
Abstract
Across the process of domestication, human selection produces changes in target attributes as well as changes that are not necessarily desired by humans due to pleiotropic or linked genes. In this paper we addressed, correlated changes between genetic diversity, damage level, defense mechanisms (resistance and tolerance), and fitness due to the domestication process of Stenocereus pruinosus (Otto ex Pfeiff.) Buxbaum and Stenocereus stellatus (Pfeiff.) Riccobono, an endemic columnar cactus of south-central Mexico. One hundred eighty individuals of S. stellatus from wild, in situ managed, and cultivated populations of Valle de Tehuacán and Mixteca Baja, Puebla, were sampled, and attributes including damage level, defense mechanisms and fitness (number of fruits) were measured. The DNA of 176 individuals was extracted to amplify and analyze five microsatellites in order to estimate genetic diversity and structure. As expected, cultivated populations showed a significantly higher damage level, as well as lower resistance and genetic diversity. Depending on the form of management, correlations between genetic diversity and the rest of the attributes exhibited different patterns. In wild populations, genetic diversity was positively correlated with damage and negatively with resistance; in situ managed populations exhibited the opposite pattern, and in cultivated populations, no correlations were found between these attributes. We propose a hypothetic model of human selection to explain the variation in these correlations. No differences in genetic diversity and tolerance were detected between regions; however, the populations of Valle de Tehuacán exhibited more damage and more resistance. In both regions, populations showed a positive correlation between fitness and resistance and a negative correlation between damage and resistance, suggesting the existence of a defense mechanism to ensure fitness. Also, non-regional differentiation suggests an eventual gene flow due to pollinators, human movement of branches, or a common ancestry before the domestication process.
Similar content being viewed by others


Introduction
Human management consists of a series of deliberate actions undertaken to transform ecological systems into socioecological systems, following the requirements and interests of the societies to which they belong (Lindig and Casas 2013; Casas et al. 2014, 2015). Management also means manipulating populations of target species to ensure their availability by selecting for attributes desired by humans (Casas et al. 1997, 2007; Blancas et al. 2013, 2014). This process, called domestication, generates morphological, physiological, and genetic divergences between populations of managed organisms and their wild relatives, known as domestication syndromes (Schwanitz 1966; Harlan 1975; Casas et al. 1999, 2015; Gepts 2003). Changes in allelic and genotypic frequencies between wild and managed populations are commonly detected, such that wild relatives of domesticated species commonly possess high levels of genetic diversity accumulated over hundreds of thousands to millions of years of natural evolution (Doebley et al. 2006; Casas et al. 2007; Pickersgill 2007). Conversely, domesticated populations exhibit lower levels of genetic diversity as a result of selection by humans over hundreds or thousands of years of domestication (Maxted et al. 2013).
This reduction in genetic diversity in domesticated populations makes them vulnerable to herbivore attack and pathogen infestation because of the loss of the chemical and structural mechanisms that protect them (Pickersgill 2007; Casas and Parra 2016; Bravo-Avilez 2017). It has been argued that this effect is correlated with increases in productivity, such as the increased size of flowers, fruits, or seeds (Benrey et al. 1998), at the expense of a reduction in the distribution of energy to other functions, such as the synthesis of secondary metabolites or defense structures (Schaller 2008). This means that plants reallocate resources to those structures or functions selected for by humans. Consequently, domesticated plants frequently exhibit more susceptibility to herbivores, pathogens, and competitors than their wild counterparts (Li et al. 2017).
Plants have two basic defense strategies against herbivores and pathogens: resistance and tolerance (Strauss and Agrawal 1999). Resistance reduces the amount of damage sustained from herbivores or pathogens because of the presence of chemical and physical defenses (Ehrlich and Raven 1964; Berenbaum et al. 1986). Since resistance implies energy allocation for the synthesis of secondary metabolites (alkaloids, tannins) or specific structures (spines, trichomes, thick cuticle), plants are faced with the dilemma of concentrating valuable resources on either growth or resistance (Bazzaz et al. 1987; Herms and Mattson 1992; Rendón and Núñez-Farfán 1998; Huot et al. 2014). Tolerance does not prevent herbivores but rather allows plants to compensate for the damage caused by natural enemies and maintain fitness in the presence of damage (Roy and Kirchner 2000). Which mechanisms are associated with the reduction or loss of these defense strategies during the process of domestication? Because the domestication process generally starts as bottleneck events or founder effects on the genetic diversity existing in wild populations, the loss of genetic diversity could be due to the loss of allelic variants that provide proteins or polysaccharides involved in these attributes (e.g., production of secondary metabolites, cuticle), causing a greater susceptibility to damage (Harlan 1975; Pickersgill 2007; Maxted et al. 2013; Bravo-Avilez et al. 2014; Casas and Parra 2016; Bravo-Avilez 2017) or a reduction in resistance (Mosquera et al. 2008). However, this loss could also be related to the existence of pleiotropic genes or gametic linkage between closely linked genetic components (Murren 2002), such as resistance and yield genes, as has been demonstrated in wheat (Hayes et al. 1996, 2003) and potato (Mosquera et al. 2008).
However, evolution under domestication of both defense strategies and their correlation with other attributes (e.g., genetic diversity or damage level) from a population approach is scarcely addressed in the literature. There are many studies on correlated responses in cultivated plants of economic importance (Hayes et al. 2003 and references inside), and most of them are analyzed from an agronomic approach. Chaudhary (2013) analyzed the possible evolution of both mechanisms between wild relatives and domesticated plants, but the taxa analyzed are evolutionarily distant, and gene flow is almost nonexistent. Huot et al. (2014) analyzed the concept of a growth-defense trade-off in plants and incorporated evidence of molecular and hormone crosstalk in this trade-off to enrich strategies for plant breeding. More recently, the existence of recessive resistance loci involved in matched-allele host–pathogen interactions was reported for wild pepper Capsicum annuum var. glabriusculum (Dunal) Heiser & Pickersgill Heiserpopulations under an incipient domestication process, demonstrating that human management has profound effects on the diversity and evolution of resistance genes (Poulicard et al. 2016). This study represents a population analysis of the evolution of resistance under domestication, but correlations with fitness components were not directly analyzed. Moreira et al. (2018) analyzed changes in plant defense compounds (constitutive and induced) involved in targeted and nontargeted attributes in three varieties of Brassica oleracea L. (one wild and two domesticated). They concluded that constitutive defenses decreased in domesticated varieties, but differences between constitutive and induced defenses did not, suggesting that human selection did not modify this trade-off. Additionally, no differences were found in the magnitude of defenses between targeted and nontargeted attributes. Thus, a reduction in defenses in domesticated cabbage is presumably the result of direct selection on these compounds rather than indirect effects via trade-offs between size and defense.
For cacti, damage caused by physical and biological factors has been reported in various species (Bravo-Avilez et al. 2019a). Recently, the effect of damage on defense mechanisms and fitness components was documented in two species of columnar cacti, Stenocereus pruinosus and Stenocereus stellatus (Pfeiffer) Riccobono, subject to different forms of management (Bravo-Avilez et al. 2014, 2019b; Bravo-Avilez 2017). However, the genetic diversity of this species has been documented before, and there is no integrated approach including both kinds of attributes (e.g., morphological and genetic). We aimed to analyze the correlated responses between genetic diversity levels and susceptibility to damage, defense mechanisms, and fitness components in wild, in situ managed, and cultivated populations of S. stellatus growing in sympatry (Casas and Caballero 1998; Casas et al. 2006; Cruse-Sanders et al. 2013). We expected variance in the levels of genetic diversity among S. stellatus populations with different forms of management (wild > in situ managed > cultivated). In the case of wild populations, high levels of genetic diversity will be positively correlated with higher resistance and negatively correlated with damage, tolerance, and fitness values. The opposite will be found in cultivated populations, where the decrease in genetic diversity will be negatively correlated with resistance and positively correlated with damage, tolerance, and fitness because human selection has eroded genetic diversity by selecting individuals with a higher number of fruits. Between regions, we expect genetic differentiation due to evidence of the use of this species in the Valle de Tehuacán region in ancient times, since at least 8000 years ago (Smith 1967). Thus, populations from the Valle de Tehuacán region will present less genetic diversity (which will be positively correlated with resistance and negatively correlated with damage levels) than those of the Mixteca Baja region.
Materials and methods
Study species
Stenocereus stellatus (Pfeiffer) Riccobono (“xoconochtli” or “pitaya de Agosto”) is a 2–6-m tall columnar cactus, branching at the base, with cylindrical, erect branches; 8–12 ribs rounded. Radial spines 7–9 (sometimes 13), 1–3 central spines. Nocturnal flowers, tubular to narrowly campanulate, white or pale rose, with reddish sepals, approximately 4–6 cm long and 3–4 cm in diameter; spiny and globular fruits; pulp exhibits different colors: red, orange, yellow, and white. It is an endemic cactus to central-southern Mexico (Casas 2001) with a distribution centered in the states of Guerrero, Morelos, Oaxaca, and Puebla (Casas and Caballero 1996, 1998; Guzmán et al. 2003). This species has been of use since ancient times, at least 8000 years (Smith 1967; Caballero et al. 1998), and continues to be domesticated in the Valle de Tehuacán and Mixteca Baja regions. It is common to find populations subject to different forms of management (wild, in situ managed, and cultivated) growing sympatrically across almost the entirety of the species’ geographic distribution (Casas et al. 1997, 1999; Casas 2002), which indicates an ancient and close interaction with human communities. Their edible fruits are in high demand, and the stems are used as forage or for living fences.
Damage, defense mechanisms, and fitness components
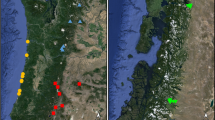
At the beginning of 2012, we sampled 180 individuals from six populations: wild (W), in situ managed (ISM), and cultivated (C) populations from two regions, Valle de Tehuacán (TV) and Mixteca Baja (MB) (Table 1; Fig. 1; supplementary file). From these individuals, 1043 branches were randomly selected to estimate a qualitative damage level based on the percentage of each branch with damage and visually classify damage types (Bravo-Avilez et al. 2014; Bravo-Avilez 2017). From these records, a subsample of 459 branches was chosen (between two and four branches from each of the 180 individuals) to analyze defense mechanisms (e.g., resistance and tolerance).

Location of wild, in situ managed and cultivated populations of S. stellatus. municipalities of Santiago Miahuatlan and Ajalpan, corresponding to the Valle de Tehuacán (TV), state of Puebla (gray color). municipality of San Pedro and San Pablo Tequixtepec, corresponding to the Mixteca Baja region (MB), state of Oaxaca (yellow color) (color figure online)
Resistance (RE) was calculated from the following attributes: number of ribs per branch (r), number of areolas (a), and length of the central spine (lcs). The latter two attributes were observed in a 150 cm2 area of the branch (30 cm × 5 cm) approximately 1.20 m from the ground. The formula is as follows:
The formula indicates that the greater the RE value is, the higher the resistance level.
The branching rate (TR) was estimated as a component of tolerance, measured as the number of new shoots produced by the branches sampled (459) in a time interval. We counted the number of shoots smaller than 30 cm produced in 2012 (t1) and again in 2013 (t2); with this, TR was calculated with the following formula:
Fitness was estimated according to the number of fruits produced by 180 individuals from mid-July to the end of September 2012.
A factorial analysis of variance was used to evaluate significant differences in damage levels, defense mechanisms, and fitness between forms of management and between regions, as well as a form of management × region interaction, using JMP® 9.0.1 software (SAS Institute Inc. 2010).
All the data from Bravo-Avilez (2017) were used to make correlations with genetic diversity.
Genetic diversity in S. stellatus populations between forms of management and regions
During the flowering season in 2012, two flower buds were collected from a subset of 176 of the 180 sampled individuals (because four individuals died). Flower buds were wrapped and labeled in aluminum foil and stored frozen at 80 °C at the Divisional Laboratory of Molecular Biology of the Universidad Autónoma Metropolitana Iztapalapa.
DNA was extracted from each individual using QIAGEN’s commercial DNeasy® Plant Mini Kit extraction protocol, with minor modifications, and DNA quality was verified using a Nanodrop 2000 spectrophotometer. DNA integrity was checked by 1% agarose gel electrophoresis with 0.2 µl of GelRedTM stain and a molecular weight marker with 100 bp bands; the gel was visualized with ultraviolet light using the MiniLumi DNR Bio-Imaging Systems biodocumentation system.
The amplification of the microsatellites or simple sequence repeats (SSR; Table 2) was performed at a volume of 12.5 µl, containing 6.25 µl of PCR Master Mix, PROMEGA, 0.75 µl of the forward primer marked with fluorescence at 10 µM, 0.75 µl of the reverse primer at 10 µM, 0.75 µl of nuclease-free water, and 4 µl of DNA.
New DNA chain synthesis was carried out using the molecular polymerase chain reaction (PCR) technique. Amplification conditions for loci (JCS; Cruse-Sander et al. 2013) started with denaturation at 94 °C for four minutes, followed by 25 successive cycles of denaturation at 94 °C for one minute, annealing according to the melting temperature for each locus (Table 2) for 60 s, and extension at 72 °C for two minutes, with a final extension at 72 °C for four minutes.
Analysis of genetic diversity between forms of management and regions
Genetic characterization of the alleles was performed using GeneMarker® software (Applied BioSystems) in a multiplex reaction and calibrated with the LIZ 500 ladder. The presence and frequency of null alleles for each locus per population were verified using the MICROCHECKER 2.2.3 program (Van Oosterhout et al. 2004). The deviation from Hardy–Weinberg equilibrium and the linkage disequilibrium for each locus per population were estimated with GENEPOP v. 1.2 (Raymond and Rousset 1995) and ARLEQUIN 3.0 (Excoffier et al. 2005), respectively.
Genetic diversity across forms of management and region was evaluated with the GENALEX 6.5 program (Peakall and Smouse 2005); to elucidate possible significant differences in the levels of genetic diversity (observed and expected heterozygosity), a Kruskal–Wallis test with a significance value of 0.05 was performed using Minitab® statistical software (Minitab 2007).
To infer the genetic structure, a Bayesian allocation algorithm was used in STRUCTURE 2.3.3 software (Pritchard et al. 2000). In addition, an ad hoc statistical test of ∆K (Evanno et al. 2005) was carried out in STRUCTURE HARVESTER software (Earl and vonHoldt 2012).
Factorial correspondence analysis (FCA) was performed using the GENETIX 4.05 program (Belkhir et al. 2004). The evaluation at hierarchical levels (difference between forms of management and between regions) was carried out by means of an analysis of molecular variance (AMOVA) with the ARLEQUIN 3.0 program (Excoffier et al. 2005). To analyze the genetic differentiation between populations and between genetic groups, Nei’s standard genetic distances (Ds) were calculated with the POPULATIONS 1.2.32 program (Langella 1999), and a phenogram was prepared with the TFPGA 1.3 program, which uses the unweighted pair group method with arithmetic means (UPGMA) algorithm (Sneath and Sokal 1973). The number of migrants (Nm) was evaluated based on the FST and RST values using the ARLEQUIN 3.0 program (Excoffier et al. 2005).
Spearman correlations between genetic diversity, damage levels, defense mechanisms, and fitness
Based on the genotype of each individual (five loci of 176 plants), the number of heterozygous loci in each individual was counted as a parameter of genetic diversity (see Table 1 in the supplementary material). Spearman correlation matrices (ρ) were developed using JMP® software (Sall et al. 2017), considering damage levels, defense mechanisms, and fitness by form of management and by region. For defense mechanisms, the RE and TR values were considered; for fitness, the total number of fruits produced per individual was used (Bravo-Avilez 2017). The correlation significance was tested at a probability level of 5% (Gomez and Gomez 1984).
Results
Damage levels, defense mechanisms, and fitness between forms of management and regions
Factorial analyses of variance of total damage (considering biological and physical factors; Table 3) indicated that damage levels were significantly higher in cultivated populations; between regions, populations from Valle de Tehuacán had a significantly higher incidence of damage compared to populations from the Mixteca Baja region. The form of management × region interaction indicated that the three populations from the Valle de Tehuacán and the cultivated population from the Mixteca Baja region presented the highest damage levels, while the wild population from the Mixteca Baja region exhibited the lowest damage level.
Regarding resistance, it was found that (1) the in situ managed populations were significantly more resistant than the others, (2) the cultivated populations presented the lowest resistance and (3) populations from the Mixteca Baja region presented greater resistance than those from the Valle de Tehuacán.
The branching rate (TR) did not exhibit significant differences between forms of management or between regions. Nevertheless, the form of management × region interaction exhibited significant differences: the wild population from the Valle de Tehuacán region and the in situ managed population from the Mixteca Baja region presented the highest TR, while the wild population from the Mixteca Baja region and the in situ managed population from the Valle de Tehuacán presented the lowest TR (Table 3).
Fitness also exhibited significant differences in the form of management × region interaction: individuals in the cultivated population from the Valle de Tehuacán produced the greatest number of fruits, while those in the population from the Mixteca Baja region, as well as the wild and in situ managed populations, produced the lowest number (Table 3).
Genetic diversity and genetic structure of S. stellatus
The Hardy–Weinberg equilibrium test per locus per population showed that Pchi 9 and JCS 68 were in equilibrium in the cultivated population of the Mixteca Baja region, while the rest of the loci presented a significant deviation from Hardy–Weinberg equilibrium (p ≤ 0.05). Two combinations with linkage disequilibrium were observed in the in situ managed population from the Mixteca Baja region, with seven in the wild population from the Valle de Tehuacán region, and three in the cultivated population from the Valle de Tehuacán region.
An average value of heterozygosity was obtained: HO = 0.422 ± 0.036. This value varied across forms of management: for the wild population, HO = 0.430 ± 0.146; for the in situ managed population, HO = 0.434 ± 0.094; and for the cultivated population, HO = 0.403 ± 0.024. Comparisons between forms of management and between regions with the Kruskal–Wallis test revealed that there were no significant differences (H = 0.09, p = 0.95 and H = 0.10, p = 0.756, respectively). With respect to expected heterozygosity (HT = 0.664 ± 0.025), significant differences were found between forms of management (H = 7.44, p = 0.024); these differences were found between in situ managed populations, with the highest value, and cultivated populations, with the lowest value (medians of HT = 0.758 and 0.617, respectively; H = 7.01, p = 0.008). Wild populations showed an intermediate value, HT = 0.695. No significant differences between regions were found for HT. The average FIS was 0.381, indicating breeding between populations.
Bayesian analysis showed that individuals of S. stellatus coming from the six populations studied were grouped into four genetic groups (K = 4; Fig. 2). This result was confirmed by the ad hoc ∆K statistical test, the factorial correspondence analysis and the phenogram based on genetic distances (Fig. 3): group I constitutes individuals in wild (WMB) and cultivated (CMB) populations from the Mixteca Baja region; group II, individuals in the in situ managed population from the Mixteca Baja region (ISMB); group III, individuals in wild (WTV) and in situ managed populations (ISTV) from the Valle de Tehuacán region; and group IV, individuals in the cultivated population from the Valle de Tehuacán region (CTV; Fig. 2). These results indicated a regional grouping of populations. However, some migrants and/or a common ancestry related to incomplete lineage classification (ILS) was found within each group (Gow et al. 2006).

Genetic structure of six populations of S. stellatus: W, wild; IS, in situ managed; C, cultivated; MB, Mixteca Baja region; VT, Valle de Tehuacán

Phenogram constructed from UPGMA analysis to estimate Nei genetic distances between six populations of S. stellatus: W, wild; IS, in situ managed; C, cultivated; MB, Mixteca Baja region (brown line); VT, Valle de Tehuacán (gray line) (color figure online)
Molecular variance analysis showed that just over 61% (p ≤ 0.001) of the variation was distributed within populations, followed by variation between populations at 24.7% (p ≤ 0.001); the smallest variation was between forms of management, at 13.59% (p ≤ 0.001). The inbreeding coefficient within populations was RIS = 0.286, and the total inbreeding coefficient was RIT = 0.383. The interpretation of these two coefficients is similar to Wright’s FIS and FIT, respectively.
Spearman correlations between genetic diversity, damage level, defense mechanisms, and fitness
Between forms of management The Spearman correlation coefficient (ρ) showed significant and variable relationships between genetic diversity and damage (Fig. 4); the coefficient was positive in wild populations (ρ = 0.46***) and negative in the in situ managed population (ρ = 0.32*). Cultivated populations did not exhibit significant differences (ρ = 0.087 ns). Wild populations showed a significant and negative correlation between genetic diversity and resistance (ρ = 0.340**) and between damage and resistance (ρ = 0.383***) and a positive correlation between damage and branching rate (ρ = 0.275*). In situ managed populations showed a positive correlation between genetic diversity and resistance (ρ = 0.402**), between genetic diversity and fitness (ρ = 0.296*), and between fitness and resistance (ρ = 0.476***). Damage was negatively correlated with resistance and branching rate (ρ = 0.414*** and ρ = 0.408***, respectively). Cultivated populations did not exhibit significant correlations (Table 4).

Correlation between levels of genetic diversity and levels of damage in a wild (r = 0.3594, p = 0.006, n = 57) and b in situ managed populations (r = 0.3077, p = 0.0167, n = 60) (b). The expected heterozygosity was considered as the parameter of genetic diversity
Between regions Populations from the Valle de Tehuacán and Mixteca Baja regions (Fig. 5) presented a positive correlation between fitness and resistance (ρ = 0.230* and ρ = 0.226*, respectively) and a negative correlation between damage and resistance (ρ = 0.254* and ρ = 0.411***).

Correlation between resistance-damage (p = 0.254, p = 0.018) (a), fitness and resistance (p = 0.230, p = 0.033) (b) in populations of Valle de Tehuacán (n = 86) and resistance-damage (p = 0.411, p = 0.001) (c), fitness and resistance (p = 0.226, p = 0.032) (d) in populations of the Mixteca Baja regions (n = 90)
Discussion
Our results showed that cultivated populations of S. stellatus exhibited a significant decrease in levels of genetic diversity and resistance, as well as an increase in damage levels, which means that these nontargeted attributes have also changed during human selection for fruit production, which is the target attribute (Casas et al. 2006; Cruse-Sanders et al. 2013). Thus, domestication syndromes (e.g., decreased genetic diversity and increased susceptibility to damage) were corroborated. Studies on the genetic diversity in other columnar cacti under domestication processes using microsatellites (Polaskia chichipe [Rol. Gross.] Backeb, Otero-Arnaiz et al. (2005); S. pruinosus [Otto ex Pfeiff.] Buxb. Parra et al. 2010) reported that cultivated populations generally have lower levels of genetic diversity than wild populations. The negative effect of domestication on genetic diversity in species of great agronomic interest has been documented in domesticated soybean (Glycine max L.), the genetic diversity of which was reduced by 50% compared to its wild relative (Hyten et al. 2006). It has also been documented that domesticated rice (Oryza sativa L.) contains only between 10 and 20% of the genetic diversity present in its wild ancestor, Oryza rufipogon Griff. (Kovach and McCouch 2008). It has been stated that domestication generally results in a reduction in the genetic diversity of managed populations because groups of domesticated individuals commonly involve only a fraction of the diversity existing in wild populations (Harlan 1975; Hawkes 1983; Doebley 1992; Pickersgill 2007). However, other genetic processes could be involved, such as gametic linkage or pleiotropy (Hayes et al. 1996, 2003; Murren 2002; Mosquera et al. 2008). It would be interesting to analyze which is the best mechanism to explain these changes in genetic diversity. Our results did not coincide with previously reported data for the same species (Casas et al. 2006; Cruse-Sanders et al. 2013; Table 5), which indicated that domesticated populations presented greater genetic diversity than wild populations. These differences correspond to the distribution of the sampled populations and their possible interrelation with pollinators and humans, as mentioned by these authors, or to unique attributes of the sampled individuals because those highly damaged individuals were probably not sampled in previous studies.
Defense mechanisms have not been previously analyzed in columnar cacti subjected to domestication. In the case of resistance, although there are no previous studies on domesticated columnar cacti, recent studies with Brassica oleracea and Capsicum annuum have found a decrease in resistance, as well as an increase in damage levels in domesticated populations (Poulicard et al. 2016; Moreira et al. 2018), as was found in the present study. It would be interesting to determine which mechanisms are associated with these changes to understand how humans, based on an empirical process of selection, have reduced resistance levels in domesticated populations (Moreira et al. 2018).
Tolerance did not exhibit significant differences due to human selection. The tolerance response may have evolved prior to the domestication process. Due to the architecture of this species, under natural conditions and subject to different stress factors, branches are commonly replaced; through the domestication process, this response has not been modified, and pruning (the process by which farmers eliminate damaged branches) also plays a role. Additionally, this attribute is not linked to other genes (pleiotropic or gametic) (Murren 2002). It seems that both defense mechanisms have evolved independently (Restif and Koella 2004).
Fitness represents the target attribute, and it would be expected that mean values would increase under human selection. We analyzed individuals with different damage levels from the worst individuals, those who were almost dead, to the best individuals. Consequently, fitness values represented not only human selection but also the environmental conditions of each form of management.
Between regions, our findings were the opposite of what was expected. Genetic diversity was similar between the two regions; however, populations from Valle de Tehuacán exhibited higher damage and resistance values. Contrary to our results, Casas et al. (2006) and Cruse-Sanders et al. (2013) reported greater genetic diversity in populations from the Mixteca Baja region compared with those from the Valle de Tehuacán region. These differences could be related to the number of markers that were used in each of these studies (Pritchard et al. 2000) and to the kind of individuals sampled. In our study, we sampled damaged or very damaged individuals who may not have been considered in another context. However, a phylogeographic study needs to be carried out to understand the historical dispersion of the species and to elucidate its origin and diversification, considering the following hypotheses. First, if the oldest remains of S. stellatus were found in the Valle de Tehuacán (MacNeish 1967; Smith 1967), then this region could correspond to its center of origin. Second, authors such as Casas et al. (2006) and Cruse-Sanders et al. (2013) reported that populations from the Mixteca Baja region present greater genetic diversity than those from the Valle de Tehuacán region. If this pattern is consistent, it is possible that the species was first domesticated in the Mixteca Baja region and then disseminated to the Valle de Tehuacán region. Third, the species has been subjected to independent domestication processes for thousands of years in different places within its distribution area (multiple domestication processes; Piperno 2011; Langlie et al. 2014), which includes populations from Guerrero, Morelos, Oaxaca, and Puebla; therefore, it is necessary to study the dynamics of this species throughout this area.
Genetic structure analysis showed that the genetic groups (K = 4) were geographically associated, which supports the hypothesis of an independent domestication process. The number of migrants (Nm) according to the FST and RST values suggests that the gene flow is regional, between wild and domesticated populations, favored by the role of bats in pollination and bats and birds in seed dispersal, and largely by the human dispersion of vegetative materials (Casas et al. 2006; Cruse-Sanders et al. 2013). These results were supported by both genetic structure analysis and genetic distance analysis, which showed that the most genetically similar populations were wild and cultivated populations from the Mixteca Baja region and wild and in situ managed populations from the Valle de Tehuacán region. Nevertheless, a common ancestral origin of all the populations from a single large population is supported by STRUCTURE analysis, which showed fragmentation by different factors, with human activities (mainly the movement of branches between different regions) being the most important. At this time (8000 years ago), there were already different human groups living in different areas of the Valle de Tehuacán and Mixteca Baja regions (Casas et al. 2006; Cruse-Sanders et al. 2013).
In the present study, we proposed an analysis of the correlated changes between target attributes and those that are not directly subject to human selection (e.g., correlated attributes). The analysis of the attribute changes during domestication has been performed under the lens of an independent evolution of each attribute and that all changes are a direct effect of human selection. A model based on phenotypic integration led us to analyze how some attributes could be subject to an indirect effect of human selection due to other genetic mechanisms (gametic linkage or pleiotropy). Some findings in the present study reinforce this approach.
In contrast to the responses expected as a result of domestication syndromes, correlations between genetic diversity and defense, damage, and fitness exhibited quite different patterns between populations with different forms of management. In wild populations, higher levels of genetic diversity were negatively correlated with resistance and positively correlated with damage; in the case of in situ managed populations, these patterns were completely the opposite. Surprisingly, in the case of cultivated populations, no significant correlation was detected between genetic diversity and resistance, tolerance, damage, and fitness.
Recent studies in natural populations have demonstrated that in wild populations, greater genetic diversity is correlated with higher damage levels (Castagneyrol et al. 2012). Possible explanations include a higher presence of generalist (rather than specialist) herbivores with mixed diets or spillover from more suitable to less suitable host plants and the presence of different plant genotypes with different functional traits, such as competitive ability (Cahill et al. 2005), forage quality for herbivorous insects (Karley et al. 2008), and resistance to herbivory (Maddox and Root 1987; Wise 2007; Agniezka et al. 2009). According to Bravo-Avilez et al. (2017,2019a,2019b,2014), eight kinds of damage were recorded in populations with different forms of management in both regions. All of these were recorded in all populations. However, wild populations of S. stellatus exhibited significantly more damage from insects and small birds, which produced small holes in the branches; ants such as Atta sp. and Crematogaster sp., which foraged the apical section of the branches, including reproductive structures; and Cactophagus spinolae (Gyllenhal) and Aleochara sp. beetle larvae of the family Hydrophilidae, which infested the roots. Cultivated populations exhibited a high incidence of damage apparently caused by Phoma spp. (Monreal-Vargas et al. 2014; Bravo-Avilez and Rendón-Aguilar in process), which produce gray spots on the branches. These results supported the findings that higher genetic diversity in wild populations was positively correlated with damage and negatively correlated with resistance. However, damage did not affect tolerance or fitness.
Based on our results, we propose a model of human selection to reconstruct different scenarios depending on the form of management (Fig. 6). Originally, this study started with the hypothesis that compared to domesticated populations, wild populations would show higher levels of genetic diversity, which is necessary for their interactions with biotic and abiotic factors (Cronin and Abrahamson 2001; Crawford et al. 2007); they share variation in defense mechanisms (resistance in principle) and, consequently, less susceptibility to damage (Rosenthal and Dirzo 1997; Gepts 2003, 2006; Chaudhary 2013; Bravo-Avilez 2017). Through human selection based on empirical management, phenotypes with higher genetic diversity that are more resistant or more tolerant and have less damage without negative effects on fitness are selected for propagation in managed in situ areas or even for cultivation. A study on genetic families of oak saplings with various levels of genetic diversity that were subjected to polyphagous herbivores in mixed and monoculture treatments showed that those families with higher genetic diversity were more damaged in mixed treatment but suffered less damage when grown in monocultures (Castagneyrol et al. 2012).

Hypothetic model of domestication process of correlated responses between populations of S. stellatus under different forms of management. Yellow line indicates the level of correlated attributes where human selection could operate (color figure online)
In in situ managed populations, farmers select for phenotypes with more resistance and less damage and did not indirectly select for a decrease in genetic diversity. Alternatively, perhaps gene flow prevented this erosion in diversity, thus fitness was not affected. The positive correlation observed between genetic diversity and fitness (ρ = 0.296*) and genetic diversity and resistance (ρ = 0.2402**) indicates that a resistance response has evolved (Fornoni and Núñez-Farfan 2000). Additionally, resistance was not correlated with branching rate. These results suggest that a resistance response has evolved in in situ managed populations of S. stellatus; thus, the most resistant plants produced the greatest number of fruits. These results agree with what was reported by Bravo-Avilez (2017), who proposed that the species opts for resistance as its main defense mechanism.
In the case of cultivated populations, no significant correlation was detected between genetic diversity and resistance or damage, which indicates that human selection eroded genetic diversity and modified ancestral phenotypic integration. It is likely that human selection would have fixed these attributes; the target attribute, which is fruit production, has been maintained until today (Fig. 6). However, statistical data must be regarded with some reservations since the Spearman correlation coefficient (ρ) was less than 0.5, which is considered a weak relationship (Martínez-Ortega et al. 2009).
The hypothesis of differences between regions was supported by an expected reduction in genetic diversity in populations from the Valle de Tehuacán region in comparison to those from the Mixteca Baja region as a consequence of historical management based on the archaeological findings of the use of S. stellatus in the Abejas Phase (5400–4300 years BP) and Palo Blanco (2150–1300 years BP) in the Valle de Tehuacán region (MacNeish 1967; Smith 1967). It was then proposed that a decrease in genetic diversity in that region would be correlated with an increase in the levels of damage to plants. However, although populations from the Valle de Tehuacán region did show significantly greater damage than those from the Mixteca Baja region (Bravo-Avilez 2017), the differences in genetic diversity were not significant. It is possible that eventual gene flow among forms of management within each region, as well as between them, could reduce the effects of regional isolation of populations on genetic diversity (Casas et al. 2007; Cruse-Sanders et al. 2013). Contrasting differences in levels of damage could indicate that different environmental conditions between the two regions favored the presence of damage in one of them (Valle de Tehuacán). Bravo-Avilez and Rendón-Aguilar (work in process) found that damage in these populations was controlled by different biological and physical factors between populations and regions. In the case of the Valle de Tehuacán region, wild and in situ managed populations were under recent pressure by human activities.
Although the size of spines and the number of ribs are good estimators of resistance in cacti, not all herbivores are deterred by these structures. Thus, it is necessary to consider, in future studies, the thickness of the cuticle, as well as the variation in secondary compounds, which have been reported in other studies (de Oliveira and da Silva Machado 2003; El-Seedi et al. 2005; da Silva et al. 2010; Falcão et al. 2012). Indeed, during this study, we wondered whether the damage was due to the reduction in the number of thorns or if damaged plants lost the thorns after suffering the damage.
Some of the herbivores were not recorded or collected. This information is necessary to clarify the ecological characteristics of herbivores, pests, and diseases (ectophagous, endophagous, and poliphagous) to explain how their interactions with S. stellatus change during the domestication process. Additionally, it is necessary to expand the study to include an analysis of abiotic components, such as climatic factors, edaphic characteristics, and floristic composition, associated with different forms of management.
Despite these limitations, this research contributed to analyzing the changes that occur during the process of domestication from phenotypic integration and population approaches. Some studies have analyzed correlations between genetic diversity and damage in domesticated species. Most were done under experimental conditions using inbred lines with specific characteristics. Others considered wild and domesticated materials, but they were analyzed under controlled conditions. In the present study, we analyzed these correlations at the population level under natural conditions, including the important role of traditional knowledge. Many studies on the domestication process, human selection, and/or domestication syndromes focus on target attributes. We believe that it is necessary to incorporate correlated attributes, such as damage and defense mechanisms, to describe the domestication process in a holistic way; even with careful sampling and data analysis, the domestication process can influence the selection of individuals and the results that we obtain.
Conclusions
The present study represents the first attempt to analyze correlations between genetic diversity, damage level, defense mechanisms (resistance and tolerance components), and fitness in populations of the columnar cacti S. stellatus under domestication processes to verify hypotheses related to domestication syndromes.
The invaluable opportunity to analyze this process in populations subject to different forms of management growing in sympatry led us to recreate a hypothetical scenario of how these changes could occur.
Genetic data confirm that genetic diversity, which is one of the correlated attributes, decreased through the domestication process, and levels of damage significantly increased, making populations more susceptible to damage (biological and physical). However, farmers selected more resistant individuals with lower levels of damage. In all cases, fitness, which represents the target attribute, is maintained. The most intensively managed population, the cultivated population, showed decreased genetic diversity and modifications to its ancestral phenotype. Nevertheless, the target attribute, fruit production, has been maintained until the present day, and even when levels of genetic diversity decrease, fruit production is maintained through different practices, such as the exchange of vegetative materials and the maintenance of natural pollination processes. It is important to emphasize that all of these selection practices have occurred under empiric knowledge based on daily experience and decisions that farmers must make.
At the regional level, populations of both regions showed the same patterns in terms of correlated responses, which suggests that human selection has moved in the same direction in both regions. Additionally, the populations did not differ in terms of levels of genetic diversity, suggesting that occasional gene flow occurs between them. Differences were found in levels of damage in the case of Valle de Tehuacán populations, suggesting that environmental factors, mainly anthropic factors, are involved in this response more than human selection per se. It is therefore necessary to consider these factors in future research because domestication processes involve not only human selection of target or correlated attributes but also the control of environmental variables.
References
Agniezka MK, Cahill JF, Keddie BA (2009) Plant genetic diversity yields increased plant productivity and herbivore performance. J Ecol 98:237–245
Bazzas FA, Chiariello NR, Coley PD, Pitelka LF (1987) Allocationg resources to reproduction and defense. Bioscience 37:58–67
Belkhir K, Borsa P, Chikhi L, Arrufaste N, Bonhomme F (2004) GENETIX 4.05, logiciel sous Windows TM pour la génétique des populations. Laboratoire Génome, Populations, Interactions, Montpellier, France
Benrey B, Callejas A, Rios L, Oyama K, Denno RF (1998) The effects of domestication of Brassica and Phaseolus on the interaction between phytophagous insects and parasitoids. Biol Control 11:130–140. https://doi.org/10.1006/bcon.1997.0590
Berenbaum MR, Zangerl AR, Nitao JK (1986) Constraints on chemical coevolution: wild parsnips and the parsnips webworm. Evolution 40:1215–1228
Blancas J, Casas A, Pérez-Salicrup D, Caballero J, Vega E (2013) Ecological and socio-cultural factors influencing plant management in Náhuatl communities of the Valle de Tehuacán, Mexico. J Ethnobiol Ethnomed. https://doi.org/10.1186/1746-4269-9-39
Blancas J, Pérez-Salicrup D, Casas A (2014) Evaluando la incertidumbre en la disponibilidad de recursos vegetales. Gaia Scientia 8:137–160
Bravo-Avilez D (2017) Variación en los niveles de daño, mecanismos de defensa y adecuación en dos cactáceas columnares del centro de México bajo domesticación. Tesis doctoral. Doctorado en Ciencias Biológicas. Universidad Autónoma Metropolitana
Bravo-Avilez D, Rendón-Aguilar B, Zavala-Hurtado JA, Fornoni J (2014) Primer registro de Cactophagus spinolae (Coleoptera: Curculionidae) sobre dos especies de Stenocereus (Cactaceae) en el centro de México. Rev Mex Biodivers 85:972–974. https://doi.org/10.7550/rmb.43764
Bravo-Avilez D, Zavala-Hurtado JA, Rendón-Aguilar B (2019a) Damage in Cactaceae, their geographic distribution and new evidences. Bot Sci 97:551–567
Bravo-Avilez D, Navarrete-Heredia JL, Rendón-Aguilar B (2019b) New hosts of insects associated with the process of rot damage in edible columnar cacti of central Mexico. Southwest Entomol 44:637–646. https://doi.org/10.3958/059.044.0309
Bravo-Avilez D, Rendón-Aguilar B (in process) Characterization of damage types and their effect on fitness in the pitayas of central Mexico: S. pruinosus and S. stellatus with different management forms. Polibotánica.
Caballero J, Casas A, Cortés L, Mapes C (1998) Patrones en el conocimiento, uso y manejo de plantas en pueblos indígenas de México. Estud Atacamenos 16:181–195. https://doi.org/10.22199/S07181043.1998.0016.00005
Cahill JF, Kembel SW, Gustafson DJ (2005) Differential genetic influences on competitive effect and response in Arabidopsis thaliana. J Ecol 93:958–967. https://doi.org/10.1111/j.1365-2745.2005.01013.x
Casas A (2002) Uso y manejo de cactáceas columnares mesoamericanas. CONABIO Biodiversitas 40:18–23
Casas A, Caballero J (1996) Traditional management and morphological variation in Leucaena esculenta (Fabaceae: Mimosoideae) in the mixtec region of Guerrero, Mexico. Econ Bot 50:167–181. https://doi.org/10.1007/BF02861449
Casas A, Pickersgill B, Caballero J, Valiente-Banuet A (1997) Etnobotany and domestication in Xoconochtli, Stenocereus stellatus (Cactaceae), in the Valle de Tehuacán and La Mixteca Baja, México. Econ Bot 51:279–292. https://doi.org/10.1007/BF02862097
Casas A, Caballero J, Valiente-Banuet A, Soriano JA, Dávila P (1999) Morphological variation and the process of domestication of Stenocereus stellatus (Cactaceae) in Central Mexico. Am J Bot 86:522–533. https://doi.org/10.2307/2656813
Casas A, Cruse-Sanders J, Morales E, Otero-Arnaiz A, Valiente-Banuet A (2006) Maintenance of phenotypic and genotypic diversity in managed populations of Stenocereus stellatus (Cactaceae) by Indigenous Peoples in Central Mexico. Biodivers Conserv 15:879–898. https://doi.org/10.1007/s10531-004-2934-7
Casas A, Otero-Arnaiz A, Pérez-Negrón E, Valiente-Banuet A (2007) In situ management and domestication of plants in Mesoamerica. Ann Bot 100:1101–1115. https://doi.org/10.1093/aob/mcm126
Casas A, Caballero J (1998) Diversidad morfológica y genética de xoconochtli Stenocereus stellatus (Pfeiffer) Riccobono (Cactaceae): conocimiento, uso y estrategias para su conservación. Informe final SNIB-CONABIO proyecto No. G023. http://www.conabio.gob.mx/institucion/proyectos/resultados/InfG023.pdf. Accessed 24 June 2020
Casas A, Parra F (2016) La domesticación como proceso evolutivo. En: Casas A, Torres-Guevara J, Parra F (eds.) Domesticación en el continente americano Volumen 1, Manejo de biodiversidad y evolución dirigida por las culturas del Nuevo Mundo. Universidad Nacional Autónoma de México/Universidad Nacional Agraria La Molina
Casas A, Camou A, Otero-Arnaiz A, Rangel-Landa S, Cruse-Sanders J, Solís L, Torres I, Delgado A, Moreno-Calles AI, Vallejo M, Guillén S, Blancas J, Parra F, Farfán-Heredia B, Aguirre-Dugua X, Arellanes Y (2014) Manejo tradicional de biodiversidad y ecosistemas en Mesoamérica: el Valle de Tehuacán. Investigación Ambiental, Ciencia y política pública
Casas A, Parra F, Blancas J (2015) Evolution of humans and by humans. In: Albuquerque UP, de Medeiros PM, Casas A (eds) Evolutionary Ethnobiology. Springer, Berlin, pp 21–36
Casas A (2001) Silvicultura y domesticación de plantas en Mesoamérica. En: Rendón-Aguilar B, Rebollar-Domínguez S, Caballero-Nieto J, Martínez-Alfaro MA (eds) Plantas, cultura y sociedad, Estudio sobre la relación entre seres humanos y plantas en los albores del siglo XXI. México, pp. 123–157
Castagneyrol B, Lagache L, Giffard B, Kremer A, Jactel H (2012) Genetic diversity increases insect herbivory on oak saplings. PLoS ONE. https://doi.org/10.1371/journal.pone.0044247
Chaudhary B (2013) Plant domestication and resistance to herbivory. Int J Plant Genom. https://doi.org/10.1155/2013/572784
Crawford KM, Crutsinger GM, Sanders NJ (2007) Host-plant genotypic diversity mediates the distribution of an ecosystem engineer. Ecology 88:2114–2120. https://doi.org/10.1890/06-1441.1
Cronin JT, Abrahamson WG (2001) Goldenrod stem galler preference and performance: effects of multiple herbivores and plant genotypes. Oecologia 127:87–96. https://doi.org/10.1007/s004420000561
Cruse-Sanders JM, Parker KC, Friar EA, Huang DI, Mashayekhi S, Prince LM, Otero-Arnaiz A, Casas A (2013) Managing diversity: domestication and gene flow in Stenocereus stellatus Riccob. (Cactaceae) in Mexico. Ecol Evol 3:1340–1355. https://doi.org/10.1002/ece3.524
da Silva MGS, Dubeux CB Jr, Assis LCDSLC, Mota DL, da Silva LLS, dos Santos MVF, dos Santos DC (2010) Anatomy of different forage cacti with contrasting insect resistance. J Arid Environ 74:718–722
de Oliveira AJB, da Silva Machado MdFP (2003) Alkaloid production by callous tissue cultures of Cereus peruvianus (Cactaceae). Appl Biochem Biotechnol 104:149–155
Doebley JF, Gaut BS, Smith BD (2006) The molecular genetics of crop domestication. Cell 127:1309–1321. https://doi.org/10.1016/j.cell.2006.12.006
Doebley J (1992) Molecular systematics and crop evolution. In: Soltis PS, Soltis D, Doyle JJ (eds) Molecular Systematics of Plants. Chapman and Hall, London, pp 202–222
Earl DA, vonHoldt BM (2012) STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Resour 4: 359–361. http://taylor0.biology.ucla.edu/structureHarvester/
Ehrlich PR, Raven PH (1964) Butterflies and plants: a study in coevolution. Evolution 18:586–608
El-Seedi HR, De Smet PAGM, Beck O, Possnert G, Bruhn JG (2005) Prehistoric peyote use: alkaloid analysis and radiocarbon dating of archaeological specimens of Lophophora from Texas. J Ethnopharmacol 101:238–242
Evanno G, Regnaut S, Goudet J (2005) Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol 14:2611–2620. https://doi.org/10.1111/j.1365-294X.2005.02553.x
Excoffier L, Laval G, Schneider S (2005) Arlequín ver. 3.0: an integrated software package for population genetics data analysis. Evol Bioinform 1:1–47. https://doi.org/10.1177/117693430500100003
Falcão HM, Oliveira MT, Mergulhão AC, Silva MV, Santos MG (2012) Ecophysiological performance of three Opuntia ficus-indica cultivars exposed to carmine cochineal under field conditions. Sci Hortic 150:419–424
Fornoni J, Núñez-Farfan J (2000) Evolutionary ecology of Datura stramonium: genetic variation and cost for tolerance to defoliation. Evolution 54:789–797
Gepts P (2003) Crop domestication as a long-term selection experiment. Plant Breeding Rev. https://doi.org/10.1002/9780470650288.ch1
Gepts P (2006) Plant genetic resources conservation and utilization: the accomplishments and future of a societal insurance policy. Crop Sci 46:2278–2292. https://doi.org/10.2135/cropsci2006.03.0169gas
Gomez KA, Gomez AA (1984) Statistical procedures for agricultural research, 2nd edn. Wiley, New York
Gow JL, Peichel CL, Taylor EB (2006) Contrasting hybridization rates between sympatric three spine sticklebacks highlight the fragility of reproductive barriers between evolutionarily young species. Mol Ecol 15:739–752
Guzmán U, Arias S, Dávila PD (2003) Catálogo de cactáceas mexicanas. Universidad Nacional Autónoma de México, Comisión Nacional para el conocimiento y uso de la Biodiversidad (CONABIO), México
Harlan J (1975) Crops and man. American Society of Agronomy, Madison, Wisconsin, USA
Hawkes JK (1983) The diversity of crop plants. Harvard University Press, London
Hayes P, Prehn D, Vivar H, Blake T, Comeau A, Henry I, Johnston M, Jones B, Steffenson B, St Pierre CA, Chen F (1996) Multiple disease resistance loci and their relationship to agronomic and quality loci in a spring barley population. J Quant Trait Loci (http://www.ncgr.org/research/jag/papers96/#paper296/jqtl22.html)
Hayes P, Castro A, Marquez-Cedillo L, Corey A, Henson C, Jones BL, Kling J, Mather D, Matus I, Rossi C, Sato K (2003) Genetic diversity for quantitatively inherited agronomic and malting quality traits. In: von Bothmer R, Knupffer H, van Hintum T, Sato K (eds) Diversity in Barley. Elsevier Science Publishers, Amsterdam Netherlands, pp 201–226
Herms DA, Mattson WJ (1992) The dilemma of plants: to grow or defend. Q Rev Biol 67:283–335
Huot B, Yaoa J, Montgomery BL, He SY (2014) Growth-defense tradeoffs in plants: a balancing act to optimize fitness molec. Plant 7:1267–1287
Hyten DL, Song Q, Zhu Y, Choi I, Nelson RL, Costa JM, Specht JE, Shoemaker RC, Cregan PB (2006) Impacts of genetic bottlenecks on soybean genome diversity. PNAS 103:16666–16671
Karley AJ, Hawes C, Ianneta PP, Squire GR (2008) Intraspecific variation in Capsella bursa-pastoris in plant quality traits for insect herbivores. Weed Res 48:147–156. https://doi.org/10.1111/j.1365-3180.2007.00617.x
Kovach MJ, McCouch SR (2008) Leveraging natural diversity: back through the bottleneck. Curr Opin Plant Biol 11:193–200. https://doi.org/10.1016/j.pbi.2007.12.006
Langella O (1999) POPULATIONS 1.2.32- Population Genetic Software http://www.mybiosoftware.com/populations-1-2-32-population-genetic-software.html
Langlie BS, Mueller NG, Spengler RN, Fritz GJ (2014) Agricultural origins from the ground up: archaeological approaches to plant domestication. Am J Bot 101:1601–1617. https://doi.org/10.3732/ajb.1400145
Li X, Garvey M, Kaplan I, Li B, Carrillo J (2017) Domestication of tomato has reduced the attraction of herbivore natural enemies to pest-damaged plants. Agr Forest Entomol 20:390–401. https://doi.org/10.1111/afe.12271
Lindig R, Casas A (2013) Experiencias docentes en integración de conocimientos ambientales en comunidades rurales. En: Camou-Guerrero A, Castillo A, García-Frapolli E (eds) Procesos de formación educativa interdisciplinaria: miradas desde las Ciencias Ambientales. Universidad Nacional Autónoma de México, pp. 125–144
MacNeish RS (1967) An interdisciplinary approach to an archaeological problem. In: Byers DS (ed) The prehistory of the Tehuacan Valley Environment and subsistence. University of Texas Press, London, pp 14–24
Maddox GD, Root RB (1987) Resistance to 16 diverse species of herbivorous insects within a population of goldenrod, Solidago altissima: genetic variation and heritability. Oecologia 72:8–14. https://doi.org/10.1007/BF00385037
Martínez-Ortega RM, Tuya-Pendás LC, Martínez-Ortega M, Pérez-Abreu A, Cánovas AM (2009) El coeficiente de correlación de los rangos de Spearman caracterización. Rev Haban Cienc Méd
Maxted N, Ford-Lloyd BV, Hawkes JG (2013) Plant genetic conservation: the in situ approach. Springer, Berlin
Minitab (2007) Minitab. Rel. 12 for Windows. Minitab, State College, PA. Minitab Inc
Monreal-Vargas CT, Méndez EE (2014) Hongos patógenos del garambullo Myrtillocactus geometrizan (Mart. ex. Pfeiff.) Console en Mexquitic de Carmona, San Luis Potosí. México Revista Iberoamericana De Ciencias 1:45–59
Moreira X, Abdala-Roberts L, Gols R, Francisco M (2018) Plant domestication decreases both constitutive and induced chemical defences by direct selection against defensive traits. Sci Rep 8:12678. https://doi.org/10.1038/s41598-018-31041-0
Mosquera T, Fernández C, Martínez L, Acuña A, Cuéllar D (2008) Genética de la resistencia de la papa (Solanum tuberosum) a patógenos. Estado De Arte Agronomía Colombiana 26:7–15
Murren CJ (2002) Phenotypic integration in plants. Plant Species Biol 17:89–99
Otero-Arnaiz A, Casas A, Hamrick JL, Cruse-Sanders J (2005) Genetic variation and evolution of Polaskia chichipe (Cactaceae) under domestication in the Valle de Tehuacán, Central Mexico. Mol Ecol 14:1603–1611. https://doi.org/10.1111/j.1365-294X.2005.02494.x
Parra F, Casas A, Peñaloza-Ramírez JM, Cortés-Palomee AC, Rocha-Ramírez V, González-Rodríguez A (2010) Evolution under domestication: ongoing artificial selection and divergence of wild and managed Stenocereus pruinosus (Cactaceae) populations in the Tehuacán Valley, Mexico. Ann Bot 106:483–496
Peakall R, Smouse PE (2005) GENALEX 6.5: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6:288–295. https://doi.org/10.1111/j.1471-8286.2005.01155.x
Pickersgill B (2007) Domestication of plants in the Americas: insights from mendelian and molecular genetics. Ann Bot 100:925–940. https://doi.org/10.1093/aob/mcm193
Piperno DR (2011) The origins of plant cultivation and domestication in the new world tropics: patterns, process, and new developments. Curr Anthropol 52:S453–S470. https://doi.org/10.1086/659998
Poulicard N, Pacios LF, Gallois J-L, Piñero D, García-Arenal F (2016) Human management of a wild plant modulates the evolutionary dynamics of a gene determining recessive resistance to virus infection. PLoS Genet 12(8):e1006214. https://doi.org/10.1371/journal.pgen.1006214
Pritchard JK, Stephens M, Donnelly P (2000) Inference of population structure using multilocus genotype data. Genetics 155:945–959
Raymond M, Rousset F (1995) GENEPOP (Version 1.2): population genetics software for exact test and ecumenicism. J Hered 86:248–249. https://doi.org/10.1093/oxfordjournals.jhered.a111573
Rendón B, Núñez-Farfán J (1998) Genética evolutiva del proceso de domesticación en plantas. Bot Sci 63:131–151
Restif O, Koella JC (2004) Concurrent evolution of resistance and tolerance to pathogens. Am Nat 164(4):E90–E102
Rosenthal JP, Dirzo R (1997) Effects of life history, domestication and agronomic selection on plant defense against insects: Evidence from maizes and wild relatives. Evol Ecol 11:337–355. https://doi.org/10.1023/A:1018420504439
Roy BA, Kirchner JW (2000) Evolutionary dynamics of pathogen resistance and tolerance. Evolution 54:51–63
Sall J, Creighton L, Lehman A (2017) JMP® start statistics a guide to statistics and data analysis using JMP®. SAS, Cary
Schaller A (2008) Induced plant resistance to herbivory. Springer, Dordrecht. https://doi.org/10.1007/978-1-4020-8182-8
Schwanitz F (1966) The origin of cultivated plants. Harvard University Press, Cambridge
Smith C (1967) Plant remains. In: Byers D (ed) The prehistory of Tehuacan Valley. University of Texas Press, Austin
Sneath P, Sokal RR (1973) Numerical Taxonomy. The Principles and Practice of Numerical Classification. W.H. Freeman & Company, San Francisco
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant tolerance to herbivory. Trends Ecol Evol 14:179–185
Van Oosterhout C, Hutchinson WF, Willis D, Shipley P (2004) MICROCHECKER software for identifying and correcting genotyphing errors in microsatellites data. Mol Ecol Notes 4:535–538. https://doi.org/10.1111/j.147
Wise MJ (2007) Evolutionary ecology of resistance to herbivory: an investigation of potential genetic constraints in the multiple-herbivore community of Solanum carolinense. New Phytol 175:773–784. https://doi.org/10.1111/j.1469-8137.2007.02143.x
Acknowledgements
We wish to thank the Laboratory of Molecular Biology of the Universidad Autónoma Metropolitana Iztapalapa for the support in the analysis of molecular samples. To the Master in Biology program, which provided a scholarship to the first author. Alejandra Serrato-Díaz for all logistic support, advice, and guidance to the first author. Dr. Luis Guevara Chumacero for the support in the loan of reagents. We deeply appreciate the permission, trust and support of all the people and authorities of the communities in the states of Puebla and Oaxaca, where the research was developed, in particular to Mrs. Martha and her husband from Miahuatlán, Puebla, and Mr. Apuleyo Hernández from Chinango, Oaxaca. To anonymous reviewers who improved the manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was financially supported by the Master in Biology program, at the Universidad Autónoma Metropolitana Iztapalapa, and personal resources of the authors.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The author(s) declared no potential competing interests with respect to the research, authorship, and/or publication of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Patiño-López, V., Bravo-Avilez, D., Vargas-Mendoza, C.F. et al. Evolution under domestication of correlated characters in populations of Stenocereus stellatus (Pfeiff.) Riccob., under different forms of management in central Mexico: genetic diversity, damage, and defense mechanisms. Genet Resour Crop Evol 69, 601–618 (2022). https://doi.org/10.1007/s10722-021-01245-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-021-01245-x