
Abstract
The photoperiod is an important environmental stress that determines flowering time. The CONSTANS (CO) and Heading date 1 (Hd1) genes are known to be central integrators of the photoperiod pathway in Arabidopsis and rice, respectively. Although they are both members of the CONSTANS-LIKE (COL) family and have two B-boxes and a CCT domain, rice also possesses novel COL genes that are not found in Arabidopsis. Here, we demonstrate that a novel COL gene, OsCO3, containing a single B-box and a CCT domain, modulates photoperiodic flowering in rice. The circadian expression pattern of OsCO3 mRNA oscillated in a different phase from Hd1 and was similar to that of OsCO3 pre-mRNA, suggesting that the diurnal expression pattern of OsCO3 transcripts may be regulated at the transcriptional level. Overexpression of OsCO3 specifically caused late flowering under short day (SD) conditions relative to wild-type rice plants. The expression of Hd3a and FTL decreased in these transgenic plants, whereas the expression of Hd1, Early heading date 1 (Ehd1), OsMADS51, and OsMADS50 did not significantly change. Our results suggest that OsCO3 primarily controls flowering time under SD conditions by negatively regulating Hd3a and FTL expression, independent of the SD-promotion pathway.





Similar content being viewed by others


Abbreviations
- AP1:
-
APETALA1
- CCT:
-
CO, CO-like, and TOC1
- CO:
-
CONSTANS
- COL:
-
CO-Like
- Ehd1:
-
Early heading date 1
- FT:
-
FLOWERING LOCUS T
- HAPs:
-
Heme Activator Proteins
- Hd1:
-
Heading date 1
- SOC1:
-
SUPPRESSOR OF OVEREXPRESSION OF CO1
References
Ben-Naim O, Eshed R, Parnis A, Teper-Bamnolker P, Shalit A, Coupland G, Samach A, Lifschitz E (2006) The CCAAT binding factor can mediate interactions between CONSTANS-like proteins and DNA. Plant J 46:462–476
Borden KL (1998) RING fingers and B-boxes: zinc-binding protein-protein interaction domains. Biochem Cell Biol (Biochimie Biol Cellulaire) 76:351–358
Cai X, Ballif J, Endo S, Davis E, Liang M, Chen D, DeWald D, Kreps J, Zhu T, Wu Y (2007) A putative CCAAT-binding transcription factor is a regulator of flowering timing in Arabidopsis. Plant Physiol 145:98–105
Cheng XF, Wang ZY (2005) Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J 43:758–768
Datta S, Hettiarachchi GH, Deng XW, Holm M (2006) Arabidopsis CONSTANS-LIKE3 is a positive regulator of red light signaling and root growth. Plant Cell 18:70–84
Doi K, Izawa T, Fuse T, Yamanouchi U, Kubo T, Shimatani Z, Yano M, Yoshimura A (2004) Ehd1, a B-type response regulator in rice, confers short-day promotion of flowering and controls FT-like gene expression independently of Hd1. Genes Dev 18:926–936
Griffiths S, Dunford RP, Coupland G, Laurie DA (2003) The evolution of CONSTANS-like gene families in barley, rice, and Arabidopsis. Plant Physiol 131:1855–1867
Hayama R, Coupland G (2004) The molecular basis of diversity in the photoperiodic flowering responses of Arabidopsis and rice. Plant Physiol 135:677–684
Hayama R, Yokoi S, Tamaki S, Yano M, Shimamoto K (2003) Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 422:719–722
Hiei Y, Ohta S, Komari T, Kumashiro T (1994) Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 6:271–282
Imaizumi T, Kay SA (2006) Photoperiodic control of flowering: not only by coincidence. Trends Plant Sci 11:550–558
Imaizumi T, Schultz TF, Harmon FG, Ho LA, Kay SA (2005) FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 309:293–297
Izawa T, Oikawa T, Sugiyama N, Tanisaka T, Yano M, Shimamoto K (2002) Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev 16:2006–2020
Jeon JS, Jang S, Lee S, Nam J, Kim C, Lee SH, Chung YY, Kim SR, Lee YH, Cho YG, An G (2000) leafy hull sterile1 is a homeotic mutation in a rice MADS box gene affecting rice flower development. Plant Cell 12:871–884
Kardailsky I, Shukla VK, Ahn JH, Dagenais N, Christensen SK, Nguyen JT, Chory J, Harrison MJ, Weigel D (1999) Activation tagging of the floral inducer FT. Science 286:1962–1965
Kim SL, Lee S, Kim HJ, Nam HG, An G (2007) OsMADS51 is a short-day flowering promoter that functions upstream of Ehd1, OsMADS14, and Hd3a. Plant Physiol 145:1484–1494
Kobayashi Y, Weigel D (2007) Move on up, it’s time for change mobile signals controlling photoperiod-dependent flowering. Genes Dev 21:2371–2384
Kobayashi Y, Kaya H, Goto K, Iwabuchi M, Araki T (1999) A pair of related genes with antagonistic roles in mediating flowering signals. Science 286:1960–1962
Kojima S, Takahashi Y, Kobayashi Y, Monna L, Sasaki T, Araki T, Yano M (2002) Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol 43:1096–1105
Ledger S, Strayer C, Ashton F, Kay SA, Putterill J (2001) Analysis of the function of two circadian-regulated CONSTANS-LIKE genes. Plant J 26:15–22
Lee S, An G (2007) Diversified mechanisms for regulating flowering time in a short-day plant rice. J Plant Biol 50:241–248
Lee H, Suh SS, Park E, Cho E, Ahn JH, Kim SG, Lee JS, Kwon YM, Lee I (2000) The AGAMOUS-LIKE 20 MADS domain protein integrates floral inductive pathways in Arabidopsis. Genes Dev 14:2366–2376
Lee S, Kim J, Han JJ, Han MJ, An G (2004) Functional analyses of the flowering time gene OsMADS50, the putative SUPPRESSOR OF OVEREXPRESSION OF CO 1/AGAMOUS-LIKE 20 (SOC1/AGL20) ortholog in rice. Plant J 38:754–764
Lee JH, Cho YS, Yoon HS, Suh MC, Moon J, Lee I, Weigel D, Yun CH, Kim JK (2005) Conservation and divergence of FCA function between Arabidopsis and rice. Plant Mol Biol 58:823–838
Martin J, Storgaard M, Andersen CH, Nielsen KK (2004) Photoperiodic regulation of flowering in perennial ryegrass involving a CONSTANS -like homolog. Plant Mol Biol 56:159–169
Michaels SD, Himelblau E, Kim SY, Schomburg FM, Amasino RM (2005) Integration of flowering signals in winter-annual Arabidopsis. Plant Physiol 137:149–156
Miyoshi K, Ito Y, Serizawa A, Kurata N (2003) OsHAP3 genes regulate chloroplast biogenesis in rice. Plant J 36:532–540
Nemoto Y, Kisaka M, Fuse T, Yano M, Ogihara Y (2003) Characterization and functional analysis of three wheat genes with homology to the CONSTANS flowering time gene in transgenic rice. Plant J 36:82–93
Oh SJ, Jeong JS, Kim EH, Yi NR, Yi SI, Jang IC, Kim YS, Suh SC, Nahm BH, Kim JK (2005) Matrix attachment region from the chicken lysozyme locus reduces variability in transgene expression and confers copy number-dependence in transgenic rice plants. Plant Cell Rep 24:145–154
Onouchi H, Igeno MI, Perilleux C, Graves K, Coupland G (2000) Mutagenesis of plants overexpressing CONSTANS demonstrates novel interactions among Arabidopsis flowering-time genes. Plant Cell 12:885–900
Putterill J, Robson F, Lee K, Simon R, Coupland G (1995) The CONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 80:847–857
Robson F, Costa MM, Hepworth SR, Vizir I, Pineiro M, Reeves PH, Putterill J, Coupland G (2001) Functional importance of conserved domains in the flowering-time gene CONSTANS demonstrated by analysis of mutant alleles and transgenic plants. Plant J 28:619–631
Samach A, Onouchi H, Gold SE, Ditta GS, Schwarz-Sommer Z, Yanofsky MF, Coupland G (2000) Distinct roles of CONSTANS target genes in reproductive development of Arabidopsis. Science 288:1613–1616
Shin BS, Lee JH, Lee JH, Jeong HJ, Yun CH, Kim JK (2004) Circadian regulation of rice (Oryza sativa L.) CONSTANS-like gene transcripts. Mol Cells 17:10–16
Strayer C, Oyama T, Schultz TF, Raman R, Somers DE, Mas P, Panda S, Kreps JA, Kay SA (2000) Cloning of the Arabidopsis clock gene TOC1, an autoregulatory response regulator homolog. Science 289:768–771
Suarez-Lopez P, Wheatley K, Robson F, Onouchi H, Valverde F, Coupland G (2001) CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 410:1116–1120
Takahashi Y, Shomura A, Sasaki T, Yano M (2001) Hd6, a rice quantitative trait locus involved in photoperiod sensitivity, encodes the alpha subunit of protein kinase CK2. Proc Natl Acad Sci USA 98:7922–7927
Tamaki S, Matsuo S, Wong HL, Yokoi S, Shimamoto K (2007) Hd3a protein is a mobile flowering signal in rice. Science 316:1033–1036
Trevaskis B, Hemming MN, Dennis ES, Peacock WJ (2007) The molecular basis of vernalization-induced flowering in cereals. Trends Plant Sci 12:352–357
Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G (2004) Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science 303:1003–1006
Wenkel S, Turck F, Singer K, Gissot L, Le Gourrierec J, Samach A, Coupland G (2006) CONSTANS and the CCAAT box binding complex share a functionally important domain and interact to regulate flowering of Arabidopsis. Plant Cell 18:2971–2984
Yan L, Loukoianov A, Blechl A, Tranquilli G, Ramakrishna W, SanMiguel P, Bennetzen JL, Echenique V, Dubcovsky J (2004) The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science 303:1640–1644
Yan L, Fu D, Li C, Blechl A, Tranquilli G, Bonafede M, Sanchez A, Valarik M, Yasuda S, Dubcovsky J (2006) The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc Natl Acad Sci USA 103:19581–19586
Yano M, Katayose Y, Ashikari M, Yamanouchi U, Monna L, Fuse T, Baba T, Yamamoto K, Umehara Y, Nagamura Y, Sasaki T (2000) Hd1, a major photoperiod sensitivity quantitative trait locus in rice, is closely related to the Arabidopsis flowering time gene CONSTANS. Plant Cell 12:2473–2484
Yano M, Kojima S, Takahashi Y, Lin H, Sasaki T (2001) Genetic control of flowering time in rice, a short-day plant. Plant Physiol 127:1425–1429
Yanovsky MJ, Kay SA (2002) Molecular basis of seasonal time measurement in Arabidopsis. Nature 419:308–312
Yoo SK, Chung KS, Kim J, Lee JH, Hong SM, Yoo SJ, Yoo SY, Lee JS, Ahn JH (2005) CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through FLOWERING LOCUS T to promote flowering in Arabidopsis. Plant Physiol 139:770–778
Acknowledgments
We are grateful to B.S. Shin and J.H. Lee for the OsCO3 cDNA clones. This study was supported by a grant from the Plant Signaling Network Research Center, Korea Science and Engineering Foundation, Republic of Korea. S-K. Kim, Y.H. Jang and H-Y. Park were supported by a BK21 research fellowship from the Korean Ministry of Education and Human Resources Development. J.H. Lee was supported by the Korean Research Foundation Grant (KRF-2007-359-C00023) funded by Korea Government (MOEHRD).
Author information
Authors and Affiliations
Corresponding author
Additional information
Soon-Kap Kim, Choong-Hyo Yun and Jeong Hwan Lee contributed equally to this work.
Electronic supplementary material
Fig6
Fig. S1 Overexpression of exogenous OsCO3 in OsCO3 T2 lines. The samples were grown under natural (LD-like) conditions. Total RNA was isolated from leaves and Northern-blot analysis was performed to confirm the overexpression of the transgene in each line. OsCO3 full cDNA was employed as a probe
Fig7
Fig. S2 The expression of exogenous and endogenous OsCO3 in OsCO3 transgenic lines grown under SD conditions. Total RNA was isolated from the samples grown for 50, 59, 66, 80, 94, and 110 days. The OsUBQ was employed as a loading control
Fig8
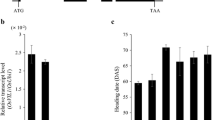
Fig. S3 Flowering times of transgenic rice plants overexpressing OsCO3 at the T3 generation. Flowering time of OsCO3 transgenic plants grown under SD conditions. In each transgenic line, mean values of 10-15 plants with standard errors were used (in some lines without error bars, 1-2 plants were used for measurement of flowering). The flowering times of non-transgenic segregating T3 plants in some lines are not shown, because all segregating plants contained the transgene (PCR analysis was performed to determine the presence of the transgene, although the genotype of each plant was not classified into heterozygote or homozygote). T and N indicate segregating T3 plants with and without the transgene, respectively
Fig9
Fig. S4 The expression of exogenous and endogenous OsCO3 in OsCO3 transgenic lines grown under SD conditions. Total RNA was isolated from the samples grown for 50, 59, 66, 80, 94, and 110 days. The OsUBQ was employed as a loading control
Fig10
Fig. S5 The expression of Hd3a, FTL, and RFT1 in OsCO3 transgenic lines. The samples are the same as those in Fig. S2. OsMADS14/RAP1B was used as a molecular marker that indicates the floral transition
Rights and permissions
About this article
Cite this article
Kim, SK., Yun, CH., Lee, J.H. et al. OsCO3, a CONSTANS-LIKE gene, controls flowering by negatively regulating the expression of FT-like genes under SD conditions in rice. Planta 228, 355–365 (2008). https://doi.org/10.1007/s00425-008-0742-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-008-0742-0