
Abstract
In an item-method directed forgetting task, attentional resources are withdrawn from forget item processing (e.g., Taylor & Fawcett in Attention, Perception, & Psychophysics, 73, 1790–1814, 2011). Taylor and Hamm (Attention, Perception, & Psychophysics, 78, 168–186, 2016) demonstrated that there is no corresponding increase in the proclivity for exogenous attention to be captured following a forget instruction. This means either that the attentional resources withdrawn from the forget item are reallocated immediately (and therefore not especially vulnerable to capture) or that it is not exogenous attention that is withdrawn. Given that endogenous attention is distinct from exogenous attention, we therefore extended the Taylor and Hamm study by using endogenous orienting rather than exogenous orienting. Words appeared individually in a peripheral location (Exp. 1) or in a central location (Exp. 2), followed by an instruction to either remember or forget. After a short (50-ms) or long (250-ms) interstimulus interval (ISI), a central cue (80% accurate) directed participants to allocate their attention to the left or right. This was followed by a discrimination target that appeared at a 1,000-ms cue–target stimulus onset asynchrony. A subsequent yes–no recognition test assessed memory for all study items. In both experiments, we observed better recognition of remember words than forget words—a directed forgetting effect. We also found a cueing effect, revealed as faster reaction times to discriminate cued targets than to discriminate uncued targets. There was not, however, an effect of memory instruction (and/or instruction–cue ISI) on the magnitude of this cueing effect. Thus, neither exogenous attention nor endogenous attention remains in an unengaged state following an instruction to forget.
Similar content being viewed by others
Optimal memory functioning relies both on the ability to remember relevant information and on the ability to intentionally forget unwanted or irrelevant information. Forgetting has neural processing benefits that aid us in the context of our limited-capacity cognitive resources (Kuhl, Dudukovic, Kahn, & Wagner, 2007; Williams, Hong, Kang, Carlisle, & Woodman, 2013). Forgetting reduces the demands placed on our cognitive control system (Kuhl et al., 2007) and frees up space in our working memory stores (Souza, Rerko, & Oberauer, 2014; Williams et al., 2013), leaving maintenance and control mechanisms available to be focused on goal-relevant processing and storage (Kuhl et al., 2007; Souza et al., 2014; Williams et al., 2013).
Both the laboratory paradigms used to elicit forgetting and the mechanisms posited to explain it differ depending on whether forgetting occurs primarily at encoding or at retrieval (see MacLeod, 1998, for a review). The present study is concerned with forgetting that occurs at encoding. Intentional forgetting at encoding is studied using an item-method directed forgetting paradigm, which presents participants with study items one at a time. Following each item, participants are instructed, with equal probability, to remember or to forget the item. Because the memory instruction appears after the word has disappeared, participants must attend to each item and use maintenance rehearsal to refresh its representation in working memory (e.g., Gardiner, Gawlik, & Richardson-Klavehn, 1994; Hsieh, Hung, Tzeng, Lee, & Cheng, 2009; Paz-Caballero, Menor, & Jiménez, 2004; van Hooff & Ford, 2011) until the instruction is provided. When the instruction is to remember, participants engage in elaborative rehearsal to commit the item to long-term memory (Gardiner et al., 1994; Hsieh et al., 2009). In contrast, when the instruction is to forget, participants employ cognitive control (Fawcett & Taylor, 2008; Ludowig et al., 2010) to actively withdraw processing resources from the forget item representation (Fawcett & Taylor, 2010; Thompson, Hamm, & Taylor, 2014). This attentional withdrawal is achieved through frontal and parietal control mechanisms (Gallant & Dyson, 2016; Rizio & Dennis, 2013; Wylie, Foxe, & Taylor, 2008; Yang, Lei, & Anderson, 2016) that limit further processing and encoding of the unwanted item (Hourihan & Taylor, 2006; Lee, Lee, & Tsai, 2007; although see Bancroft, Hockley, & Farquhar, 2013). Due to this selective rehearsal of remember items over forget items, forget items are less likely to be encoded, and therefore also less likely to be remembered later (see, e.g., Fawcett, Lawrence, & Taylor, 2016).Footnote 1 This is evidenced by a directed forgetting effect, which is operationalized as better recognition or recall of remember items than forget items (see MacLeod, 1998, for a review).
Given that the directed forgetting effect depends, in part, on the removal of limited-capacity resources from unwanted forget item processing, a key question is whether this withdrawal has longer-term downstream consequences for subsequent information processing. This would be the case, for example, if the attentional resources freed from unwanted forget item processing remained available for subsequent exogenous capture by sudden changes in the environment and/or for endogenous allocation to other goal-directed activities. As we describe below, the question of whether a forget instruction frees exogenous attention for capture has already been answered in the negative (Taylor & Hamm, 2016). However, as we also describe, exogenous and endogenous attention are not simply different ways of orienting the same pool of attentional resources, but rather are two distinct attentional systems. The fact that exogenous attention is not more vulnerable to capture following a forget instruction implies either that the withdrawn resources are immediately reallocated (and, hence, no more vulnerable to capture than on remember trials) or that the withdrawal of attention involves changes in the endogenous attentional system rather than the exogenous attentional system. We put the latter possibility to the test in the present investigation.
Exogenous versus endogenous attentional systems
Exogenous orienting is initiated when attention is captured by salient sensory events in the environment (Hopfinger & West, 2006), such as abrupt onsets in the visual periphery (e.g., Jonides, 1981; Jonides & Yantis, 1988; Yantis & Jonides, 1984). This bottom-up capture of attention is relatively automatic (e.g., Hopfinger & West, 2006; McCormick, 1997) and fast (Cheal & Lyon, 1991; Müller & Rabbitt, 1989). Exogenous attention leads to stimulus enhancement and a reduction of external noise (Lu & Dosher, 2000) and influences object-based perceptual processing (see Chica, Bartolomeo, & Lupiáñez, 2013). In contrast, endogenous orienting is intentional and effortful; it is a voluntary response to an informative instruction (Hopfinger & West, 2006). Unlike exogenous attention, endogenous attention leads to stimulus enhancement — but not to external noise reduction (Lu & Dosher, 2000) — and it influences spatial-based rather than object-based perceptual processing (see Chica et al., 2013). As compared to exogenous attention, endogenous attention is easier to suppress, less resistant to interruption, and more affected by cognitive load (Jonides, 1981; Müller & Rabbitt, 1989).
Whereas early models conceptualized endogenous and exogenous attention as two modes for orienting a single attentional system (e.g., Jonides, 1981; Posner, 1980), more recent evidence compels the conclusion that exogenous and endogenous attention are two independent but interacting attentional systems (see Chica et al., 2013, for a comprehensive review), characterized by separate limited-capacity pools of attentional resources (e.g., Berger, Henik, & Rafal, 2005). This underscores the importance of testing the downstream effects of memory instructions on both systems—the endogenous attentional system as well as the exogenous attentional system.
Downstream consequences of memory instructions
With respect to the exogenous attentional system, Taylor and Hamm (2016) tested the prediction that forget instructions (relative to remember instructions) make exogenous resources more available for capture by abrupt onsets in the visual periphery. Their rationale for doing so was based on an argument made by Taylor and Fawcett (2011), that a forget instruction initiates a withdrawal of exogenous, rather than endogenous, attention. Taylor and Fawcett posited this explanation to account for larger inhibition of return (IOR; Posner & Cohen, 1984) effects following instructions to forget than following instructions to remember—but only when the instructions were presented auditorily. When the instructions consisted of a visual onset at center, the IOR difference between remember and forget trials was eliminated. Reasoning that IOR was equivalently “unmasked” (cf. Danziger & Kingstone, 1999), Taylor and Fawcett suggested that the capture of exogenous attention by the central visual onset subverted the differential withdrawal of exogenous attention that would otherwise occur in response to a forget instruction as compared to a remember instruction.
Taylor and Fawcett’s (2011) argument for a differential withdrawal of exogenous attention from forget and remember items was admittedly post hoc. Nevertheless, it suggested prioritizing the exogenous attentional system in the search for downstream consequences of a forget instruction—hence, Taylor and Hamm’s (2016) investigation. In their first experiment, Taylor and Hamm presented study words at center, and in a second and third experiment, they presented words to the left and right. In all three experiments, a memory instruction followed immediately after the disappearance of the word. Not knowing how long it might take for a memory instruction to be enacted or for it to influence subsequent attentional capture by a cue, they imposed both a relatively short (50 ms) and a relatively long (250 ms) instruction–cue interstimulus interval (ISI). After this interval had elapsed, they presented a spatially nonpredictive peripheral onset cue that was intended to capture exogenous attention. This capture of attention was confirmed by measuring reaction times (RTs) to a localization (Exps. 1 and 2) or discrimination (Exp. 3) target that appeared at a fixed 100-ms cue–target stimulus onset asynchrony (SOA) in either the cued or the uncued location. In this way, the temporal relation between the cue and target onset remained constant; all that varied was whether the cue was preceded at a relatively short or a relatively long interval by a remember or a forget instruction. The goal was to determine whether the withdrawal of processing resources from a forget instruction might free exogenous resources for immediate capture by an onset cue in the visual periphery.
Perceptual processing of a target is enhanced when it appears at an attended rather than an unattended location (e.g., Hopfinger & West, 2006; Liu, Pestilli, & Carrasco, 2005). Consequently, if a cue captures attention at its location, targets that appear subsequently at this cued location should elicit faster responses than targets that appear elsewhere, at an uncued location. This uncued–cued RT difference is referred to as a cueing effect. The larger the cueing effect, the more effective the cues are at orienting attention. Accordingly, Taylor and Hamm (2016) posited that if exogenous attention were more vulnerable to capture following forget instructions than following remember instructions, the cueing effect would be larger on forget trials than on remember trials. But this was not what they found. They observed a reliable cueing effect, demonstrating the efficacy of the onset cues in capturing exogenous attention. But this cueing effect was not larger following forget instructions than following remember instructions (at either instruction–cue ISI). Taylor and Hamm concluded that memory intentions formed at encoding produce no discernable downstream consequences for the exogenous attentional system.
Taylor and Hamm’s (2016) failure to find a larger exogenous cueing effect following forget instructions than following remember instructions means one of two things: Either the attentional resources withdrawn from the forget item are reallocated immediately (and therefore not more vulnerable to capture than on remember trials), or it is not exogenous attentional resources that are withdrawn following a forget instruction. Indeed, it is not at all clear why implementing top-down control to limit unwanted memory encoding would engage a reflexive, rather than a controlled, withdrawal of attentional resources—despite Taylor and Fawcett’s (2011) claims to the contrary. In any case, the prediction derived from Taylor and Fawcett’s argument—that endogenous attention should not be influenced by memory instructions—is a falsifiable claim that demands an empirical test (e.g., Popper, 1968). This is especially true given work from Lee’s laboratory (see below; Lee, 2012; Lee & Lee, 2011), which points to a potential role for endogenous attention in item-method directed forgetting.
Lee and colleagues have demonstrated that memory instructions are most effective when the cognitive system is taxed by a high load (Lee, 2012) or by the need to divide attention (Lee & Lee, 2011). At first blush, it might seem counterintuitive that participants are most likely to implement a forget instruction when their cognitive system is taxed—especially when one considers that implementing a forget instruction is itself effortful (e.g., Cheng, Liu, Lee, Hung, & Tzeng, 2012; Fawcett & Taylor, 2008). However, expending the effort to forget helps minimize interference from these unwanted items. More importantly for present purposes, the fact that implementing a memory instruction is under strategic guidance and can be influenced by perceived task difficulty speaks to a potential role of the endogenous attentional system. This is because strategic control is within the purview of endogenous attention and not exogenous attention. This underscores the importance of extending the Taylor and Hamm (2016) study to determine whether memory instruction interacts with endogenous cueing, even though it did not interact with exogenous cueing.
The present study
Modeling our methods closely on those of Taylor and Hamm (2016), we presented participants with study words, one at a time. In Experiment 1, these words appeared to the left or right of fixation; in Experiment 2, they all appeared at center. Following each word, we presented one of two tones, with equal probability, as instructions to either remember or forget the preceding word. This was followed by a 50-ms or a 250-ms instruction–cue ISI. These ISIs were of the same durations used by Taylor and Hamm and were meant to hedge our bets with respect to the possible time course over which endogenous attentional resources might become differentially available following interpretation and implementation of the forget and remember instructions. Following the delay, a cue intended to encourage the allocation of endogenous attention appeared at center. This cue directed participants to orient their attention to the left (“<<”) or to the right (“>>”).Footnote 2
To give participants incentive to orient their attention voluntarily, this cue was 80% predictive of the upcoming target location. Like Taylor and Hamm (2016), we did not restrict eye movements, but instead allowed for the possibility of joint effects of endogenous covert and endogenous overt attention. We confirmed the effectiveness of these cues at eliciting an attentional shift by measuring RTs to discriminate the up–down orientation of targets (“/\” or “\/”) that appeared after a fixed 1,000-ms cue–target SOA (i.e., 1,000 ms separated the onset of the cue and the onset of the target); this SOA was selected to ensure that the cues had more than sufficient time to maximally engage available attentional resources (e.g., Cheal & Lyon, 1991; Müller & Rabbitt, 1989). If endogenous attention is more available to be allocated following forget instructions than following remember instructions, this should be revealed as a larger cueing effect when the cue is preceded — whether at a relatively short and/or at a relatively long ISI — by a forget instruction, rather than by a remember instruction. In other words, if memory instructions have downstream consequences for endogenous orienting, there should be evidence of a two-way interaction between memory instruction and cueing effects, with the possibility of a three-way interaction with instruction–cue ISI.
Experiment 1
In Experiment 1, study words appeared with equal probability to the left or right of fixation. The goal of this experiment was to determine whether there is a larger endogenous cueing effect on forget trials than on remember trials. The methods closely resembled those of Taylor and Hamm (2016), except that we used cues that were intended to engage the endogenous attentional system, rather than the exogenous attentional system. To this end, we employed central cues that were 80% predictive of the upcoming target location and displayed these cues for a longer duration than Taylor and Hamm had used (500 ms rather than 100 ms), to provide time for participants to interpret their meaning (e.g., Müller, 1986, reported that central arrows presented for 300 ms were subject to directional confusions that did not occur for peripheral cues of much shorter duration). We also increased the fixed cue–target SOA (1,000 ms instead of 100 ms) to accommodate the slower time course of endogenous orienting compared to exogenous orienting (Cheal & Lyon, 1991; Müller & Rabbitt, 1989).
The magnitude of endogenous cueing effects varies directly with the ratio of cued:uncued targets (Riggio & Kirsner, 1997). The necessity of having more cued than uncued targets precluded using a target localization task: Participants would be able to maintain high target localization accuracy simply by matching their responses to the validity ratio, without necessarily having to shift their attention to the cued location. We therefore elected not to use the localization task that Taylor and Hamm (2016) employed in their Experiments 1 and 2; we instead used the target discrimination task that they employed in their Experiment 3.
Method
Participants
An initial 48 undergraduate students from Dalhousie University participated in exchange for psychology course credit. Two of these participants were replaced with new participants because of experimenter concerns regarding the degree of effort put forth by one of these individuals, and because the other individual reported prior knowledge of the directed forgetting paradigm. The data from these two participants were not analyzed, either before or after these individuals were selected for replacement. The final sample size consisted of 48 participants: 46 from the initial sample, plus two new recruits.
The research protocol was determined by the Dalhousie Social Sciences and Research Ethics Board to be in compliance with the Canadian Tri-Council Policy on Ethical Conduct for Research with Human Participants.
Stimuli and apparatus
Psyscope X (https://doi.org/http://psy.ck.sissa.it/index.html; see Cohen, MacWhinney, Flatt, & Provost, 1993), running on 24-in. Macintosh iMac computers equipped with Apple Universal Serial Bus QWERTY keyboards, was used to present stimuli and collect keyboard responses.
We employed the same list of 320 nouns that was used by Taylor and Hamm (2016); the characteristics of those words are described in their article. Custom software was used to randomly distribute 160 words to a study wordlist and 160 words to a recognition foil list whose presentation was limited to the final recognition test.
During the study phase trials, three boxes, outlined with a 1-point black line, were presented along the horizontal meridian of the computer monitor. In Psyscope, the boxes were set to 175 points wide and 100 points high; at a viewing distance of 57 cm from the computer monitors, the three boxes were 4.8° of visual angle wide and 3.5° of visual angle tall. The middle box was positioned in the center of the computer monitor, and each box was separated by 10.4° of visual angle measured from its center to the center of the box next to it.
Stimuli were presented in black on a white background. The only exception was the fixation stimulus, which consisted of black crosshairs (“+”) that changed to green for correct target responses or to red for incorrect target responses. The endogenous cue stimulus consisted of two less-than signs that directed attention to the left (“<<”), or two greater-than signs that directed attention to the right (“>>”). The target stimulus consisted of forward and backward slash characters arranged to converge at the top (“/\”), to require an “up” response from participants, or at the bottom (“\/”), to require a “down” response. All study and recognition words were written in lowercase letters. The fixation, cue, and target stimuli, along with the study words, appeared in the center of the stimulus box in which they were presented. The memory instructions were communicated via high-frequency (1170-Hz) and low-frequency (260-Hz) tones played to both ears through Sony MDR-XD100 headphones.
Procedure
All participants completed the experiment procedures individually, in a single session of 1-h maximum duration. Before beginning the experiment, participants provided informed consent and were given a verbal and written overview of the experiment. This was followed by tone familiarization trials, target practice trials, study trials, and recognition test trials. Before each set of trials, printed instructions were presented on the computer monitor. At these times, participants were given the opportunity to ask the experimenter to return to the room for clarification; no participant elected to do so.
Tone familiarization
Participants were first presented with ten tone familiarization trials. Fixation crosshairs (“+”) were presented in the center of the computer monitor on a white background for 500 ms. The crosshairs were then replaced with a text description (e.g., “High Tone—REMEMBER”) for 1,000 ms. Midway through this presentation, the corresponding tone played over the headphones for 500 ms. For half of the participants, the high tone was assigned to the remember instruction and the low tone was assigned to the forget instruction; this assignment was reversed for the other half of the participants. Familiarization trials were separated by an intertrial interval (ITI) of 1,000 ms, during which the computer monitor remained completely white.
Target practice
Following the tone familiarization trials, participants received a step-by-step demonstration of study trial events. After this, they were given 32 practice trials selected at random from the full experimental design. These trials were identical to the experimental trials, except that no study word needed to be remembered or forgotten. Instead, the word word appeared on every trial. The purpose of these trials was to give participants practice making speeded responses to accurately discriminate the target orientation, without the demands of a concurrent memory task. Participants were instructed to place their left index finger on the “f” key of the keyboard and their right index finger on the “j” key and to press one of these keys as quickly and as accurately as possible to report the orientation of each target stimulus. Half of the participants were instructed to press the “f” key when the target was oriented with the vertex up (“/\”) and to press the “j” key when the target was oriented with the vertex down (“\/”); the other half of the participants received the opposite instructions.
Study
Following the target practice trials, participants were introduced to the study trials. They were informed that the procedures were identical to those for the target practice, except that they would be presented with a different word on each trial. They were told that a memory instruction presented via a high or low tone would follow each word presentation, and they were reminded of the meaning of those tones. Participants were instructed to try to remember words that were followed by the remember instruction and told that they could forget words that were followed by the forget instruction. Participants were informed that their memory would be tested later; they were not informed that the recognition test would include forget words and foil words in addition to remember words.
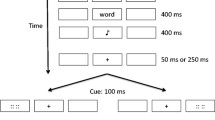
As depicted in the top panel of Fig. 1, each study trial began with the presentation of three empty stimulus boxes for 2,000 ms. A fixation crosshair (“+”) then appeared in the center of the middle stimulus box for 500 ms. After this, the crosshair disappeared and a study word was presented for 400 ms. This study word appeared with equal probability in the left or right stimulus box and was followed immediately by the presentation for 400 ms of an auditory memory instruction (i.e., a high or low tone, with equal probability). The stimulus boxes remained empty during the auditory memory instruction. Following the memory instruction, a random half of the trials imposed a 50-ms ISI, and the other half imposed a 250-ms ISI prior to the onset of the cue. The cue appeared in the center of the middle stimulus box for 500 ms before being replaced by a black fixation crosshair that remained visible for the remainder of the trial. The cue was as likely to direct attention to the left (“<<”) as to the right (“>>”). At a fixed 1,000-ms stimulus onset asynchrony (SOA)—that is, 1,000 ms after the onset of the cue—the target appeared in the center of either the left or the right box and remained visible for 100 ms. This target appeared in the cued location on 80% of trials and in the uncued location on 20% of trials. Cued and uncued targets were as likely to be oriented with the vertex up (“/\”) as down (“\/”). Participants were allowed up to 1,500 ms to make a speeded response to report the target orientation. At the end of this response window, visual feedback was presented for 500 ms. This feedback consisted of the fixation crosshair color changing to green if participants made a correct discrimination response to the target, or to red if they made an incorrect response. If the participant failed to respond by pressing either the “f” or the “j” key within the discrimination response window, everything on the computer monitor disappeared and was replaced with a question mark (“?”) in the center for 500 ms.

(Top) Methods used in the study trials of Experiment 1. Note that only one target is represented (“/\”), whereas in the experiment there were two targets (“/\,” “\/”) that occurred equally often and required a speeded discrimination response; the timeline depicts trial events only until the end of the target presentation. Cued targets are those that appear in the location indicated by the cue; uncued targets are those that appear in the location opposite that indicated by the cue. See the text for more details. (Bottom) Mean target reaction times (RTs) to make a correct target discrimination response on the Experiment 1 study trials, as a function of memory instruction (remember, forget), instruction–cue interstimulus interval (ISI; short = 50 ms, long = 250 ms), and target location (cued, uncued). Error bars represent Fisher’s least significant difference for the plotted effect
There were 160 study trials, comprised of 20 trials presented in a 4:1 ratio of cued:uncued targets within each cell of the following factorial design: Memory Instruction (remember, forget) × Instruction–Cue ISI (short, long) × Cue Direction (left, right) × Target Orientation (up, down). Both cued and uncued targets were as likely to appear on the left as on the right, and both were as likely to be oriented up as down; this controlled for spatial compatibility effects, given that half of all target trials in each condition therefore required a spatially compatible keypress response, whereas the other half required a spatially incompatible keypress response. For the purpose of our analyses, we collapsed over cue direction and target orientation, to reconceptualize our design as Memory Instruction (remember, forget) × Instruction–Cue ISI (short, long) × Target Location (cued, uncued). Cued trials were ones in which the target appeared at the location indicated by the cue (80% of trials); uncued trials were ones in which the target appeared at the location opposite that indicated by the cue (20% of trials).
Recognition trials
Following the study trials, participants were given a total of 320 recognition trials that randomly intermixed the presentation of 80 remember words, 80 forget words, and 160 foil words. Participants were instructed to press the “y” key for words that they recognized from the study trials, regardless of whether they had been instructed to remember or forget these words, and the “n” key for words that they did not recognize. These instructions remained visible at the top of the computer monitor during completion of the test trials. Words were presented one at a time, along with the prompt “Do you recognize this word? (y/n)” displayed beneath each word; both remained visible until participants submitted a response. Keyboard presses were echoed on the monitor to the right of the prompt. Participants could self-correct entries using the backspace key. When participants were ready to submit a response and advance to the next trial, they pressed the RETURN key. A “y” response to remember and forget words constituted a recognition hit; a “y” response to a foil word constituted a false alarm.
Data analysis
Data collation and analyses were conducted using R Studio 1.1.383 running R 3.4.3 (R Core Team, 2017) and various R packages including: plyr (Wickham, 2011), dplyr (Wickham & Francois, 2016), tidyr (Wickham, 2017b), and stringr (Wickham, 2017a). R package ez 4.4-0 (Lawrence, 2016) was used to calculate descriptive statistics (ezStats) and within-subjects analyses of variance (ezANOVA), and to create plots (ezPlot) that were then modified using ggplot2 (Wickham, 2009).
Applying the approach described by Masson (2011), a Bayesian information criterion (BIC) approximation of Bayesian posterior probabilities was generated using the output from ezANOVA. In this article, we refer to the approximated posterior probability of the alternative effect (i.e., nonzero effect) as pH1 and the approximated posterior probability of the null effect as pH0. The two probabilities summed together equal 1; we will therefore report whichever probability is largest, using the verbal descriptors suggested by Raftery (1995): .50–.75 = weak, .75–.95 = positive, .95–.99 = strong, > .99 = very strong. We interpret our results based on the posterior probability estimates; however, for readers inexperienced with this approach to data analysis, we also present the F-test statistics and generalized eta-squared (ges) as a measure of effect size.
Preliminary data analyses determined the mean foil false alarm rates on the recognition test for each participant, and also the overall accuracy in discriminating the target orientation on study trials. Accurate target responses were those on which participants correctly identified the target orientation by pressing the corresponding response key within 100–1,500 ms of target onset. Data were excluded from all subsequent analyses for participants whose false alarm rate was > 2 standard deviations above the mean of all participants and/or whose mean target accuracy was > 2 standard deviations below the mean of all participants.
Trial-by-trial data (as opposed to collapsed condition means) were used to calculate the descriptive statistics (ezStats) and inferential statistics (ezANOVA), and to create plots (ezPlot).
Results
Preliminary analyses led to the removal of data contributed by two participants. Both of these participants had average foil false alarm rates that were > 2 standard deviations above the mean of all participants; one of these participants also had average target accuracy > 2 standard deviations below the mean of all participants. All subsequent analyses were conducted using the data contributed by the remaining 46 participants.
Recognition trials
Participants made recognition “yes” responses to 60% of remember words, 33% of forget words, and 13% of unstudied foil words. An analysis of these data revealed very strong evidence for an effect of word type (remember, forget, foil), pH1 > .99 [F(2, 90) = 163.77, MSE = 150.39, p < .01, ges = .60]. Pairwise comparisons provided very strong evidence for more hits to forget words than false alarms to unstudied foils, pH1 > .99 [F(1, 45) = 141.37, MSE = 62.73, p < .01, ges = .38] and, more importantly, for a directed forgetting effect, with more hits to remember words than to forget words, pH1 > .99 [F(1, 45) = 94.62, MSE = 170.38, p < .01, ges = .36]. This suggests that participants made use of the memory instructions provided during the study trials and enables us to ask the key question of whether endogenous cueing effects were influenced by these instructions.
Study trials
For the study trials, mean RTs were calculated for correct target discrimination responses made within 100–1,500 ms of target onset. Target discrimination accuracy is summarized in Table 1.
Target RTs
Mean target RTs are shown in the bottom panel of Fig. 1 and were analyzed as a function of memory instruction (remember, forget), instruction–cue ISI (short = 50 ms, long = 250 ms), and target location (cued, uncued). This analysis provided positive evidence against an effect of memory instruction, pH0 = .84 [F < 1, MSE = 4,274.15, p > .46, ges < .01] and weak evidence against an effect of instruction–cue ISI, pH0 = .68 [F(1, 45) = 2.32, MSE = 4,504.16, p > .13, ges < .01]. There was, however, very strong evidence for an effect of target location, with overall faster RTs to targets in the cued location (M = 653 ms) than to targets in the uncued location (M = 739 ms), pH1 > .99 [F(1, 45) = 48.54, MSE = 13,795.18, p < .01, ges = .09]. This cueing effect demonstrates that attention was allocated in accordance with the cues.
We found weak evidence against a two-way interaction between memory instruction and instruction–cue ISI, pH0 = .55 [F(1, 45) = 3.46, MSE = 4,024.77, p > .06, ges < .01]. Countering our hypothesis, there was also positive evidence against the critical interaction between memory instruction and target location, pH0 = .86 [F < 1, MSE = 2,622.76, p > .70, ges < .01]: On remember trials, RTs were 86 ms faster to targets in the cued location than to targets in the uncued location; on forget trials, RTs were 85 ms faster to targets in the cued location than to targets in the uncued location. There was positive evidence against a two-way interaction between instruction–cue ISI and target location, pH0 = .83 [F < 1, MSE = 3,669.88, p > .41, ges < .01], and against a three-way interaction between memory instruction, instruction–cue ISI, and target location, pH0 = .85 [F < 1, MSE = 3768.16, p > .57, ges < .01]. In other words, the cueing effect did not interact with any other variable(s).
Target accuracies
Target accuracies were analyzed in a manner analogous to the analysis of target RTs. This analysis provided positive evidence against an effect of memory instruction, pH0 = .87 [F < 1, MSE = 122.51, p > .95, ges < .01], and against an effect of instruction–cue ISI, pH0 = .87 [F < 1, MSE = 98.37, p > .75, ges < .01]. There was, however, very strong evidence for an effect of target location, pH1 > .99 [F(1, 45) = 40.16, MSE = 249.72, p < .01, ges = .12]. Countering a speed–accuracy trade-off, this effect reflects the fact that not only were cued targets discriminated more quickly than uncued targets (see Fig. 1), they also were discriminated more accurately: The overall accuracy was 92% for discriminating cued targets and 81% for discriminating uncued targets.
We also found positive evidence against all two-way interactions: between memory instruction and instruction–cue ISI, pH0 = .85 [F < 1, MSE = 199.40, p > .57, ges < .01]; between memory instruction and target location, pH0 = .85 [F < 1, MSE = 87.96, p > .53, ges < .01]; and between instruction–cue ISI and target location, pH0 = .86 [F < 1, MSE = 133.69, p > .69, ges < .01]. There was also weak evidence against a three-way interaction between memory instruction, instruction–cue ISI, and target location, pH0 = .69 [F(1, 45) = 2.22, MSE = 79.94, p > .14, ges < .01].
Discussion
Performance on the recognition test provided very strong evidence for a directed forgetting effect, indicating that participants were able to effectively implement the memory instructions. The 60% hit rate for remember words and the 33% hit rate for forget words were comparable to the rates reported in the Taylor and Hamm (2016) study whose methods informed the present investigation (their recognition hits ranged from 61%–65% for remember items and 22%–30% for forget items). The present hit rates were also comparable to those in other dual-task implementations of an item-method directed forgetting task (i.e., ranging from 46%–74% for remember items and from 31%–53% for forget items; e.g., Fawcett & Taylor, 2008, 2010, 2012; Taylor, 2005; Taylor & Fawcett, 2011; Thompson et al., 2014). The magnitude of the directed forgetting effect (i.e., remember hits – forget hits = 27%) also fell within the range of the effects from those studies (i.e., ~ 12%–40%). Thus, there is no reason to believe that the successful implementation of the memory instructions was influenced by the requirement that participants use top-down control to voluntarily direct their endogenous attention to the location predicted by the central cue. The very large endogenous cueing effect would also seem to counter any suggestion that voluntary allocation was impeded by the concurrent memory task: Averaged over remember and forget trials, RTs were 85 ms faster to cued targets than to uncued targets.
Now consider the main question of interest. Despite the fact that participants used the memory instructions to commit remember items to memory and to disregard forget items, we found no evidence that the implementation of these instructions influenced the availability of endogenous attention. The central arrow cues were very effective in encouraging the allocation of attentional resources to the cued location. However, they were no more effective on forget trials than on remember trials. In both cases, participants were able to direct endogenous attentional resources to the cued location ahead of the target. This enabled overall faster responses to cued targets than to uncued targets, without regard to prior trial events. This runs counter to our hypothesis that endogenous cueing effects would be larger following forget instructions than following remember instructions, which is predicted if endogenous attentional resources are liberated by the forget instruction and remain available to be allocated in the cueing task.
In their study, Taylor and Hamm (2016) presented words in peripheral locations and—after an interposed memory instruction and exogenous cue—required participants either to localize the onset of a target to the left or right (Exp. 2) or to discriminate its orientation (Exp. 3). Despite the change in target task from localization to discrimination, the results remained the same: They found no interaction of memory instruction with exogenous cueing. The results from our Experiment 1 extend this finding by demonstrating that there is also no interaction of memory instruction with endogenous cueing. However, before drawing this conclusion too strongly, we thought it worthwhile to conduct a second experiment in which we eased the cognitive demands of the task.
In Experiment 1, it is likely that participants had to repeatedly shift their attention to perform the task. Attention was required to shift from center to the periphery in order to read, attend, and establish a mental representation of the study word; then attention had to return to center in time to process the endogenous cue; finally attention had to again shift to the periphery in accordance with the cue. In Experiment 2, we sought to ease these demands by presenting study words at center, as Taylor and Hamm (2016) did in their Experiment 1. This would allow attention to remain focused at center until shifted voluntarily in accordance with the endogenous cue.
Experiment 2
In Experiment 2, all study words were presented at center rather than in the periphery. In all other respects, the methods replicated those of Experiment 1. The goal, again, was to determine whether endogenous cueing effects would be larger following forget instructions than following remember instructions.
Method
Participants
Initially, a total of 48 undergraduate students from Dalhousie University participated in exchange for psychology course credit. One participant reported previous experience in a directed forgetting task and so was replaced by a new recruit; the data from the excluded participant were not analyzed either before or after replacement.
Stimuli and apparatus
The stimuli and apparatus were the same as those used in Experiment 1.
Procedure
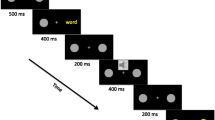
The experimental procedures replicated those of Experiment 1, with the exception that all study words were presented at center, as depicted in the top panel of Fig. 2.

(Top) Methods used in the study trials of Experiment 2. Note that only one target is represented (“/\”), whereas in the experiment there were two targets (“/\,” “\/”) that occurred equally often and required a speeded discrimination response; the timeline depicts trial events only until the end of the target presentation. Cued targets are those that appear in the location indicated by the cue; uncued targets are those that appear in the location opposite that indicated by the cue. See the text for more details. (Bottom) Mean target reaction times (RTs) to make a correct target discrimination response on the Experiment 2 study trials, as a function of memory instruction (remember, forget), instruction–cue interstimulus interval (ISI; short = 50 ms, long = 250 ms), and target location (cued, uncued). Error bars represent Fisher’s least significant difference for the plotted effect
Results
Preliminary analyses led to the removal of data contributed by three participants. Two of these participants had average foil false alarm rates that were > 2 standard deviations above the mean of all participants; one participant had an average target accuracy > 2 standard deviations below the mean of all participants. All subsequent analyses were conducted using data contributed by the remaining 45 participants.
Recognition trials
Participants made recognition “yes” responses to 70% of remember words, 36% of forget words, and 12% of unstudied foil words. An analysis of these data revealed very strong evidence for an effect of word type (remember, forget, foil), pH1 > .99 [F(2, 88) = 291.36, MSE = 128.12, p < .01, ges = .73]. Pairwise comparisons provided very strong evidence for more hits to forget words than false alarms to unstudied foils, pH1 > .99 [F(1, 44) = 149.44, MSE = 83.08, p < .01, ges = .44], and also for a directed forgetting effect, with more hits to remember words than to forget words, pH1 > .99 [F(1, 44) = 185.54, MSE = 138.58, p < .01, ges = .53]. Thus, as was the case for Experiment 1, the results of Experiment 2 suggest that participants made use of the memory instructions provided during the study trials. This enabled us to ask the key question of whether endogenous cueing effects were influenced by these instructions.
Study trials
As was the case for Experiment 1, mean RTs were calculated for correct target discrimination responses made within 100–1,500 ms of target onset. Target discrimination accuracy is summarized in Table 1.
Target RTs
Mean target RTs are shown in the bottom panel of Fig. 2 and were analyzed as a function of memory instruction (remember, forget), instruction–cue ISI (short = 50 ms, long = 250 ms), and target location (cued, uncued). This analysis provided positive evidence against an effect of memory instruction, pH0 = .84 [F < 1, MSE = 3,188.22, p > .55, ges < .01], and against an effect of instruction–cue ISI, pH0 = .86 [F < 1, MSE = 4,419.31, p > .71, ges < .01]. However, similar to Experiment 1, we observed very strong evidence for an effect of target location, with overall faster RTs to targets presented in a cued location (M = 656 ms) rather than in an uncued location (M = 761 ms), pH1 > .99 [F(1, 44) = 86.71, MSE = 10,301.40, p < .01, ges = .13]. This cueing effect confirms that participants were able to allocate their attention in accordance with the endogenous cues.
There was positive evidence against a two-way interaction between memory instruction and instruction–cue ISI, pH0 = .85 [F < 1, MSE = 1,975.45, p > .57, ges < .01]. Critically, there also was positive evidence against an interaction between memory instruction and target location, pH0 = .79 [F(1, 44) = 1.16, MSE = 2,435.33, p > .28, ges < .01]: On remember trials, RTs were 108 ms faster to cued targets than to uncued targets; on forget trials, RTs were 101 ms faster to cued targets than to uncued targets. We also found positive evidence against a two-way interaction between instruction–cue ISI and target location, pH0 = .80 [F < 1, MSE = 3,797.66, p > .32, ges < .01], and against a three-way interaction between memory instruction, instruction–cue ISI, and target location, pH0 = .86 [F < 1, MSE = 2,366.22, p > .63, ges < .01]. Thus, replicating the results of Experiment 1, a cueing effect emerged that did not interact with any other variable(s).
Target accuracies
An analogous analysis of target accuracies provided positive evidence against an effect of memory instruction, pH0 = .83 [F < 1, MSE = 172.51, p > .43, ges < .01], and against an effect of instruction–cue ISI, pH0 = .87 [F < 1, MSE = 90.96, p > .78, ges < .01]. As had also been the case for Experiment 1, however, there was very strong evidence for an effect of target location, pH1 > .99 [F(1, 44) = 37.84, MSE = 278.10, p < .01, ges = .15]. Again countering a speed–accuracy trade-off, cued targets not only were discriminated more quickly than uncued targets (see Fig. 2), they also were discriminated more accurately: The overall accuracy was 94% for discriminating cued targets, versus 83% for discriminating uncued targets.
We found positive evidence against all two-way interactions: between memory instruction and instruction–cue ISI, pH0 = .81 [F < 1, MSE = 101.55, p > .35, ges < .01]; between memory instruction and target location, pH0 = .87 [F < 1, MSE = 117.49, p > .81, ges < .01]; and between instruction–cue ISI and target location, pH0 = .84 [F < 1, MSE = 120.24, p > .52, ges < .01]. There was also positive evidence against a three-way interaction between memory instruction, instruction–cue ISI, and target location, pH0 = .87 [F < 1, MSE = 131.55, p > .92, ges < .01].
Discussion
In Experiment 2, all study words were presented at center rather than in the periphery. In all other respects, the method was identical to Experiment 1. The rationale for presenting words at center was to reduce the demands otherwise created by having to shift attention to the peripheral word location and then back to center in time to receive the endogenous cue. We reasoned that easing the demands on attention in the interval prior to endogenous cue onset might uncover an interaction between memory instruction and endogenous cueing that was otherwise obscured in Experiment 1. On this, our data were clear: Even when words were presented at center, there continued to be compelling evidence against an interaction of memory instruction and endogenous cueing. Indeed, the pattern of results in Experiment 2 closely replicated that of Experiment 1.
Like Experiment 1, Experiment 2 provided very strong evidence for a directed forgetting effect. Where the directed forgetting effect is measured as the difference in hit rates between remember and forget words, the magnitude was 27% in Experiment 1 and a larger 34% in Experiment 2. Although there was weak evidence against a difference in the magnitudes of these directed forgetting effects, pH0 = .57 [F(1, 89) = 3.95, MSE = 303.32, p = .05, ges = .04], we analyzed remember word hit rates and forget word hit rates separately as a function of experiment. There was positive evidence that hit rates to remember words were higher in Experiment 2 (70%) than in Experiment 1 (60%), pH1 = .76 [F(1, 89) = 6.97, MSE = 330.26, p < .01, ges = .07]. There was, however, positive evidence against a difference in hit rates to forget words in Experiment 2 (36%) as compared to Experiment 1 (33%), pH0 = .87 [F < 1, MSE = 256.08, p > .41, ges < .01]. In other words, the numerically larger magnitude of the directed forgetting effect for centrally presented study words was attributable primarily to increased recognition of remember words, rather than to decreased recognition of forget words.
The fact that remember words were recognized at a lower rate in Experiment 1 (peripheral presentation) than in Experiment 2 (central presentation) is consistent with research demonstrating that attention-demanding processes interfere with remembering (Barrouillet, Portrat, Vergauwe, Diependaele, & Camos, 2011; Camos & Portrat, 2015). Barrouillet et al. argued that the more cognitively demanding a task is, the more time attention must be allocated to processing the task and the less time attention is available for maintaining a memory representation. Following this argument, participants likely had less time to refresh the remember words in memory in Experiment 1 than in Experiment 2 (i.e., due to the need in Experiment 1 to shift their attention to the periphery before returning it to center to receive the endogenous cue). The fact that the remember hit rates were sensitive to the change in study word presentation from the periphery (Exp. 1) to center (Exp. 2) implies that this manipulation was effective in reducing the cognitive demands of the task.
In addition to a directed forgetting effect, Experiment 2 also revealed an overall endogenous cueing effect. As was the case in Experiment 1, the results of Experiment 2 provided very strong evidence for a cueing effect, with faster responses to targets presented in a cued location rather than in an uncued location. Numerically, the magnitude of this cueing effect was larger in Experiment 2 (104 ms) than in Experiment 1 (86 ms). However, there was positive evidence against a difference in the magnitude of the cueing effect as a function of experiment, pH0 = .87 [F < 1, MSE = 6,169.60, p > .39, ges < .01]. Indeed, even when we analyzed the data separately for cued and uncued trials, there was positive evidence against a difference between experiments for both cued-target RTs, pH0 = .90 [F < 1, MSE = 13,565.24, p > .94, ges < .01] and uncued-target RTs, pH0 = .89 [F < 1, MSE = 16,672.05, p > .55, ges < .01]. In other words, the endogenous cues were effective at directing attention to the peripheral locations in advance of the target and were equally successful at doing so in Experiments 1 and 2.
Critically, despite having relieved participants of the need to repeatedly shift attention in advance of the endogenous cue, the central word presentation of Experiment 2 did not uncover an interaction between memory instruction and endogenous cueing. The magnitude of the endogenous cueing effects was no larger on forget trials (M = 101 ms) than on remember trials (M = 108 ms). The conclusion of Experiment 2 therefore reinforces that of Experiment 1: A forget instruction does not make endogenous attentional resources any more available for subsequent goal-directed allocation than does a remember instruction. This conclusion is reinforced by an analysis of target RTs that included memory instruction (remember, forget), instruction–cue ISI (short = 50 ms, long = 250 ms), and target location (cued, uncued) as within-subjects factors and experiment as a between-subjects factor. Reiterating the pattern of results found within each experiment on the within-subjects factors, we found positive evidence against main effects of both memory instruction, pH0 = .86 [F < 1, MSE = 3,737.29, p > .34, ges < .01], and instruction–cue ISI, pH0 = .79 [F(1, 89) = 1.80, MSE = 4,462.21, p > .18, ges < .01], but very strong evidence in support of an overall effect of target location, with faster RTs to cued targets than to uncued targets, pH1 > .99 [F(1, 89) = 128.73, MSE = 12,067.92, p < .01, ges = .11]. Across the two experiments, there was positive evidence against all interactions between the within-subjects factors: memory instruction and instruction–cue ISI, pH0 = .82 [F(1, 89) = 1.47, MSE = 3,011.63, p > .22, ges < .01]; the critical interaction between memory instruction and target location, pH0 = .89 [F < 1, MSE = 2,530.10, p > .64, ges < .01]; instruction-cue ISI and target location, pH0 = .81 [F(1,89) = 1.64, MSE = 3,733.05, p > .20, ges < .01]; as well as between memory instruction, instruction–cue ISI, and target location, pH0 = .90 [F < 1, MSE = 3,075.07, p > .87, ges < .01]. In addition, this analysis provided positive evidence against a main effect of experiment, pH0 = .90 [F < 1, MSE = 109,005.50, p > .72, ges < .01], and against all interactions with experiment: memory instruction and experiment, pH0 = .90 [F < 1, MSE = 3,737.29, p > .87, ges < .01]; instruction–cue ISI and experiment, pH0 = .87 [F < 1, MSE = 4,462.21, p > .41, ges < .01]; target location and experiment, pH0 = .87 [F < 1, MSE = 12,076.92, p > .38, ges < .01]; memory instruction, instruction–cue ISI, and experiment, pH0 = .64 [F(1, 89) = 3.36, MSE = 3,011.63, p > .06, ges < .01]; memory instruction, target location, and experiment, pH0 = .85 [F(1, 89) = 1.04, MSE = 2,530.10, p > .30, ges < .01]; instruction–cue ISI, target location, and experiment, pH0 = .90 [F < 1, MSE = 3,733.05, p > .89, ges < .01]; and memory instruction, instruction–cue ISI, target location, and experiment, pH0 = .88 [F < 1, MSE = 3,075.07, p > .47, ges < .01].
General discussion
Intentional forgetting involves cognitive control processes that limit the rehearsal of unwanted or irrelevant information for the purpose of directing limited-capacity resources to more important or relevant tasks (Kuhl et al., 2007; Souza et al., 2014; Williams et al., 2013). This involves an active withdrawal of attentional resources from the representation in working memory of unwanted forget item traces (Fawcett & Taylor, 2010, 2012; Taylor, 2005; Taylor & Fawcett, 2011; Thompson et al., 2014; see also Rizio & Dennis, 2013, and Lee, 2018). The present results reinforce Taylor and Hamm’s (2016) supposition that this attentional withdrawal lasts only long enough to stop unwanted item rehearsal. In the longer-term, neither are exogenous attentional resources more available for bottom-up capture (Taylor & Hamm, 2016) nor are endogenous attentional resources more available for top-down allocation.
Top-down prioritization of the target task?
We interpret the equivalent endogenous cueing effects following forget and remember instructions as evidence that no persistent changes in resource availability outlasted the instantiation of the memory instruction. At first blush, however, one might wonder whether it could instead have been the case that endogenous attention was equally available following forget and remember instructions simply because participants made it available. Indeed, people can explicitly decide how to allocate their attention between multiple tasks based on the relative priority of those tasks (e.g., Brumby, Salvucci, & Howes, 2009; Jansen, van Egmond, & de Ridder, 2016; Janssen & Brumby, 2010; Janssen, Brumby, & Garnett, 2012; Wang, Proctor, & Pick, 2007). It follows that if our participants chose to prioritize performance on the target task, this might have led them to immediately suspend attention to remember word rehearsal in order to maintain a high level of performance on the target task, effectively equating attentional withdrawal on forget and remember trials. We do not think this likely, however.
It is typical to see dual-task trade-offs when one task is prioritized over another (e.g., Janssen et al., 2012). But in the present study, we found no evidence of a dual-task trade-off: Performance on the directed forgetting task was similar to that in other studies implementing an item-method directed forgetting paradigm (e.g., Fawcett & Taylor, 2008, 2010, 2012; Taylor, 2005; Taylor & Fawcett, 2011; Thompson et al., 2014). Moreover, combining the data across both experiments revealed a negligible positive correlation between the overall endogenous cueing effect and the magnitude of the directed forgetting effect [r(89) = .06, pH0 = .89, p > .57]. If it were the case that performance on the memory task was traded off for performance on the endogenous cueing task, this correlation should have been robust and negative (i.e., with larger cueing effects associated with smaller directed forgetting effects). On this basis, we do not believe that prioritization of the endogenous orienting task can explain the fact that endogenous cues were equally effective following remember and forget instructions.
Efficacy versus speed of endogenous orienting
When interpreting target RTs as a function of cue–target SOA, two theoretical constructs bear consideration. The first is asymptote: As the cue–target SOA is increased from 0, the cueing effect increases to an asymptotic level that reflects maximal preparedness for the target—the point along the time course function at which the RT difference is greatest for cued targets compared to uncued targets. The more available attentional resources are for endogenous allocation, and the more efficacious the cue is in orienting those resources, the higher the asymptote. The second construct is the rate of rise to asymptote. This reflects the speed of orienting. The faster that available resources can be maximally oriented to a cued location, the sooner asymptote is reached.
In the present study we purposely fixed our cue–target SOA at 1,000 ms—an interval during which endogenous orienting is expected to have reached asymptote and stabilized (e.g., Cheal & Lyon, 1991; Müller & Rabbitt, 1989; Ristic & Kingstone, 2006). Our rationale for doing so was that if a forget instruction makes endogenous attentional resources relatively more available, this should result in higher asymptotic performance on forget trials than on remember trials, as reflected in a larger overall cueing effect. Had such a difference occurred, however, we would have been obligated to demonstrate that the difference was, in fact, due to different asymptotic levels rather than different rise times. Indeed, if two functions rise at different rates to the same asymptote, the faster-rising function will produce a larger cueing effect than the slower-rising function when sampled at pre-asymptotic intervals. Barring such a difference in the magnitude of the cueing effect on forget and remember trials, however, there is no compelling reason to manipulate cue–target SOA. Not only does our hypothesis concern a difference in the asymptotic availability of attentional resources on forget trials as compared to remember trials, there was also no a priori rationale for expecting forget instructions to alter the speed of orienting attentional resources to a cued peripheral location. This is an important point because, in the absence of persistent effects of forget instructions on the availability of endogenous attention, one might wonder why we did not decrease the cue–target SOA to at least characterize any short-term changes that occurred immediately post-instruction. As we hope we have made clear, early immediate effects of memory instructions on the overall availability of endogenous attentional resources cannot be ascertained simply by decreasing the cue–target SOA.
Immediate versus downstream consequences of memory intentions
Whereas we deliberately fixed the cue–target SOA at 1,000 ms, we did vary the interval between the memory instruction and the endogenous cue. Whether this post-instruction interval was 50 or 250 ms, participants were able to orient attention to the cued location—and able to do so equally well on remember trials and forget trials. Consequently, targets that appeared at the cued location were discriminated more quickly than targets that appeared at the uncued location, with no discernable effect of the instruction–cue ISI. This suggests that the attentional processes that underlie the interpretation and implementation of forget instructions versus remember instructions differ for a duration that is less than the 450 ms that comprised the combined duration of our instruction (400 ms) and the shortest instruction–cue ISI (50 ms). The implication is that the initial withdrawal of attention from forget item processing releases limited-capacity resources only long enough to stop unwanted forget item rehearsal before switching to cumulative rehearsal of previous-trial remember items. Indeed, this may account for the fact that the attentional blink—a deficit in second-target processing that follows first-target processing (e.g., Raymond, Shapiro, & Arnell, 1992)—is smaller following a forget instruction than following a remember instruction, but only within a temporal window that extends 400–500 ms postinstruction (Taylor, 2018).
If instructions to forget prompt changes in attention that last for only 400–500 ms—less than the 450- to 650-ms interval that separated the onset of our memory instruction (400-ms duration) from the onset of our endogenous cue (after a 50- or 250-ms delay)—it raises the question of whether this interval should have been decreased. We think not. Decreasing the interval between instruction onset and endogenous cue onset is not instructive with respect to our hypothesis that memory instruction has persistent downstream consequences for the efficacy of orienting the endogenous attentional system. Moreover, designing such a study would face challenges from pragmatic considerations of stimulus timing. This is because to minimize directional confusions, an endogenous cue must be presented for at least 300 ms (Müller, 1986). It happens that 300 ms is also the shortest effective memory instruction duration of which we are aware (Bancroft et al., 2013). Even if these minimum stimulus durations were implemented, the only way that a 300-ms endogenous cue could follow within 400–500 ms of a 300-ms memory instruction would be for the two to temporally overlap for 100–200 ms. This risks interference between the perceptual and control processes needed to perceive, interpret, and implement the memory instruction and those needed to perceive, interpret, and implement the endogenous orienting cue—quite apart from any overlap in endogenous attention per se.
To complicate matters, even if actual temporal overlap were eliminated by reducing the duration of the memory instruction to 100 ms—considerably shorter than in any published study of which we are aware—this would not necessarily assuage concerns about the design. There are two reasons for this. First, the neural activity that distinguishes forget trial processing and remember trial processing extends 400–500 ms after instruction onset (Hsieh et al., 2009). This suggests that a shorter instruction presentation duration would not necessarily result in a shorter processing duration. Second, subsequent memory effects (ERP waveforms that distinguish successful and unsuccessful encoding attempts) are associated with posterior late wave potentials that occur after about 600–1,000 ms on remember trials, but not on forget trials (see Hsieh et al., 2009; van Hooff & Ford, 2011). It follows that to present an endogenous cue within 400–500 ms of memory instruction onset would likely disrupt the memory processes needed to encode and retain remember words in particular. This would, in turn, undermine successful measurement of a directed forgetting effect. In short, it is not at all clear that characterizing early changes in attention following a memory instruction can be accomplished using a behavioral paradigm.
Conclusion
Regardless of its inability to characterize the nature of short-term changes in attention that occur following instructions to remember and forget, the present investigation is conclusive with regard to the critical question of whether there are persistent changes in endogenous attention: There are not. This finding complements that of Taylor and Hamm (2016) by demonstrating that forget instructions do not influence the downstream availability of either exogenous attentional resources or endogenous attentional resources. This bolsters the claim made by Taylor and Hamm that memory intentions influence attention only long enough to regulate encoding processes: Any attentional resources that are withdrawn from forget item processing are most likely reallocated within the memory task—presumably to the cumulative rehearsal of preceding remember items—rather than being freed for capture or allocation to other external stimulus events.
Change history
21 February 2019
In this issue, there is an error in the citation information on the opening page of each article HTML. The year of publication should be 2019 instead of 2001. The Publisher regrets this error.
Notes
The effect of selective rehearsal on encoding is what makes the item-method paradigm relevant for our purposes. Nevertheless, it is worth noting that the poorer subsequent memory for forget items than for remember items might not be accounted for entirely by mechanisms that operate at encoding—additional mechanisms may function to reduce forget item retrieval as well (e.g., Marevic & Rummel, 2018; Rummel, Marevic, & Kuhlmann, 2016; although see Taylor, Cutmore, & Pries, 2018).
Given the complexity of the dual memory–attention task, we chose these cues on the grounds that they would be relatively easy to interpret without actually being arrows. Nevertheless, we acknowledge that their presumed ease of interpretation is related to the perception that these stimuli “point” to the left or right; indeed, this is how they were described to participants. This introduced the possibility that these stimuli might have had an exogenous effect on attention, in addition to the endogenous influence that we intended to measure (e.g., Ristic & Kingstone, 2006). This possibility was not particularly worrying, however, given that Taylor and Hamm (2016) provided evidence against an interaction of memory instruction and exogenous cueing. It follows that any interaction between memory instruction and cueing in the present investigation would therefore be most reasonably attributed to endogenous orienting prompted by the high predictive validity (80%) of these cues.
References
Bancroft, T. D., Hockley, W. E., & Farquhar, R. (2013). The longer we have to forget the more we remember: The ironic effect of postcue duration in item-based directed forgetting. Journal of Experimental Psychology: Learning, Memory, and Cognition, 39, 691–699. https://doi.org/10.1037/a0029523
Barrouillet, P., Portrat, S., Vergauwe, E., Diependaele, K., & Camos, V. (2011). Further evidence for temporal decay in working memory: Reply to Lewandowsky and Oberauer (2009). Journal of Experimental Psychology, 37, 1302–1317.
Berger, A., Henik, A., & Rafal, R. (2005). Competition between endogenous and exogenous orienting of visual attention. Journal of Experimental Psychology: General, 134, 207–221. https://doi.org/10.1037/0096-3445.134.2.207
Brumby, D. P., Salvucci, D. D., & Howes, A. (2009). Focus on driving: How cognitive constraints shape the adaptation of strategy when dialing while driving. In S. Greenberg, S. E. Hudson, K. Hinkley, M. R. Morris, & D. R. Olsen Jr. (Eds.), Proceedings of the SIGCHI Conference on Human Factors in Computing Systems (pp. 1629–1638). New York: ACM Press.
Camos, V., & Portrat, S. (2015). The impact of cognitive load on delayed recall. Psychonomic Bulletin & Review, 22, 1029–1034.
Cheal, M., & Lyon, D. R. (1991). Central and peripheral precuing of forced-choice discrimination. Quarterly Journal of Experimental Psychology, 43, 859–880. https://doi.org/10.1080/14640749108400960
Cheng, S., Liu, I., Lee, J. R., Hung, D. L., & Tzeng, O. J.-L. (2012). Intentional forgetting might be more effortful than remembering: An ERP study of item-method directed forgetting. Biological Psychology, 89, 283–292.
Chica, A. B., Bartolomeo, P., & Lupiáñez, J. (2013). Two cognitive and neural systems for endogenous and exogenous spatial attention. Behavioural Brain Research, 237, 107–123.
Cohen, J., MacWhinney, B., Flatt, M., & Provost, J. (1993). PsyScope: An interactive graphic system for designing and controlling experiments in the psychology laboratory using Macintosh computers. Behavioral Research Methods, Instruments, & Computers, 25, 257–271. https://doi.org/10.3758/BF03204507
Danziger, S., & Kingstone, A. (1999). Unmasking the inhibition of return phenomenon. Perception & Psychophysics, 61, 1024–1037. https://doi.org/10.3758/BF03207610
Fawcett, J. M., Lawrence, M. A., & Taylor, T. L. (2016). The representational consequences of intentional forgetting: Impairments to both the probability and fidelity of long-term memory. Journal of Experimental Psychology: General, 145, 56–81. https://doi.org/10.1037/xge0000128
Fawcett, J. M., & Taylor, T. L. (2008). Forgetting is effortful: Evidence from reaction time probes in an item-method directed forgetting task. Memory & Cognition, 36, 1168–1181. https://doi.org/10.3758/MC.36.6.1168
Fawcett, J. M., & Taylor, T. L. (2010). Directed forgetting shares mechanisms with attentional withdrawal but not with stop-signal inhibition. Memory & Cognition, 38, 797–808. https://doi.org/10.3758/MC.38.6.797
Fawcett, J. M., & Taylor, T. L. (2012). The control of working memory resources in intentional forgetting: Evidence from incidental probe word recognition. Acta Psychologica, 139, 84–90.
Gallant, S. N., & Dyson, B. J. (2016). Neural modulation of directed forgetting by valence and arousal: An event-related potential study. Brain Research, 1648, 306–316.
Gardiner, J. M., Gawlik, B., & Richardson-Klavehn, A. (1994). Maintenance rehearsal affects knowing, not remembering: Elaborative rehearsal affects remembering, not knowing. Psychonomic Bulletin & Review, 1, 107–110. https://doi.org/10.3758/BF03200764
Hopfinger, J. B., & West, V. M. (2006). Interactions between endogenous and exogenous attention on cortical visual processing. NeuroImage, 31, 774–789.
Hourihan, K. L., & Taylor, T. L. (2006). Cease remembering: Control processes in directed forgetting. Journal of Experimental Psychology: Human Perception and Performance, 32, 1354–1365.
Hsieh, L. T., Hung, D. L., Tzeng, O. J. L., Lee, J. R., & Cheng, S. K. (2009). An event-related potential investigation of the processing of Remember/Forget cues and item encoding in item-method directed forgetting. Brain Research, 1250, 190–201.
Jansen, R. J., van Egmond, R., & de Ridder, H. (2016). Task prioritization in dual-tasking: Instructions versus preferences. PLoS ONE, 11, e0158511. https://doi.org/10.1371/journal.pone.0158511
Janssen, C. P., & Brumby, D. P. (2010). Strategic adaptation to performance objectives in a dual-task setting. Cognitive Science, 34, 1548–1560.
Janssen, C. P., Brumby, D. P., & Garnett, R. (2012). Natural break points: The influence of priorities and cognitive and motor cues on dual-task interleaving. Journal of Cognitive Engineering and Decision Making, 6, 5–29.
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eye’s movement. In J. Long & A. Baddeley (Eds.), Attention and performance IX (pp. 187–203). Hillsdale: Erlbaum.
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43, 346–354. https://doi.org/10.3758/BF03208805
Kuhl, B. A., Dudukovic, N. M., Kahn, I., & Wagner, A. D. (2007). Decreased demands on cognitive control reveal the neural processing benefits of forgetting. Nature Neuroscience, 10, 908–914. https://doi.org/10.1038/nn1918
Lawrence, M. A. (2016). ez: Easy analysis and visualization of factorial experiments. R package version 4.4. http://CRAN.R-project.org/package=ez
Lee, Y. S. (2012). Cognitive load hypothesis of item-method directed forgetting. Quarterly Journal of Experimental Psychology, 65, 1110–1122.
Lee, Y. S. (2018). Withdrawal of spatial overt attention following intentional forgetting: evidence from eye movements. Memory, 26, 503–513.
Lee, Y. S., & Lee, H. M. (2011). Divided attention facilitates intentional forgetting: Evidence from item-method directed forgetting. Consciousness and Cognition, 20, 618–626. https://doi.org/10.1016/j.concog.2010.09.008
Lee, Y. S., Lee, H. M., & Tsai, S. H. (2007). Effects of post-cue interval on intentional forgetting. British Journal of Psychology, 98, 257–272.
Liu, T., Pestilli, F., & Carrasco, M. (2005). Transient attention enhances perceptual performance and fMRI response in human visual cortex. Neuron, 45, 469–477.
Lu, Z.-L., & Dosher, B. A. (2000). Spatial attention: Different mechanisms for central and peripheral temporal precues? Journal of Experimental Psychology: Human Perception and Performance, 26, 1534–1548. https://doi.org/10.1037/0096-1523.26.5.1534
Ludowig, E., Möller, J., Bien, C. G., Münte, T. F., Elger, C. E., & Rosburg, T. (2010). Active suppression in the mediotemporal lobe during directed forgetting. Neurobiology of Learning and Memory, 93, 352–361.
MacLeod, C. M. (1998). Directed forgetting. In J. M. Golding & C. M. MacLeod (Eds.), Intentional forgetting: Interdisciplinary approaches (pp. 1–57). Mahwah: Erlbaum.
Marevic, I., & Rummel, J. (2018). Retrieval-mediated directed forgetting in the item-method paradigm: The effect of semantic cues. Psychological Research, (in press).
Masson, M. E. J. (2011). A tutorial on a practical Bayesian alternative to null-hypothesis significance testing. Behavior Research Methods, 43, 679–690. https://doi.org/10.3758/s13428-010-0049-5
McCormick, P. A. (1997). Orienting attention without awareness. Journal of Experimental Psychology: Human Perception and Performance, 23, 168–180. https://doi.org/10.1037/0096-1523.23.1.168
Müller, H. J. (1986). The effect of selective spatial attention on peripheral discrimination thresholds (Doctoral dissertation). Durham University, Durham, UK.
Müller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary orienting of visual attention: Time course of activation and resistance to interruption. Journal of Experimental psychology: Human Perception and Performance, 15, 315–330. https://doi.org/10.1037/0096-1523.15.2.315
Paz-Caballero, M. D., Menor, J., & Jiménez, J. M. (2004). Predictive validity of event-related potentials (ERPs) in relation to the directed forgetting effects. Clinical Neurophysiology, 115, 369–377.
Popper, K. R. (1968). The logic of scientific discovery (Revised). London: Hutchinson & Co.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32, 3–25. https://doi.org/10.1080/00335558008248231
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. In H. Bouma & D. G. Bouwhuis (Eds.), Attention and performance X: Control of language processes (pp. 531–556). Hillsdale: Erlbaum.
R Core Team. (2017). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Retrieved from http://www.R-project.org
Raftery, A. E. (1995). Bayesian model selection in social research. Sociological Methodology, 25, 111–163. https://doi.org/10.2307/271063
Raymond, J. E., Shapiro, K. L., & Arnell, K. M. (1992). Temporary suppression of visual processing in an RSVP task: An attentional blink? Journal of Experimental Psychology: Human Perception and Performance, 18, 849–860. https://doi.org/10.1037/0096-1523.18.3.849
Riggio, L., & Kirsner, K. (1997). The relationship between central cues and peripheral cues in covert visual orientation. Attention, Perception, & Psychophysics, 59, 885–899.
Ristic, J., & Kingstone, A. (2006). Attention to arrows: Pointing to a new direction. Quarterly Journal of Experimental Psychology, 59, 1921–1930. https://doi.org/10.1080/17470210500416367
Rizio, A. A., & Dennis, N. A. (2013). The neural correlates of cognitive control: Successful remembering and intentional forgetting. Journal of Cognitive Neuroscience, 25, 297–312.
Rummel, J., Marevic, I., & Kuhlmann, B. G. (2016). Investigating storage and retrieval processes of directed forgetting: A model-based approach. Journal of Experimental Psychology: Learning, Memory, and Cognition, 42, 1526–1543.
Souza, A. S., Rerko, L., & Oberauer, K. (2014). Unloading and reloading working memory: Attending to one item frees capacity. Journal of Experimental Psychology: Human Perception and Performance, 40, 1237–1256. https://doi.org/10.1037/a0036331
Taylor, T. L. (2005). Inhibition of return following instructions to remember and forget. Quarterly Journal of Experimental Psychology, 58, 613–629.
Taylor, T. L. (2018). Remember to blink: Reduced attentional blink following instructions to forget. Attention, Perception, & Psychophysics, 80, 1489–1503. https://doi.org/10.3758/s13414-018-1528-5
Taylor, T. L., Cutmore, L., & Pries, L. (2018). Item-method directed forgetting: Effects at retrieval? Acta Psychologica, 183, 116–123.
Taylor, T. L., & Fawcett, J. M. (2011). Larger IOR effects following forget than following remember instructions depend on exogenous attentional withdrawal and target localization. Attention, Perception, & Psychophysics, 73, 1790–1814. https://doi.org/10.3758/s13414-011-0146-2
Taylor, T. L., & Hamm, J. P. (2016). Selection for encoding: No evidence of greater attentional capture following forget than remember instructions. Attention, Perception, & Psychophysics, 78, 168–186.
Thompson, K. M., Hamm, J. P., & Taylor, T. L. (2014). Effects of memory instruction on attention and information processing: Further investigation of inhibition of return in item-method directed forgetting. Attention, Perception, & Psychophysics, 76, 322–334.
van Hooff, J. C., & Ford, R. M. (2011). Remember to forget: ERP evidence for inhibition in an item-method directed forgetting paradigm. Brain Research, 1392, 80–92.
Wang, D. D., Proctor, R. W., & Pick, D. F. (2007). Acquisition and transfer of attention allocation strategies in a multiple-task work environment. Human Factors, 49, 995–1004.
Wickham, H. (2009). ggplot2: Elegant graphics for data analysis. New York: Springer. http://ggplot2.org
Wickham, H. (2011). The split–apply–combine strategy for data analysis. Journal of Statistical Software, 40, 1–29. http://www.jstatsoft.org/v40/i01/
Wickham, H. (2017a). stringr: Simple, consistent wrappers for common string operations (R package version 1.2.0). Retrieved from https://CRAN.R-project.org/package=stringr
Wickham, H. (2017b). tidyr: Easily tidy data with “spread()” and “gather()” functions (R package version 0.6.3). Retrieved from https://CRAN.R-project.org/package=tidyr
Wickham, H., & Francois, R. (2016). dplyr: A grammar of data manipulation (R package version 0.5.0). Retrieved from https://CRAN.R-project.org/package=dplyr
Williams, M., Hong, S. W., Kang, M.-S., Carlisle, N. B., & Woodman, G. F. (2013). The benefit of forgetting. Psychonomic Bulletin & Review, 20, 348–355. https://doi.org/10.3758/s13423-012-0354-3
Wylie, G. R., Foxe, J. J., & Taylor, T. L. (2008). Forgetting as an active process: An fMRI investigation of item-method-directed forgetting. Cerebral Cortex, 18, 670–682. https://doi.org/10.1093/cercor/bhm101
Yang, T., Lei, X., & Anderson, M. (2016). Decreased inhibitory control of negative information in directed forgetting. International Journal of Psychophysiology, 100, 44–51.
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10, 601–621. https://doi.org/10.1037/0096-1523.10.5.601
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Rubinfeld, L.M., Taylor, T.L. & Hamm, J.P. Selection for encoding: No evidence of better endogenous orienting following forget than following remember instructions. Atten Percept Psychophys 81, 237–252 (2019). https://doi.org/10.3758/s13414-018-1587-7
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1587-7