of CWI impairment caused by cellulose biosynthesis inhibition, but is possibly required for perception of a different type of CWD or redundancy for certain FER activities does exist within the family.
The WAK subfamily contains 5 members in Arabidopsis, which are characterized by their conserved Epidermal growth factor (EGF)-containing extracellular domain, a transmembrane region and a conserved kinase domain (Kohorn 2016). Arabidopsis also encodes 21 WAK-like (WAKL) kinases, which show little similarity to the WAKs except in the EGF repeats and kinase domains, but it is not clear whether they associate with the cell wall (Verica and He 2002). WAK1, preferentially expressed in the vasculature, and WAK2, preferentially expressed in organ junctions, abscission zones and meristems, are the most abundant WAKs. WAK2 mutants exhibit shorter roots and reduction of vacuolar invertase levels, whereas the hyperactive receptor WAK2cTAP induced stress responses, which were suppressed in a PECTIN-METHYLESTERASE3 (PME3) loss-of-function allele, suggesting that de-esterified pectin activates the WAK2 hyperactive allele. (Kohorn et al. 2012, 2009, 2006). This is in accordance with other data showing that WAKs can bind oligogalacturonides (OGs) and (preferentially) de-esterified pectin (Kohorn 2016). Other components of the signaling cascades in which WAKs are involved include MITOGEN-ACTIVATED PROTEIN KINASE 3 (MPK3), MPK6 and downstream targets of the signaling processes such as a vacuolar invertase, ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) and PHYTOALEXIN DEFICIENT4 (PAD4), required for pathogen resistance as well as affecting turgor pressure (Kohorn 2016). While our knowledge regarding WAKs and WAKLs has improved significantly over recent years, we still do not fully understand their respective biological functions and mode of action nor identified their interaction partners. One possible way to resolve this situation could be to combine CRISPR-CAS-based manipulation of their respective activities with (phospho-)proteomics studies.
Representative PAMPs and DAMPs relevant for CWI maintenance
As sessile organisms, plants have developed mechanisms to resist a variety of stresses. Pathogen infection can cause CWD during infection and activate the CWI maintenance mechanism (Hamann 2012; Novaković et al. 2018). Biotic stress responses are activated during plant infection by microbes and pathogens. Plant infection can occur through natural openings like stomata, open wounds, or direct infiltration by the usage of cell wall (CW)-degrading enzymes (Muthamilarasan and Prasad 2013). CW-degrading enzymes help to penetrate into the host tissues and make carbon sources and nutrients available to infecting pathogens (Cantu et al. 2008). Plants have evolved a multi-level protection system to prevent successful infection. The first level of protection consists of the physical-structural defense determined by cell wall composition and structure, which influence infection success or failure (Bacete et al. 2018). The second level is the response to signal molecules such as PAMPs and DAMPs. Plants can recognize specific invaders by their molecular patterns and activate specific defense mechanisms in response (Fig. 2; Abdul Malik et al. 2020; DeFalco and Zipfel, 2021; Zhou and Zhang 2020). DAMPs are produced by damaged cells or secreted from intact cells undergoing pathogen invasion. They typically consist of wall glycans, cytosolic proteins, protein fragments, peptides, nucleotides and amino acids (Hou et al. 2019). In contrast, PAMPs are secreted by invaders. PAMPs and DAMPs are recognized by pattern recognition receptors (PRRs), which are plasma membrane-localized receptor-like kinases (RLKs) or receptor-like proteins (RLPs) which activate plant immune responses (Li et al. 2020; Newman et al. 2013). The PRR family is divided into subfamilies according to their ability to perceive signals. Leucine-rich repeat receptor-like kinases (LRR-RLK), such as FLAGELLIN SENSING 2 (FLS2) and EF-TU RECEPTOR (EFR), perceive signals deriving from FLAGELLIN (flg22 epitope) and EF-Tu (elf18/elf26 epitopes). The PRR lysin-motif (LysM) RLK CHITIN ELICITOR RECEPTOR KINASE-1 (CERK1) binds to fungal chitin-oligomers and bacterial peptidoglycans (Couto and Zipfel 2016). P/DAMPs-triggered immunity (PTI) both involve a drastic increase in ROS production, activation of MAPK modules and Ca2+-DEPENDENT PROTEIN KINASES (CDPKs), modulation of hormone-based signaling, reorganization of the cytoskeleton and changes in gene expression (Ferrari et al. 2007; Giovannoni et al. 2021; Gravino et al. 2015; Kawasaki et al. 2017; Marti et al. 2021; Wong et al. 2007). These procedures also activate other defense mechanisms, which modulate CWI, cell wall composition/structure and help the plant to resist wall degradation. At the same time, activated signaling pathways also modify allocation of resources between growth and defense to maximise resources available to support defense responses (Lorrai and Ferrari 2021). CWI maintenance involves also accumulation of phytohormones such as JA, SA, and ethylene, increased production of ROS and activation of Ca2+-based and MAPK signaling modules. These inputs lead to ectopic production of cell wall components such as callose and lignin, originally implicated in defense responses (Denness et al. 2011; Ellis and Turner 2001; Engelsdorf et al. 2018; Hamann et al. 2009; Kohorn et al. 2009; Nakagawa et al. 2007; Tsang et al. 2011). Here we will focus on selected examples for PAMPs and DAMPs, because they can be used as tools to understand how CWI maintenance and PTI are controlling plant defense responses in a coordinated manner to successfully resist pathogen infection.

PAMPs and DAMPs activate diverse pathways in response to environmental stimuli. PAMPs and DAMPs are sensed at the plasma membrane by receptors from diverse families. Dependent on the specific perturbation and foreign or endogenous molecule detected specific defense responses are activated. Similar responses to biotic stress are observed in Arabidopsis and other plants, whereas chitin detection differs between Arabidopsis and rice (see text and Fig. 4)
Flagellin is the main building block of the organs responsible for moving almost all flagellated bacteria and the primary ligand of receptors like FLS2 (Schuster and Khan 1994). Flagellin perception depends on FLS2 heterodimerization with the LRR-RLK co-receptor BRASSINOSTEROID INSENSITIVE 1-ASSOCIATED KINASE 1 (BAK1) followed by activation of signal translation elements such as BOTRYTIS-INDUCED KINASE 1 (BIK1) and MAPK species, PBS1-LIKE KINASES 1 (PBL1), and SUGAR TRANSPORT PROTEIN 13 (STP13), which leads to accumulation of hydrogen peroxide, reorganization of the cytoskeleton and increased hexose uptake (Henty-Ridilla et al. 2013; Lee and Seo 2021; Li and Staiger 2018; Macho and Zipfel 2014; Rasmussen et al. 2012; Zhang et al. 2010).
Plant defense responses induced by perception of chitin involve mainly secretion of chitinases (hydrolytic enzymes). However, a minor strategy involves recognition of fungal chitin by LYSIN MOTIF-CONTAINING RECEPTOR-LIKE KINASE 5 (LYK5) and CERK1 (Miya et al. 2007). LYK5 has a higher affinity to chitin than CERK1, which enhances the chitin-mediated formation of LYK5/CERK1 complex and CERK1 phosphorylation (Cao et al. 2014; Miya et al. 2007; Petutschnig et al. 2010). The signals generated by this complex are translated by RECEPTOR-LIKE CYTOPLASMIC KINASES (RLCK) VII-4 and PBL27, which in turn activate a MAPK signaling module (Bi et al. 2018; Rao et al. 2018; Yamada et al. 2016). CALCIUM-DEPENDENT PROTEIN KINASE 5 (CPK5) was recently implicated in the signaling process, because it can directly phosphorylate LYK5 and thus modulate chitin-induced defense responses in plants (Huang et al. 2020). Interestingly, chitin is not the only ligand that activates the CERK1/LYK5 complex. Callose-derived DAMPs, such as non-branched β-1,3-glucan oligosaccharides, promote immune responses in Arabidopsis through this complex as well (Mélida et al. 2018). Intriguingly, the juxta-membrane domains of CERK1, BAK1 and FLS2 all regulate the kinase activities of these receptors and play conserved roles in chitin signaling. This exemplifies the functionally similar organization of the receptor-based signaling processes mediating plant responses to chitin (Zhou et al. 2020).
Lipo-polysaccharides (LPS) form another relevant group of PAMPs (Kutschera et al. 2019; Shang-Guan et al. 2018; Sun et al. 2017). They act as ligands for the LECTIN RECEPTOR-LIKE KINASES (LecRLKs). LecRLKs bind non-enzymatically to specific carbohydrates, which function as links between the plasma membrane and cell wall (Gouget et al. 2006; Vaid et al. 2013). LecRLKs consist of an extracellular lectin domain, a transmembrane region and a cytoplasmic kinase region (Vaid et al. 2013). The classification of lectins is based on amino acid sequence, structure, as well as properties of the lectin domains and has led to a division of all lectins into three sub-classes. The first-class, C-type lectins require calcium ions for carbohydrate binding. The second-class, G-type lectins are Galanthus nivalis agglutinin-related lectins. The last-class are the L-type lectins, which were first discovered in Legume seeds (Bellande et al. 2017; Vaid et al. 2013). LPS can activate G-type PRR LIPOOLIGOSACCHARIDE-SPECIFIC REDUCED ELICITATION (LORE), which in turn phosphorylates the cytoplasmic receptor kinases PBL34, PBL35, and PBL36, leading to activation of a MAPK signaling pathway (Luo et al. 2020; Ranf et al. 2015; Sun et al. 2020). L-types LecRK-IX.2 and LecRK-I.9 (DOES NOT RESPOND TO NUCLEOTIDES 1 [DORN1]), which perceive flg22 and extracellular ATP, respectively; induce PTI responses including Ca2+ influx, direct phosphorylation and activation of RESPIRATORY BURST OXIDASE HOMOLOGS D (RBOHD) leading to ROS production and activation of the MAPK signaling module (Choi et al. 2014; Luo et al. 2017; Wang et al. 2018a, b).
PLANT ELICITOR PEPTIDES (PEPs) are well-established DAMPs and play an important role in PTI (Bartels and Boller 2015; Huffaker 2015; Lori et al. 2015). PEP precursors are sequestered at the vacuolar membrane. When cells are damaged, they are activated by METACASPASE4 (MC4) and released only in the damaged cells (Hander et al. 2019). PEPs bind to PEP-RECEPTOR1 and 2 (PEPR1 and PEPR2) and activate different responses. This includes interaction of PEPR2 with the proton pump AUTOINHIBITED H + -ATPase 2 (AHA2; Shen et al. 2020). AHA2 pumps protons into the apoplast, thus decreasing the pH in the apoplastic space (Shen et al. 2020). The PEP1/PEPR2 complex also interacts with BIK1 and RBOHD/F, activating a robust burst of ROS in response to bacterial pathogens (Jing et al. 2020). Intriguingly, application of PEPs suppresses in PEPR-dependent manner CWD-induced phytohormone production (Engelsdorf et al. 2018). Simultaneously, PEPR1 and PEPR2 loss of function mutants show enhanced JA, SA accumulation in response to CWI impairment (Engelsdorf et al. 2018). These results indicate that PTI and CWI maintenance-controlled processes are coordinated and PEP-based signaling processes are required for this coordination. One possible reason for this coordination could be that CWI maintenance acts as back-up defense mechanism activating phytohormone-based defenses in case the normal PTI-based responses are inhibited by pathogen-derived modulators.
DAMPs can also be cellulose-derived, since treatments with cellulose fragments (disaccharide to heptasaccharide) result in activation of PTI in a similar manner as OGs (Johnson et al. 2018; Souza et al. 2017). The most active compound is cellotriose, which induces ROS production, phosphorylation of MAPKs and expression of defense genes (Johnson et al. 2018; Locci et al. 2019). Such fragments could be generated by LYTIC POLYSACCHARIDE MONOOXYGENASES (LPMOs), enzymes used by phytopathogens to cleavage cellulose polymers (Vaaje-Kolstad et al. 2010). Two LRR-RLKs, namely STRESS INDUCED FACTOR 2 and 4 (SIF2 and SIF4), were implicated as critical factors in response to LPMO. LPMO treatments led to enhanced levels of ethylene, JA, SA, camalexin and increased resistance to the necrotrophic fungus Botrytis cinerea (Zarattini et al. 2021). In addition, physical interaction of SIF2 with the FLS2-BAK1 complex activates PTI signaling (Chan et al. 2020; Yuan et al. 2018). This results in direct phosphorylation of the SLOW ANION CHANNEL1 (SLAC1), which is necessary for abscisic acid (ABA)-mediated stomatal closure that enhances bacterial resistance (Chan et al. 2020).
One of the most studied groups of DAMPs are pectin-derived OGs, which activate PTI (Nothnagel et al. 1983) and were the first DAMPs discovered (Hahn et al. 1981). The core element of plant pectin is a polymer homogalacturonan (HG) that consists of α-1–4 linearly linked galacturonic acid molecules, which are methylesterified on C6 and often acetylesterified on C2 and/or C3 residues (Ridley et al. 2001). Production of OGs could be based on polygalacturonases that hydrolyze the α-1,4 linkages of low methyl-esterified homogalacturonans and have a substrate preference for linear polygalacturonic acids (Benedetti et al. 2015). However, a more recent report showed that treatment with Botrytis cinerea resulted in 80% of OGs being produced by pathogen-derived pectin lyases and acetyl- and methylesterified (Voxeur et al. 2019). Importantly, HG turnover also generates OGs, which could act as DAMPs starting immune responses or also have a signaling role during development (Denoux et al. 2008; Moscatiello et al. 2006). OGs promote expression of genes encoding chitinases, glucanases, and polygalacturonase-inhibiting proteins (De Lorenzo et al. 2001; Ridley et al. 2001); drastic increase of ROS production, accumulation of phytoalexins, activation of phospholipase C, CDPK and the cytoplasmic kinases ROG1 and ROG2 leading to increased expression of defense-related genes (Delteil et al. 2016; Ferrari et al. 2007; Galletti et al. 2008; Kohorn et al. 2016). The activity of OGs is dependent on the length, with a molecule size of 10–15 monomers apparently forming the most active forms (Ferrari et al. 2007). The pectin-hydrolyzing enzyme ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE 1 (ADPG1), which is required during development (Ogawa et al. 2009), has been reported recently to be upregulated in lignin-modified plants (Gallego-Giraldo et al. 2020). The changes in lignin composition apparently initiate a signaling cascade involving the CWI maintenance mechanism and induction of ADPG1 expression, releasing DAMPs (possibly OGs) and initiating immune responses (Voxeur and Höfte 2020). OGs also bind to WAK1 and 2, key regulators of cell expansion, biotic stress responses, wounding and metal tolerance (Kohorn et al. 2006; Tripathi et al. 2021). However, WAKs bind not only small pectin fragments but also long polymers cross-linked by Ca2+ in the cell wall and activate pectin-induced defense responses raising questions about specific functions of particular pectin types (Kohorn et al. 2009). To keep the deleterious effects of OGs under control, Arabidopsis encodes Berberine bridge enzyme-like (BBE-like) proteins whose oxidative activity specifically targets and inactivates the eliciting activity of non-oxidized OGs (Benedetti et al. 2018). Modification of OGs converts reducing galacturonic acids to oxidized galactaric acids. The change in homogalacturonan fragments retards degradation and use by microbial pathogens as carbon sources (Benedetti et al. 2018). It is unknown whether other molecules use similar oxidative mechanisms to regulate signaling and metabolic turn over of cell walls.
Transcriptional regulation of cell wall metabolism
Transcriptional regulation acting downstream of receptor-like kinases, small peptides or P/DAMPS has been linked mostly to immune responses. Examples are the interaction between FER and RALF peptides which lead to phosphorylation of the transcription factor (TF) MYC2 influencing JA signaling (Guo et al. 2018); the regulation of EDS1 and PAD4 by WAKs mediated by MAPKs mentioned above (Kohorn et al. 2016) and the activation of defense-related genes by P/DAMPS (B. Li et al. 2016a, b, c). The production of callose regulated by the TF MYB51 in response to flg22 illustrates how PTI regulates a particular element of cell wall metabolism (Clay et al. 2009). Many forms of CWD, both abiotic and biotic, activate common signaling processes involving hormone crosstalk, ROS and calcium signaling, meaning that the transcriptional machinery, controlled by these signaling processes, will be activated in response to CWD as well (Le Gall et al. 2015; Li et al. 2016a, b, c; Liao et al. 2017; Novaković et al. 2018). While CWI maintenance is likely to control transcriptional regulation of genes involved in cell wall metabolism, the mechanisms linking CWD perception to transcriptional changes remain to be elucidated. In addition to input from CWI monitoring, transcriptional regulation of cell wall biosynthesis genes needs to also integrate inputs deriving from perception of other stresses and developmental cues (Fig. 3). This ensures correct synthesis and delivery of cell wall components for specific cell types, developmental stages and during interactions with the environment (Kozlova et al. 2020; Rao and Dixon 2017; Wang et al. 2016; Wolf et al. 2012).

Transcriptional regulation of genes involved in responses to cell wall metabolism. An overview of relevant molecular components and structures in the cytoplasm and nucleus. Stress leads to generation of signals, which are relayed to the nucleus to modulate expression of transcriptional regulators, controlling expression of genes mediating responses to CWI impairment and biotic stress
Little is known about transcriptional regulation during primary cell wall formation. In Arabidopsis, the Ethylene Response Factors (ERF) TFs ERF034, ERF035, ERF038 and ERF039 belong to group IIId of the ERF APETALA2/ETHYLENE-RESPONSIVE ELEMENT BINDING PROTEIN (AP2/EREBP) supergene family and function as transcriptional activators of CESA1, CESA3 and CESA6, which are involved in primary (but not secondary) cell wall formation (Saelim et al. 2019). Ectopic expression of ERFs 035–038 and ERF040 (belonging to groups IIId and IIIe) in the double mutant nac secondary wall thickening promoting factor 1 and 3 (nst1and nst3), lacking secondary cell walls, produced multilayered, thickened fiber cell walls without ultra violet autofluorescence indicative for the presence lignin, therefore resembling primary cell walls (Sakamoto et al. 2018).
Secondary cell wall regulation is better understood and involves coordination of multiple regulatory layers consisting of distinct classes of transcription factors (Kumar et al. 2016). Using high-spatial-resolution gene expression data and enhanced yeast one-hybrid assays, a gene regulatory network involving TFs and enzymes involved in secondary cell wall biosynthesis was mapped (Taylor-Teeples et al. 2015). E2 FACTOR C (E2Fc, from the E2F family of TFs, whose members are key regulators of the cyclinD/retinoblastoma/E2F pathway) seems to act upstream of the several TF families acting as secondary cell wall formation regulators: NAM (NO APICAL MERISTEM), ATAF (ARABIDOPSIS TRANSCRIPTION ACTIVATION FACTOR), CUC (CUP-SHAPED COTYLEDON) and VASCULAR-RELATED NAC DOMAIN 6 and 7 (VND6 and VND7) belonging to the NAC family of TFs. Moderate levels of E2FC activate VND7 whereas extremely low or high levels of E2FC repress VND7 expression, in agreement with previous reports showing that E2FC can act as a transcriptional activator and repressor (de Jager et al. 2001; del Pozo et al. 2007; Heckmann et al. 2011; Kosugi and Ohashi, 2002). TFs belonging to this regulatory network were further classified in tiers, based on their regulatory hierarchy (Kumar et al. 2016). Tier 1 genes regulate transcription of structural genes, tier 2 regulate tier 1 and structural genes whereas tier 3 regulate tier 1 and 2 genes. VND6 and VND7 are positioned high in the hierarchy and together with NST1, NST2 and SECONDARY WALL-ASSOCIATED NAC DOMAIN PROTEIN 1 (SND1) form Tier 3. Tiers 1 and 2 contain TFs of the MYB TF family-like KNOTTED1-like homeobox (KNOX). TFs from the Homeodomain-leucine Zipper class III (HD-ZIP III) family are also positioned high in the regulatory hierarchy and are important for cell differentiation (Du and Wang, 2015). HD-ZIP III expression is regulated by microRNAs miR165/166, ensuring proper vasculature patterning (Fig. 3; Carlsbecker et al. 2010; Miyashima et al. 2011). These transcriptional networks regulate genes required for cell wall formation include cellulose biosynthesis genes such as CESA4, CESA7 and CESA8 or genes involved in hemicellulose and lignin biosynthesis. These selected examples provide some perspective on the complexity of the transcriptional networks responsible for regulation of cell wall metabolism and form the foundation to be used for understanding the mode of action of the corresponding processes in other plant species. More importantly, the examples also highlight our lack of knowledge regarding the mechanisms connecting initial perception of CWI impairment with transcriptional regulation of metabolic processes bringing about changes in structure and composition of cell walls, in particular primary cell walls.
Epigenetic regulation of cell wall biosynthesis
Epigenetic control is important for cell fate maintenance by tissue-specific regulation of gene expression during differentiation processes (Lafos et al. 2011). Epigenetic modifications are induced by abiotic and biotic factors and can result in improved long-term adaptability of plants to unfavorable environmental conditions (Kumar et al. 2017). Epigenetic regulation in dicots and especially in Arabidopsis thaliana has been extensively studied in the context of development and stress responses (Liang et al. 2020; Pikaard and Mittelsten Scheid, 2014).
The acetyltransferase GENERAL CONTROL NONDEREPRESSIBLE 5 (GCN5) modulates directly expression of genes required for cell wall loosening such as CHITINASE-LIKE 1 (CTL1), POLYGALACTURONASE INVOLVED IN EXPANSION 3 (PGX3), and MYB DOMAIN PROTEIN 54 (MYB54) in response to salt stress (Zheng et al. 2019). The importance is confirmed by GCN5 knockout mutants resulting in dwarfed plants, deformed flowers, decreased root length, and reduced cellulose content (Hu et al. 2015; Zheng et al. 2019). GCN5 binds directly to sequences in the promoter of CTL1, which mediates interactions between cellulose microfibrils and hemicelluloses (Zheng et al. 2019). The H3K4-histone methyltransferase ARABIDOPSIS HOMOLOG of TRITHORAX1 (ATX1) acts as a positive regulator on the secondary cell wall formation during stem growth by activating expression of NAC TF family members SECONDARY WALL-ASSOCIATED NAC DOMAIN PROTEIN1 (SND1) and NAC SECONDARY WALL THICKENING PROMOTING FACTOR1 (NST1; Wang et al. 2021). EXPANSIN (EXP) genes play an essential role during fruit ripening and their expression is regulated by histone modifications such as H3K9/K14 acetylation and H3K27 tri-methylation (Mu et al. 2021). While the available evidence indicates that regulation of cell wall metabolism by epigenetic modification happens, neither are the underlying principals and regulatory mechanisms well understood, nor do we know to what extent this regulatory mechanism is relevant in the context of CWI-mediated changes in cell wall metabolism.
Regulation of CWI maintenance in other plant species
Receptor-like kinases: abundance and diversification in different plant species
Homologues of Arabidopsis RLK genes have been identified in other plant species (Couto and Zipfel, 2016; Honkanen et al. 2016), but conservation and diversification of their specific functions remain largely unexplored (Fig. 4). A recent study suggests that the CrRLK1L kinase signaling pathway is specific for land plants, since analysis of algae genomes identified neither orthologues for CrRLK1 members nor for RALFs peptides (Mecchia et al. 2020). The only CrRLK1L kinase orthologue (MpFER) found in Marchantia polymorpha, exhibits the highest similarity to FER from Arabidopsis, highlighting again its particular importance (despite having been referred to as MpTHE before) (Honkanen et al. 2016; Mecchia et al. 2020). MpFER is required for correct rhizoid formation and expansion, maintaining the morphological integrity of the gametophyte and plant fertility, biological processes similar to those requiring AtFER. However, MpFER does not seem to repress cellular growth after CWI impairment, as AtTHE1. Isoxaben, a cellulose biosynthesis inhibitor, still affected the development of the gemmae in wild type and RNA interference lines targeting MpFER (Mecchia et al. 2020), suggesting that the CWI maintenance function arose later during plant evolution.

Receptors and biotic stress responses in other plants. Similar strategies to sense CWI impairment seem to be active in Arabidopsis and other plants. Since the number of family members encoding membrane-bound receptors and peptides has diversified in other plants, the specific interactions between receptors, co-receptors and ligands as well as the downstream effects of CWI impairment, can differ from the ones observed in Arabidopsis. FLR1 and FLR2 (Oryza sativa) are homologues of AtFER with specialized functions in growth and immunity. WAKs family members have increased and specialized in crops including Brachypodium, Zea maize, wheat and Oryza sativa; where they function in diverse developmental and immune responses. Similar to their function in Arabidopsis, PEPs and PEPRs have been implicated in defense responses in Zea maize and Oryza sativa. SIT1 (L-type LecRLK) and CEBiP/CERK complex and interactors represented in the figure are based on Oryza sativa studies
The two main rice varietal groups Oryza sativa indica and Oryza sativa japonica contain the same number of CrRLK1L kinases as Arabidopsis but very little information exists regarding their respective functions (Yang et al. 2020). FERONIA-LIKE RECEPTOR 2 and 11 (FLR2 and FLR11) seem to participate in plant immunity responses since flr2 and flr11 knock-out mutants exhibit enhanced resistance to rice blast infection without significant negative impact on growth. ROS production was increased at the penetration site after rice blast inoculation in leaf cells, while reduced ROS production and bigger lesion areas were observed in FRL2 overexpression lines (Yang et al. 2020). FRL2 seems to have a critical role in plant height, while FLR1 has a minor role but is required for fertility instead. FLR1 and FLR2 both positively regulate expression of genes encoding enzymes (ENT-KAURENE OXIDASE 2, GIBERELLIN 20-OXIDASE 2 and GIBERELLIN 30-OXIDASE 2) required for Gibberellin biosynthesis (Li et al. 2016a, b, c). Single mutants of the enzymes showed reduced shoot length, which was rescued to different extents by introducing flr1 and flr2, suggesting that FLR1 and FLR2 are involved in different regulatory pathways.
A comprehensive genome-wide comparative analysis of the CrRLK1L subfamily in 62 species found that on average eudicots contain 22 CrRLK1L proteins whereas monocots only 13, possibly explained by the larger genome size of eudicots (Solis-Miranda et al. 2020). Additionally, a comparison between Arabidopsis and four legume species revealed that expression patterns of most CrRLK1L genes were conserved despite the increased number of members in legumes (especially in the MEDOS clade). Based on the expression patterns of several legume CrRLK1Ls they have been implicated in root nodulation.
The number of WAK kinases has increased profoundly in other plant species compared to Arabidopsis. Whereas this family consists only of 5 WAKs and 21 WAKLs in Arabidopsis (the distinction between WAKs and WAKLs is less clear in other species), the number has increased to 29 in cotton, 125 in rice, 341 in wheat, more than 100 in maize, 91 in Barley and 115 in Brachypodium distachyon (hereafter Brachypodium)(Dou et al. 2021; Tripathi et al. 2021; Wu et al. 2020; Zhang et al. 2021). Their ability to interact with pectin has been confirmed independently for several species, despite the low pectin abundance in grass cell walls (about 20% in primary cell walls of Arabidopsis but only 5–10% in grasses; Chen et al. 2021a, b; Gigli-Bisceglia et al. 2020; Wu et al. 2020) suggesting that WAKs have a highly conserved role in connecting pectic compounds in plant cell walls to intracellular responses. In Brachypodium, high expression levels of several WAK genes (specifically BdWAK2, 10, 42, 72 and 108) in rapidly growing tissue implicates them in cell expansion (Wu et al. 2020). Vacuolar invertase-dependent regulation of turgor pressure was hypothesized to be the mechanism responsible for WAK-mediated cell wall expansion in Arabidopsis but remains to be experimentally confirmed (Kohorn 2016; Kohorn et al. 2012). While WAKs have been implicated in abiotic and biotic stress responses by activating phytohormone biosynthesis, such as JA, and hypersensitive responses signaling cascades leading to programmed cell death, as exemplified by the activity of BdWAK2 (Wu et al. 2020), the mechanisms responsible remain unknown. Interestingly, manipulation of wheat WAK and WAK-like genes confers pathogen resistance by different mechanisms. Wheat WAK2 enhances resistance to the fungus Fusarium graminearum through interactions with pectin (Gadaleta et al. 2019). In contrast, gene-for-gene resistance to the fungus Zymoseptoria tritici producing Septoria tritici blotch (STB) is provided by WAKL4, also named Stb6, through recognition of apoplastic avirulent effectors (Saintenac et al. 2018).
Relevant signaling processes during response to biotic stress
The mode of action of the signaling cascades activated by flagellin in Arabidopsis and rice exhibits similarities (Fig. 4). The CrRLK1L FERONIA family in rice has 12 FERONIA-LIKE RECEPTORS (OsFLR) members (Yang et al. 2020). Overexpression of OsFLS2 results in enhanced immune responses to flg22 and flagellin (Takai et al. 2008). OsFLR1 has a crucial role in parasitism of Meloidogyne incognita where it interacts with parasite-derived RALFs and manipulates plant cell expansion and cell wall modification to facilitate parasitism (Zhang et al. 2020a). This is in contrast to the situation in Triticum aestivum (Ta), where transient silencing of TaFER1 and TaFER2 had no obvious impact on Fusarium graminearum infection (Wood et al. 2020).
In contrast, the mechanism modulating the response to chitin in rice differs from the one in Arabidopsis. Fungus derived chitin is recognized by LysM-RLP CHITIN OLIGOSACCHARIDE ELICITOR-BINDING PROTEIN (OsCEBiP), which afterward binds to (OsCERK1) whereas in Arabidopsis, AtCERK1 binds chitin directly. The OsCEBiP/OsCERK1 complex in turn activates signaling components such as OsRacGEF (Akamatsu et al. 2013; Malinovsky et al. 2014; Shimizu et al. 2010). OsRAC1 is a small GTPase that activates immune responses such as ROS production by interacting with NADPH oxidase and a ROS scavenger to modulate the extent of ROS production (Wong et al. 2007). For proper response, both HEAT SHOCK PROTEIN 90 and its co-chaperone HSP70-HSP90 ORGANIZING PROTEIN (HOP) / STRESS-INDUCED PROTEIN 1 (STI1) are required because they are responsible for efficient transport of OsCERK1 and OsRAC1GEF from the endoplasmatic reticulum to the plasma membrane (Akamatsu et al. 2013; Chen et al. 2010; Nakashima et al. 2008). OsRAC1 is phosphorylated and activates then immune responses (including lignin biosynthesis involving CINNAMOYL-COA REDUCTASE 1 (OsCCR1)) through a MAPK-signaling module (Akamatsu et al. 2013; Kawasaki et al. 2017, 2006; Tang et al. 2017). RLPs like RECEPTOR-LIKE CYTOPLASMIC KINASE 185 (OsRLCK185) and RECEPTOR-LIKE CYTOPLASMIC KINASE 176 (OsRLCK176) are also positive regulators of immune responses in rice and interact directly with the OsCEBiP/OsCERK1 complex (Ao et al. 2014; Yamaguchi et al. 2019, 2013). Since PTI-controlled responses activated by chitin are quite pronounced, pathogens try to suppress them by secretion of LysM-containing proteins, which bind chitin oligomers creating complexes to avoid chitin recognition by the plant (de Jonge et al. 2010; Mentlak et al. 2012).
The LPS protein family has 75 members (32 G-type, 42 L-type, and 1 C-type) in Arabidopsis, and 173 members (100 G-type, 72 L-type, and 1 C-type) in rice (Vaid et al. 2012). In rice, ABNORMAL POLLEN 1 (AP1) belongs to the L-type LecRLKs and modulates carbohydrate metabolism required for pollen maturation (He et al. 2021). Another L-type LecRLK (SALT INTOLERANCE 1, SIT1) mediates salt-induced, ethylene-based signaling processes by modulating ethylene production and promoting ROS accumulation in rice (Li et al. 2014a, b). A G-type lecRLK is required for interactions between Populus trichocarpa and the ectomycorrhizal fungus Laccaria bicolor (Labbé et al. 2019). Simultaneously, it was shown that expression of LecRLK1 from Populus trichocarpa in non-host switchgrass roots allows colonization by Laccaria bicolor and improves productivity of this potential bioenergy crop exposed to abiotic stress (Qiao et al. 2021).
In Zea mays, ZmWAK genes play essential roles during infection by fungal pathogens Sporisorium reilianum and Exserohilum turcicum by regulating biosynthesis of defense-related benzoxazinoids (Hurni et al. 2015; Yang et al. 2019a, b; Zhang et al. 2017). Mutations in OsWAK genes (such as Xa4) lead to enhanced resistance against the bacterial blight (Xanthomonas oryzae pv. oryzae-Xoo), increased cell wall strength while reducing plant height to a small extent and not interfering with grain yield (Krattinger and Keller, 2017; Ning et al. 2017). In wheat, TaWAK7D is modulating resistance against Rhizoctonia cerealis, by controlling expression of several pathogenesis-related genes (Qi et al. 2021). Comparative transcriptome analysis showed that the OsWAK activates TFs belonging to the bZIP, WRKY, MYB, DOF, and HSF families during bacterial infection, suggesting the TFs may be responsible for the enhanced resistance (Bakade et al. 2021).
DAMPs like the tri-saccharide 31-β-d-Cellobiosyl-glucose or the tetrasaccharide 31-β-d-Cellotriosyl-glucose arise when pathogen-derived endoglucanases digest rice hemicellulose. They also activate PTI in rice via OsCERK1 (Mélida et al. 2018; Yang et al. 2021). In rice, OGs do also activate WAK-mediated responses to biotic stress. These responses include enhanced cellulose biosynthesis (Hu et al. 2017), ROS production, and expression of genes involved in pathogen response (Delteil et al. 2016).
PEP-mediated processes have also been described in monocots (Lori et al. 2015). Infection with Cochliobolus heterostrophus and Colletotrichum graminicola activates ZmPEPR1, increases expression of defense-related genes and metabolites, while ZmPEP1 binds to ZmPEPR1 (Huffaker et al. 2011; Lori et al. 2015). Exogenous application of OsPEPs elicits multiple defense responses in rice cell cultures (Shinya et al. 2018). Treatment of rice cells with OsPep3 during infection by Mythimna loreyi enhances defense responses, including activation of MAPKs and production of defense-related hormones and metabolites (Shinya et al. 2018). At the same time, OsPEPR1 overexpression increases the sensitivity of rice plants to these stress signals (Shinya et al. 2018).
Transcriptional regulation of cell wall metabolism in different species
Our current knowledge regarding the regulation of primary cell wall formation in plants is based mainly on studies of Arabidopsis with the obvious consequence being that the process remains unexplored in many other species. Interestingly, coexpression experiments suggest that the rice homologue of AtERF34 is involved in secondary cell wall regulation, instead of primary cell wall regulation as in Arabidopsis (Hirano et al. 2013). Similar to Arabidopsis, transcriptional regulation of secondary cell wall formation in other plant species requires a diverse set of TF families including NAC, MYB, HD-ZIP III and KNOX (Kumar et al. 2016; Taylor-Teeples et al. 2015).
In rice, the KNOX TF OsKNAT7 coordinates secondary cell wall biosynthesis and cell expansion via different interactors and downstream factors compared to its Arabidopsis homologue, as also indicated by the differences in mutant phenotypes observed (Wang et al. 2019; Yu 2019a). Thicker walls in rice are caused by higher cellulose and xylan content, whereas in Arabidopsis this is caused by increased lignin. Additionally, rice mutants exhibit larger grain size and cells in spikelet bracts, effects not observed in Arabidopsis. Rice KNAT7 acts upstream of MYB61, thereby inhibiting secondary cell wall biosynthesis whereas in Arabidopsis KNAT7 is active downstream of MYB61 (Wang et al. 2019; Yu 2019b). It is unclear whether a feedback loop exists between both TFs or if secondary cell wall regulation evolved differently across species. OsKNAT7 interacts with GROWTH REGULATING FACTOR 4 (GRF4) and NAC31 to repress cell expansion and wall thickness. Furthermore, the cotton (Gossypium hirsutum) gene KNOTTED1-LIKE, member of the GhKNOX family, regulates fiber development, initiation and elongation possibly by forming heterodimers with OVATE FAMILY PROTEIN 4 (Gong et al. 2014).
In Eucalyptus trees (Eucalyptus grandis), the NAC TF EgNAC141 activates genes required for lignin biosynthesis (Sun et al. 2021). EgNAC141 belongs to a group of genes identified through a combination of phylogenetics and large-scale expression profiling (Hussey et al. 2015). These genes are preferentially expressed in the xylem and have no Arabidopsis orthologues. NAC TFs involved in lignin production are characterized by their tissue- or cell-specific expression, as exemplified by EgNAC141, which is apparently expressed 1000 times higher in the xylem and stem than in other tissues. Another NAC-domain containing protein, Populus trichocarpa PdWND3A, which seems to be the homologue of the Arabidopsis AtVND4 and 5 TFs, regulates vessel size in the stem (Yang et al. 2019a, b). Upregulation of ferulate 5-hydroxylase 1 gene expression in PdWND3A overexpressing lines increases lignin content. The enzyme encoded mediates the chemical conversion from coniferaldehyde to 5-OH coniferaldehyde in the syringil monolignol biosynthesis pathway. It remains unclear if particular aspects of lignin biosynthesis are specifically regulated by PdWND3A as both lignin biosynthesis and the ratio of lignin monomers syringil/guaiacyl are affected.
Expression of the TF HD-ZIP III family is regulated by microRNAs in Arabidopsis and rice (Carlsbecker et al. 2010; Miyashima et al. 2011; Zhang et al. 2018). OsmiR166b is located in a yield-related QTL interval and affects rice grain yield (Fang et al. 2013). One of its targets, OsHox32, regulates lignin and cellulose biosynthesis by suppressing expression of the cell wall biosynthetic genes CINNAMYL ALCOHOL DEHYDROGENASE 2 (OsCAD2) and OsCESA7 (H. Chen et al. 2021a, b). Knockdown lines of OsmiR166b and overexpression of OsHox32 exhibit culms with cavities, brittle culms and reduced cell wall thickness in leaves. These results suggest that OsmiR166b and OsHox32 form a regulatory module modulating culm formation. These selected examples illustrate how the functions of several TFs are conserved between different plant species, while others differ (Feller et al. 2011; Jiang, 2019; Wang et al. 2018a, b; Yokoyama and Nishitani, 2004). However, it is important to note that we observe both cases where individual genes have changed activities (ERF34) and where organisation of gene activities (KNAT7/MYB61) are modified.
Epigenetic regulation of cell wall metabolism in crop plants
Epigenetic regulation in dicots and especially in Arabidopsis thaliana has been extensively studied in the context of development and stress responses (Yamaguchi 2019). In contrast, knowledge regarding epigenetic processes in monocots is limited. In protoplasts from Oryza, chromatin decondensation/reorganization and histone modification seem to be tightly connected to proteins required for de novo formation of cell walls (Mujahid et al. 2013; Tan et al. 2011). However, the mechanism regulating de novo wall formation in protoplasts differs probably from the normal situation, where the synthesis of new cell walls occurs during cell division, raising questions to what extent the regulatory mechanisms are the same (Mujahid et al. 2013). Salinity stress increased in Zea mays expression of histone acetyltransferase genes such as HISTONE ACETYLTRANSFERASES B (ZmHATB) and ZmGCN5, which can result in increased acetylation of H3K9 on at histone H4K5 (Li et al. 2014a, b). The increased histone acetylation is associated with an enhanced expression of EXPANSIN B2 (ZmEXPB2) and XYLOGLUCAN ENDOTRANSGLYCOSYLASE (ZmXET1) genes (Li et al. 2014a, b; Wolny et al. 2021). These results are intriguing because salinity stress induces mainly elevated expression of ZmEXPB2, ZmEXPB6, and ZmEXPB8 isomers (Geilfus et al. 2010). In wheat, increased dimethylation at histone H3K9 and decreased levels of trimethylation at histone H3K4 and acetylation at histone H3K9 lead to a negative impact on transcriptional modulation of three EXPANSIN A1 (TaEXPA1) homologs, gene silencing (Hu et al. 2013). Thus, monocots seem to sustain stable expression of β-Expansins in response to stress through epigenetic modification. In rice, GCN5 is highly expressed in the root meristem and is required for cell division and growth (Zhou et al. 2017). GCN5 is recruited by WUSCHEL (WUS)-RELATED HOMEOBOX11 (WOX11) and regulates root-specific genes involved in energy metabolism, hormone response and cell wall biosynthesis. To summarize the currently available information indicates that epigenetic modification is relevant for regulation of at least certain aspects of cell wall metabolism and that research opportunities exist to explore the mechanisms responsible.
Conclusions
By monitoring the status of their cell walls, plants are able to adapt successfully to adverse environmental conditions. Sensing environmental stimuli at the wall triggers cellular responses responsible for successful adaptation to the surrounding environment. Progress has been made in Arabidopsis to identify the function of several CWI maintenance sensors and the processes they regulate. Nevertheless, the molecular mechanisms, interacting partners and regulatory pathways of most of the putative CWI sensors have still not been elucidated in detail. AtFER function has been characterized extensively mainly because of the pleiotropic effects observed in the loss-of-function mutants, indicating that this protein has key functions in many different biological processes. This is to be expected bearing in mind the contributions of cell walls to many different aspects of plant life. AtTHE1 has also received attention because of its apparently more specific function in CWI maintenance in response to CWD. Interestingly, amongst higher plants, the CrRLK1L gene family has fairly conserved numbers and expression domains for its members, suggesting that their function might also be conserved. In contrast, the number of WAK family members, which also seem to be able to detect cell wall impairment, is increased significantly in other plant species compared to Arabidopsis. However, WAKs and WAKLs continue to be involved in cell elongation and responses to biotic stress mechanisms in different plant species. Therefore, the reason for the increased number of genes and potential benefits of having them remains an open question. One possibility is that expression of several of these genes is restricted to specialized tissues and organs not found in other species.
Knowledge regarding the mode of action of the CWI maintenance mechanism and its relevance in other plant species is in its infancy. Larger genomes and multiple homologous genes, likely with redundant function, will pose difficulties in identifying the key molecular components for CWI maintenance. Investigating both ends of the evolutionary tree, close species to Arabidopsis on one side and ferns on the other, should help us discover common molecular mechanisms involved in the response to CWD and CWI maintenance and more importantly identify opportunities to use the resulting knowledge to improve performance of food and bioenergy crop plants.
In Arabidopsis, our knowledge regarding transcriptional regulation of primary cell wall metabolism has started to grow recently. This is exemplified by the ERF family of transcription factors, which regulate expression of CESA genes, thus controlling cellulose production. Regarding other metabolic processes required for primary cell wall formation, our understanding could be improved. However, comparing this knowledge to information regarding transcriptional regulation of the same metabolic processes in other species, highlights that the situation is there even worse, thus creating exciting research opportunities. In contrast, information, knowledge and understanding regarding transcriptional regulation of secondary cell wall metabolism in Arabidopsis and other plants is much more advance as exemplified by our knowledge regarding the networks of TFs from different families acting in a hierarchical manner to control secondary cell wall formation. For several Arabidopsis homologues the functions seem to be preserved in other species suggesting they could form leads to analyse the regulatory processes in these species in a knowledge-based manner.
Data availability
Not applicable.
Code availability
Not applicable.
References
Abdul Malik NA, Kumar IS, Nadarajah K (2020) Elicitor and receptor molecules: orchestrators of plant defense and immunity. Int J Mol Sci 21:963. https://doi.org/10.3390/ijms21030963
Akamatsu A, Wong HL, Fujiwara M, Okuda J, Nishide K, Uno K, Imai K, Umemura K, Kawasaki T, Kawano Y, Shimamoto K (2013) An OsCEBiP/OsCERK1-OsRacGEF1-OsRac1 module is an essential early component of chitin-induced rice immunity. Cell Host Microbe 13:465–476. https://doi.org/10.1016/j.chom.2013.03.007
Alonso-Simón A, García-Angulo P, Mélida H, Encina A, Álvarez JM, Acebes JL (2011) The use of FTIR spectroscopy to monitor modifications in plant cell wall architecture caused by cellulose biosynthesis inhibitors. Plant Signal Behav 6:1104–1110. https://doi.org/10.4161/psb.6.8.15793
Ao Y, Li Z, Feng D, Xiong F, Liu J, Li J-F, Wang M, Wang J, Liu B, Wang H-B (2014) OsCERK1 and OsRLCK176 play important roles in peptidoglycan and chitin signaling in rice innate immunity. Plant J 80:1072–1084. https://doi.org/10.1111/tpj.12710
Bacete L, Hamann T (2020) The role of mechanoperception in plant cell wall integrity maintenance. Plants. https://doi.org/10.3390/plants9050574
Bacete L, Mélida H, Miedes E, Molina A (2018) Plant cell wall-mediated immunity: cell wall changes trigger disease resistance responses. Plant J 93:614–636. https://doi.org/10.1111/tpj.13807
Bakade R, Ingole KD, Deshpande S, Pal G, Patil SS, Bhattacharjee S, Prasannakumar MK, Ramu VS (2021) Comparative transcriptome analysis of rice resistant and susceptible genotypes to Xanthomonas oryzae pv. oryzae identifies novel genes to control bacterial leaf blight. Mol Biotechnol 63:719–731. https://doi.org/10.1007/s12033-021-00338-3
Bartels S, Boller T (2015) Quo vadis, Pep? Plant elicitor peptides at the crossroads of immunity, stress, and development. J Exp Bot 66:5183–5193. https://doi.org/10.1093/jxb/erv180
Basu D, Haswell ES (2020) The mechanosensitive ion channel MSL10 potentiates responses to cell swelling in Arabidopsis seedlings. Curr Biol 30:2716-2728.e6. https://doi.org/10.1016/j.cub.2020.05.015
Bellande K, Bono J-J, Savelli B, Jamet E, Canut H (2017) Plant lectins and lectin receptor-like kinases: how do they sense the outside? Int J Mol Sci 18:E1164. https://doi.org/10.3390/ijms18061164
Benedetti M, Pontiggia D, Raggi S, Cheng Z, Scaloni F, Ferrari S, Ausubel FM, Cervone F, De Lorenzo G (2015) Plant immunity triggered by engineered in vivo release of oligogalacturonides, damage-associated molecular patterns. Proc Natl Acad Sci USA 112:5533–5538. https://doi.org/10.1073/pnas.1504154112
Benedetti M, Verrascina I, Pontiggia D, Locci F, Mattei B, De Lorenzo G, Cervone F (2018) Four Arabidopsis berberine bridge enzyme-like proteins are specific oxidases that inactivate the elicitor-active oligogalacturonides. Plant J 94:260–273. https://doi.org/10.1111/tpj.13852
Bi G, Zhou Z, Wang W, Li L, Rao S, Wu Y, Zhang X, Menke FLH, Chen S, Zhou J-M (2018) Receptor-like cytoplasmic kinases directly link diverse pattern recognition receptors to the activation of mitogen-activated protein kinase cascades in Arabidopsis. Plant Cell 30:1543–1561. https://doi.org/10.1105/tpc.17.00981
Boisson-Dernier A, Roy S, Kritsas K, Grobei MA, Jaciubek M, Schroeder JI, Grossniklaus U (2009) Disruption of the pollen-expressed FERONIA homologs ANXUR1 and ANXUR2 triggers pollen tube discharge. Development 136:3279–3288. https://doi.org/10.1242/dev.040071
Cantu D, Vicente AR, Labavitch JM, Bennett AB, Powell ALT (2008) Strangers in the matrix: plant cell walls and pathogen susceptibility. Trends Plant Sci 13:610–617. https://doi.org/10.1016/j.tplants.2008.09.002
Cao Y, Liang Y, Tanaka K, Nguyen CT, Jedrzejczak RP, Joachimiak A, Stacey G (2014) The kinase LYK5 is a major chitin receptor in Arabidopsis and forms a chitin-induced complex with related kinase CERK1. Elife. https://doi.org/10.7554/eLife.03766
Carlsbecker A, Lee J-Y, Roberts CJ, Dettmer J, Lehesranta S, Zhou J, Lindgren O, Moreno-Risueno MA, Vatén A, Thitamadee S, Campilho A, Sebastian J, Bowman JL, Helariutta Y, Benfey PN (2010) Cell signalling by microRNA165/6 directs gene dose-dependent root cell fate. Nature 465:316–321. https://doi.org/10.1038/nature08977
Chae HB, Kim MG, Kang CH, Park JH, Lee ES, Lee S-U, Chi YH, Paeng SK, Bae SB, Wi SD, Yun B-W, Kim W-Y, Yun D-J, Mackey D, Lee SY (2021) Redox sensor QSOX1 regulates plant immunity by targeting GSNOR to modulate ROS generation. Mol Plant 14:1312–1327. https://doi.org/10.1016/j.molp.2021.05.004
Chan C, Panzeri D, Okuma E, Tõldsepp K, Wang Y-Y, Louh G-Y, Chin T-C, Yeh Y-H, Yeh H-L, Yekondi S, Huang Y-H, Huang T-Y, Chiou T-J, Murata Y, Kollist H, Zimmerli L (2020) STRESS INDUCED FACTOR 2 regulates Arabidopsis stomatal immunity through phosphorylation of the anion channel SLAC1. Plant Cell 32:2216–2236. https://doi.org/10.1105/tpc.19.00578
Chen L, Hamada S, Fujiwara M, Zhu T, Thao NP, Wong HL, Krishna P, Ueda T, Kaku H, Shibuya N, Kawasaki T, Shimamoto K (2010) The Hop/Sti1-Hsp90 chaperone complex facilitates the maturation and transport of a PAMP receptor in rice innate immunity. Cell Host Microbe 7:185–196. https://doi.org/10.1016/j.chom.2010.02.008
Chen H, Fang R, Deng R, Li J (2021a) The OsmiRNA166b-OsHox32 pair regulates mechanical strength of rice plants by modulating cell wall biosynthesis. Plant Biotechnol. https://doi.org/10.1111/pbi.13565
Chen P, Giarola V, Bartels D (2021b) The Craterostigma plantagineum protein kinase CpWAK1 interacts with pectin and integrates different environmental signals in the cell wall. Planta 253:92. https://doi.org/10.1007/s00425-021-03609-0
Choi J, Tanaka K, Liang Y, Cao Y, Lee SY, Stacey G (2014) Extracellular ATP, a danger signal, is recognized by DORN1 in Arabidopsis. Biochem J 463:429–437. https://doi.org/10.1042/BJ20140666
Clay NK, Adio AM, Denoux C, Jander G, Ausubel FM (2009) Glucosinolate metabolites required for an Arabidopsis innate immune response. Science 323:95–101. https://doi.org/10.1126/science.1164627
Codjoe JM, Miller K, Haswell ES (2021) Plant cell mechanobiology: greater than the sum of its parts. Plant Cell. https://doi.org/10.1093/plcell/koab230
Couto D, Zipfel C (2016) Regulation of pattern recognition receptor signalling in plants. Nat Rev Immunol 16:537–552. https://doi.org/10.1038/nri.2016.77
de Jager SM, Menges M, Bauer M (2001) Arabidopsis E2F1 binds a sequence present in the promoter of S-phase-regulated gene AtCDC6 and is a member of a multigene family with differential activities. Plant Mol Biol 47(4):555–568. https://doi.org/10.1023/a:1011848528377
de Jonge R, van Esse HP, Kombrink A, Shinya T, Desaki Y, Bours R, van der Krol S, Shibuya N, Joosten MHAJ, Thomma BPHJ (2010) Conserved fungal LysM effector Ecp6 prevents chitin-triggered immunity in plants. Science 329:953–955. https://doi.org/10.1126/science.1190859
De Lorenzo G, D’Ovidio R, Cervone F (2001) The role of polygalacturonase-inhibiting proteins (PGIPs) in defense against pathogenic fungi. Annu Rev Phytopathol 39:313–335. https://doi.org/10.1146/annurev.phyto.39.1.313
Decreux A, Messiaen J (2005) Wall-associated kinase WAK1 interacts with cell wall pectins in a calcium-induced conformation. Plant Cell Physiol 46:268–278. https://doi.org/10.1093/pcp/pci026
DeFalco TA, Zipfel C (2021) Molecular mechanisms of early plant pattern-triggered immune signaling. Mol Cell 81:3449–3467. https://doi.org/10.1016/j.molcel.2021.07.029
del Pozo JC, Diaz-Trivino S, Cisneros N, Gutierrez C (2007) The E2FC-DPB transcription factor controls cell division, endoreplication and lateral root formation in a SCFSKP2A-dependent manner. Plant Signal Behav 2:273–274. https://doi.org/10.4161/psb.2.4.3897
Delteil A, Gobbato E, Cayrol B, Estevan J, Michel-Romiti C, Dievart A, Kroj T, Morel J-B (2016) Several wall-associated kinases participate positively and negatively in basal defense against rice blast fungus. BMC Plant Biol 16:17. https://doi.org/10.1186/s12870-016-0711-x
Denness L, McKenna JF, Segonzac C, Wormit A, Madhou P, Bennett M, Mansfield J, Zipfel C, Hamann T (2011) Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species- and jasmonic acid-dependent process in Arabidopsis. Plant Physiol 156:1364–1374. https://doi.org/10.1104/pp.111.175737
Denoux C, Galletti R, Mammarella N, Gopalan S, Werck D, De Lorenzo G, Ferrari S, Ausubel FM, Dewdney J (2008) Activation of defense response pathways by OGs and Flg22 elicitors in Arabidopsis seedlings. Mol Plant 1:423–445. https://doi.org/10.1093/mp/ssn019
Deslauriers SD, Larsen PB (2010) FERONIA is a key modulator of brassinosteroid and ethylene responsiveness in Arabidopsis hypocotyls. Mol Plant 3:626–640. https://doi.org/10.1093/mp/ssq015
Desprez T, Vernhettes S, Fagard M, Refrégier G, Desnos T, Aletti E, Py N, Pelletier S, Höfte H (2002) Resistance against herbicide isoxaben and cellulose deficiency caused by distinct mutations in same cellulose synthase isoform CESA6. Plant Physiol 128:482–490. https://doi.org/10.1104/pp.010822
Dievart A, Gottin C, Périn C, Ranwez V, Chantret N (2020) Origin and diversity of plant receptor-like kinases. Annu Rev Plant Biol 71:131–156. https://doi.org/10.1146/annurev-arplant-073019-025927
Dou L, Li Z, Shen Q, Shi H, Li H, Wang W, Zou C, Shang H, Li H, Xiao G (2021) Genome-wide characterization of the WAK gene family and expression analysis under plant hormone treatment in cotton. BMC Genomics 22:85. https://doi.org/10.1186/s12864-021-07378-8
Du Q, Wang H (2015) The role of HD-ZIP III transcription factors and miR165/166 in vascular development and secondary cell wall formation. Plant Signal Behav 10:e1078955. https://doi.org/10.1080/15592324.2015.1078955
Duan Q, Kita D, Li C, Cheung AY, Wu H-M (2010) FERONIA receptor-like kinase regulates RHO GTPase signaling of root hair development. Proc Natl Acad Sci USA 107:17821–17826. https://doi.org/10.1073/pnas.1005366107
Dünser K, Gupta S, Herger A, Feraru MI, Ringli C, Kleine-Vehn J (2019) Extracellular matrix sensing by feronia and Leucine-Rich Repeat Extensins controls vacuolar expansion during cellular elongation in Arabidopsis thaliana. EMBO J. https://doi.org/10.15252/embj.2018100353
Ellis C, Turner JG (2001) The Arabidopsis mutant cev1 has constitutively active jasmonate and ethylene signal pathways and enhanced resistance to pathogens. Plant Cell 13:1025–1033. https://doi.org/10.1105/tpc.13.5.1025
Engelsdorf T, Gigli-Bisceglia N, Veerabagu M, McKenna JF, Vaahtera L, Augstein F, Van der Does D, Zipfel C, Hamann T (2018) The plant cell wall integrity maintenance and immune signaling systems cooperate to control stress responses in Arabidopsis thaliana. Sci Signal 11:eaao3070. https://doi.org/10.1126/scisignal.aao3070
Escobar-Restrepo J-M, Huck N, Kessler S, Gagliardini V, Gheyselinck J, Yang W-C, Grossniklaus U (2007) The FERONIA receptor-like kinase mediates male-female interactions during pollen tube reception. Science 317:656–660. https://doi.org/10.1126/science.1143562
European Commission. Directorate-General for Research and Innovation (2018) A sustainable bioeconomy for Europe: strengthening the connection between economy, society and the environment. Updated Bioeconomy Strategy
Ezquer I, Salameh I, Colombo L, Kalaitzis P (2020) Plant cell walls tackling climate change: biotechnological strategies to improve crop adaptations and photosynthesis in response to global warming. Plants 9:212. https://doi.org/10.3390/plants9020212
Fang R, Li L, Li J (2013) Spatial and temporal expression modes of MicroRNAs in an elite rice hybrid and its parental lines. Planta 238:259–269. https://doi.org/10.1007/s00425-013-1881-5
Feller A, Machemer K, Braun EL, Grotewold E (2011) Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J 23:1–8
Feng W, Kita D, Peaucelle A, Cartwright HN, Doan V, Duan Q, Liu M-C, Maman J, Steinhorst L, Schmitz-Thom I, Yvon R, Kudla J, Wu H-M, Cheung AY, Dinneny JR (2018) The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca2+ signaling. Curr Biol 28:666-675.e5. https://doi.org/10.1016/j.cub.2018.01.023
Ferrari S, Galletti R, Denoux C, De Lorenzo G, Ausubel FM, Dewdney J (2007) Resistance to Botrytis cinerea induced in Arabidopsis by elicitors is independent of salicylic acid, ethylene, or jasmonate signaling but requires PHYTOALEXIN DEFICIENT3. Plant Physiol 144:367–379. https://doi.org/10.1104/pp.107.095596
Franck CM, Westermann J, Boisson-Dernier A (2018) Plant malectin-like receptor kinases: from cell wall integrity to immunity and beyond. Annu Rev Plant Biol 69:301–328. https://doi.org/10.1146/annurev-arplant-042817-040557
Gadaleta A, Colasuonno P, Giove SL, Blanco A, Giancaspro A (2019) Map-based cloning of QFhbmgb-2A identifies a WAK2 gene responsible for Fusarium Head Blight resistance in wheat. Sci Rep 9:6929. https://doi.org/10.1038/s41598-019-43334-z
Gallego-Giraldo L, Liu C, Pose-Albacete S, Pattathil S, Peralta AG, Young J, Westpheling J, Hahn MG, Rao X, Knox JP, Meester BD, Boerjan W, Dixon RA (2020) ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE 1 (ADPG1) releases latent defense signals in stems with reduced lignin content. Proc Natl Acad Sci USA 117:3281–3290. https://doi.org/10.1073/pnas.1914422117
Galletti R, Denoux C, Gambetta S, Dewdney J, Ausubel FM, De Lorenzo G, Ferrari S (2008) The AtrbohD-mediated oxidative burst elicited by oligogalacturonides in Arabidopsis is dispensable for the activation of defense responses effective against Botrytis cinerea. Plant Physiol 148:1695–1706. https://doi.org/10.1104/pp.108.127845
García-Angulo P, Alonso-Simón A, Encina A, Álvarez JM, Acebes JL (2012) Cellulose biosynthesis inhibitors: comparative effect on bean cell cultures. Int J Mol Sci 13:3685–3702. https://doi.org/10.3390/ijms13033685
Geilfus C-M, Zörb C, Mühling KH (2010) Salt stress differentially affects growth-mediating β-expansins in resistant and sensitive maize (Zea mays L.). Plant Physiol Biochem 48:993–998. https://doi.org/10.1016/j.plaphy.2010.09.011
Gigli-Bisceglia N, Engelsdorf T, Hamann T (2020) Plant cell wall integrity maintenance in model plants and crop species-relevant cell wall components and underlying guiding principles. Cell Mol Life Sci 77:2049–2077. https://doi.org/10.1007/s00018-019-03388-8
Giovannoni M, Marti L, Ferrari S, Tanaka-Takada N, Maeshima M, Ott T, De Lorenzo G, Mattei B (2021) The plasma membrane-associated Ca2+ -binding protein, PCaP1, is required for oligogalacturonide and flagellin-induced priming and immunity. Plant Cell Environ. https://doi.org/10.1111/pce.14118
Gong S-Y, Huang G-Q, Sun X, Qin L-X, Li Y, Zhou L, Li X-B (2014) Cotton KNL1, encoding a class II KNOX transcription factor, is involved in regulation of fibre development. J Exp Bot 65:4133–4147. https://doi.org/10.1093/jxb/eru182
Gonneau M, Desprez T, Martin M, Doblas VG, Bacete L, Miart F, Sormani R, Hématy K, Renou J, Landrein B, Murphy E, Van De Cotte B, Vernhettes S, De Smet I, Höfte H (2018) Receptor kinase THESEUS1 is a rapid alkalinization factor 34 receptor in Arabidopsis. Curr Biol 28:2452-2458.e4. https://doi.org/10.1016/j.cub.2018.05.075
Gouget A, Senchou V, Govers F, Sanson A, Barre A, Rougé P, Pont-Lezica R, Canut H (2006) Lectin receptor kinases participate in protein-protein interactions to mediate plasma membrane-cell wall adhesions in Arabidopsis. Plant Physiol 140:81–90. https://doi.org/10.1104/pp.105.066464
Gravino M, Savatin DV, Macone A, De Lorenzo G (2015) Ethylene production in Botrytis cinerea- and oligogalacturonide-induced immunity requires calcium-dependent protein kinases. Plant J Cell Mol Biol 84:1073–1086. https://doi.org/10.1111/tpj.13057
Guo H, Li L, Ye H, Yu X, Algreen A, Yin Y (2009a) Three related receptor-like kinases are required for optimal cell elongation in Arabidopsis thaliana. Proc Natl Acad Sci USA 106:7648–7653. https://doi.org/10.1073/pnas.0812346106
Guo H, Ye H, Li L, Yin Y (2009b) A family of receptor-like kinases are regulated by BES1 and involved in plant growth in Arabidopsis thaliana. Plant Signal Behav 4:784–786. https://doi.org/10.4161/psb.4.8.9231
Guo H, Nolan TM, Song G, Liu S, Xie Z, Chen J, Schnable PS, Walley JW, Yin Y (2018) FERONIA receptor kinase contributes to plant immunity by suppressing jasmonic acid signaling in Arabidopsis thaliana. Curr Biol 28:3316-3324.e6. https://doi.org/10.1016/j.cub.2018.07.078
Ha CM, Rao X, Saxena G, Dixon RA (2021) Growth-defense trade-offs and yield loss in plants with engineered cell walls. New Phytol 231:60–74. https://doi.org/10.1111/nph.17383
Hahn MG, Darvill AG, Albersheim P (1981) Host–pathogen interactions : XIX. The endogenous elicitor, a fragment of a plant cell wall polysaccharide that elicits phytoalexin accumulation in soybeans. Plant Physiol 68:1161–1169. https://doi.org/10.1104/pp.68.5.1161
Hamann T (2012) Plant cell wall integrity maintenance as an essential component of biotic stress response mechanisms. Front Plant Sci. https://doi.org/10.3389/fpls.2012.00077
Hamann T, Bennett M, Mansfield J, Somerville C (2009) Identification of cell-wall stress as a hexose-dependent and osmosensitive regulator of plant responses. Plant J Cell Mol Biol 57:1015–1026. https://doi.org/10.1111/j.1365-313X.2008.03744.x
Hander T, Fernández-Fernández ÁD, Kumpf RP, Willems P, Schatowitz H, Rombaut D, Staes A, Nolf J, Pottie R, Yao P, Gonçalves A, Pavie B, Boller T, Gevaert K, Van Breusegem F, Bartels S, Stael S (2019) Damage on plants activates Ca2+-dependent metacaspases for release of immunomodulatory peptides. Science 363:7486. https://doi.org/10.1126/science.aar7486
Haruta M, Sabat G, Stecker K, Minkoff BB, Sussman MR (2014) A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 343:408–411. https://doi.org/10.1126/science.1244454
He Z, Zou T, Xiao Q, Yuan G, Liu M, Tao Y, Zhou D, Zhang X, Deng Q, Wang S, Zheng A, Zhu J, Liang Y, Yu X, Wang A, Liu H, Wang L, Li P, Li S (2021) An L-type lectin receptor-like kinase promotes starch accumulation during rice pollen maturation. Dev Camb Engl 148:196378. https://doi.org/10.1242/dev.196378
Heckmann S, Lermontova I, Berckmans B, De Veylder L, Bäumlein H, Schubert I (2011) The E2F transcription factor family regulates CENH3 expression in Arabidopsis thaliana: regulation of AtCENH3 expression. Plant J 68:646–656. https://doi.org/10.1111/j.1365-313X.2011.04715.x
Hématy K, Sado P-E, Van Tuinen A, Rochange S, Desnos T, Balzergue S, Pelletier S, Renou J-P, Höfte H (2007) A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr Biol 17:922–931. https://doi.org/10.1016/j.cub.2007.05.018
Henty-Ridilla JL, Shimono M, Li J, Chang JH, Day B, Staiger CJ (2013) The plant actin cytoskeleton responds to signals from microbe-associated molecular patterns. PLoS Pathog 9:e1003290. https://doi.org/10.1371/journal.ppat.1003290
Hirano K, Aya K, Morinaka Y, Nagamatsu S, Sato Y, Antonio BA, Namiki N, Nagamura Y, Matsuoka M (2013) Survey of genes involved in rice secondary cell wall formation through a co-expression network. Plant Cell Physiol 54:1803–1821. https://doi.org/10.1093/pcp/pct121
Höfte H (2015) The Yin and Yang of cell wall integrity control: brassinosteroid and FERONIA signaling. Plant Cell Physiol 56:224–231. https://doi.org/10.1093/pcp/pcu182
Honkanen S, Jones VAS, Morieri G, Champion C, Hetherington AJ, Kelly S, Proust H, Saint-Marcoux D, Prescott H, Dolan L (2016) The mechanism forming the cell surface of tip-growing rooting cells is conserved among land plants. Curr Biol 26:3238–3244. https://doi.org/10.1016/j.cub.2016.09.062
Hou S, Liu Z, Shen H, Wu D (2019) Damage-associated molecular pattern-triggered immunity in plants. Front Plant Sci. https://doi.org/10.3389/fpls.2019.00646
Hu Z, Han Z, Song N, Chai L, Yao Y, Peng H, Ni Z, Sun Q (2013) Epigenetic modification contributes to the expression divergence of three TaEXPA1 homoeologs in hexaploid wheat (Triticum aestivum). New Phytol 197:1344–1352. https://doi.org/10.1111/nph.12131
Hu Z, Song N, Zheng M, Liu X, Liu Z, Xing J, Ma J, Guo W, Yao Y, Peng H, Xin M, Zhou D-X, Ni Z, Sun Q (2015) Histone acetyltransferase GCN5 is essential for heat stress-responsive gene activation and thermotolerance in Arabidopsis. Plant J 84:1178–1191. https://doi.org/10.1111/tpj.13076
Hu K, Cao J, Zhang J, Xia F, Ke Y, Zhang H, Xie W, Liu H, Cui Y, Cao Y, Sun X, Xiao J, Li X, Zhang Q, Wang S (2017) Improvement of multiple agronomic traits by a disease resistance gene via cell wall reinforcement. Nat Plants 3:17009. https://doi.org/10.1038/nplants.2017.9
Huang C, Yan Y, Zhao H, Ye Y, Cao Y (2020) Arabidopsis CPK5 phosphorylates the chitin receptor LYK5 to regulate plant innate immunity. Front Plant Sci 11:702. https://doi.org/10.3389/fpls.2020.00702
Huffaker A (2015) Plant elicitor peptides in induced defense against insects. Curr Opin Insect Sci 9:44–50. https://doi.org/10.1016/j.cois.2015.06.003
Huffaker A, Dafoe NJ, Schmelz EA (2011) ZmPep1, an ortholog of Arabidopsis elicitor peptide 1, regulates maize innate immunity and enhances disease resistance. Plant Physiol 155:1325–1338. https://doi.org/10.1104/pp.110.166710
Hurni S, Scheuermann D, Krattinger SG, Kessel B, Wicker T, Herren G, Fitze MN, Breen J, Presterl T, Ouzunova M, Keller B (2015) The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc Natl Acad Sci USA 112:8780–8785. https://doi.org/10.1073/pnas.1502522112
Hussey SG, Saïdi MN, Hefer CA, Myburg AA, Grima-Pettenati J (2015) Structural, evolutionary and functional analysis of the nac domain protein family in Eucalyptus. New Phytol 206:1337–1350. https://doi.org/10.1111/nph.13139
Intergovernmental Panel on Climate Change, 2019. IPCC special report on climate.
Jiang X (2019) Rapid functional divergence after small-scale gene duplication in grasses. BMC Evol Biol 19(1):97. https://doi.org/10.1186/s12862-019-1415-2
Jing Y, Shen N, Zheng X, Fu A, Zhao F, Lan W, Luan S (2020) Danger-associated peptide regulates root immune responses and root growth by affecting ROS formation in Arabidopsis. Int J Mol Sci 21:E4590. https://doi.org/10.3390/ijms21134590
Johnson JM, Thürich J, Petutschnig EK, Altschmied L, Meichsner D, Sherameti I, Dindas J, Mrozinska A, Paetz C, Scholz SS, Furch ACU, Lipka V, Hedrich R, Schneider B, Svatoš A, Oelmüller R (2018) A Poly(A) ribonuclease controls the cellotriose-based interaction between piriformospora indica and its host Arabidopsis. Plant Physiol 176:2496–2514. https://doi.org/10.1104/pp.17.01423
Kawasaki T, Koita H, Nakatsubo T, Hasegawa K, Wakabayashi K, Takahashi H, Umemura K, Umezawa T, Shimamoto K (2006) Cinnamoyl-CoA reductase, a key enzyme in lignin biosynthesis, is an effector of small GTPase Rac in defense signaling in rice. Proc Natl Acad Sci USA 103:230–235. https://doi.org/10.1073/pnas.0509875103
Kawasaki T, Yamada K, Yoshimura S, Yamaguchi K (2017) Chitin receptor-mediated activation of MAP kinases and ROS production in rice and Arabidopsis. Plant Signal Behav 12:e1361076. https://doi.org/10.1080/15592324.2017.1361076
Kessler SA, Shimosato-Asano H, Keinath NF, Wuest SE, Ingram G, Panstruga R, Grossniklaus U (2010) Conserved molecular components for pollen tube reception and fungal invasion. Science 330:968–971. https://doi.org/10.1126/science.1195211
Kohorn BD (2016) Cell wall-associated kinases and pectin perception. J Exp Bot 67:489–494. https://doi.org/10.1093/jxb/erv467
Kohorn BD, Kobayashi M, Johansen S, Friedman HP, Fischer A, Byers N (2006) Wall-associated kinase 1 (WAK1) is crosslinked in endomembranes, and transport to the cell surface requires correct cell-wall synthesis. J Cell Sci 119:2282–2290. https://doi.org/10.1242/jcs.02968
Kohorn BD, Johansen S, Shishido A, Todorova T, Martinez R, Defeo E, Obregon P (2009) Pectin activation of MAP kinase and gene expression is WAK2 dependent: wall associated kinase-dependent pectin activation. Plant J 60:974–982. https://doi.org/10.1111/j.1365-313X.2009.04016.x
Kohorn BD, Kohorn SL, Todorova T, Baptiste G, Stansky K, McCullough M (2012) A dominant allele of Arabidopsis pectin-binding wall-associated kinase induces a stress response suppressed by MPK6 but Not MPK3 mutations. Mol Plant 5:841–851. https://doi.org/10.1093/mp/ssr096
Kohorn BD, Hoon D, Minkoff BB, Sussman MR, Kohorn SL (2016) Rapid oligo-galacturonide induced changes in protein phosphorylation in Arabidopsis. Mol Cell Proteomics MCP 15:1351–1359. https://doi.org/10.1074/mcp.M115.055368
Kosugi S, Ohashi Y (2002) Interaction of the Arabidopsis E2F and DP proteins confers their concomitant nuclear translocation and transactivation. Plant Physiol 128:833–843. https://doi.org/10.1104/pp.010642
Kozlova LV, Nazipova AR, Gorshkov OV, Petrova AA, Gorshkova TA (2020) Elongating maize root: zone-specific combinations of polysaccharides from type I and type II primary cell walls. Sci Rep 10:10956. https://doi.org/10.1038/s41598-020-67782-0
Krattinger SG, Keller B (2017) Resistance: double gain with one gene. Nat Plants 3:17019. https://doi.org/10.1038/nplants.2017.19
Kumar M, Campbell L, Turner S (2016) Secondary cell walls: biosynthesis and manipulation. J Exp Bot 67:515–531. https://doi.org/10.1093/jxb/erv533
Kumar S, Beena AS, Awana M, Singh A (2017) Physiological, biochemical, epigenetic and molecular analyses of wheat (Triticum aestivum) genotypes with contrasting salt tolerance. Front Plant Sci. https://doi.org/10.3389/fpls.2017.01151
Kutschera A, Dawid C, Gisch N, Schmid C, Raasch L, Gerster T, Schäffer M, Smakowska-Luzan E, Belkhadir Y, Vlot AC, Chandler CE, Schellenberger R, Schwudke D, Ernst RK, Dorey S, Hückelhoven R, Hofmann T, Ranf S (2019) Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants. Science 364:178–181. https://doi.org/10.1126/science.aau1279
Labbé J, Muchero W, Czarnecki O, Wang J, Wang X, Bryan AC, Zheng K, Yang Y, Xie M, Zhang J, Wang D, Meidl P, Wang H, Morrell-Falvey JL, Cope KR, Maia LGS, Ané J-M, Mewalal R, Jawdy SS, Gunter LE, Schackwitz W, Martin J, Le Tacon F, Li T, Zhang Z, Ranjan P, Lindquist E, Yang X, Jacobson DA, Tschaplinski TJ, Barry K, Schmutz J, Chen J-G, Tuskan GA (2019) Mediation of plant–mycorrhizal interaction by a lectin receptor-like kinase. Nat Plants 5:676–680. https://doi.org/10.1038/s41477-019-0469-x
Lafos M, Kroll P, Hohenstatt ML, Thorpe FL, Clarenz O, Schubert D (2011) Dynamic regulation of H3K27 trimethylation during Arabidopsis differentiation. PLOS Genet 7:e1002040. https://doi.org/10.1371/journal.pgen.1002040
Lamers J, van der Meer T, Testerink C (2020) How plants sense and respond to stressful environments. Plant Physiol 182:1624–1635. https://doi.org/10.1104/pp.19.01464
Le Gall H, Philippe F, Domon J-M, Gillet F, Pelloux J, Rayon C (2015) Cell wall metabolism in response to abiotic stress. Plants 4:112–166. https://doi.org/10.3390/plants4010112
Lee HG, Seo PJ (2021) Transcriptional activation of SUGAR TRANSPORT PROTEIN 13 mediates biotic and abiotic stress signaling. Plant Signal Behav 16:1920759. https://doi.org/10.1080/15592324.2021.1920759
Li J, Staiger CJ (2018) Understanding cytoskeletal dynamics during the plant immune response. Annu Rev Phytopathol 56:513–533. https://doi.org/10.1146/annurev-phyto-080516-035632
Li C-H, Wang G, Zhao J-L, Zhang L-Q, Ai L-F, Han Y-F, Sun D-Y, Zhang S-W, Sun Y (2014a) The receptor-like kinase SIT1 Mediates salt sensitivity by activating MAPK3/6 and regulating ethylene homeostasis in rice. Plant Cell 26:2538–2553. https://doi.org/10.1105/tpc.114.125187
Li H, Yan S, Zhao L, Tan J, Zhang Q, Gao F, Wang P, Hou H, Li L (2014b) Histone acetylation associated up-regulation of the cell wall related genes is involved in salt stress induced maize root swelling. BMC Plant Biol 14:105. https://doi.org/10.1186/1471-2229-14-105
Li B, Meng X, Shan L, He P (2016a) Transcriptional regulation of pattern-triggered immunity in plants. Cell Host Microbe 19:641–650. https://doi.org/10.1016/j.chom.2016.04.011
Li C, Wang L, Cui Y, He L, Qi Y, Zhang J, Lin J, Liao H, Lin Q, Yang T, Yu F, Liu X (2016b) Two FERONIA-like receptor (FLR) genes are required to maintain architecture, fertility, and seed yield in rice. Mol Breed 36:151. https://doi.org/10.1007/s11032-016-0580-x
Li C, Wu H-M, Cheung AY (2016c) FERONIA and her Pals: functions and mechanisms. Plant Physiol 171:2379–2392. https://doi.org/10.1104/pp.16.00667
Li P, Lu Y-J, Chen H, Day B (2020) The lifecycle of the plant immune system. Crit Rev Plant Sci 39:72–100. https://doi.org/10.1080/07352689.2020.1757829
Liang Z, Riaz A, Chachar S, Ding Y, Du H, Gu X (2020) Epigenetic modifications of mRNA and DNA in plants. Mol Plant 13:14–30. https://doi.org/10.1016/j.molp.2019.12.007
Liao H, Tang R, Zhang X, Luan S, Yu F (2017) FERONIA receptor kinase at the crossroads of hormone signaling and stress responses. Plant Cell Physiol 58:1143–1150. https://doi.org/10.1093/pcp/pcx048
Lindner H, Müller LM, Boisson-Dernier A, Grossniklaus U (2012) CrRLK1L receptor-like kinases: not just another brick in the wall. Curr Opin Plant Biol 15:659–669. https://doi.org/10.1016/j.pbi.2012.07.003
Locci F, Benedetti M, Pontiggia D, Citterico M, Caprari C, Mattei B, Cervone F, De Lorenzo G (2019) An Arabidopsis berberine bridge enzyme-like protein specifically oxidizes cellulose oligomers and plays a role in immunity. Plant J Cell Mol Biol 98:540–554. https://doi.org/10.1111/tpj.14237
Lori M, van Verk MC, Hander T, Schatowitz H, Klauser D, Flury P, Gehring CA, Boller T, Bartels S (2015) Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: interfamily incompatibility of perception but compatibility of downstream signalling. J Exp Bot 66:5315–5325. https://doi.org/10.1093/jxb/erv236
Lorrai R, Ferrari S (2021) Host cell wall damage during pathogen infection: mechanisms of perception and role in plant–pathogen interactions. Plants Basel Switz 10:399. https://doi.org/10.3390/plants10020399
Luo X, Xu N, Huang J, Gao F, Zou H, Boudsocq M, Coaker G, Liu J (2017) A lectin receptor-like kinase mediates pattern-triggered salicylic acid signaling. Plant Physiol 174:2501–2514. https://doi.org/10.1104/pp.17.00404
Luo X, Wu W, Liang Y, Xu N, Wang Z, Zou H, Liu J (2020) Tyrosine phosphorylation of the lectin receptor-like kinase LORE regulates plant immunity. EMBO J 39:e102856. https://doi.org/10.15252/embj.2019102856
Macho AP, Zipfel C (2014) Plant PRRs and the activation of innate immune signaling. Mol Cell 54:263–272. https://doi.org/10.1016/j.molcel.2014.03.028
Malinovsky FG, Fangel JU, Willats WGT (2014) The role of the cell wall in plant immunity. Front Plant Sci. https://doi.org/10.3389/fpls.2014.00178
Marti L, Savatin D-V, Gigli-Bisceglia N, de Turris V, Cervone F, De Lorenzo G (2021) The intracellular ROS accumulation in elicitor-induced immunity requires the multiple organelle-targeted Arabidopsis NPK1-related protein kinases. Plant Cell Environ 44:931–947. https://doi.org/10.1111/pce.13978
Mecchia MA, Rövekamp M, Giraldo-Fonseca A, Meier D, Gadient P, Bowman JL, Grossniklaus U (2020) Characterization of the single FERONIA homolog in Marchantia polymorpha reveals an ancestral role of Cr RLK1L receptor kinases in regulating cell expansion and morphological integrity (preprint). Evol Biol. https://doi.org/10.1101/2020.12.23.424085
Mélida H, Sopeña-Torres S, Bacete L, Garrido-Arandia M, Jordá L, López G, Muñoz-Barrios A, Pacios LF, Molina A (2018) Non-branched β-1,3-glucan oligosaccharides trigger immune responses in Arabidopsis. Plant J Cell Mol Biol 93:34–49. https://doi.org/10.1111/tpj.13755
Mentlak TA, Kombrink A, Shinya T, Ryder LS, Otomo I, Saitoh H, Terauchi R, Nishizawa Y, Shibuya N, Thomma BPHJ, Talbot NJ (2012) Effector-mediated suppression of chitin-triggered immunity by magnaporthe oryzae is necessary for rice blast disease. Plant Cell 24:322–335. https://doi.org/10.1105/tpc.111.092957
Merz D, Richter J, Gonneau M, Sanchez-Rodriguez C, Eder T, Sormani R, Martin M, Hématy K, Höfte H, Hauser M-T (2017) T-DNA alleles of the receptor kinase THESEUS1 with opposing effects on cell wall integrity signaling. J Exp Bot 68:4583–4593. https://doi.org/10.1093/jxb/erx263
Miedes E, Vanholme R, Boerjan W, Molina A (2014) The role of the secondary cell wall in plant resistance to pathogens. Front Plant Sci. https://doi.org/10.3389/fpls.2014.00358
Miya A, Albert P, Shinya T, Desaki Y, Ichimura K, Shirasu K, Narusaka Y, Kawakami N, Kaku H, Shibuya N (2007) CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proc Natl Acad Sci USA 104:19613–19618. https://doi.org/10.1073/pnas.0705147104
Miyashima S, Koi S, Hashimoto T, Nakajima K (2011) Non-cell-autonomous microRNA165 acts in a dose-dependent manner to regulate multiple differentiation status in the Arabidopsis root. Development 138:2303–2313. https://doi.org/10.1242/dev.060491
Miyazaki S, Murata T, Sakurai-Ozato N, Kubo M, Demura T, Fukuda H, Hasebe M (2009) ANXUR1 and 2, sister genes to FERONIA/SIRENE, are male factors for coordinated fertilization. Curr Biol 19:1327–1331. https://doi.org/10.1016/j.cub.2009.06.064
Molina A, Miedes E, Bacete L, Rodríguez T, Mélida H, Denancé N, Sánchez-Vallet A, Rivière M-P, López G, Freydier A, Barlet X, Pattathil S, Hahn M, Goffner D (2021) Arabidopsis cell wall composition determines disease resistance specificity and fitness. Proc Natl Acad Sci 118:e2010243118. https://doi.org/10.1073/pnas.2010243118
Moscatiello R, Mariani P, Sanders D, Maathuis FJM (2006) Transcriptional analysis of calcium-dependent and calcium-independent signalling pathways induced by oligogalacturonides. J Exp Bot 57:2847–2865. https://doi.org/10.1093/jxb/erl043
Mu Q, Li X, Luo J, Pan Q, Li Y, Gu T (2021) Characterization of expansin genes and their transcriptional regulation by histone modifications in strawberry. Planta 254:21. https://doi.org/10.1007/s00425-021-03665-6
Mujahid H, Tan F, Zhang J, Nallamilli BRR, Pendarvis K, Peng Z (2013) Nuclear proteome response to cell wall removal in rice (Oryza sativa). Proteome Sci 11:26. https://doi.org/10.1186/1477-5956-11-26
Nakagawa Y, Katagiri T, Shinozaki K, Qi Z, Tatsumi H, Furuichi T, Kishigami A, Sokabe M, Kojima I, Sato S, Kato T, Tabata S, Iida K, Terashima A, Nakano M, Ikeda M, Yamanaka T, Iida H (2007) Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots. Proc Natl Acad Sci USA 104:3639–3644. https://doi.org/10.1073/pnas.0607703104
Nakashima A, Chen L, Thao NP, Fujiwara M, Wong HL, Kuwano M, Umemura K, Shirasu K, Kawasaki T, Shimamoto K (2008) RACK1 functions in rice innate immunity by interacting with the Rac1 immune complex. Plant Cell 20:2265–2279. https://doi.org/10.1105/tpc.107.054395
Newman M-A, Sundelin T, Nielsen JT, Erbs G (2013) MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front Plant Sci. https://doi.org/10.3389/fpls.2013.00139
Ning Y, Liu W, Wang G-L (2017) Balancing immunity and yield in crop plants. Trends Plant Sci 22:1069–1079. https://doi.org/10.1016/j.tplants.2017.09.010
Nothnagel EA, McNeil M, Albersheim P, Dell A (1983) Host–pathogen interactions : XXII. A galacturonic acid oligosaccharide from plant cell walls elicits phytoalexins. Plant Physiol 71:916–926. https://doi.org/10.1104/pp.71.4.916
Novaković L, Guo T, Bacic A, Sampathkumar A, Johnson KL (2018) Hitting the wall—sensing and signaling pathways involved in plant cell wall remodeling in response to abiotic stress. Plants. https://doi.org/10.3390/plants7040089
Ogawa M, Kay P, Wilson S, Swain SM (2009) ARABIDOPSIS DEHISCENCE ZONE POLYGALACTURONASE1 (ADPG1), ADPG2, and QUARTET2 are Polygalacturonases required for cell separation during reproductive development in Arabidopsis. Plant Cell 21:216–233. https://doi.org/10.1105/tpc.108.063768
Ortiz-Morea FA, Liu J, Shan L, He P (2021) Malectin-like receptor kinases as protector deities in plant immunity. Nat Plants. https://doi.org/10.1038/s41477-021-01028-3
Petutschnig EK, Jones AME, Serazetdinova L, Lipka U, Lipka V (2010) The lysin motif receptor-like kinase (LysM-RLK) CERK1 is a major chitin-binding protein in Arabidopsis thaliana and subject to chitin-induced phosphorylation. J Biol Chem 285:28902–28911. https://doi.org/10.1074/jbc.M110.116657
Pikaard CS, Mittelsten Scheid O (2014) Epigenetic regulation in plants. Cold Spring Harb Perspect Biol 6:a019315. https://doi.org/10.1101/cshperspect.a019315
Prevedel R, Diz-Muñoz A, Ruocco G, Antonacci G (2019) Brillouin microscopy: an emerging tool for mechanobiology. Nat Methods 16:969–977. https://doi.org/10.1038/s41592-019-0543-3
Qi H, Zhu X, Guo F, Lv L, Zhang Z (2021) The wall-associated receptor-like kinase TaWAK7D is required for defense responses to rhizoctonia cerealis in wheat. Int J Mol Sci 22:5629. https://doi.org/10.3390/ijms22115629
Qiao Z, Yates TB, Shrestha HK, Engle NL, Flanagan A, Morrell-Falvey JL, Sun Y, Tschaplinski TJ, Abraham PE, Labbé J, Wang Z-Y, Hettich RL, Tuskan GA, Muchero W, Chen J-G (2021) Towards engineering ectomycorrhization into switchgrass bioenergy crops via a lectin receptor-like kinase. Plant Biotechnol J. https://doi.org/10.1111/pbi.13671
Qu S, Zhang X, Song Y, Lin J, Shan X (2017) THESEUS1 positively modulates plant defense responses against Botrytis cinerea through GUANINE EXCHANGE FACTOR4 signaling: THE1 functions in plant defense responses. J Integr Plant Biol 59:797–804. https://doi.org/10.1111/jipb.12565
Ranf S, Gisch N, Schäffer M, Illig T, Westphal L, Knirel YA, Sánchez-Carballo PM, Zähringer U, Hückelhoven R, Lee J, Scheel D (2015) A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat Immunol 16:426–433. https://doi.org/10.1038/ni.3124
Rao X, Dixon RA (2017) Brassinosteroid mediated cell wall remodeling in grasses under abiotic stress. Front Plant Sci 8:806. https://doi.org/10.3389/fpls.2017.00806
Rao S, Zhou Z, Miao P, Bi G, Hu M, Wu Y, Feng F, Zhang X, Zhou J-M (2018) Roles of receptor-like cytoplasmic kinase VII members in pattern-triggered immune signaling. Plant Physiol 177:1679–1690. https://doi.org/10.1104/pp.18.00486
Rasmussen MW, Roux M, Petersen M, Mundy J (2012) MAP kinase cascades in Arabidopsis innate immunity. Front Plant Sci 3:169. https://doi.org/10.3389/fpls.2012.00169
Ridley BL, O’Neill MA, Mohnen D (2001) Pectins: structure, biosynthesis, and oligogalacturonide-related signaling. Phytochemistry 57:929–967. https://doi.org/10.1016/s0031-9422(01)00113-3
Rui Y, Dinneny JR (2020) A wall with integrity: surveillance and maintenance of the plant cell wall under stress. New Phytol 225:1428–1439. https://doi.org/10.1111/nph.16166
Rydahl MG, Hansen AR, Kračun SK, Mravec J (2018) Report on the current inventory of the toolbox for plant cell wall analysis: proteinaceous and small molecular probes. Front Plant Sci 9:581. https://doi.org/10.3389/fpls.2018.00581
Saelim L, Akiyoshi N, Tan TT, Ihara A, Yamaguchi M, Hirano K, Matsuoka M, Demura T, Ohtani M (2019) Arabidopsis Group IIId ERF proteins positively regulate primary cell wall-type CESA genes. J Plant Res 132:117–129. https://doi.org/10.1007/s10265-018-1074-1
Saintenac C, Lee W-S, Cambon F, Rudd JJ, King RC, Marande W, Powers SJ, Bergès H, Phillips AL, Uauy C, Hammond-Kosack KE, Langin T, Kanyuka K (2018) Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat Genet 50:368–374. https://doi.org/10.1038/s41588-018-0051-x
Sakamoto S, Somssich M, Nakata MT, Unda F, Atsuzawa K, Kaneko Y, Wang T, Bågman A-M, Gaudinier A, Yoshida K, Brady SM, Mansfield SD, Persson S, Mitsuda N (2018) Complete substitution of a secondary cell wall with a primary cell wall in Arabidopsis. Nat Plants 4:777–783. https://doi.org/10.1038/s41477-018-0260-4
Scheible WR, Eshed R, Richmond T, Delmer D, Somerville C (2001) Modifications of cellulose synthase confer resistance to isoxaben and thiazolidinone herbicides in Arabidopsis Ixr1 mutants. Proc Natl Acad Sci USA 98:10079–10084. https://doi.org/10.1073/pnas.191361598
Schuster SC, Khan S (1994) The bacterial flagellar motor. Annu Rev Biophys Biomol Struct 23:509–539. https://doi.org/10.1146/annurev.bb.23.060194.002453
Shang-Guan K, Wang M, Htwe NMPS, Li P, Li Y, Qi F, Zhang D, Cao M, Kim C, Weng H, Cen H, Black IM, Azadi P, Carlson RW, Stacey G, Liang Y (2018) Lipopolysaccharides trigger two successive bursts of reactive oxygen species at distinct cellular locations. Plant Physiol 176:2543–2556. https://doi.org/10.1104/pp.17.01637
Shen N, Jing Y, Tu G, Fu A, Lan W (2020) Danger-associated peptide regulates root growth by promoting protons extrusion in an AHA2-dependent manner in Arabidopsis. Int J Mol Sci 21:E7963. https://doi.org/10.3390/ijms21217963
Shih H-W, Miller ND, Dai C, Spalding EP, Monshausen GB (2014) The receptor-like kinase FERONIA is required for mechanical signal transduction in Arabidopsis seedlings. Curr Biol 24:1887–1892. https://doi.org/10.1016/j.cub.2014.06.064
Shimizu T, Nakano T, Takamizawa D, Desaki Y, Ishii-Minami N, Nishizawa Y, Minami E, Okada K, Yamane H, Kaku H, Shibuya N (2010) Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J 64:204–214. https://doi.org/10.1111/j.1365-313X.2010.04324.x
Shinya T, Yasuda S, Hyodo K, Tani R, Hojo Y, Fujiwara Y, Hiruma K, Ishizaki T, Fujita Y, Saijo Y, Galis I (2018) Integration of danger peptide signals with herbivore-associated molecular pattern signaling amplifies anti-herbivore defense responses in rice. Plant J Cell Mol Biol 94:626–637. https://doi.org/10.1111/tpj.13883
Solis-Miranda J, Fonseca-García C, Nava N, Pacheco R, Quinto C (2020) Genome-wide identification of the CrRLK1L subfamily and comparative analysis of its role in the legume-rhizobia symbiosis. Genes 11:793. https://doi.org/10.3390/genes11070793
Somerville C (2004) Toward a systems approach to understanding plant cell walls. Science 306:2206–2211. https://doi.org/10.1126/science.1102765
Souza CA, Li S, Lin AZ, Boutrot F, Grossmann G, Zipfel C, Somerville SC (2017) Cellulose-derived oligomers act as damage-associated molecular patterns and trigger defense-like responses1. Plant Physiol 173:2383–2398. https://doi.org/10.1104/pp.16.01680
Stegmann M, Monaghan J, Smakowska-Luzan E, Rovenich H, Lehner A, Holton N, Belkhadir Y, Zipfel C (2017) The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 355:287–289. https://doi.org/10.1126/science.aal2541
Sun J, Li L, Wang P, Zhang S, Wu J (2017) Genome-wide characterization, evolution, and expression analysis of the leucine-rich repeat receptor-like protein kinase (LRR-RLK) gene family in Rosaceae genomes. BMC Genomics 18:763. https://doi.org/10.1186/s12864-017-4155-y
Sun Y, Qiao Z, Muchero W, Chen J-G (2020) Lectin receptor-like kinases: the sensor and mediator at the plant cell surface. Front Plant Sci 11:596301. https://doi.org/10.3389/fpls.2020.596301
Sun Y, Jiang C, Jiang R, Wang F, Zhang Z, Zeng J (2021) A novel NAC transcription factor from eucalyptus, EgNAC141, positively regulates lignin biosynthesis and increases lignin deposition. Front Plant Sci 12:642090. https://doi.org/10.3389/fpls.2021.642090
Takai R, Isogai A, Takayama S, Che F-S (2008) Analysis of flagellin perception mediated by flg22 receptor OsFLS2 in rice. Mol Plant–microbe Interact MPMI 21:1635–1642. https://doi.org/10.1094/MPMI-21-12-1635
Tan F, Zhang K, Mujahid H, Verma DPS, Peng Z (2011) Differential histone modification and protein expression associated with cell wall removal and regeneration in rice (Oryza sativa). J Proteome Res 10:551–563. https://doi.org/10.1021/pr100748e
Tang D, Wang G, Zhou J-M (2017) Receptor kinases in plant-pathogen interactions: more than pattern recognition. Plant Cell 29:618–637. https://doi.org/10.1105/tpc.16.00891
Tateno M, Brabham C, DeBolt S (2016) Cellulose biosynthesis inhibitors—a multifunctional toolbox. J Exp Bot 67:533–542. https://doi.org/10.1093/jxb/erv489
Taylor-Teeples M, Lin L, de Lucas M, Turco G, Toal TW, Gaudinier A, Young NF, Trabucco GM, Veling MT, Lamothe R, Handakumbura PP, Xiong G, Wang C, Corwin J, Tsoukalas A, Zhang L, Ware D, Pauly M, Kliebenstein DJ, Dehesh K, Tagkopoulos I, Breton G, Pruneda-Paz JL, Ahnert SE, Kay SA, Hazen SP, Brady SM (2015) An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature 517:571–575. https://doi.org/10.1038/nature14099
Tripathi RK, Aguirre JA, Singh J (2021) Genome-wide analysis of wall associated kinase (WAK) gene family in barley. Genomics 113:523–530. https://doi.org/10.1016/j.ygeno.2020.09.045
Tsang DL, Edmond C, Harrington JL, Nühse TS (2011) Cell wall integrity controls root elongation via a general 1-aminocyclopropane-1-carboxylic acid-dependent, ethylene-independent pathway. Plant Physiol 156:596–604. https://doi.org/10.1104/pp.111.175372
Ursache R, Fujita S, Tendon VD, Geldner N (2021) Combined fluorescent seed selection and multiplex CRISPR/Cas9 assembly for fast generation of multiple Arabidopsis mutants. Plant Methods 17(1):111
Vaahtera L, Schulz J, Hamann T (2019) Cell wall integrity maintenance during plant development and interaction with the environment. Nat Plants 5:924–932. https://doi.org/10.1038/s41477-019-0502-0
Vaaje-Kolstad G, Westereng B, Horn SJ, Liu Z, Zhai H, Sørlie M, Eijsink VGH (2010) An oxidative enzyme boosting the enzymatic conversion of recalcitrant polysaccharides. Science 330:219–222. https://doi.org/10.1126/science.1192231
Vaid N, Pandey PK, Tuteja N (2012) Genome-wide analysis of lectin receptor-like kinase family from Arabidopsis and rice. Plant Mol Biol 80:365–388. https://doi.org/10.1007/s11103-012-9952-8
Vaid N, Macovei A, Tuteja N (2013) Knights in action: lectin receptor-like kinases in plant development and stress responses. Mol Plant 6:1405–1418. https://doi.org/10.1093/mp/sst033
Verica JA, He Z-H (2002) The cell wall-associated kinase (WAK) and WAK-like kinase gene family. Plant Physiol 129:455–459. https://doi.org/10.1104/pp.011028
Vilarrasa-Blasi J, Vellosillo T, Jinkerson RE, Fauser F, Xiang T, Minkoff BB, Wang L, Kniazev K, Guzman M, Osaki J, Sussman MR, Jonikas MC, Dinneny JR (2021) Identification of green lineage osmotic stress pathways. bioRxiv. https://doi.org/10.1101/2021.07.19.453009
Voxeur A, Höfte H (2020) Pectin-derived immune elicitors in response to lignin modification in plants. Proc Natl Acad Sci USA 117:4442–4444. https://doi.org/10.1073/pnas.2000509117
Voxeur A, Habrylo O, Guénin S, Miart F, Soulié M-C, Rihouey C, Pau-Roblot C, Domon J-M, Gutierrez L, Pelloux J, Mouille G, Fagard M, Höfte H, Vernhettes S (2019) Oligogalacturonide production upon Arabidopsis thaliana–Botrytis cinerea interaction. Proc Natl Acad Sci USA 116:19743–19752. https://doi.org/10.1073/pnas.1900317116
Wang T, McFarlane HE, Persson S (2016) The impact of abiotic factors on cellulose synthesis. J Exp Bot 67(2):543–552
Wang L, Wilkins KA, Davies JM (2018a) Arabidopsis DORN1 extracellular ATP receptor; activation of plasma membrane K+ -and Ca2+ -permeable conductances. New Phytol 218:1301–1304. https://doi.org/10.1111/nph.15111
Wang P, Moore BM, Panchy NL, Meng F, Lehti-Shiu MD (2018b) Factors influencing gene family size variation among related species in a plant family, Solanaceae. Genome Biol Evol 18:1–7
Wang S, Yang H, Mei J, Liu X, Wen Z, Zhang L, Xu Z, Zhang B, Zhou Y (2019) Rice homeobox protein KNAT7 integrates the pathways regulating cell expansion and wall stiffness. Plant Physiol 181:669–682. https://doi.org/10.1104/pp.19.00639
Wang X, Wang D, Xu W, Kong L, Ye X, Zhuang Q, Fan D, Luo K (2021) Histone methyltransferase ATX1 dynamically regulates fiber secondary cell wall biosynthesis in Arabidopsis inflorescence stem. Nucleic Acids Res 49:190–205. https://doi.org/10.1093/nar/gkaa1191
Widhalm JR, Jaini R, Morgan JA, Dudareva N (2015) Rethinking how volatiles are released from plant cells. Trends Plant Sci 20:545–550. https://doi.org/10.1016/j.tplants.2015.06.009
Wolf S, Hématy K, Höfte H (2012) Growth control and cell wall signaling in plants. Annu Rev Plant Biol 63:381–407
Wolny E, Skalska A, Braszewska A, Mur LAJ, Hasterok R (2021) Defining the cell wall, cell cycle and chromatin landmarks in the responses of brachypodium distachyon to salinity. Int J Mol Sci. https://doi.org/10.3390/ijms22020949
Wong HL, Pinontoan R, Hayashi K, Tabata R, Yaeno T, Hasegawa K, Kojima C, Yoshioka H, Iba K, Kawasaki T, Shimamoto K (2007) Regulation of rice NADPH oxidase by binding of Rac GTPase to its N-terminal extension. Plant Cell 19:4022–4034. https://doi.org/10.1105/tpc.107.055624
Wong A, Hu N, Tian X, Yang Y, Gehring C (2021) Nitric oxide sensing revisited. Trends Plant Sci 26:885–897. https://doi.org/10.1016/j.tplants.2021.03.009
Wood AKM, Walker C, Lee W-S, Urban M, Hammond-Kosack KE (2020) Functional evaluation of a homologue of plant rapid alkalinisation factor (RALF) peptides in Fusarium graminearum. Fungal Biol 124:753–765. https://doi.org/10.1016/j.funbio.2020.05.001
Wu X, Bacic A, Johnson KL, Humphries J (2020) The role of brachypodium distachyon wall-associated kinases (WAKs) in cell expansion and stress responses. Cells 9:2478. https://doi.org/10.3390/cells9112478
Yamada K, Yamaguchi K, Shirakawa T, Nakagami H, Mine A, Ishikawa K, Fujiwara M, Narusaka M, Narusaka Y, Ichimura K, Kobayashi Y, Matsui H, Nomura Y, Nomoto M, Tada Y, Fukao Y, Fukamizo T, Tsuda K, Shirasu K, Shibuya N, Kawasaki T (2016) The Arabidopsis CERK1-associated kinase PBL27 connects chitin perception to MAPK activation. EMBO J 35:2468–2483. https://doi.org/10.15252/embj.201694248
Yamaguchi K, Yamada K, Ishikawa K, Yoshimura S, Hayashi N, Uchihashi K, Ishihama N, Kishi-Kaboshi M, Takahashi A, Tsuge S, Ochiai H, Tada Y, Shimamoto K, Yoshioka H, Kawasaki T (2013) A receptor-like cytoplasmic kinase targeted by a plant pathogen effector is directly phosphorylated by the chitin receptor and mediates rice immunity. Cell Host Microbe 13:347–357. https://doi.org/10.1016/j.chom.2013.02.007
Yamaguchi K, Yoshimura Y, Nakagawa S, Mezaki H, Yoshimura S, Kawasaki T (2019) OsDRE2 contributes to chitin-triggered response through its interaction with OsRLCK185. Biosci Biotechnol Biochem 83:281–290. https://doi.org/10.1080/09168451.2018.1543012
Yang P, Praz C, Li B, Singla J, Robert CAM, Kessel B, Scheuermann D, Lüthi L, Ouzunova M, Erb M, Krattinger SG, Keller B (2019a) Fungal resistance mediated by maize wall-associated kinase ZmWAK-RLK1 correlates with reduced benzoxazinoid content. New Phytol 221:976–987. https://doi.org/10.1111/nph.15419
Yang Y, Yoo CG, Rottmann W, Winkeler KA, Collins CM, Gunter LE, Jawdy SS, Yang X, Pu Y, Ragauskas AJ, Tuskan GA, Chen J-G (2019b) PdWND3A, a wood-associated NAC domain-containing protein, affects lignin biosynthesis and composition in Populus. BMC Plant Biol 19:486. https://doi.org/10.1186/s12870-019-2111-5
Yang Z, Xing J, Wang L, Liu Y, Qu J, Tan Y, Fu X, Lin Q, Deng H, Yu F (2020) Mutations of two FERONIA-like receptor genes enhance rice blast resistance without growth penalty. J Exp Bot 71:2112–2126. https://doi.org/10.1093/jxb/erz541
Yang C, Liu R, Pang J, Ren B, Zhou H, Wang G, Wang E, Liu J (2021) Poaceae-specific cell wall-derived oligosaccharides activate plant immunity via OsCERK1 during Magnaporthe oryzae infection in rice. Nat Commun. https://doi.org/10.1038/s41467-021-22456-x
Yokoyama R, Nishitani K (2004) Genomic basis for cell-wall diversity in plants. a comparative approach to gene families in rice and Arabidopsis. Plant Cell Physiol 45:1111–1121. https://doi.org/10.1093/pcp/pch151
Yoo CG, Meng X, Pu Y, Ragauskas AJ (2020) The critical role of lignin in lignocellulosic biomass conversion and recent pretreatment strategies: a comprehensive review. Bioresour Technol 301:122784. https://doi.org/10.1016/j.biortech.2020.122784
Yoshimura K, Iida K, Iida H (2021) MCAs in Arabidopsis are Ca2+-permeable mechanosensitive channels inherently sensitive to membrane tension. Nat Commun 12:6074. https://doi.org/10.1038/s41467-021-26363-z
Yu Y (2019) OsKNAT7 bridges secondary cell wall formation and cell growth regulation. Plant Physiol 181:385–386. https://doi.org/10.1104/pp.19.01018
Yu F, Qian L, Nibau C, Duan Q, Kita D, Levasseur K, Li X, Lu C, Li H, Hou C, Li L, Buchanan BB, Chen L, Cheung AY, Li D, Luan S (2012) FERONIA receptor kinase pathway suppresses abscisic acid signaling in Arabidopsis by activating ABI2 phosphatase. Proc Natl Acad Sci 109:14693–14698. https://doi.org/10.1073/pnas.1212547109
Yuan N, Yuan S, Li Z, Zhou M, Wu P, Hu Q, Mendu V, Wang L, Luo H (2018) Stress induced factor 2, a leucine-rich repeat kinase regulates basal plant pathogen defense. Plant Physiol 176:3062–3080. https://doi.org/10.1104/pp.17.01266
Zarattini M, Corso M, Kadowaki MA, Monclaro A, Magri S, Milanese I, Jolivet S, de Godoy MO, Hermans C, Fagard M, Cannella D (2021) LPMO-oxidized cellulose oligosaccharides evoke immunity in Arabidopsis conferring resistance towards necrotrophic fungus B. cinerea. Commun Biol 4:727. https://doi.org/10.1038/s42003-021-02226-7
Zhang J, Li W, Xiang T, Liu Z, Laluk K, Ding X, Zou Y, Gao M, Zhang X, Chen S, Mengiste T, Zhang Y, Zhou J-M (2010) Receptor-like cytoplasmic kinases integrate signaling from multiple plant immune receptors and are targeted by a Pseudomonas syringae effector. Cell Host Microbe 7:290–301. https://doi.org/10.1016/j.chom.2010.03.007
Zhang N, Zhang B, Zuo W, Xing Y, Konlasuk S, Tan G, Zhang Q, Ye J, Xu M (2017) Cytological and molecular characterization of ZmWAK-mediated head-smut resistance in maize. Mol Plant-Microbe Interact MPMI 30:455–465. https://doi.org/10.1094/MPMI-11-16-0238-R
Zhang J, Zhang H, Srivastava AK, Pan Y, Bai J, Fang J, Shi H, Zhu J-K (2018) Knockdown of rice MicroRNA166 confers drought resistance by causing leaf rolling and altering stem xylem development. Plant Physiol 176:2082–2094. https://doi.org/10.1104/pp.17.01432
Zhang Y, Malzahn AA, Sretenovic S, Qi Y (2019) The emerging and uncultivated potential of CRISPR technology in plant science. Nat Plants 5:778–794. https://doi.org/10.1038/s41477-019-0461-5
Zhang X, Peng H, Zhu S, Xing J, Li X, Zhu Z, Zheng J, Wang L, Wang B, Chen J, Ming Z, Yao K, Jian J, Luan S, Coleman-Derr D, Liao H, Peng Y, Peng D, Yu F (2020a) Nematode-encoded RALF peptide mimics facilitate parasitism of plants through the FERONIA receptor kinase. Mol Plant 13:1434–1454. https://doi.org/10.1016/j.molp.2020.08.014
Zhang X, Yang Z, Wu D, Yu F (2020b) RALF–FERONIA signaling: linking plant immune response with cell growth. Plant Commun 1:100084. https://doi.org/10.1016/j.xplc.2020b.100084
Zhang H, Li H, Zhang X, Yan W, Deng P, Zhang Y, Peng S, Wang Y, Wang C, Ji W (2021) Wall-associated receptor kinase and the expression profiles in wheat responding to fungal stress (preprint). Plant Biol. https://doi.org/10.1101/2021.07.11.451968
Zhao C, Jiang W, Zayed O, Liu X, Tang K, Nie W, Li Y, Xie S, Li Y, Long T, Liu L, Zhu Y, Zhao Y, Zhu J-K (2021) The LRXs-RALFs-FER module controls plant growth and salt stress responses by modulating multiple plant hormones. Natl Sci Rev 8:149. https://doi.org/10.1093/nsr/nwaa149
Zheng M, Liu X, Lin J, Liu X, Wang Z, Xin M, Yao Y, Peng H, Zhou D-X, Ni Z, Sun Q, Hu Z (2019) Histone acetyltransferase GCN5 contributes to cell wall integrity and salt stress tolerance by altering the expression of cellulose synthesis genes. Plant J 97:587–602. https://doi.org/10.1111/tpj.14144
Zhong S, Li L, Wang Z, Ge Z, Li Q, Bleckmann A, Wang J, Song Z, Shi Y, Liu T, Li L, Zhou H, Wang Y, Zhang L, Wu H-M, Lai L, Gu H, Dong J, Cheung AY, Dresselhaus T, Qu L-J (2022) RALF peptide signaling controls the polytubey block in Arabidopsis. Science 375:290–296. https://doi.org/10.1126/science.abl4683
Zhou J-M, Zhang Y (2020) Plant immunity: danger perception and signaling. Cell 181:978–989. https://doi.org/10.1016/j.cell.2020.04.028
Zhou S, Jiang W, Long F, Cheng S, Yang W, Zhao Y, Zhou D-X (2017) Rice homeodomain protein WOX11 recruits a histone acetyltransferase complex to establish programs of cell proliferation of crown root meristem. Plant Cell 29:1088–1104. https://doi.org/10.1105/tpc.16.00908
Zhou Q, Liu J, Wang J, Chen S, Chen L, Wang J, Wang H-B, Liu B (2020) The juxtamembrane domains of Arabidopsis CERK1, BAK1, and FLS2 play a conserved role in chitin-induced signaling. J Integr Plant Biol 62:556–562. https://doi.org/10.1111/jipb.12847
Zhu H, Li C, Gao C (2020) Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat Rev Mol Cell Biol 21:661–677. https://doi.org/10.1038/s41580-020-00288-9
Funding
Open access funding provided by NTNU Norwegian University of Science and Technology (incl St. Olavs Hospital - Trondheim University Hospital). The work was supported by The Research Council of Norway (NFR, Grant No. 315325).
Author information
Authors and Affiliations
Contributions
All authors have contributed to writing of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Baez, L.A., Tichá, T. & Hamann, T. Cell wall integrity regulation across plant species. Plant Mol Biol 109, 483–504 (2022). https://doi.org/10.1007/s11103-022-01284-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-022-01284-7