
Abstract
High-severity fires in dry conifer forests of the United States Southwest have created large (>1000 ha) treeless areas that are unprecedented in the regional historical record. These fires have reset extensive portions of Southwestern ponderosa pine (Pinus ponderosa Lawson & C. Lawson var. scopulorum Engelm.) forest landscapes. At least two recovery options following high-severity fire are emerging. One option is for post-fire successional pathways to move toward a return to the pre-fire forest type. Alternatively, an area may transition to persistent non-forested ecosystems. We studied regeneration patterns of ponderosa pine following eight fires in Arizona and New Mexico, USA, that burned in dry conifer forests dominated by ponderosa pine during a recent 18-year regional drought period, 1996 to 2013. Our a priori hypotheses were: 1) the most xeric areas within these severely burned dry conifer forests are least likely to regenerate to the pre-fire forest type due to persistent post-fire moisture stress; and 2) areas farther away from conifer seed sources have a lower likelihood of regeneration, even if these areas are climatically favorable for post-fire ponderosa pine establishment. We evaluated our hypotheses using empirical data and generalized linear mixed-effects models. We found that low-elevation, xeric sites are more limiting to conifer regeneration than higher-elevation mesic sites. Areas >150 m from a seed source are much less likely to have ponderosa pine regeneration. Spatial interpolations of modeled post-fire regeneration of ponderosa pine across the study landscapes indicate expansive areas with low likelihood of pine regeneration following high-severity fire. We discuss multiple post-fire successional pathways following high-severity fire, including potentially stable transitions to non-forest vegetation types that may represent long-term type conversions. These findings regarding landscape changes in Southwest forests in response to fires and post-fire regeneration patterns during early-stage climate warming contribute to the development of better-informed ecosystem management strategies for forest adaptation or mitigation under projected hotter droughts in this region.
Resumen
Los fuegos de alta severidad en bosques secos de coníferas del suroeste de los Estados Unidos han creado grandes áreas denudadas de árboles (>1000 ha) que no tienen precedentes en los registros históricos regionales. Estos fuegos han remodelado extensas porciones del paisaje en bosques de pino ponderosa (Pinus ponderosa Lawson & C. Lawson var. scopulorum Engelm.). Al menos dos opciones de recuperación están emergiendo luego de fuegos de alta severidad. Una de las opciones implica un rumbo sucesional post-fuego tendiente a retornar al tipo de bosque previo al fuego. Alternativamente, un área puede hacer la transición hacia ecosistemas sin bosques de manera permanente. Estudiamos los patrones de regeneración de pino ponderosa después de la ocurrencia de ocho incendios en Arizona y Nuevo Méjico, EEUU, acaecidos en bosques secos de coníferas dominados por pino ponderosa, en un período reciente de sequía regional de 18 años, desde 1996 a 2013. Nuestras hipótesis a priori fueron: 1) es improbable que las áreas más xéricas dentro de estos bosques severamente quemados regeneren el tipo de bosque anterior al fuego, debido a un estrés de humedad permanente post fuego; 2) las áreas más alejadas de fuentes de semillas de coníferas tienen una probabilidad menor de regeneración, aun si estas áreas fueran climáticamente más favorables para el establecimiento post fuego de pino ponderosa. Evaluamos nuestras hipótesis utilizando datos empíricos y modelos lineales generalizados de efectos mixtos. Encontramos que elevaciones bajas (sitios xéricos) son más limitantes para la regeneración de coníferas que las elevaciones altas, representativas de sitios mésicos. Áreas que se encuentran a >150 m de una fuente de semillas tienen una menor probabilidad de tener regeneración de pino ponderosa. Interpolaciones espaciales de modelos de regeneración post fuego de pino ponderosa en los paisajes de estudio, indicaron que las posibles áreas de expansión tienen baja probabilidad de regeneración de este pino después de fuegos severos. Nosotros discutimos múltiples caminos sucesionales luego de fuegos de alta severidad, incluyendo transiciones potencialmente estables hacia tipos de vegetación sin bosque que pueden representar conversiones a largo plazo. Estos hallazgos, relacionados con cambios a nivel de paisaje en los bosques del suroeste, en respuesta a fuegos y a los patrones de regeneración post fuego en las primeras fases del calentamiento global, contribuyen al desarrollo de estrategias de manejo de ecosistemas con mejor información, para la adaptación de los bosques o su mitigación, en el caso de sequías cálidas pronosticadas para esta región.
Similar content being viewed by others
Introduction
Since the mid 1990s, the frequency and size of high-severity wildfire activity in the southwestern United States has been increasing in dry conifer forests that are dominated or co-dominated by ponderosa pine (Pinus ponderosa Lawson & C. Lawson var. scopulorum Engelm.; Westerling et al. 2006, Dennison et al. 2014, Abatzoglou and Williams 2016, Allen 2016). Factors contributing to this increase in high-severity fire activity include uncharacteristically dense and homogeneous forest structure due to a century of fire exclusion (Covington 2000), natural fluctuations in climate driven by the El Niño Southern Oscillation and other multi-annual to multi-decadal patterns of ocean variability (Swetnam and Betancourt 1990, 1998; Margolis and Swetnam 2013), and increasing forest drought stress from climate warming (Williams et al. 2013, Tarancon et al. 2014, Allen et al. 2015). The recent emergence of relatively large patches of high-severity fire has created extensive areas with drastically reduced live forest canopy cover, with few or no remnant live trees to serve as seed sources for forest regeneration. Southwestern US ponderosa pine forests generally evolved with low-severity, high-frequency, surface fire regimes (Covington and Moore 1994, Swetnam and Baisan 1996), documented by numerous tree-ring fire histories from ponderosa pine forests in most mountain ranges across Arizona and New Mexico (Falk et al. 2011). Before circa 1900 AD, these low-severity surface fires characterized most Southwest ponderosa pine and dry mixed-conifer forests, burning extensively every 5 to 25 years (Swetnam and Baisan 1996, Touchan et al. 1996, Grissino-Mayer and Swetnam 2000, Reynolds et al. 2013, Margolis and Malevich 2016). High-severity fire did occur in ponderosa pine, but multiple lines of evidence indicate that patches generally were small (from clusters of a few trees to <100 ha), discontinuous, and relatively uncommon (Iniguez et al. 2009, Margolis and Balmat 2009, Fulé et al. 2013, Fornwalt et al. 2016). This contrasts with the large (>1000 ha) high-severity fire patches that have emerged recently in Southwest ponderosa pine forests (e.g., 2011 Las Conchas Fire, 2011 Wallow Fire; Allen 2016).
The scale of these recent tree-killing forest disturbances is unprecedented in the Southwest since historic record keeping began around 1900, almost certainly is unprecedented since the megadrought of late 1500s (Swetnam and Betancourt 1998), and the size of recent high-severity fire patches in Southwestern ponderosa pine forests quite possibly is unprecedented (Fulé et al. 2014) since before modern patterns of climate, vegetation, and fire regimes established 9000 to 6000 years ago (Anderson et al. 2008). During the most recent regional drought, from about 1996 to 2013, multiple large, high-severity fires in the dry conifer forests of Arizona and New Mexico have created historically anomalous high-severity burn patches of many thousands of contiguous hectares (A. Thode, Northern University, Flagstaff, Arizona, USA, unpublished data). This drought represents some of the driest and warmest years of the past century, with mostly negative Palmer Drought Severity Index (PDSI) values across the study region since 1999 (Figure 1). Thus, these large, recent high-severity burn patches, many of which are devoid of any surviving conifer trees post fire (i.e., no on-site seed source for obligate seeders like ponderosa pine), also have been recovering under hot and dry post-fire climate conditions. In response to this combination of historically unprecedented factors, these recent high-severity burn areas may be following post-fire ecological trajectories that move away from persistent forest conditions, transitioning instead toward shrublands or grasslands (i.e., type conversion). Our study examines this hypothesis.

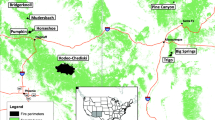
Locations of the eight sampled fires (red polygons) that burned with high severity in dry conifer forests of Arizona and New Mexico, USA (1996 to 2006). Inset shows drought conditions (Palmer Drought Severity Index, PDSI) in the study area from 1904 to 2013 and the year of the sampled fires (red dots). The fires all burned after 1996, when the region entered a period of significant prolonged drought (PDSI data from: https://doi.org/www.wrcc.dri.edu/wwdt/about.html, accessed January 2014).
Limits to Post-Fire Pine Regeneration
There is increasing evidence of limitations in the capacity of ponderosa pine forests to regenerate following large high-severity (i.e., tree-killing) fires in combination with drought conditions in a warmer climate (Bonnet et al. 2005, Lentile et al. 2007, Keyser et al. 2008, Moser et al. 2010, Puhlick et al. 2012, Feddema et al. 2013, Collins and Roller 2013, Savage et al. 2013, Rother et al. 2015, Chambers et al. 2016, Owen et al. 2017). Severely burned patches are often warmer and drier than adjacent landscape patches with intact forest canopy (Meyer et al. 2001). For some burned areas, post-fire regeneration has been observed in areas with mature forest canopy, while seedling regeneration was absent or drastically reduced in areas that burned at high severity, presumably because of warmer and drier conditions in the severely burned areas (Lentile et al. 2005, Crotteau et al. 2013). Open areas created by high-severity forest fire and drought are often colonized by, and can become dominated by, drought-tolerant species of shrubs or grasses (Barton 2002, Savage and Mast 2005, Foxx et al. 2013, Abella and Fornwalt 2015, Guiterman et al. 2017, Barton and Poulos 2018). Areas of hyper-dense conifer regeneration also have been documented post fire in the US Southwest, but such regeneration can be especially susceptible to substantial mortality during re-burns (Savage and Mast 2005).
In the dry conifer forests of the western US and European Mediterranean Basin, post-fire conifer regeneration is low or non-existent at the lowest elevations of the pre-fire species range, which may permanently reduce the range of a species (Gracia et al. 2002, Vilà-Cabrera et al. 2012, Crotteau et al. 2013, Dodson and Root 2013). In both of these seasonally warm-dry regions, post-fire conifer regeneration in open areas is often found in favorable microsites, such as next to logs or under the cover of litter (Bonnet et al. 2005, Castro et al. 2011, Roccaforte et al. 2012, Marañón-Jiménez et al. 2013), and near available seed sources (Rother and Veblen 2016).
The combination of post-fire drought, climate warming, and large high-severity patch size may limit ponderosa pine regeneration in the US Southwest. Ponderosa pine has a heavy, small-winged seed that limits the range of seed dispersal. Multiple studies have found that ponderosa pine establishment is primarily limited to <200 m from a seed source or an intact forest edge (Haire and McGarigal 2010, Dodson and Root 2013, Chambers et al. 2016, Haire et al. 2017). Therefore, the size and shape of high-severity patches alone, which determines the distance to seed sources, may limit the rate of recovery of ponderosa pine forests (Collins et al. 2017).
Hypotheses
We asked the question: do topography and landscape-level patterns in fire severity predict pine regeneration following high-severity fire in US Southwest ponderosa pine forests? To answer this question, we developed hypotheses based on ecological factors known to influence post-fire conifer recovery and applied them to high-severity fires in US Southwest ponderosa pine forests. Our hypotheses were:
-
H1) The likelihood of ponderosa pine regeneration decreases near warm and dry margins of local ponderosa pine landscape distributions, (e.g., lower elevations, ridgetops, and southwestern aspects).
-
H2) The likelihood of ponderosa pine regeneration decreases with greater distance to seed source.
We used empirical data and general linear mixed-effects models to test our hypotheses. We created a spatially explicit representation of the modeling results to show the likelihood of ponderosa pine regeneration across our study sites. We draw on our results to consider management implications of recent and projected trends of more extreme warming, droughts, and fire activity on forest restoration and post-fire succession.
Methods
Study Sites
In 2013, we collected field data in high-severity patches of eight wildfires that burned ponderosa pine forests in Arizona and New Mexico between 1996 and 2006 (Table 1, Figure 1). We selected fires that burned during this 11-year period because: a) this period represented a historically extreme warm-dry climate, with associated extreme fire weather and fire behavior, which are projected to become typical in the near future of the Southwest (Williams et al. 2013, 2014); and b) the fires that occurred during this period provided sufficient time, prior to our 2013 fieldwork year, to allow examination of initial patterns in post-fire vegetation recovery.
We accessed fire perimeters from the US Forest Service spatial fire perimeter database (US Forest Service 2013) and extracted the relevant fire perimeters. To determine the spatial extent of high-severity fire, we used the Relative Differenced Normalized Burn Ratio (RdNBR; Thode and Miller 2007, Miller et al. 2009), derived from the Monitoring Trends in Burn Severity dataset (MTBS.gov), which uses pre- and post-fire Landsat imagery to determine tree canopy change caused by fire (Eidenshink et al. 2007). High-severity patches were identified by contiguous RdNBR pixels with values greater than 640, which represent areas with 95% or greater tree mortality (Miller and Thode 2007).
Field Data Collection
Within the mapped perimeter of each fire, we used the Euclidian distance tool in ArcGIS (ESRI Inc., Redlands, California, USA) to create sampling bands extending into the severely burned area at fixed distances from the forest edge. These bands were established at distance intervals of 0 m to 50 m, 50 m to 100 m, 100 m to 150 m, 150 m to 250 m, and greater than 250 m from the unburned forest. We selected the largest high-severity patch in each fire for field sampling because these largest patches of high-severity fire represent the greatest departure from the historic range of variability in dry-conifer forests (Fulé et al. 2014). Within these patches, we randomly determined field sample site locations within each sample band. Table 1 lists the number of field sample site locations per fire, which is roughly proportional to the area of high-severity fire. Field sampling occurred in the summer of 2013.
At each sample site, we established a multi-part vegetation plot, composed of four subplots. Starting with a central circular subplot, three additional circular subplots were radially arranged 13 m away from the central subplot center, with each radial subplot at 120° intervals, starting from an initial randomly selected bearing. Each subplot included an interior plot of 1.5 m radius (total area = 28.3 m2) nested inside a larger plot with a 3 m radius (total area = 113.1 m2). At each subplot, vegetative cover was classified within the interior 1.5 m radius constituent plot based on physiognomic characteristics—grasses, forbs, shrubs, trees, or logs—and assigned to one of seven different percentage cover classes: absent (0%), 1% to 5%, 5% to 25%, 25% to 50%, 50% to 75%, 75% to 95%, and 95% to 100%. We categorized exposed ground surface as litter, bare ground, or rock using the same percentage classes. Within the outer 3 m radius subplots, we measured all individuals of all tree species ≥15 cm tall, as research suggests that, once a tree grows to that height, it is likely to survive to maturity (Flathers et al. 2016, Waring et al. 2016). For each tree seedling, we recorded species and height. We also determined whether biotic or abiotic nurse structures were positioned to aid in seedling germination or growth (i.e., the structure was within 30 cm of the seedling and could shade the seedling). We directly measured distance to the nearest ponderosa pine seed source at each plot using a rangefinder if possible or, where the distance to the nearest seed source was >500 m (the distance limit of the rangefinder) or the straight-line view was obstructed, we used 1 m resolution aerial imagery from the National Agriculture Imagery Program in a GIS (USFSA 2013) to estimate distance. We defined seed source as one or more cone-producing, mature ponderosa pine, or the nearest edge of the unburned forest containing mature ponderosa pine.
Predictor Variables
We developed spatial data for topographic variables to characterize moisture and temperature gradients to evaluate H1—that ponderosa pine regeneration is less likely in the hottest and driest portions of its current range. We chose three independent topographic landscape metrics as proxies for local climate variability: elevation, aspect, and topographic position index (TPI), which delineates ridges, valleys, and slopes (Parker 1982). The Pumpkin, Horseshoe, and Hochderffer fires were analyzed together, hereafter referred to as the Flagstaff Group, as all occurred in the same general area north of Flagstaff, Arizona. Similarly, the BS and Bear fires were analyzed together as the Gila Group, because they occurred adjacent to each other in the Gila National Forest in New Mexico. In the GIS, we buffered each individual fire, or group of fires, by 2 km to identify the minimum and maximum values for topographic variables across an area larger than the fire perimeter itself.
We employed the following methods to generate spatial data for the topographic variables.
Elevation. We determined the local elevational range of ponderosa pine for each fire by combining a 30 m digital elevation model (DEM) with the ponderosa pine distributions layers from the Southwest ReGap project (Prior-Magee et al. 2007). For fires that occurred before 2000 (the year of the Landsat imagery used), we interpolated small sections of the local ponderosa pine distribution from the surrounding mapped vegetation types.
Aspect. Based on research documenting aspect-driven variability in forest productivity in the region, we created a variable called aspect index (AI) for which areas with 45° aspects have the highest likelihood of pine regeneration, and lands with opposing southwest aspects (225°) have the lowest likelihood of ponderosa pine regeneration (Trimble and Weitzman 1956, Beers et al. 1966). To normalize aspect from 0 to 1 we used azimuth values from the LANDFIRE aspect data set (LANDFIRE 2013) and determined the minimum distance in degrees of each pixel from 45° (i.e., all values ≤180°; labeled az°). We then converted each azimuth to radians and took the cosine of that value. Finally, we converted the values to a scale of 0 to 1 and inverted that scale so that 0 represented the aspects of 225° and 1 represented aspects of 45°. The following equation is a summary of the steps:
where AI = aspect index, and az° = azimuth.
Topographic position index. The topographic position index (TPI) was developed using the methods outlined in Jenness et al. (2013). TPI is correlated with soil development and has been shown to relate to soil moisture availability (Parker 1982). The TPI uses a 30 m DEM and a moving window to assess the topographic position of each individual pixel and assigns a relative value to each pixel in which low numbers indicate valleys or canyons, and higher values represent steep upper slopes and exposed ridge tops. For each fire, TPI was reclassified to a range from zero to one, representing the hypothesized relative likelihood of post-fire ponderosa pine regeneration; ridge-top fires were determined to be the driest fires and assigned values near zero (i.e., least likely to have pine regeneration), and valleys were determined to be the wettest areas so they were assigned values near one (i.e., most likely to favor pine regeneration).
We normalized each topographic variable by rescaling values between zero and one for each fire, or fire group, to ensure equal weighting and to allow easier comparison of the influence of these variables on the likelihood of ponderosa pine regeneration following high-severity fire. For each topographic variable, values near 0 represented the hottest and driest sites with the lowest hypothesized likelihood of pine regeneration, while values near 1 represented the wettest or coolest sites that we assumed are more likely to regenerate to the pre-fire forest type. We did not normalize distance to seed source because it is, by nature, not comparable to topographic variables, its range does not vary among fires, and previous research has established it as an important and independent determinant of ponderosa pine regeneration (Lentile et al. 2005, Haire and McGarigal 2010, Chambers et al. 2016).
For each fire or fire group, we created raster files reflecting the normalized values for the three topographic variables to complement the distance-to-seed-source raster file used to determine sampling locations. We used these spatial data products to create maps of predicted pine regeneration for each fire or fire group.
Model Comparison and Averaging
Using an information-theoretic approach (Burnham and Anderson 2002), we developed and compared statistical models relative to combinations of the physical variables (elevation, TPI, and aspect), and distance to seed source, to explain patterns in the regeneration of ponderosa pine observed across our study landscapes following large high-severity fires. We classified regeneration, for the purposes of model development and comparison, as a plot with one or more post-fire ponderosa pine individuals greater than 15 cm tall. One tree per plot, the equivalent of 88 trees ha−1, approximates the low end of historical forest densities in the region (Moore et al. 1999).
We used logistic regression to model regeneration, employing the lme4 package (Bates et al. 2015) within the statistical program R (R Core Team 2013). We assumed a binomial distribution and incorporated fire, or fire group, as a random effect to account for natural variation among fires that was not attributable to the predictor variables described above (Crawley 2012). We developed all possible models drawing on this set of variables.
We compared statistical models using Akaike’s Information Criterion (AIC; Burnham and Anderson 2002), so we did not compute P-values. We used an intercept-only model (including random effects) and the difference in Akaike’s Information Criterion (ΔAIC) values to evaluate how well the models approximated, or fit, the data (Burnham and Anderson 2002). Those with AIC values at least 10 units less than the intercept-only model were deemed good models, while those with ΔAIC less than 2 were considered the set of best-fit models (Burnham and Anderson 2002). For models with AAIC < 2, we used model averaging to estimate regression coefficients, following the methods of Burnham and Anderson (2002). Model averaging allows for a better model fit by determining the weighted contribution of each variable, based on their importance to each of the best-fit models. We used Z-statics (variable coefficient divided by the standard error) to compare the relative importance of explanatory variables, following model averaging (Neter et al. 1996).
We created maps of predicted relative likelihood of pine regeneration, drawing on the model-averaged variable coefficients. We applied the inverse logit (ilogit) function from the Faraway package (Faraway 2014) in R to back-transform model coefficients from the binomial distribution, then used ArcGIS’s raster calculator, along with raster files reflecting normalized values for topographic variables and unmodified distance-to-seed source, to depict likelihood of regeneration. We calculated the relative likelihood of pine regeneration independently for each fire or fire group, allowing us to incorporate the appropriate random effects term for each fire or fire group using the intercept value derived from the random effects of each fire.
Results
Observed Ponderosa Pine Regeneration
We found ponderosa pine regeneration at 43 of the 175 field plots (25%; Table 1). For the 100 plots that were <150 m from a seed source, 41 plots (41%) had natural ponderosa pine regeneration. Only two of the 75 plots >150 m from a seed source (3%) contained natural ponderosa pine regeneration (Figure 2). We did not find any natural ponderosa pine regeneration >225 m from a seed source (Figure 2). The median distance of pine regeneration to a seed source was 67 m, whereas the median distance of all sampled plots (containing regeneration or not) to a seed source was 145 m.

The density of post-fire ponderosa pine regeneration as a function of distance to seed source. The median distance to a seed source for natural regeneration was 67 m, median distance for all sampled plots was 124 m.
Nurse structures were important for ponderosa pine regeneration. We found that 44% of ponderosa pine seedlings germinated under a nurse structure. Logs and large branches served as the most common nurse structure (61%), followed by shrubs and bunch grasses (27%), live trees (10%), and rocks or other inorganic material (2%).
Modeling Limits to Ponderosa Pine Regeneration
Our best model of pine regeneration was 48 AIC units lower than the intercept-only model, indicating a considerably better fit to field data. Four models had AAIC less than 2 (Table 2) and were considered the set of bestfit models. All four models included distance-to-seed source, elevation, and the random effect term (i.e., fire). Table 2 shows the subset of models that included distance to seed source.
Distance to seed source was the most important of the model-averaged predictor variables (∣Z∣ = 4.22; Table 3). As distance-to-seed source increased, likelihood of pine regeneration decreased, as reflected in the negative beta value. Elevation was the most important topographic variable (∣Z∣ = 0.95) and was positively related to pine regeneration, as indicated by a positive beta value. There was a weaker relationship with aspect (∣Z∣ = 0.86), which was negatively correlated with the likelihood of pine regeneration, indicating that, as aspect changed along the scale from 0 (dry) to 1 (wet), the likelihood of pine regeneration decreased. Topographic position index had a positive relationship with pine regeneration, but it was of low importance in the model-averaged model (∣Z∣ = 0.83). Coefficients (beta values), standard errors, and ∣Z∣ statistics are shown for all variables in Table 3.
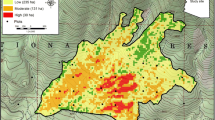
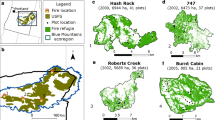
Predictive maps of relative likelihood of pine regeneration reveal areas where forest regeneration is most and least likely to occur (Figure 2). Large patches with a low likelihood of pine regeneration reflect the limited potential for seed dispersal (Figure 3) in areas distant from mature trees at the edge of the high-severity fire. Some areas near seed sources, however, show relatively low regeneration likelihood due to low elevation, which is believed to restrict ponderosa pine regeneration at the warmest ends of its local range.

Spatial outputs of our final (averaged) model for the likelihood of forest regeneration in high-severity burn patches for five of eight fires. Note the large (red) patches with very low likelihood of forest regeneration. These patches mainly occur in areas far away from any potential seed source, although locations near a seed source but at lower (drier) elevations are less likely to regenerate than those at higher (wetter) elevations. Values are categorized by quantiles. Note the differences in scale between the study areas.
Dominant Vegetation following High-Severity Fire
Grassland was the most common vegetation cover type in high-severity fire patches 7 to 17 years post fire. Grasses dominated cover on 60% of the plots (Figure 4). Across all plots, mean grass canopy cover was 44%, varying from absent to complete coverage (≥95%). Complete plot coverage was more common in areas dominated by rhizomatous grasses such as western wheatgrass (Pascopyrum smithii [Rydb.] Á. Löve). Areas dominated by bunchgrasses, mainly Arizona fescue (Festuca arizonica Vasey) and mountain muhly (Muhlenbergia montana [Nutt.] Hitchc.), tended to have more exposed ground due to the interspaces between individual grass bunches. Cheatgrass (Bromus tectorum L.) and smooth brome (B. inermis Leyss.), common invasive grasses in US Southwest forests, appeared on 26% and 11% of field plots, respectively.

Mean post-fire proportion of cover type in 2013 for our study areas that burned with high-severity fire between 1996 and 2006.
Shrubland was the second most common vegetation cover type. Shrubs dominated 36% of all plots, with tall shrub species, particularly New Mexico locust (Robinia neomexicana A. Gray) and multiple oak species (Quercus L. spp.), present on over a third (36%) of all plots. Tall shrub cover was variable, ranging from absent to completely covering sample plots in some areas. We commonly observed areas of grasses, forbs, or bare ground between the shrub canopies. At xeric locations, we commonly found oak shrub species such as Gambel oak (Quercus gambelii Nutt.), shrub live oak (Q. turbinella Greene), and wavy-leaf oak (Quercus × pauciloba Rydb. [pro sp.] [gambelii × turbinella]), and occasionally New Mexico locust.
Short shrub species dominated just 2% of all plots and averaged just 6% cover over all plots. We found members of the genera Arctostaphylos Adans. and Ceanothus L. including Arctostaphylos patula Green, A. uva-ursi (L.) Spreng., and Ceanothus fendleri A. Gray. to be the most common short shrub species, though these two genera were never co-located on the same plot. In relatively xeric areas, there was little vegetation between the sparse short shrub (Arctostaphylos spp.) canopies, and minimal fine-textured topsoil present. In contrast, short shrub communities in more mesic environments (typically dominated by Ceanothus species) tended to have a higher percentage of canopy cover, with grasses and forbs between shrub canopies.
Discussion
For our best-fit model, the most important predictor variable was distance-to-seed source. This lends support to our hypothesis that areas near a seed source are significantly more likely to regenerate than areas far away (Figure 2). Extensive areas that once supported ponderosa pine forests are now unsuitable for seedling germination and establishment. Our predictive model, employing model-averaged variable coefficients, illustrates how pine regeneration will be limited in hotter and drier locations, which typically occur at lower elevations. These results are consistent with the hypothesis that sites at relatively low elevations are generally the most water-stressed and are therefore the least likely to regenerate naturally following a high-severity fire.
There was a high level of variability in pine regeneration among the different fires, as indicated by the relatively high weight of the random-effect variable that differentiated among the fire locations included in this study. We attribute some of this variability to unmeasured site characteristics such as differences in soil types, land use history, and local precipitation patterns. Additionally, the episodic nature of ponderosa pine recruitment (Savage et al. 1996, Brown and Wu 2005), with years of very low germination followed by infrequent years of successful seedling establishment, likely influenced our results. It is possible that the recent “hotter drought” period in the US Southwest (e.g., Williams et al. 2013, 2014, 2015) has made these episodic regeneration events even rarer. Non-climatic factors, such shifts in fire regimes, vulnerability to herbivory, and competition, may further restrict reestablishment of pre-fire forest stands (Ouzts et al. 2015, Coop et al. 2016), although Owen et al. (2017) found that the presence of Gambel oak had no influence on ponderosa pine germination. Effects of shrub competition and facilitation on conifer establishment and growth following high-severity fire needs further exploration.
Even under the most optimistic projections of natural regeneration, large high-severity fire patches, such as those we studied, are likely to remain largely without forest cover, for many decades to centuries, due to the lack of nearby seed sources. The establishment of herbaceous and shrub communities post fire, combined with projected climate warming and increased forest drought stress, is likely to further constrain successful pine regeneration in the US Southwest regardless of high-severity fire patch size and distance to seed source (Puhlick et al. 2012). Concordance between our data and other projections of tree species responses to climate change (e.g., Gray and Hamann 2012, Notaro et al. 2012, Stevens-Rumann et al. 2017) suggests that climate-mediated limits to pine regeneration are already widespread across many of our study sites. Additionally, post-fire regenerated conifers are vulnerable to mortality caused by subsequent fires (Coppoletta et al. 2016), particularly young trees that germinate near a flammable nurse structure (e.g., logs), as was the case in 43% of our observations of post-fire ponderosa pine regeneration (Figure 5).

Post-fire ponderosa pine regeneration killed when nurse logs burned (Gila National Forest, New Mexico, USA). Photo by C. Haffey, 2012.
Observed changes from forest to shrubland or grassland cover types could be a short-term intermediate landscape successional stage in the regeneration of pre-fire forest conditions (Falk 2013). Alternatively, shrubland and grassland communities could become well-established, self-organizing ecosystems (Hobbs et al. 2009), indicative of long-term ecosystem type conversion to non-forest. This outcome is more likely if new ecological feedbacks develop (Johnstone et al. 2016). For example, more intense fires, occurring on moderate return intervals, could preclude re-establishment of pine forest and sustain an early-successional shrub community over the coming decades to centuries (Coop et al. 2016, Guiterman et al. 2017, Barton and Poulos 2018). The particular processes that alter ecological trajectories within a specific landscape will determine which areas, from sub-stand level (<100 ha) to near landscape scale (>500 ha), that ultimately type convert within any given patch of high-severity fire. The patches of type conversion will alter the overall landscape structure and affect many ecological processes, including future fire, hydrologic, and carbon cycles. The effects will cascade across the landscape and the connected ecological and social systems (Keane et al. 2002). As climate continues to warm, it will become even more important to understand the inter-relationships of species and ecosystems so that society can better manage or adapt to the consequences of ecological change (Walther 2010).
Management in an Uncertain Future
There is mounting evidence that ongoing climate change is driving US Southwest landscapes toward “no-analogue” futures of chronically hotter drought conditions and increasingly extreme fire activity (Westerling et al. 2006; Williams et al. 2013, 2014), resulting in rapid change and great uncertainty concerning species composition (McDowell et al. 2015) and ecological dynamics (Johnstone et al. 2016). Given these rapid and dramatic changes, land managers and policy makers are in need of tools that provide not only an assessment of changing conditions, but also a predictive capacity to help in the visualization of likely outcomes in post-fire ecosystems (e.g., Figure 3). We need to rapidly increase our knowledge about post-fire landscapes, including our understanding of the recovery trajectories that lead to long-term type conversion. Acting on our knowledge of forest and fire ecology will require a focus not only on the forests themselves, but also on the diverse ecosystems and communities that are coupled to healthy forests (e.g., Stortz et al. 2017).
The modeling approach presented here is a simple, preliminary step toward generating spatially explicit information about where post-fire type conversion is most likely to occur and most likely to be long lasting. It also identifies locations that are most likely to regenerate as ponderosa pine forests, even under changed climate conditions. These initial insights provide opportunities to test explicit predictions of post-fire dynamics and build a framework for adaptive management that anticipates ecosystem change. When implemented in broader planning discussions, these developments can enrich public discourse and identify management strategies that foster a landscape perspective and accelerate the development of landscape and community resilience to fire and incremental adaptation to changing climate.
Just as efforts to mitigate high-severity fire risk through active forest restoration (Allen et al. 2002, Stephens et al. 2013, Stevens-Rumann et al. 2013) have benefited ecosystems and the communities dependent on them, management that is informed by a clearer understanding of post-fire type vegetation dynamics can benefit people, communities, and native biota in areas where climate change precludes forest regeneration (Jackson and Hobbs 2009, Stortz et al. 2017).
Restoration efforts are desperately needed in areas where the re-establishment of forests is possible in a warmer climate. In these areas, however, it may be necessary to intervene in the successional pathway to bump systems out of a cycle of continued degradation. In upland systems, fuels management may be necessary to mitigate the risk that the treeless patches will continue to expand following subsequent fires. In other areas, targeted tree-planting and assisted migration could help create resilient forests in the future. Areas where regeneration is unlikely might be better addressed through efforts to ensure that post-fire type conversion moves toward more desirable and productive non-forest cover types. For example, in watersheds where forest is unlikely to return, managers could facilitate the post-fire establishment of dense groundcover composed of native herbaceous species, rather than shrubland with invasive annual grasses such as cheatgrass, which provide little resistance to soil erosion and contribute to recurrent fire that precludes establishment of native vegetation.
Given the high likelihood of continued increases in the size and frequency of high-severity fire throughout the US Southwest, we should anticipate permanent type conversion in large swaths of currently forested areas. These changes in cover type will result in complex landscape changes, some of which will not be reversible. Given this unprecedented change, it is critical that forest managers look beyond the forest itself and manage the post-fire landscape to increase the likelihood that the loss of forest will not lead to permanent loss of valuable ecosystem services, including watershed stability, hydrological function, the establishment of appropriate fire regimes, and the conservation of biological diversity. Increasingly, this will require managers to focus on the entire landscape, including the non-forested habitats that are an inevitable part of healthy post-fire ecosystems.
Literature Cited
Abatzoglou, J.T., and A.P. Williams. 2016. Impact of anthropogenic climate change on wildfire across western US forests. Proceedings of the National Academy of Sciences 113: 11770–11775. doi: https://doi.org/10.1073/pnas.1607171113
Abella, S.R., P.J. Fornwalt. 2015. Ten years of vegetation assembly after a North American mega fire. Global Change Biology 21: 789–802. doi: https://doi.org/10.1111/gcb.12722
Allen, C.D. 2016. Forest ecosystem reorganization underway in the southwestern US: a preview of widespread forest changes in the Anthropocene? Pages 103–122 in: V.A. Sample, R.P. Bixler, and C. Miller, editors. Forest conservation and management in the Anthropocene: adaptation of science, policy and practices. University Press of Colorado, Boulder, Colorado, USA.
Allen, CD, D.D. Breshears, and N.G. McDowell. 2015. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6: 1–55. doi: https://doi.org/10.1890/ES15-00203.1
Allen, C.D., M. Savage, D.A. Falk, K.F. Suckling, T.W. Swetnam, T. Schulke, P.B. Stacey, P. Morgan, M. Hoffman, and J.T. Klingel. 2002. Ecological restoration of Southwestern ponderosa pine ecosystems: a broad perspective. Ecological Applications 12: 1418–1433. doi: https://doi.org/10.1890/1051-0761(2002)012[1418:EROSPP]2.0.CO;2
Anderson, R.S., C.D. Allen, J.L. Toney, R.B. Jass, and A.N. Bair. 2008. Holocene vegetation and fire regimes in subalpine and mixed conifer forests, southern Rocky Mountains, USA. International Journal of Wildland Fire 17: 96. doi: https://doi.org/10.1071/WF07028
Barton, A.M. 2002. Intense wildfire in southeastern Arizona: transformation of a Madrean oakpine forest to oak woodland. Forest Ecology and Management 165: 205–212. doi: https://doi.org/10.1016/S0378-1127(01)00618-1
Barton, AM, and H.M. Poulos. 2018. Pine vs. oaks revisited: conversion of Madrean pine-oak forest to oak shrubland after high-severity wildfire in the Sky Islands of Arizona. Forest Ecology and Management 414: 28–40. doi: https://doi.org/10.1016/j.foreco.2018.02.011
Bates, D., M. Mächler, B. Bolker, and S. Walker. 2015. fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1): 1–48. doi: https://doi.org/10.18637/jss.v067.i01
Beers, T.W., P.E. Dress, and L.C. Wensel. 1966. Notes and observations: aspect transformation in site productivity research. Journal of Forestry 64: 691–692.
Bonnet, V.H., A.W. Schoettle, and W.D. Shepperd. 2005. Postfire environmental conditions influence the spatial pattern of regeneration for Pinus ponderosa. Canadian Journal of Forest Research 35: 37–47. doi: https://doi.org/10.1139/x04-157
Brown, P.M., and R. Wu. 2005. Climate and disturbance forcing of episodic tree recruitment in a Southwestern ponderosa pine landscape. Ecology 86: 3030–3038. doi: https://doi.org/10.1890/05-0034
Burnham, K.P., and D.R. Anderson. 1998. Model selection and multi-model inference: a practical information-theoretic approach, second edition. Springer, New York, New York, USA. doi: https://doi.org/10.1007/978-1-4757-2917-7
Castro, J., C.D. Allen, M. Molina-Morales, S. Marañón-Jiménez, Á. Sánchez-Miranda, and R. Zamora. 2011. Salvage logging versus the use of burnt wood as a nurse object to promote post-fire tree seedling establishment. Restoration Ecology 19: 537–544. doi: https://doi.org/10.1111/j.1526-100X.2009.00619.x
Chambers, M.E., P.J. Fornwalt, S.L. Malone, and A. Battaglia. 2016. Patterns of conifer regeneration following high severity wildfire in ponderosa pine-dominated forests of the Colorado Front Range. Forest Ecology and Management 378: 57–67. doi: https://doi.org/10.1016/j.foreco.2016.07.001
Collins, B.M., and G.B. Roller. 2013. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landscape Ecology 28: 1801–1813. doi: https://doi.org/10.1007/s10980-013-9923-8
Collins, B.M., J.T. Stevens, J.D. Miller, S.L. Stephens, P.M. Brown, and M.P. North. 2017. Alternative characterization of forest fire regimes: incorporating spatial patterns. Landscape Ecology 32: 1543–1552. doi: https://doi.org/10.1007/s10980-017-0528-5
Coop, J.D., S.A. Parks, S.R. McClernan, and L.M. Holsinger. 2016. Influences of prior wildfires on vegetation response to subsequent fire in a reburned Southwestern landscape. Ecological Applications 26: 346–354. doi: https://doi.org/10.1890/15-0775
Coppoletta, M., K.E. Merriam, and B.M. Collins. 2016. Post-fire vegetation and fuel development influences fire severity patterns in reburns. Ecological Applications 26: 686–699. doi: https://doi.org/10.2737/RDS-2015-0039
Covington, W.W. 2000. Helping Western forests heal. Nature 408: 135–136. doi: https://doi.org/10.1038/35041641
Covington, W.W., and M.M. Moore. 1994. Southwestern ponderosa forest structure: changes since Euro-American settlement. Journal of Forestry 92: 39–47.
Crotteau, J.S., J.M. Varner, and M.W. Ritchie. 2013. Post-fire regeneration across a fire severity gradient in the southern Cascades. Forest Ecology and Management 287: 103–112. doi: https://doi.org/10.1016/j.foreco.2012.09.022
Dennison, P.E., S.C. Brewer, J.D. Arnold, and M.A. Moritz. 2014. Large wildfire trends in the western United States, 1984–2011. Geophysical Research Letters 41: 2928–2933. doi: https://doi.org/10.1002/2014GL059576
Dodson, E.K., and H.T. Root. 2013. Conifer regeneration following stand-replacing wildfire varies along an elevation gradient in a ponderosa pine forest, Oregon, USA. Forest Ecology and Management 302: 163–170. doi: https://doi.org/10.1016/j.foreco.2013.03.050
Eidenshink, J., B. Schwind, K. Brewer, Z. Zhu, B. Quayle, and S. Howard. 2007. A project for monitoring trends in burn severity. Fire Ecology 3: 3–21. doi: https://doi.org/10.4996/fireecology.0301003
Falk, D.A. 2013. Are Madrean ecosystems approaching tipping points? Anticipating interactions of landscape disturbance and climate change. Pages 40–47 in: G.J. Gottfried, P.F. Ffolliott, B.S. Gebow, L.G. Eskew, and L.C. Collins, editors. Merging science and management in a rapidly changing world: biodiversity and management of the Madrean Archipelago III. USDA Forest Service, Tucson, Arizona, USA.
Falk, D.A., E.K. Heyerdahl, P.M. Brown, C. Farris, P.Z. Fulé, D. McKenzie, T.W. Swetnam, H. Taylor, and M.L. Van Horne. 2011. Multi-scale controls of historical forest-fire regimes: new insights from fire-scar networks. Frontiers in Ecology and the Environment 9: 446–454. doi: https://doi.org/10.1890/100052
Faraway, J.J. 2014. Linear models with R. CRC Press, Boca Raton, Florida, USA.
Feddema, J.J., J.N. Mast, and M. Savage. 2013. Modeling high-severity fire, drought and climate change impacts on ponderosa pine regeneration. Ecological Modelling 253: 56–69. doi: https://doi.org/10.1016/j.ecolmodel.2012.12.029
Flathers, K.N., T.E. Kolb, J.B. Bradford, K.M. Waring, and W.K. Moser. 2016. Long-term thinning alters ponderosa pine reproduction in northern Arizona. Forest Ecology and Management 374: 154–165. doi: https://doi.org/10.1016/j.foreco.2016.04.053
Fornwalt, P.J., L.S. Huckaby, S.K. Alton, M.R. Kaufmann, P.M. Brown, and A.S. Cheng. 2016. Did the 2002 Hayman Fire, Colorado, USA, burn with uncharacteristic severity? Fire Ecology 12: 117–132. doi: https://doi.org/10.4996/fireecology.1203117
Foxx, T.S., L.A. Hansen, R. Oertel, C. Haffey, and K. Beeley. 2013. The La Mesa Fire: studies and observations from 1975 through 2012. Los Alamos National Laboratory, Los Alamos, New Mexico, USA. doi: https://doi.org/10.2172/1084501
Fulé, P.Z., T.W. Swetnam, P.M. Brown, D. A. Falk, D. L. Peterson, C. D. Allen, G. H. Aplet, M. A. Battaglia, D. Binkley, and C. Farris. 2013. Unsupported inferences of high-severity fire in historical dry forests of the western United States: response to Williams and Baker. Global Ecology and Biogeography 23: 825–830. doi: https://doi.org/10.1111/geb.12136
Gracia, M., J. Retana, and P. Roig. 2002. Mid-term successional patterns after fire of mixed pine-oak forests in NE Spain. Acta Oecologica 23: 405–411. doi: https://doi.org/10.1016/S1146-609X(02)01169-4
Gray, L.K., and A. Hamann. 2012. Tracking suitable habitat for tree populations under climate change in western North America. Climatic Change 117: 289–303. doi: https://doi.org/10.1007/s10584-012-0548-8
Grissino-Mayer, H.D., and T.W. Swetnam. 2000. Century-scale climate forcing of fire regimes in the American Southwest. The Holocene 10: 213–220. doi: https://doi.org/10.1191/095968300668451235
Guiterman, C.H., E.Q. Margolis, C.D. Allen, D.A. Falk, and T.W. Swetnam. 2017. Long-term persistence and fire resilience of oak shrubfields in dry conifer forests of northern New Mexico. Ecosystems 20: 1–17. doi: https://doi.org/10.1007/s10021-017-0192-2
Haire, S., J. Coop, and C. Miller. 2017. Characterizing spatial neighborhoods of refugia following large fires in northern New Mexico USA. Land 6: 19. doi: https://doi.org/10.3390/land6010019
Haire, S.L., and K. McGarigal. 2010. Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landscape Ecology 25: 1055–1069. doi: https://doi.org/10.1007/s10980-010-9480-3
Hobbs, R.J., E. Higgs, and J.A. Harris. 2009. Novel ecosystems: implications for conservation and restoration. Trends in Ecology and Evolution 24(11): 599–605. doi: https://doi.org/10.1016/j.tree.2009.05.012
Iniguez, J.M., T.W. Swetnam, and C.H. Baisan. 2009. Spatially and temporally variable fire regime on Rincon Peak, Arizona, USA. Fire Ecology 5: 3–21. doi: https://doi.org/10.4996/fireecology.0501003
Jackson, S.T., and R.J. Hobbs. 2009. Ecological restoration in the light of ecological history. Science 325: 567–569. doi: https://doi.org/10.1126/science.1172977
Jenness, J., B. Brost, and P. Beier. 2013. Land facet CorridorDesigner. <https://doi.org/corridordesign.org/downloads>. Accessed 4 April 2018.
Johnstone, J.F., C.D. Allen, J.F. Franklin, L.E. Frelich, B.J. Harvey, P.E. Higuera, M.C. Mack, R.K. Meentemeyer, M.R. Metz, G.L. Perry, T. Schoennagel, and M.G. Turner. 2016. Changing disturbance regimes, ecological memory, and forest resilience. Frontiers in Ecology and the Environment 14: 369–378. doi: https://doi.org/10.1002/fee.1311
Keane, R.E., K.C. Ryan, T.T. Veblen, C.D. Allen, J. Logan, and B. Hawkes. 2002. Cascading effects of fire exclusion in Rocky Mountain ecosystems: a literature review. USFS General Technical Report RMRS-GTR-91, US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fort Collins, Colorado, USA. doi: https://doi.org/10.2737/RMRS-GTR-91
Keyser, T.L., L.B. Lentile, F.W. Smith, and W.D. Shepperd. 2008. Changes in forest structure after a large, mixed-severity wildfire in ponderosa pine forests of the Black Hills, South Dakota, USA. Forest Science 54: 328–338.
LANDFIRE. 2013. LANDFIRE Digital Elevation Model, US Department of the Interior, Geological Survey. <https://doi.org/landfire.cr.usgs.gov/viewer/>. Accessed 1 January 2013.
Lentile, L.B., P. Morgan, A.T. Hudak, M.J. Bobbitt, S.A. Lewis, A.M.S. Smith, and P.R. Robichaud. 2007. Post-fire burn severity and vegetation response following eight large wildfires across the western United States. Fire Ecology 3(1): 91–108. doi: https://doi.org/10.4996/fireecology.0301091
Lentile, L.B., F.W. Smith, and W.D. Shepperd. 2005. Patch structure, fire-scar formation, and tree regeneration in a large mixed-severity fire in the South Dakota Black Hills, USA. Can J For Res 35: 2875–2885. doi: https://doi.org/10.1139/x05-205
Marañón-Jiménez, S., J. Castro, J.I. Querejeta, E. Fernández-Ondoño, and C.D. Allen. 2013. Post-fire wood management alters water stress, growth, and performance of pine regeneration in a Mediterranean ecosystem. Forest Ecology and Management 308: 231–239. doi: https://doi.org/10.1016/j.foreco.2013.07.009
Margolis, E.Q., and J. Balmat. 2009. Fire history and fire-climate relationships along a fire regime gradient in the Santa Fe Municipal Watershed, NM, USA. Forest Ecology and Management 258: 2416–2430. doi: https://doi.org/10.1016/j.foreco.2009.08.019
Margolis, E.Q., and S.B. Malevich. 2016. Historical dominance of low-severity fire in dry and wet mixed-conifer forest habitats of the endangered terrestrial Jemez Mountains salamander (Plethodon neomexicanus). Forest Ecology and Management 375: 12–26. doi: https://doi.org/10.1016/j.foreco.2016.05.011
Margolis, E.Q., and T.W. Swetnam. 2013. Historical fire-climate relationships of upper elevation fire regimes in the south-western United States. International Journal of Wildland Fire 22: 588. doi: https://doi.org/10.1071/WF12064
McDowell, N.G., A.P. Williams, C. Xu, W.T. Pockman, L.T. Dickman, S. Sevanto, R. Pangle, J.-M. Limousin, J. Plaut, D.S. Mackay, J. Ogee, J.C. Domec, C.D. Allen, R.A. Fisher, X. Jiang, J.D. Muss, D.D. Breshears, S.A. Rauscher, and C. Koven. 2015. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nature Climate Change 6: 295–300. doi: https://doi.org/10.1038/nclimate2873
McDowell, N., W.T. Pockman, C.D. Allen, D.D. Breshears, N. Cobb, T. Kolb, J. Plaut, J. Sperry, A. West, D.G. Williams, and E.A. Yepez. 2008. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytologist 178: 719–739. doi: https://doi.org/10.1111/j.1469-8137.2008.02436.x
Meyer, C.L., T.D. Sisk, and W.W. Covington. 2001. Microclimatic changes induced by ecological restoration of ponderosa pine forests in northern Arizona. Restoration Ecology 9: 443–452. doi: https://doi.org/10.1046/j.1526-100X.2001.94013.x
Miller, J.D., E.E. Knapp, C.H. Key, C.N. Skinner, C.J. Isbell, R.M. Creasy, and J.W. Sherlock. 2009. Calibration and validation of the relative differenced Normalized Burn Ratio (RdNBR) to three measures of fire severity in the Sierra Nevada and Klamath Mountains, California, USA. Remote Sensing of Environment 113: 645–656. doi: https://doi.org/10.1016/j.rse.2008.11.009
Miller, J.D., and A.E. Thode. 2007. Quantifying burn severity in a heterogeneous landscape with a relative version of the delta Normalized Burn Ratio (dNBR). Remote Sensing of Environment 109: 66–80. doi: https://doi.org/10.1016/j.rse.2006.12.006
Moore, M.M., W.W. Covington, and P.Z. Fulé. 1999. Reference conditions and ecological restoration: a Southwestern ponderosa pine perspective. Ecological Applications 9: 1266–1277. doi: https://doi.org/10.1890/1051-0761(1999)009[1266:RCAERA]2.0.CO;2
Moser, B., C. Temperli, G. Schneiter, and T. Wohlgemuth. 2010. Potential shift in tree species composition after interaction of fire and drought in the Central Alps. European Journal of Forest Research 129: 625–633. doi: https://doi.org/10.1007/s10342-010-0363-6
Neter, J., M.H. Kutner, C.J. Nachtsheim, and W. Wasserman. 1996. Applied linear statistical models. Irwin, Chicago, Illinois, USA.
Notaro, M., A. Mauss, and J.W. Williams. 2012. Projected vegetation changes for the American Southwest: combined dynamic modeling and bioclimatic-envelope approach. Ecological Applications 22: 1365–1388. doi: https://doi.org/10.1890/11-1269.1
Ouzts, J., T. Kolb, D. Huffman, and A. Sánchez Meador. 2015. Post-fire ponderosa pine regeneration with and without planting in Arizona and New Mexico. Forest Ecology and Management 354: 281–290. doi: https://doi.org/10.1016/j.foreco.2015.06.001
Owen, S.M., C.H. Sieg, A.J. Sánchez Meador, P.Z. Fulé, J.M. Iniguez, L.S. Baggett, P.J. Fornwalt, and M.A. Battaglia. 2017. Spatial patterns of ponderosa pine regeneration in high-severity burn patches. Forest Ecology and Management 405: 134–149. doi: https://doi.org/10.1016/j.foreco.2017.09.005
Parker, A.J. 1982. The topographic relative moisture index: an approach to soil-moisture assessment in mountain terrain. Physical Geography 3: 160–168.
Prior-Magee, J.S., K.G. Boykin, D.F. Bradford, W.G. Kepner, K.A. Thomas, and B.C. Thompson. 2007. The Southwest Regional Gap Analysis Project. <https://doi.org/swregap.nmsu.edu/default.htm>. Accessed 4 April 2018.
Puhlick, J.J., D.C. Laughlin, and M.M. Moore. 2012. Factors influencing ponderosa pine regeneration in the southwestern USA. Forest Ecology and Management 264: 10–19. doi: https://doi.org/10.1016/j.foreco.2011.10.002
R Core Team. 2013. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
Reynolds, R.T., A.J.S Meador, J.A. Youtz, T. Nicolet, M.S. Matonis, P.L. Jackson, D.G. Delorenzo, and A.D. Graves. 2013. Restoring composition and structure in Southwestern frequent-fire forests: a science-based framework for improving ecosystem resiliency. USDA Forest Service, General Technical Report RMRS-GTR-310, Rocky Mountain Research Station, Fort Collins, Colorado, USA.
Roccaforte, J.P., P.Z. Fulé, W.W. Chancellor, and D.C. Laughlin. 2012. Woody debris and tree regeneration dynamics following severe wildfires in Arizona ponderosa pine forests. Canadian Journal of Forest Research 42: 593–604. doi: https://doi.org/10.1139/x2012-010
Rother, M.T., and T.T. Veblen. 2016. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 7(2): e01594. doi: https://doi.org/10.1002/ecs2.1594
Rother, M.T., T.T. Veblen, and L.G. Furman. 2015. A field experiment informs expected patterns of conifer regeneration after disturbance under changing climate conditions. Canadian Journal of Forest Research 45(11): 1607–1616. doi: https://doi.org/10.1139/cjfr-2015-0033
Savage, M., P.M. Brown, and J. Feddema. 1996. The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience 3: 310–318. doi: https://doi.org/10.1080/11956860.1996.11682348
Savage, M., and J.N. Mast. 2005. How resilient are Southwestern ponderosa pine forests after crown fires? Canadian Journal of Forest Research 35: 967–977. doi: https://doi.org/10.1139/x05-028
Savage, M., J.N. Mast, and J.J. Feddema. 2013. Double whammy: high-severity fire and drought in ponderosa pine forests of the Southwest. Canadian Journal of Forest Research 43: 570–583. doi: https://doi.org/10.1139/cjfr-2012-0404
Sisk, T.D., M. Savage, D.A. Falk, C.D. Allen, E. Muldavin, and P. McCarthy. 2005. A landscape perspective for forest restoration. Journal of Forestry 103(6): 319–320.
Stephens, S.L., J.K. Agee, P.Z. Fulé, M.P. North, W.H. Romme, T.W. Swetnam, and M.G. Turner. 2013. Managing forests and fire in changing climates. Science 342: 41–42. doi: https://doi.org/10.1126/science.1240294
Stevens-Rumann, C.S., K.B. Kemp, P.E. Higuera, B.J. Harvey, M.T. Rother, D.C. Donato, P. Morgan, and T.T. Veblen. 2017. Evidence for declining forest resilience to wildfires under climate change. Ecology Letters 21(2): 243–522. doi: https://doi.org/10.1111/ele.12889
Stevens-Rumann, C., K. Shive, P.Z. Fulé, and C.H. Sieg. 2013. Pre-wildfire fuel reduction treatments result in more resilient forest structure a decade after wildfire. International Journal of Wildland Fire 22(8): 1108–1117. doi: https://doi.org/10.1071/WF12216
Stortz, S., C.M. Haffey, and C. Kimball. 2017. East Jemez Landscape Futures needs assessment and recommendations: identifying cross-boundary opportunities for management in altered landscapes. <https://doi.org/docs.wixstatic.com/ugd/dcbcff_eecc0efa6a244c839b67a4a49ea31095.pdf>. Accessed 29 March 2018.
Swetnam, T.W., and C.H. Baisan. 1996. Historical fire regime patterns in the southwestern United States since AD 1700. Pages 11–32 in: C.D. Allen, editor. Fire effects in Southwestern forest: proceedings of the second La Mesa Fire Symposium. USFS General Technical Report RM-GTR-286, USDA Forest Service, Rocky Mountain Research Station, Los Alamos, New Mexico, USA.
Swetnam, T.W., and J.L. Betancourt. 1990. Fire-Southern Oscillation relations in the southwestern United States. Science 249: 1017–1020. doi: https://doi.org/10.1126/science.249.4972.1017
Swetnam, T.W., and J.L. Betancourt. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate 11: 3128–3147. doi: https://doi.org/10.1175/1520-0442(1998)011<3128:MDAERT>2.0.CO;2
Touchan, R., C.D. Allen, and T.W. Swetnam. 1996. Fire history and climatic patterns in ponderosa pine and mixed-conifer forests of the Jemez Mountains, northern New Mexico. Pages 33–46 in: C.D. Allen, editor. Fire effects in Southwestern forest: proceedings of the second La Mesa Fire Symposium. USFS General Technical Report RM-GTR-286, USDA Forest Service, Rocky Mountain Research Station, Los Alamos, New Mexico, USA.
Trimble, G.R., and S. Weitzman. 1956. Site index studies of upland oaks in the northern Appalachians. Forest Science 2: 162–173.
USFSA [US Farm Service Agency]. 2013. Southwestern Region land & resource management, geospatial data portal. <https://doi.org/www.fs.usda.gov/main/r3/landmanagement/gis>. Accessed 1 January 2013.
Vilà-Cabrera, A., A. Rodrigo, J. Martínez-Vilalta, and J. Retana. 2012. Lack of regeneration and climatic vulnerability to fire of Scots pine may induce vegetation shifts at the southern edge of its distribution. Journal of Biogeography 39: 488–496. doi: https://doi.org/10.1111/j.1365-2699.2011.02615.x
Walther, G.-R. 2010. Community and ecosystem responses to recent climate change. Philosophical Transactions of the Royal Society of London, Series B, Biological Sciences 365: 2019–2024. doi: https://doi.org/10.1098/rstb.2010.0021
Waring, K.M., K.J. Hansen, and W. Flatley. 2016. Evaluating prescribed fire effectiveness using permanent monitoring plot data: a case study. Fire Ecology 12: 1–25. doi: https://doi.org/10.4996/fireecology.1203002
Westerling, A.L., H.G. Hidalgo, D.R. Cayan, and T.W. Swetnam. 2006. Warming and earlier spring increase western US forest wildfire activity. Science 313: 940–943. doi: https://doi.org/10.1126/science.1128834
Williams, A.P., C.D. Allen, A.K. Macalady, D. Griffin, C.A. Woodhouse, D.M. Meko, T.W. Swetnam, S.A. Rauscher, R. Seager, H.D. Grissino-Mayer, J.S. Dean, E.R. Cook, C. Gangodagamage, M. Cai, and N.G. McDowell. 2013. Temperature as a potent driver of regional forest drought stress and tree mortality. Nature Climate Change 3: 8–13. doi: https://doi.org/10.1038/nclimate1693
Williams, A.P., R. Seager, J.T. Abatzoglou, B.I. Cook, J.E. Smerdon, and E.R. Cook. 2015. Contribution of anthropogenic warming to California drought during 2012–2014. Geophysical Research Letters 42: 6819–6828. doi: https://doi.org/10.1002/2015GL064924
Williams, A.P., R. Seager, M. Berkelhammer, A.K. Macalady, M.A. Crimmins, T.W. Swetnam, A. T. Trugman, N. Buenning, N. Hryniw, N.G. McDowell, D. Noone, C.I. Mora, and T. Rahn. 2014. Causes and implications of extreme atmospheric moisture demand during the record-breaking 2011 wildfire season in the southwestern United States. Journal of Applied Meteorology and Climatology 53: 2671–2684. doi: https://doi.org/10.1175/JAMC-D-14-0053.1
Acknowledgements
Support for this project came for the Lab of Landscape Ecology and Conservation Biology (LLECB), the Olajos-Goslow Endowment, and the Landscape Conservation Initiative at Northern Arizona University, as well as the US Geological Survey’s Western Mountain Initiative. We would like to thank R. Nanni for her fieldwork assistance, B. Dickson for support developing statistical models, LLECB members for great discussion, and R. Meier for cartographic support. Comments from W. Palen and two anonymous reviewers greatly improved the manuscript. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Haffey, C., Sisk, T.D., Allen, C.D. et al. Limits to Ponderosa Pine Regeneration following Large High-Severity Forest Fires in the United States Southwest. fire ecol 14, 143–163 (2018). https://doi.org/10.4996/fireecology.140114316
Published:
Issue Date:
DOI: https://doi.org/10.4996/fireecology.140114316