
Abstract
This study embedded attentional cues in the study phase of an item-method directed forgetting task. We used an unpredictive onset cue (Experiment 1), a predictive onset cue (Experiment 2), or a predictive central cue (Experiments 3–6) to direct attention to the left or right. In Experiments 1–5, this was followed by a pink or blue study word that required a speeded colour discrimination; in Experiment 6, it was followed by a pink or blue word or nonword that required a lexical decision. Each study word was followed by an instruction to Remember or Forget. A yes–no recognition test confirmed better recognition of to-be-remembered words than to-be-forgotten words; a cueing effect confirmed the effectiveness of predictive cues in allocating attentional resources. There was, however, no evidence that the directed forgetting effect differed for attended and unattended words: Encoding depends more on the memory intention formed after a study word has disappeared than on the availability of processing resources when that word first appears.
Similar content being viewed by others

Memory intentions can be formed at encoding or at retrieval. We are interested only in those formed at encoding. To study encoding intentions, participants are presented with an item-method directed forgetting paradigm, in which words are presented one at a time, and each is followed by an instruction to remember or forget (see MacLeod, 1998, for a review). Because participants do not know in advance whether each word will need to be remembered or forgotten, they must attend to each item as it is presented and maintain its representation in working memory until the memory instruction appears (cf. Gardiner, Gawlik, & Richardson-Klavehn, 1994). If the instruction is to remember, participants engage in elaborative rehearsal to commit the word to long-term memory (e.g., Hsieh, Hung, Tzeng, Lee, & Cheng, 2009), using semantic relational processing (e.g., Montagliani & Hockley, 2019). If the instruction is to forget, participants engage frontal control mechanisms (e.g., Cheng, Liu, Lee, Hung, & Tzeng, 2012; Hsieh et al., 2009; Rizio & Dennis, 2013; van Hoof & Ford, 2011; Wylie, Foxe, & Taylor, 2008) to withdraw attentional resources (Fawcett & Taylor, 2010; Lee, 2018; Taylor, 2005; Taylor & Fawcett, 2011; Thompson, Hamm, & Taylor, 2014), likely as a means for terminating further unwanted processing and rehearsal of the to-be-forgotten (TBF) item (cf. Hourihan & Taylor, 2006; see also Fellner, Waldhauser, & Axmacher, 2019). In this way, to-be-remembered (TBR) items receive more rehearsal than TBF items (e.g., Basden, 1996; Basden, Basden, & Gargano, 1993; Bjork, 1989; see MacLeod, 1998, for a review). Accordingly, subsequent explicit tests reveal better memory for TBR items than TBF items, a difference known as a directed forgetting effect.
Given that the intention to forget depends on removing attentional resources in an effort to prevent further item processing and rehearsal, we tested whether the corollary is also true: Does the prior allocation of attention to a TBF item interfere with the ability to intentionally forget that item? To address this question, the current study embedded a Posner cueing paradigm (Posner, 1980) into the study phase of an item-method directed forgetting paradigm. In a Posner cueing paradigm, a cue is used to capture or direct attention to the left or right visual periphery and reaction times (RTs) are measured to items that appear subsequently in the cued or in the uncued location. The rationale is that attention facilitates processing of items that appear within its locus. Accordingly, to the extent that attention is successfully allocated to the cued location, RTs are faster to respond to items that appear subsequently in that location, rather than in an uncued location—an RT difference known as a cueing effect.
On each of a series of study trials, we thus presented a cue that was intended to capture or direct attention to the left or right. A coloured study word then appeared in the cued or in the uncued location, and participants were required to make a speeded response to report the colour of this word; their RTs to do so were used to measure a cueing effect. The study word was then followed by an instruction to remember or forget. After all study trials were presented, participants were tested for their recognition of study items—those that had been designated TBF as well as those that had been designated TBR. Performance on the recognition test was used to measure a directed forgetting effect.
We expected to find a cueing effect, with faster colour discrimination RTs to study words that appeared in a cued location compared with those that appeared in an uncued location. This would confirm that attentional resources were relatively available to support immediate study word processing at the cued location, compared with at the uncued location. Indeed, at the uncued location, item processing would be delayed by the need to first disengage attentional resources from the cued location and then to reallocate them to the word location (Posner, 1980, 1988; Posner, Walker, Friedrich, & Rafal, 1984). Ultimately, of course, attention would be allocated to all study items—after all, participants still needed to read each word and maintain it in working memory until they learned whether it was designated TBR or TBF. However, we reasoned that attention would be available sooner and for a more prolonged duration for cued study words than for uncued study words. The question was whether this processing advantage for cued compared with uncued words would influence the likelihood of remembering and forgetting those items when later instructed to do so.
Although increased processing time can improve recognition of both TBR and TBF items (e.g., Hockley, Ahmad, & Nicholson, 2016), we expected the prolonged availability of attentional resources to have relatively less effect on TBR items than on TBF items. This is because the commitment of TBR items to long-term memory depends primarily on postinstruction elaboration—that is, processing that occurs after participants have learned that the word must be committed to memory. In contrast, the successful exclusion of TBF items from long-term memory depends on whether the instruction to forget comes soon enough (Hourihan & Taylor, 2006; see also Timmins, 1974) and is of sufficiently short duration (e.g., Bancroft, Hockley, & Farquhar, 2013) to prevent ongoing rehearsal from forming an indelible trace. Indeed, even in the post-instruction interval, forgetting improves when attentional resources are relatively unavailable (Lee & Lee, 2011). Accordingly, we reasoned that facilitated item processing at a cued location is more likely to disrupt the mechanisms needed to intentionally forget than it is to enhance those needed to intentionally remember. In other words, facilitated study word processing at the cued location compared with the uncued location should have relatively little impact on TBR item recognition, whereas it should increase TBF item recognition and thereby reduce the directed forgetting effect. The result would be a smaller directed forgetting effect for attended study words than for unattended study words. The goal of this study was to test this prediction.
Experiment 1: Exogenous cues
In Experiment 1, we used exogenous cues (Jonides, 1981). These cues consisted of the onset of a visual stimulus (“:: ::”) in a location to the left or right of the computer monitor. The location of the cue did not predict the study word location; instead, the study word was as likely to appear in the cued location as it was to appear in the uncued location. Because there was no incentive for participants to attend to the cues, these cues were expected to produce a cueing effect only to the extent that they were effective in capturing attention to the cued location automatically. Presuming they did so, we expected to use the recognition test results to ascertain whether this attention capture influenced the magnitude of the directed forgetting effect. This would be revealed as an interaction between cue condition and memory instruction.
Method
Participants
A total of 48 Dalhousie University students were recruited as an initial sample. During debriefing, three participants indicated that they did not comply fully with all task instructions; their data were excluded before conducting any analyses and replaced with data from three new recruits. Participants were tested individually in a session that lasted approximately 60 minutes. They received optional course credit in exchange for their participation.
Stimuli and apparatus
PsyScope X (cf. Cohen, MacWhinney, Flatt, & Provost, 1993) was loaded on 27-in Apple iMac computers and used to present stimuli and record responses from USB QWERTY keyboards. Except where noted otherwise, all text items presented as stimuli in the study and recognition phases were displayed in black on a uniform white background, using 24-point default PsyScope font.
During the study phase, three outline boxes were presented across the horizontal meridian of the computer monitor. Using the PsyScope tools, these boxes were drawn with a 1-point line and were set to 200 points wide and 150 points tall (approximately 5.5 degrees of visual angle × 3.7 degrees of visual angle when viewed from a distance of 65 cm). A fixation stimulus consisted of crosshairs (“+”) centred in the middle stimulus box; the cue consisted of four colons interrupted by two spaces (“:: ::”), centred in the left or right stimulus box; and, the memory instruction consisted of a high-frequency tone (1170 Hz) or a low-frequency tone (260 Hz) played over both channels of Sony MDR-XD100 stereo headphones.
Study and foil words were drawn from the same 320-item list used by Taylor and Hamm (2016). Prior to testing each participant, custom software was used to randomly distribute these 320 words to eight lists of 20 study items each and one list of 160 unstudied (foil) items. Each of the eight study lists was assigned to one cell of the study trial design.
Procedure
Before beginning the experiment, participants were asked to provide informed consent and were given a detailed overview of the experiment before the researcher left the room. Participants then proceeded to complete tone familiarization, practice, study, and recognition trials. Before each set of trials, participants received detailed written instructions and an invitation to recall the researcher into the room if they had any questions; none elected to do so.
Tone familiarization trials
Ten tone familiarization trials gave participants practice associating each tone with its corresponding memory instruction. A random half of these trials presented a high tone and the other half presented a low tone. Each trial started with the presentation of fixation crosshairs (“+”) in the centre of the computer monitor for 500 ms. This fixation stimulus was replaced by a verbal descriptor of the tone-instruction mapping (e.g., “High Tone—REMEMBER”) that remained visible at center for 1,000 ms. Halfway through this presentation interval, the corresponding tone sounded for 500 ms.
Target practice trials
A total of 32 trials gave participants practice making speeded responses to discriminate the study word colour, but without a concurrent memory load. These trials were identical to the study trials (see below), except that word appeared as the study item on every trial and there was no requirement to commit any items to memory.
Study trials
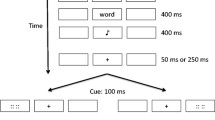
A total of 160 study trials were presented. As depicted in Fig. 1, each trial started with a 2,000-ms delay, during which the computer monitor remained blank except for the three outline stimulus boxes centred across the horizontal aspect. After this delay, fixation crosshairs (“+”) appeared for 500 ms in the middle of the centre stimulus box. Except for during cue presentation, these fixation crosshairs remained visible through the remainder of the trial.Footnote 1

Schematic representation of the study trials in Experiment 1
The cue appeared for 100 ms in the middle of the left or right stimulus box. This was followed immediately by a pink or blue study word in the left or right stimulus box. Where this study word also served as the attention target, this meant that the cue–target stimulus onset asynchrony (SOA) was fixed at 100 ms, within the typical time course of exogenous orienting (e.g., Cheal & Lyon, 1991; Müller & Rabbitt, 1989).
Participants were required to report the colour of this study word by pressing the “j” or “f” key on the computer keyboard as quickly and as accurately as possible; colour–key mapping was counterbalanced across participants. The coloured study word remained visible for 400 ms before it disappeared and was followed by the 400-ms presentation of a high or low tone. This tone served as a Remember or a Forget instruction. Participants were instructed that a Remember tone meant they were to commit the study word to memory for a later test and that a Forget tone meant they could forget the study word. The tone–instruction mapping was counterbalanced within each level of the colour–key mapping. Participants were forewarned that the word might disappear, and the memory instruction appear, before they had time to report the colour of the study word; they were allowed a maximum of 1,500 ms from the first appearance of the study word to report its colour. Feedback was presented for 500 ms at the end of the trial and consisted of a change in the fixation stimulus from black to green to signal a correct discrimination response; a change in the fixation stimulus from black to red to signal an incorrect discrimination response; and, a change in the shape of the fixation stimulus, from black crosshairs to a question mark (?) to indicate that no press of the “f” or “j” key had been registered within the time limit. The purpose of providing feedback was to ensure that participants remained engaged with the colour discrimination task and could know whether their responses were being made correctly and within time limits. The feedback was presented at the end of each trial so as not to interfere with processing the memory instruction.
The study trials represented the randomized presentation of a fully factorial design: Cue Location (Left, Right) × Word Location (Left, Right) × Word Colour (Pink, Blue) × Memory Instruction (Remember, Forget). For the purpose of analysis, Cue Location and Word Location were collapsed into a new factor called Cue Condition: The Cued level of this factor comprised trials where the word appeared in the same location as the preceding cue (e.g., Left–Left, Right–Right) and the Uncued level comprised trials where the word appeared in the location opposite the preceding cue (e.g., Left–Right, Right–Left).
Recognition trials
After the last study trial, participants received instructions for the recognition trials. They were informed that words would appear one at a time on each trial and that they were to press the “y” key for “yes” to report words that they recognized from the earlier study trials and the “n” key for “no.” They were explicitly instructed that they should respond “y” to all words that they recognized, regardless of the associated memory instruction presented during the study trials. These instructions remained visible at the top of the computer monitor throughout the recognition trials.
Participants input each response using the computer keyboard. Keyboard input was echoed to the monitor. Participants could self-correct an input error by pressing the backspace key; they pressed the “return” key to submit their response and proceed to the next trial. There was no time limit for submitting a response.
The recognition trials were composed of the randomized presentation of the 160 studied words and 160 unstudied foil words. Recognition hits were defined as “y” responses to studied words; false alarms were defined as “y” responses to unstudied foil words.
Data analysis
Data were processed, analyzed, and plotted using R Studio 1.1.463 running R 3.5.3 (R Core Team, 2019) and using packages plyr (Wickham, 2011), dplyr (Wickham, Francois, Henry, & Müller, 2019), tidyr (Wickham & Henry, 2019), stringr (Wickham, 2019), ez (Lawrence, 2016), ggplot2 (Wickham, 2016). Using the methods described by Masson (2011), we used output from ezANOVA to generate a Bayesian Information Criterion (BIC) approximation to Bayesian posterior probabilities. Where pH1 refers to the approximated posterior probability of the alternative (i.e., nonzero effect) given the data and pH0 refers to the approximated posterior probability of a null effect given the data, these values sum to 1.0. As such, we will report only the effect with the larger probability, describing the data as providing evidence for an effect when pH1 is the larger value and against an effect when pH0 is the larger value. We will qualify these conclusions using the verbal descriptors suggested by Raftery (1995): 0.50–0.75 = “weak”; 0.75–0.95 = “positive”; 0.95–0.99 = “strong”; >0.99 = “very strong.” We will interpret our results in light of this Bayesian approach, but for interested readers we will also provide the F-test statistics and generalized eta squared (ges) as a measure of effect size.
Before conducting our critical analyses, we calculated for each participant the mean proportion of false alarms on the recognition test. We also calculated on a subject-by-subject basis the mean error rates for reporting the word colour on study trials. Trials were coded as errors if the responses were made using an incorrect key, within the first 100 ms of word onset (i.e., an anticipation), or after 1,500 had elapsed. We excluded from all subsequent analyses the data for any participants whose mean false-alarm rate was more than two standard deviations higher than the mean of all participants and/or whose mean colour discrimination accuracy was more than two standard deviations lower than the mean of all participants.
Following these exclusions, trial-by-trial data were used to calculate descriptive and inferential statistics and to plot the results, with the full design specified as a parameter in the relevant ez function (see Lawrence, 2016).
Results
The data from two participants were excluded based on high false-alarm rates and the data from another two participants were excluded based on low colour discrimination accuracy. This left a final sample size of 44 participants.
Colour discrimination RTs
The mean RTs to correctly report the study word colour are shown in Fig. 2a. We analyzed these data as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). This analysis provided positive evidence against a main effect of Cue Condition, consistent with the near-identical mean RTs to report the colour of study words that appeared at Cued (M = 709 ms) and Uncued (M = 710 ms) locations, F < 1, pH0 = 0.87. In other words, contrary to expectation, there was no evidence that the cue was effective in capturing attention to its onset location. This analysis also provided positive evidence against a main effect of Memory Instruction, F(1, 43) = 1.03, MSE = 1298.47, p > .31, ges < .01, pH0 = 0.80, and against an interaction of Cue Condition and Memory Instruction, F < 1, pH0 = 0.86.

Results of Experiment 1. a Mean correct RTs (ms) to discriminate the colour of the study word, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied words (i.e., hits), as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different
Colour discrimination accuracy
To ensure that the analysis of discrimination RTs was not compromised by a speed–accuracy trade-off, study word colour discrimination accuracy was likewise analyzed as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). These data are depicted in Table 1. This analysis revealed positive evidence against an effect of Cue Condition, F < 1, pH0 = 0.87, against an effect of Memory Instruction, F < 1, pH0 = 0.86, and against an interaction, F < 1, pH0 = 0.85. There was thus no evidence to suggest that the RT data were compromised by changes in accuracy.
Recognition hits
The mean percentage of recognition “yes” responses to studied words (hits) is shown in Fig. 2b as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For visual comparison, also shown is the mean false-alarm rate for unstudied Foils (dotted line).
With an average of 42% hits to study words that appeared in a Cued location and 43% to those that appeared in an Uncued location, an analysis of the recognition hit rate provided positive evidence against a main effect of Cue Condition, F < 1, pH0 = 0.82. In other words, there was no evidence that words that appeared in a Cued location were recognized at an overall higher rate than those that appeared in an Uncued location. There was, however, very strong evidence for an effect of Memory Instruction, F(1, 43) = 77.47, MSE = 757.19, p < .001, ges = 0.47, pH1 > .99. This confirms a directed forgetting effect, with overall better recognition of TBR words (M = 61%) than TBF words (M = 25%). Finally, there was positive evidence against an interaction of Cue Condition and Memory Instruction, F(1, 43) < 1, pH0 = 0.84.
Discussion
The recognition hit rates confirmed a directed forgetting effect, with better overall recognition of TBR words than TBF words. This confirms that participants were able to use the memory instructions to exert control over encoding. There was, however, no cueing effect in the RT data: Participants were equally fast to discriminate the study word colour whether the word appeared in the cued location or in the uncued location. Absent evidence of a cueing effect in the RTs, it is impossible to ascertain whether the directed forgetting effect differed for cued and uncued study words.
At first blush, one might be inclined to wonder whether the failure to find a cueing effect in the colour discrimination RTs was somehow related to the fact that these RTs had mean latencies over 700 ms. These responses seem unusually long when compared with a Donder’s-type choice reaction task (e.g., Gottsdanker & Shragg, 1985) or to RTs in a cueing task that requires a discrimination response (e.g., Chica, Lupiánez, & Bartolomeo, 2006). However, they do not seem unusually long in the context of an item-method directed forgetting paradigm. In other studies that required a response to a probe or target item embedded in the study trials of an item-method task, simple RTs to report the onset of a stimulus averaged between 400 and 500 ms (Fawcett & Taylor, 2008); RTs to localize a target were over 500 ms (Thompson et al., 2014); RTs to respond to a go/no-go target averaged 600–700 ms (Fawcett & Taylor, 2010); and, RTs to report the colour of a probe word varied between 700 and 800 ms (Fawcett & Taylor, 2012). In this context, colour discrimination RTs greater than 700 ms do not seem particularly unusual, especially given that the current paradigm had the added complexity of requiring participants to read and maintain in working memory the same item for which they were reporting the colour discrimination.
Thus, it is not immediately clear why we failed to find an effect of the exogenous cues. One possibility is that participants adopted an attentional set that favoured the appearance of only coloured items, rendering any other stimulus event relatively ineffective at capturing attention (Bacon & Egeth, 1994; Folk, Remington, & Johnston, 1992; Leber & Egeth, 2006; although, see Theeuwes, 1994, 2004). A second possibility is that the concurrent memory task commanded control over the endogenous (voluntary) attentional system to such an extent that the effects of endogenous orienting obscured any effects of exogenous (automatic) capture (Berger, Henik, & Rafal, 2005). And a third possibility is that our decision to remove the fixation stimulus during cue presentation meant that attentional resources successfully captured by the cue were redistributed by the onset of the fixation stimulus (at the same time as the peripheral word). This would have the effect of reducing both the RT benefits of having the study word appear in a cued location and RT costs of having the study word appear in an uncued location.
In any case, without evidence of attentional capture to the cued location, we cannot on the basis of Experiment 1 address our core question of whether the directed forgetting effect is smaller for cued study words than for uncued study words. For this reason, Experiment 2 replicated the methods of Experiment 1, except that the peripheral onset cues were given 80% predictive validity for the upcoming study word location. We reasoned that making the peripheral cues spatially predictive of the upcoming study word would encourage participants to attend to the onset, rather than to filter it out. This should have the added benefit of preventing an endogenous override of attention capture by the peripheral cue, since the goal of attending to each study word is in concert with the goal of using the cue to predict the study word location. It was less clear whether concerns over the reappearance of the fixation stimulus could also be addressed by adding predictive validity. But we decided that this was an empirical question: If Experiment 2 continued to show no cueing effects in RT, we would need to explore the role of the fixation stimulus disappearance/reappearance; however, if Experiment 2 did show cueing effects (see Footnote 1), we could rule this out as a concern.
Experiment 2: Peripheral cues
Experiment 2 was identical to Experiment 1, except that the peripheral onset cues correctly predicted the upcoming study word location with 80% accuracy.
Method
Participants
A total of 48 Dalhousie University students were recruited as an initial sample. Six participants from this original sample were replaced with new participants prior to any data analysis. Reasons for replacement were varied, but included failure to adhere to task demands, prior experience in a directed forgetting task, and a request to participate for pedagogical purposes only and not have data retained for research.
Stimuli and apparatus
The stimuli and apparatus were identical to those described for Experiment 1.
Procedure
The procedure was identical to Experiment 1, except for the following. Whereas the study word was equally likely to appear at the Cued and Uncued locations in Experiment 1, in Experiment 2 the study word appeared at the Cued location with 80% probability. Thus, of the 160 study trials, there was a total of 128 trials on which the study word appeared in the Cued location and 32 on which the study word appeared in the Uncued location. Within each type of trial (Cued, Uncued), the cue appeared equally often in the left and in the right stimulus box; the word was printed equally often in blue or in pink; and, the word was followed equally often by a Remember and by a Forget instruction. To accommodate the change in predictive validity, we increased the cue duration to 500 ms and the cue–target SOA (i.e., the time between onset of the cue and onset of the coloured study word) to 1,000 ms. These changes accommodate the longer time course for endogenous attentional orienting, compared with exogenous attention capture (e.g., Cheal & Lyon, 1991; Müller & Rabbitt, 1989).
Results
The data from 3 participants were excluded: One based on high false alarms, one based on low colour discrimination accuracy, and one based on both high false alarms and low colour discrimination accuracy. Subsequent analyses were conducted on data contributed by the 45 remaining participants.
Colour discrimination RTs
The mean RTs to correctly report the study word colour are shown in Fig. 3a. An analysis of these data provided very strong evidence for an effect of Cue Condition, F(1, 44) = 34.43, MSE = 3785.77, p < .05, ges = 0.04, pH1 > .99. This reflects a cueing effect, with overall faster RTs to study words that appeared in a Cued location (M = 740 ms) rather than in an Uncued location (M = 794 ms). There was, however, positive evidence against both a main effect of Memory Instruction, F < 1, pH0 = 0.87, and an interaction of Cue Condition with Memory Instruction, F < 1, pH0 = 0.87. The fact that there were no effects of Memory Instruction—either alone or in interaction with Cue Condition—is not terribly surprising given that the colour discrimination response was initiated by a study word that had disappeared by the time the memory instruction appeared.

Results of Experiment 2. a Mean correct RTs (ms) to discriminate the colour of the study word, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied words (i.e., hits), as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different
Colour discrimination accuracy
Data for the study word colour discrimination accuracy are shown in Table 1. The analysis of these data revealed weak evidence against an effect of Cue Condition, F(1, 44) = 3.66, MSE = 32.63, p > .06, ges = 0.02, pH0 = 0.53; positive evidence against an effect of Memory Instruction, F < 1, pH0 = 0.86; and, positive evidence against their interaction, F < 1, pH0 = 0.87. There was therefore no suggestion that the analysis of RTs was compromised by effects in target discrimination accuracy.
Recognition hits
The mean percentage of recognition “yes” responses is shown in Fig. 3b as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For visual comparison, also shown as a dotted line is the mean false-alarm rate for unstudied Foils.
An analysis of the recognition hit rate to studied words provided positive evidence against a main effect of Cue Condition, with an average of 45% hits to study words that appeared in a Cued location and 43% to those that appeared in an Uncued location, F(1, 44) = 1.20, MSE = 75.31, p > .27, ges < .01, pH0 = 0.79. Given that the RTs to perform the colour discrimination were faster to study words in the Cued compared with the Uncued location, we can surmise that attention had been allocated in accordance with the cue. Yet recognition did not differ for words that appeared in the attended versus unattended locations. There was, however, very strong evidence for an effect of Memory Instruction, F(1, 44) = 149.39, MSE = 406.60, p < .001, ges = 0.51, pH1 > .99. This confirms that a directed forgetting effect also occurred in Experiment 2, with overall better recognition of TBR words (62%) than TBF words (26%). Nevertheless, there was weak evidence against an interaction of Cue Condition and Memory Instruction, F(1, 44) = 1.96, MSE = 58.92, p > .16, ges < .01, pH0 = 0.72, indicating that the locus of attention at study word onset had no discernible effect on the magnitude of the directed forgetting effect.
Discussion
As was the case for Experiment 1, the results of Experiment 2 revealed a reliable directed forgetting effect, with better subsequent recognition of TBR words than TBF words. This confirms that participants were able to use the memory instructions to regulate encoding. The question, then, is whether their ability to do so differed depending on whether study words appeared at attended or unattended locations.
The results of Experiment 1 could not speak to this central question because there was no evidence that the spatially nonpredictive onset cues (i.e., exogenous cues) were successful in capturing attention. The results of Experiment 2, however, were more instructive. In Experiment 2, RTs were faster when reporting the colour of a study word that appeared in a cued location rather than in an uncued location. This cueing effect confirms that the peripheral cues used in Experiment 1 were effective at orienting attention when they were given high spatial predictability (80%) in Experiment 2. Nevertheless, our predictions were not borne out with respect to the potential role that attention might play in item memorability: Despite evidence that attention was successfully allocated to the peripheral location in advance of the study word, there was no evidence that item memorability differed for attended and unattended TBR and TBF items.
Before discussing our results further, we thought it valuable to replicate this finding. Whereas Experiment 2 presented spatially predictive peripheral onset cues, Experiment 3 presented central cues that likewise were spatially predictive of the upcoming word location.
Experiment 3: Endogenous cues
Experiment 3 was identical to Experiment 2, except that it used symbolic cues presented at centre that predicted the study word location with 80% accuracy. This ensured that the predictive cues did not themselves attract attention to the periphery by virtue of their abrupt onset in a peripheral location. This not only allowed for an independent replication of Experiment 2, it also ensured that exogenous attention and endogenous attention did not remain conflated. This is an important consideration, given that exogenous attention and endogenous attention are not two modes for orienting the same attentional resources, but are, instead, two independent but interacting attentional systems (see Chica, Bartolomeo, & Lupiáñez, 2013, for a review), each with its own pool of limited-capacity attentional resources (e.g., Berger et al., 2005).
Method
Participants
A total of 48 Dalhousie University students were recruited as an initial sample. Four participants from this original sample were replaced with new participants prior to any data analysis due to failure to adhere to task instructions.
Stimuli and apparatus
The stimuli and apparatus were identical to those used for Experiments 1 and 2, with the following exception. The cue stimulus was changed from a peripheral onset to a central cue consisting of two less-than or two greater-than signs that directed attention to the left (“<<”) and right (“>>”), respectively.
Procedure
The procedure was identical to Experiment 2, except that the cue was always presented in the middle of the centre stimulus box.
Results
The data from six participants were excluded: four based on high false alarms and two based on low colour discrimination accuracy. Subsequent analyses were conducted on data contributed by the 42 remaining participants.
Colour discrimination RTs
The mean RTs to correctly report the study word colour are shown in Fig. 4a. These data were analyzed as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). Similar to the predictive peripheral cues of Experiment 2, the predictive central symbolic cues used in Experiment 3 led to very strong evidence for an effect of Cue Condition, F(1, 41) = 43.54, MSE = 2602.73, p < .01, ges = 0.03, pH1 > .99. This resulted from overall faster RTs to study words that appeared in a Cued location (M = 707 ms) rather than in an Uncued location (M = 759 ms), indicative of an overall cueing effect. There was weak evidence against a main effect of Memory Instruction, F(1, 41) = 3.45, MSE = 1331.30, p > .07, ges < 0.01, pH0 = 0.54, and positive evidence against an interaction of Cue Condition and Memory Instruction, F < 1, pH0 = 0.84. Again, the fact that there was no compelling evidence of an effect of Memory Instruction on colour discrimination RTs—either alone or in interaction with Cue Condition—is expected, given that the colour discrimination response was initiated by the study word, which had disappeared by the time the memory instruction appeared.

Results of Experiment 3. a Mean correct RTs (ms) to discriminate the colour of the study word, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied words (i.e., hits), as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different
Colour discrimination accuracy
Study word colour discrimination accuracy was analyzed as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). These data are shown in Table 1. Unlike for Experiment 2, the results of Experiment 3 revealed very strong evidence for an effect of Cue Condition, F(1, 41) = 24.16, MSE = 40.76, p < .01, ges = 0.06, pH1 > 0.99. Importantly, this result counters a speed-accuracy trade-off: Whereas RTs were faster to discriminate the colour of study words that appeared in the Cued location rather than in the Uncued location, these responses were also more accurate (MCued = 90%, MUncued = 85%). There was positive evidence against an effect of Memory Instruction, F < 1, pH0 = 0.85, and weak evidence against the interaction of Cue Condition and Memory Instruction, F(1, 41) = 3.72, MSE = 71.79, p > .06, ges = 0.02, pH0 = 0.51.
Recognition hits
The mean percentage of recognition “yes” responses is shown in Fig. 4b as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). As was the case for Experiments 1 and 2, the mean false-alarm rate to unstudied Foils is also depicted.
An analysis of the recognition hit rate to studied words provided weak evidence against a main effect of Cue Condition, with an average of 40% hits to study words that appeared in a Cued location and 38% to those that appeared in an Uncued location, F(1, 41) = 1.58, MSE = 72.25, p > .21, ges < .01, pH0 = 0.75. In other words, subsequent recognition memory performance was unaffected by whether study words appeared in the initial focus of attention or outside of that focus. As was true for Experiment 2, however, there was very strong evidence for an effect of Memory Instruction, F(1, 41) = 72.20, MSE = 694.64, p < .01, ges = 0.42, pH1 > .99. This confirms a directed forgetting effect, with overall better recognition of TBR words (56%) than TBF words (22%). As was the case for Experiment 2, there was no evidence that the focus of attention differentially affected recognition of TBR and TBF items, as revealed by positive evidence against an interaction of Cue Condition and Memory Instruction, F < 1, pH0 = 0.85.
Discussion
The results of Experiment 3 showed a directed forgetting effect, with greater recognition of TBR words than of TBF words. There was also evidence that participants were faster to report the colour of a study word that appeared in a cued location rather than in an uncued location. This suggests that participants were effective in allocating their attention in accordance with the cue even when it was changed from a peripheral onset (Experiment 2) to a central instruction (Experiment 3). However, as was the case for Experiment 2, there was no compelling evidence in Experiment 3 that the locus of attention affected the magnitude of the directed forgetting effect.
Taken together, the results of Experiments 2 and 3 suggest that memory intentions influence the allocation of attention, but that the allocation of attention does not influence either overall item memorability or the successful implementation of memory intentions. That said, however, it is also possible that any effect of attention on directed forgetting is overridden by the onset of the word itself in the visual periphery. As a singleton onset, a study word presented alone in the visual periphery might capture attention resources automatically (Jonides & Yantis, 1988; Yantis & Jonides, 1984). Even if such capture is not sufficient to completely override the effect of the endogenous cue (as evidenced by the significant cueing effect in both Experiments 2 and 3), it might be sufficient to override any effect of the endogenous cue on subsequent recognition. To test this, Experiment 4 repeated exactly the methods of Experiment 3 except that a string of xs of equal length to the study word appeared as a distractor in the location opposite that word. This prevented the study word from being an onset singleton.
Experiment 4: Central cues with distractors
Experiment 4 was identical to Experiment 3, except that the coloured study word was presented simultaneously with a string of lower case xs.
Method
Participants
A total of 48 Dalhousie University students were recruited as an initial sample. One participant from this original sample was replaced prior to data analysis due to reported experience in prior studies of directed forgetting.
Stimuli and apparatus
The stimuli and apparatus were identical to those used for Experiment 3.
Procedure
The procedure was identical to Experiment 3, except that a distractor string of xs always appeared simultaneously with the study word. This distractor string was matched for character length to the study word; appeared in the location opposite the study word; and, was presented in black (so that the participant could not use information about the distractor to inform the colour discrimination response).
Results
The data from two participants were excluded: one based on high false alarms and one based on low colour discrimination accuracy. Subsequent analyses were conducted on data contributed by the 46 remaining participants.
Colour discrimination RTs
The mean RTs to correctly report the study word colour are shown in Fig. 5a. As in Experiment 3, the central predictive cues used in Experiment 4 led to very strong evidence for an effect of Cue Condition, F(1, 45) = 63.21, MSE = 3041.13, p < .01, ges = 0.05, pH1 > .99. This was due to overall faster RTs to study words that appeared in a Cued location (M = 743 ms) rather than in an Uncued location (M = 808 ms), indicative of an overall cueing effect. There was weak evidence for a main effect of Memory Instruction, F(1, 45) = 5.94, MSE = 1172.43, p < .02, ges < 0.01, pH1 = 0.72, with RTs to discriminate the study word colour slightly slower when the word was followed by a Forget instruction (M = 781 ms) rather than by a Remember instruction (M = 769 ms). This suggests that the appearance of the memory instruction during execution of the colour discrimination response might have had some influence on the speed of that response. Finally, there was weak evidence against an interaction of Cue Condition and Memory Instruction, F(1, 45) = 3.35, MSE = 1806.16, p > .07, ges < 0.01, pH0 = .57.

Results of Experiment 4. a Mean correct RTs (ms) to discriminate the colour of the study word, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied words (i.e., hits), as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different.
Colour discrimination accuracy
Study word colour discrimination accuracy data are shown in Table 1. As was also true for Experiment 3, the results of Experiment 4 revealed very strong evidence for an effect of Cue Condition, F(1, 45) = 43.25, MSE = 57.50, p < .01, ges = 0.15, pH1 > 0.99: Whereas RTs were faster to discriminate the colour of study words that appeared in the Cued location rather than in the Uncued location, they were also more accurate (MCued = 92%, MUncued = 84%). There was weak evidence against an effect of Memory Instruction, F(1, 45) = 2.08, MSE = 42.60, p > .15, ges < 0.01, pH0 = .71, and positive evidence against an interaction of Cue Condition and Memory Instruction, F(1, 45) = 1.33, MSE = 46.79, p > .25, ges < .01, pH0 = 0.78.
Recognition hits
The mean percentage of recognition “yes” responses is shown in Fig. 5b as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). The mean false-alarm rate to unstudied foils is also depicted.
An analysis of the recognition hit rate to studied words provided weak evidence against a main effect of Cue Condition, with an average of 42% hits to study words that appeared in a Cued location and 39% to those that appeared in an Uncued location, F(1, 45) = 2.92, MSE = 75.78, p > .09, ges < .01, pH0 = 0.61. Thus, despite evidence of a cueing effect in the colour discrimination RTs, there was no evidence that subsequent recognition was affected by whether study words appeared in an attended rather than in an unattended location. There was, of course, very strong evidence for an effect of Memory Instruction, F(1, 45) = 136.52, MSE = 387.89, p < .01, ges = 0.45, pH1 > 0.99. This confirms a directed forgetting effect, with overall better recognition of TBR words (57%) than TBF words (23%). Consistent with the preceding three experiments, there was weak evidence against an interaction of Cue Condition and Memory Instruction, F(1, 45) = 3.44, MSE = 76.41, p > .07, ges < 0.01, pH0 = 0.56, arguing that the magnitude of the directed forgetting effect was not discernibly different for items at attended and unattended locations.
Discussion
Adding a distractor to the location opposite the study word produced a pattern of results that largely echoed those of Experiment 3. There continued to be a directed forgetting effect, indicating that participants utilized the memory instructions. There was also a cueing effect, indicating that participants were successful at orienting their attention to the cued location in advance of the study word. Nevertheless, having a study word appear in the attended location, rather than in an unattended location, had no obvious effect on the magnitude of the directed forgetting effect. Thus, whether the study word appeared alone as an abrupt onset singleton (Experiment 3) or appeared along with a length-matched x-string (Experiment 4), the pattern of results was largely the same. Taken together with the results of Experiment 4, the results of Experiments 2 and 3 cannot be explained by competition with endogenous orienting by automatic capture of attention to a study word singleton.
The results of Experiments 2–4 argue that any processing advantage afforded to cued study words, compared with uncued study words, produces no discernible effect on the magnitude of the directed forgetting effect. This argues that attention does not, in itself, lead to encoding of a word presented in its focus—either in accordance with a subsequent Remember instruction or in opposition to a subsequent Forget instruction. This might be the case for one of two reasons. One possibility is that memory encoding is affected by the allocation of attention only after a memory intention is formed. A second possibility is that encoding is affected by attentional allocation at the time of study word onset, but that this effect is weaker than that of the subsequent memory intention.
Experiment 5 was intended to distinguish between these explanations. We replicated exactly the methods of Experiment 4, except that we replaced the x-string distractors with a different distractor word on every trial. On the recognition test, we assessed memory not only for the study words that participants intended to remember and forget, but also for the distractor words for which no explicit memory intention had been formed but to which attention had nevertheless been directed (i.e., when the study word appeared in the Uncued location, it meant that attention had initially been directed to the distractor in the opposite location). In the absence of a potentially stronger explicit memory intention, is there any evidence that the prior allocation of attention to a word location influences its encoding into long-term memory?
Experiment 5: Central cues with word distractors
Experiment 5 was identical to Experiment 4, except that the coloured study word was presented simultaneously with a word distractor. This allowed us to examine the influence of attention locus on intentional remembering and forgetting of the study word as well as on incidental encoding of the distractor word.
Method
Participants
Data were collected from a total of 74 Dalhousie University students.Footnote 2 Data from one participant were excluded prior to any analyses being conducted, due to self-reported colour-blindness; the participant was aware of the need to discriminate colours, but wished to participate in any case. This participant was not replaced.
Stimuli and apparatus
The stimuli and apparatus were identical to those used for Experiment 4, except that a larger list of words was required. We used the MRC psycholinguistics database (http://websites.psychology.uwa.edu.au/school/MRCDatabase/uwa_mrc.htm; Coltheart, 1981; Wilson, 1988) to draw words that we used to create a list of 320 matched-length pairs of nouns. These words had an average concreteness rating of 471; an average familiarity rating of 552; an average Kučera–Francis word frequency rating of 117; and a range of 3–7 letters, with a mean of 5. Prior to testing each participant, custom software was used to randomly swap word pairs and then distribute these pairs to eight lists of 20 study-distractor pairs and one list of 160 unstudied (foil) items. Each of the eight study-distractor pair lists was assigned to one cell of the study trial design, which was defined by Memory Instruction (Remember, Forget) × Cue Location (Left, Right) × Word Location (Left, Right) × Word Colour (Blue, Pink). The first member of each word pair served as the study word; the second member served as the distractor word.
Procedure
The procedure was identical to Experiment 4, except that the distractor was a length-matched word that appeared in black in the location opposite the study word. To keep the number of recognition trials equivalent to the preceding experiments (and not overwhelm or overburden participants), only one member of each pair was tested. Where a distractor is defined by the study word with which it was presented, the 320 recognition trials comprised the presentation of 40 TBR words, 40 TBF words, 40 TBR distractors, 40 TBF distractors, and 160 unstudied foil items.
Results
In addition to the participant with colour deficiency, the data from three participants were excluded: two based on high false alarms and one based on low colour discrimination accuracy. Subsequent analyses were conducted on data contributed by the 70 remaining participants.
Colour discrimination RTs
The mean RTs to correctly report the study word colour are shown in Fig. 6a. As was true for Experiments 3 and 4, the central predictive cues used in Experiment 5 led to very strong evidence for an effect of Cue Condition, F(1, 69) = 95.07, MSE = 6775.99, p < .01, ges = 0.08, pH1 > .99. This was due to overall faster RTs to study words that appeared in a Cued location (M = 705 ms) rather than in an Uncued location (M = 801 ms), indicative of an overall cueing effect. As was also true in Experiment 4, there was weak evidence for a main effect of Memory Instruction, F(1, 69) = 4.48, MSE = 1482.84, p < .04, ges < 0.01, pH1 = 0.52; RTs to discriminate the study word colour were slightly slower when the word was followed by a Forget instruction (M = 758 ms) rather than by a Remember instruction (M = 748 ms). Critically, there was positive evidence against an interaction of Cue Condition and Memory Instruction, F(1, 69) = 1.68, MSE = 1999.15, p > .19, ges < 0.01, pH0 = 0.78. This indicates that the central cues were equally effective at directing attention to the cued location on Remember and Forget trials.

Results of Experiment 5. a Mean correct RTs (ms) to discriminate the colour of the study word, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied Words and Distractors, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different
Colour discrimination accuracy
Study word colour discrimination accuracy data are shown in Table 1. As was also true for Experiments 3 and 4, the results of Experiment 5 revealed very strong evidence for an effect of Cue Condition, F(1, 69) = 62.25, MSE = 140.58, p < .01, ges = 0.19, pH1 > 0.99: Whereas RTs were faster to discriminate the colour of study words that appeared in the Cued location rather than in the Uncued location, they were also more accurate (MCued = 89%, MUncued = 78%). There was weak evidence for an effect of Memory Instruction, F(1, 69) = 4.62, MSE = 58.19, p < .04, ges < 0.01, pH1 = 0.54, and weak evidence against an interaction of Cue Condition and Memory Instruction, F(1, 69) = 2.96, MSE = 57.92, p > .08, ges < .01, pH0 = 0.66.
Recognition hits
The mean percentage of recognition “yes” responses was first analyzed as a function of Cue Condition (Cued, Uncued), Memory Instruction (Remember, Forget), and Test Item (Study Word, Distractor). The intention of this preliminary analysis was to confirm that memory performance was affected by whether the tested item was the Study Word or Distractor word. Indeed, there was very strong evidence for an effect of Test Item, F(1, 69) = 130.56, MSE = 306.06, p < .01, ges = 0.16, pH1 > .99, and for an interaction of Test Item with Memory Instruction, F(1, 69) = 62.69, MSE = 185.01, p < .01, ges = 0.05, pH1 > .99. These findings justified performing separate analyses for the recognition of study words and study trial distractors.
Study words
The mean percentage of recognition “yes” responses is shown in Fig. 6b (top) as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). The mean false alarm rate to unstudied foils is also depicted.
An analysis of the recognition hit rate to study words provided weak evidence against a main effect of Cue Condition, with an average of 35% hits to study words that appeared in a Cued location and 33% to those that appeared in an Uncued location, F(1, 69) = 3.14, MSE = 169.86, p > .08, ges < .01, pH0 = 0.64. Thus, despite evidence of a cueing effect in the colour discrimination RTs, there was no evidence that subsequent recognition of study words was affected by whether these words appeared in an attended rather than in an unattended location. There was, of course, very strong evidence for an effect of Memory Instruction, F(1, 69) = 73.75, MSE = 335.35, p < .01, ges = 0.17, pH1 > 0.99. This confirms a directed forgetting effect, with overall better recognition of TBR words (43%) than TBF words (25%). Consistent with the preceding four experiments, there was positive evidence against an interaction of Cue Condition and Memory Instruction, F(1, 69) = 1.98, MSE = 128.86, p > .16, ges < 0.01, pH0 = 0.76, arguing that recognition of study words was not differentially influenced by attention to TBR and TBF items.
Distractor words
The mean percentage of recognition “yes” responses to study trial distractor words is shown in Fig. 6b (bottom). An analysis of the these data provided weak to positive evidence against all main effects and interactions: Cue Condition, F(1, 69) = 2.69, MSE = 98.60, p > .10, ges < .01, pH0 = 0.69; Memory Instruction, F(1, 69) < 1, MSE = 98.12, p > .61, ges < 0.01, pH0 = 0.88; and, the interaction of Cue Condition and Memory Instruction, F(1,69) < 1, MSE = 71.55, p > .62, ges < 0.01, pH0 = 0.88. Thus, despite evidence of a cueing effect in RTs—which confirms that attention was initially directed to the study word on Cued trials (and, therefore, to the distractor word on Uncued trials)—there was no evidence that attention paid to a distractor word affected the likelihood of memory formation.
Discussion
Adding a distractor word to the location opposite a study word supports our claim that the allocation of attention does not play a discernible role in memory encoding in the absence of a memory intention. Across Experiments 2–5, both peripheral and central cues were effective at directing attention to a cued location in advance of study word onset. Nevertheless, any processing benefit afforded the cued study word neither enhanced subsequent commitment of that item to memory when it was designated TBR nor prevented its exclusion from memory when it was designated TBF. Importantly, this was not simply because a stronger memory intention overrode weaker encoding by attention: In Experiment 5, there was also no evidence for an effect of attention on the incidental encoding of distractor words for which there was no explicit memory intention.
Pooling data over Experiments 2–5
Even though none of Experiments 2–5 revealed the predicted interaction of attentional locus and directed forgetting in the recognition hit rates, all showed a cueing effect and all showed a directed forgetting effect. Given that all four of these experiments shared similar methods—they all presented spatially predictive cues and required participants to discriminate the colour of the study word—we thought it worthwhile to repeat our analyses after pooling these data. Density functions for the colour discrimination RTs are shown in Fig. 7, as a function of Cue Condition and Memory Instruction. The plots are aligned on zero, which corresponds with the appearance of the study word and a vertical dotted line at 400 ms on the x-axis marks the appearance of the memory instruction. There was very strong evidence for an effect of Cue Condition, F(1, 202) = 213.62, MSE = 4710.44, p < .01, ges = 0.05, pH1 > .99, consistent with the cueing effect observed in each of Experiments 2–5. This can be seen in Fig. 7 as a rightward shift in the density distributions for discrimination RTs in the Uncued condition, compared with the Cued condition. As can also be seen in Fig. 7, across this pooled data set, the majority of colour discrimination RTs occurred after the memory instruction. Perhaps not surprisingly, then, there was positive evidence for an effect of Memory Instruction, F(1, 202) = 9.11, MSE = 1391.57, p < .01, ges < .01, pH1 = 0.86, reflecting the fact that colour discrimination RTs were overall a little slower on Forget trials (M = 761 ms) than on Remember trials (M = 753 ms). Over this pooled data set, however, there was positive evidence against an interaction of Cue Condition and Memory Instruction in this analysis of discrimination RTs, F(1, 202) < 1, MSE = 1586.06, p > .78, ges < 0.01, pH0 = 0.93.

Density functions for the study word colour discrimination RTs, combined across Experiments 2–5. Data are plotted as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). Remember trial functions are plotted in black and Forget trial functions are plotted in white; grey depicts where these functions overlap. Plots are aligned on 0 at the x-axis, which corresponds with the appearance of the coloured study word that required a discrimination response. The vertical dashed line that intercepts the x-axis at 400 ms represents the appearance of the Memory Instruction
The corresponding analysis of discrimination accuracy revealed very strong evidence for an effect of Cue Condition, F(1, 202) = 107.29, MSE = 89.68, p < .01, ges = .01, pH1 > .99, with overall higher accuracy in the Cued condition (M = 91%)—for which the RTs were also fastest—than in the Uncued condition (M = 84%), thereby countering a speed–accuracy trade-off. There was only weak evidence for an effect of Memory Instruction, F(1, 202) = 6.26, MSE = 46.40, p < .02, ges < .01, pH1 = 0.61, with slightly more accurate responding on Forget trials (M = 88%) than on Remember trials (M = 87%). There was also only weak evidence for an interaction of Cue Condition and Memory instruction, F(1, 202) = 7.44, MSE = 50.57, p < .01, ges < .01, pH1 = .73: The difference in accuracy between Cued and Uncued trials was 8% for Remember trials and 6% for Forget trials.
In the analysis of recognition hits for studied words, the pooled data set provided weak evidence against an effect of Cue Condition, F(1, 202) = 4.75, MSE = 71.78, p < .04, ges < 0.01, pH0 = .57, with nearly identical hit rates to Cued (M = 36%) and Uncued (M = 35%) study words. Reflecting the robust directed forgetting effect that occurred in each of Experiments 2–5, in this pooled data set there was very strong evidence for an effect of Memory Instruction, F(1, 202) = 277.76, MSE = 505.90, p < .01, ges = 0.29, pH1 > .99. Indeed, the overall hit rate was 49% for TBR items and 23% for TBF items, reflecting a directed forgetting effect of 26% (i.e., TBR–TBF), the magnitude of which is typical of other studies from our lab that have combined measures of attention and memory (e.g., range ~12%–40%; see Fawcett & Taylor, 2008, 2010, 2012; Rubinfeld, Taylor, & Hamm, 2019; Taylor, 2005; Taylor & Fawcett, 2011; Thompson et al., 2014). Critically, there was positive evidence against an interaction of Cue Condition with Memory Instruction, F(1, 202) < 1, MSE = 61.60, p > .38, ges < .01, pH0 = 0.91. In other words, even when data were pooled across Experiments 2–5, there was no evidence to suggest that the directed forgetting effect was influenced by whether the study word appeared in an attended or in an unattended location.
This conclusion was corroborated by a correlational analysis. To perform this analysis, we calculated—on a subject-by-subject basis—the magnitude of the cueing effect obtained for the colour discrimination RTs (i.e., Uncued RTs–Cued RTs) and the magnitude of the directed forgetting effect obtained for recognition hits (i.e., TBR hits–TBF hits). This analysis provided only weak evidence for the predicted negative correlation, r = −.18, t = 2.64(201), p < .01, pH0 = .58, and the correlation did not survive after using Cook’s distance (Cohen & Cohen, 1983) to reject outliers on both measures. Thus, despite the fact that endogenous attentional facilitation is sustained rather than transient in nature (Carrasco, 2011, for a review), the increased processing opportunity provided by the prior allocation of attention to the study word location appears to afford little memorial advantage: Attention seems to matter less than intention. At least, this is true when attention is allocated to a study word location in advance of a perceptual discrimination.
Experiment 6: Central cues and a lexical decision task
The results of the preceding experiments suggest that the prior allocation of attention to the study word location speeds the discrimination of the study word colour but has no discernible influence on the directed forgetting effect. However, it remains possible that we did not observe a meaningful interaction of attentional cueing and directed forgetting across these studies because the attention task and the memory task were focused on different aspects of word processing: The attention task required participants to focus on the perceptual qualities of the study word (i.e., colour), whereas selective rehearsal in the directed forgetting task required participants to process the word itself. In a final experiment, we addressed this possibility by requiring participants to perform a lexical decision task rather than a colour discrimination task during the study trials.
Adapting the methods of Experiment 4, each study trial in Experiment 6 presented a central cue that predicted the most likely location of a coloured letter string. This pink or blue letter string, which appeared opposite an x-string distractor, was configured as a word or a nonword. Attentional cueing was measured as faster and/or more accurate lexical decisions for strings that appeared in the cued location rather than in the uncued location. The subsequent instruction to Remember or Forget applied only to word strings.
Our interim conclusion from Experiments 2–5 is that the prior allocation of attention has no discernible effect on subsequent encoding control. If this is true only when an attentional set has been established for a perceptual (colour) discrimination, then the lexical decision task in Experiment 6 should reveal the hitherto elusive interaction between attentional cueing and directed forgetting. If, however, attentional cueing measured by a lexical decision task continues to show no interaction with directed forgetting, this would underscore the generalizability of our findings from Experiments 2–5 and emphasize that the prior allocation of attention to a study word location neither facilitates intentional remembering nor impairs intentional forgetting.
Method
Participants
Due to the global COVID-19 pandemic, data collection for Experiment 6 took place online. Sona-Systems was used to recruit participants from eligible online classes at Dalhousie University. A link in Sona-Systems directed interested students to the experiment URL, which was hosted on the Pavlovia.org server. To account for high online attrition rates, the Sona-Systems software was set to allow up to 90 participants to volunteer for this study, with the goal of collecting at least 48 complete data files, consistent with the sample size used in Experiment 4 (on which the general methods of Experiment 6 were based). Periodic manual checks of the Pavlovia server were made to determine the number of collected data files, and recruitment was stopped as soon as we became aware that this number exceeded the minimum goal of 48. This resulted in a total of 58 files being downloaded, only 54 of which were determined to contain data for all phases of the experiment and retained for analysis.
Stimuli and apparatus
The stimuli and apparatus were altered to accommodate online data collection. The software used to present stimuli and collect responses was developed using PsychoPy Builder (Peirce et al., 2019) and modified using customized Python script within the Builder interface. Within Builder, this software was auto-translated to JavaScript, which was uploaded to and hosted on the Pavlovia.org server.
Before beginning online recruitment, custom software written in Python 3 was used to randomly draw 192 word and matched-length x-string pairs from the list used in Experiment 4. Over 100 iterations, these items were randomized and assigned to experiment conditions to create unique stimulus files numbered 0–99. For each stimulus file iteration, a total of 128 of word pairs were selected for study trial presentation, with the remaining 64 pairs allocated to the foil list. Of the 128 word pairs selected for presentation as study items, half were reconfigured as nonwords. This was accomplished by searching word strings letter-by-letter, and replacing each vowel (including “y”) in turn with a random draw from a consonant set that excluded the letters “x” (because this was used to define the distractor string) and “y” (which was treated as a vowel). The resulting stimulus files were uploaded to the Pavlovia server and made accessible to the experiment software. During recruitment, Sona-Systems assigned a random number to each participant that was used as the participant identifier on Pavlovia; the experiment software used the last two digits of this random number to select the matching numbered iteration of the stimulus file. In this way, there was the possibility of a unique stimulus configuration for every participant.
We used a consistent response key mapping in the lexical decision task, such that participants were required to press the “w” key to report a word and the “n” key to report a nonword, with no counterbalancing of this assignment. This intuitive mapping was meant to reduce potential memory demands on participants. The lack of counterbalancing was not a worry, given our intention to collapse the lexical decision data over word/nonword decisions.
In a similar vein, rather than use arbitrary tone mappings for the memory instructions, the Remember instruction consisted of a string of five uppercase Rs (i.e., RRRRR) and the Forget instruction consisted of a string of five uppercase Fs (i.e., FFFFF). This change likewise reduced the memory load for participants and had the added benefit of avoiding potential sound quality/volume issues across devices used for online access. A consistent mapping of letter strings to Remember/Forget instructions is common within the directed forgetting literature (e.g., Dewhurst, Anderson, Howe, & Clough, 2019; Hourihan, Ozubko, & MacLeod, 2009; MacLeod, 1989; Quinlan, Taylor, & Fawcett, 2010; van Hoof & Ford, 2011).
Informed consent was obtained before beginning the experiment and full debriefing was provided after the experiment. The text of the informed consent and debriefing forms was presented in black on a full-screen white background to distinguish the content from the experiment trials for which all stimuli were presented in white on a full-screen black background, except where otherwise indicated. Within the PsychoPy programming environment, we used the default Arial font and specified font height as a proportion of the vertical screen extent, such that the size of stimuli remained constant relative to the size of the monitors used to view the stimuli online. Text on study trials was presented at a height of 0.055 and test items on the recognition trials were presented at a height of 0.1. The stimulus boxes on study trials were drawn in a 1-point opaque white line and set to a height that was 4 times that of the study text stimuli and to a width that was 1.5 times the height; one box was centred on the computer monitor and the other two displaced 0.4 to the left and right. During recruitment, participants were instructed that that the study required an internet-connected device equipped with a physical keyboard; the 54 files that were retained for analysis identified the operating system as MacIntel for 35 participants and Win32 for the other 19 participants, all with a reported 60 Hz refresh rate.
Procedure
The procedure was identical to Experiment 4, except for the following. With the change to a letter string for the memory instruction, there was no need to present tone familiarization trials in Experiment 6. Whereas the word word was presented on every trial of the practice block in Experiments 1–5, to ensure that participants understood that a word response was required to any configuration of letters presented as a word (and not only to word), the coloured letter string in the practice block of Experiment 6 was configured as the word dog on half of the trials and as the nonword dqg on the other half.
In the study block, a different study item was presented on each trial and was as likely to be configured as a word as a nonword. Although half of the word and nonword items were coloured blue and half were coloured pink, colour was task-irrelevant; we retained the pink/blue colouring to maintain consistency with the methods used in Experiments 1–5, as well as to assist participants in distinguishing the study item from the white distractor x-string in the opposite location. In response to each coloured letter string, participants were required to make a speeded lexical decision to report whether that string was a word (“w”) or a nonword (“n”). They received feedback as described for Experiments 1–5.
Participants were instructed that the memory instruction that followed the coloured study item was relevant only to word configurations: They were not required to remember and forget nonword letter strings. This focused attention squarely on differentiating words from nonwords—not only so that participants could distinguish words from nonwords to perform the lexical decision task that was used to assess attention allocation, but so that they would continue doing so for the directed forgetting task. Our goal was to provide a strong test of whether a common focus on word processing in Experiment 6 would elicit an interaction of attention and directed forgetting.
There was a total of 128 study trials comprised of 64 word/x-string pairs and 64 nonword/x-string pairs. There were thus fewer study (and recognition) trials in Experiment 6 than in Experiment 4—a change deemed necessary to accommodate the time and attention demands associated with online data collection. Study items were distributed such that there were eight words and eight nonwords in each cell of a factorial design: Cue Location (Left, Right) × Study Item Colour (Pink, Blue) × Memory Instruction (Remember, Forget). Of the eight item pairs within each of these cells, six were presented in the location indicated by the cue and two were presented in the location opposite that indicated by the cue, such that the cue was a valid predictor of study item location on 75% of trials. This slightly reduced validity rate (75% in Experiment 6 vs. 80% in Experiments 2–5) compensated for the reduction in number of study trials while still providing incentive for participants to attend to the cued location. The x-string member of each study item pair always appeared in white in the location opposite the coloured study item. For the purpose of data analysis, this design was collapsed and reconceptualized as a 2 (Cue Condition: Cued, Uncued) × 2 (Memory Instruction: Remember, Forget).
Because the memory instruction had no meaning with respect to nonwords, the recognition task tested only words from the study trials.Footnote 3 There were thus a total of 128 recognition trials, composed of the presentation of 32 TBR words, 32 TBF words, and 64 unstudied foil words.
Results
The data from five participants were excluded: two based on high false alarms, two based on low lexical decision accuracy, and one based on both criteria. Subsequent analyses were conducted on the 49 remaining data sets.
Lexical decision RTs
The mean RTs to make a correct word/nonword decision are shown in Fig. 8a. As was true in Experiments 3–5, the central predictive cues used in Experiment 6 led to very strong evidence for an effect of Cue Condition, F(1, 48) = 75.28, MSE = 3624.81, p < .01, ges = 0.10, pH1 > .99. This was due to overall faster RTs to make a lexical decision about letter strings that appeared in a Cued location (M = 709 ms) rather than in an Uncued location (M = 784 ms), indicative of an overall cueing effect. There was weak evidence for a main effect of Memory Instruction, F(1, 48) = 4.17, MSE = 1028.29, p < .05, ges < 0.01, pH1 = 0.52, with slightly slower RTs to make a lexical decision about letter strings followed by a Forget instruction (M = 751 ms) than those followed by a Remember instruction (M = 742 ms). This suggests that the appearance of the memory instruction during execution of the lexical decision response might have had some influence on the speed of that response. Finally, there was positive evidence against an interaction of Cue Condition and Memory Instruction, F < 1, pH0 = .87.

Results of Experiment 6. a Mean correct RTs (ms) to make a lexical decision, as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). b Mean recognition “yes” responses to studied words (i.e., hits), as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). For reference, the mean recognition “yes” responses to unstudied foil words (i.e., false alarms) is depicted as a dotted line. The error bars represent Fisher’s Least Significant Difference on the two-way interaction; nonoverlapping bars can be interpreted as being significantly different
Lexical decision accuracy
Mean lexical decision accuracy data are shown in Table 1. An analysis of these data revealed strong evidence for an effect of Cue Condition, F(1, 48) = 28.21, MSE = 180.84, p < .01, ges = 0.17, pH1 > 0.99: Not only were lexical decisions faster to Cued letter strings, they were also more accurate to those strings (M = 92%) compared with Uncued strings (M = 82%). There was positive evidence against an effect of Memory Instruction, F < 1, pH0 = 0.85, and against an interaction of Cue Condition and Memory Instruction, F < 1, pH0 = 0.88.
Recognition hits
The mean percentage of recognition “yes” responses is shown in Fig. 8b as a function of Cue Condition (Cued, Uncued) and Memory Instruction (Remember, Forget). The mean false-alarm rate to unstudied foils is also depicted.
An analysis of the recognition hit rate to studied words provided weak evidence against a main effect of Cue Condition, with an average of 60% hits to study words that appeared in a Cued location and 55% to those that appeared in an Uncued location, F(1, 48) = 3.58, MSE = 310.33, p > .06, ges < .01, pH0 = 0.55. Thus, despite evidence of a cueing effect for both the speed and accuracy of making a lexical decision, there was no compelling evidence that subsequent recognition was affected by whether study words appeared in an attended rather than in an unattended location. There was positive evidence for an effect of Memory Instruction, F(1, 48) = 10.29, MSE = 330.58, p < .01, ges = 0.03, pH1 = 0.94. This confirms a directed forgetting effect, with overall better recognition of TBR words (62%) than TBF words (54%). Although the magnitude of this effect was smaller in Experiment 6 (8%) than across the pooled data for Experiments 2–5 (23%), a sign test conducted using an online calculator (https://memory.psych.mun.ca/models/stats/sign_test.shtml) revealed that of those participants who showed a nonzero difference in the overall hit rate to TBR and TBF items (47/49), a significant number (36/47) showed a difference in the expected direction of TBR > TBF, p < .01. Critically, despite having participants make a lexical decision about the study item rather than discriminate the colour of that item, the results of Experiment 6 echoed those of Experiments 2–5, with weak evidence against an interaction of Cue Condition and Memory Instruction, F(1, 48) = 3.96, MSE = 197.45, p > .05, ges < 0.01, pH0 = 0.50, arguing that the magnitude of the directed forgetting effect was not discernibly different for items at attended and unattended locations.
Discussion
The results of Experiment 6 showed a directed forgetting effect, with greater recognition of TBR words than of TBF words. The magnitude of this effect was smaller than for the preceding experiments. This was likely due to the fact that relative to the preceding experiments, Experiment 6 included fewer trials and used consistent stimulus mappings (e.g., to the lexical decision response and to the memory instruction) that were intended to reduce the cognitive load on participants. The fewer the TBR trials and the lighter the cognitive load, the less inclined participants seem to be to make the effort needed to forget, and consequently the more TBF items that tend to be remembered and the smaller the directed forgetting effect (Lee, 2012; Lee & Lee, 2011). Regardless of this smaller magnitude directed forgetting effect in Experiment 6, the overall pattern of results was otherwise unchanged from Experiments 2–5. Indeed, there was evidence of a cueing effect in the lexical decision task, with overall faster and more accurate responses made to letter strings that appeared in a cued location rather than in an uncued location. And, despite the change from a colour discrimination to a lexical decision task, there continued to be no compelling evidence that the locus of attention interacted with the directed forgetting effect, underscoring the generalizability of the findings reported for Experiments 2–5.
General discussion
When participants implement instructions to remember and forget, they do so by leveraging attentional resources. Not only do they elaborate TBR items to commit them to memory, they also expend cognitive effort (Cheng et al., 2012; Fawcett & Taylor, 2008; Fawcett, Taylor, & Nadel, 2013; although see Tan, Ensor, Hockley, Harrison, & Wilson, 2020) to actively withdraw attention (Lee, 2018; Taylor, 2005, 2018; Taylor & Fawcett, 2011; Thompson et al., 2014) from the representation of TBF items to cease unwanted item processing (Hourihan & Taylor, 2006). To the extent that successful control over long-term memory encoding thus depends on regulating attentional resources, it stands to reason that regulating attentional resources prior to study word onset might likewise influence the success of exerting subsequent intentional control over encoding. There is no requirement that a bidirectional relation exist, but the possibility of such a relation is worth testing. Doing so was the goal of the current investigation.
On the grounds that attention enhances processing of items within its focus (e.g., Hawkins et al., 1990; Luck, 1995; Mangun, 1995), we reasoned that study words that appear in the spotlight of attention might be better remembered than those that appear outside of its central beam and that this might interact with memory intentions to enhance efforts to intentionally remember or—more likely—interfere with efforts to intentionally forget. This is not, however, what we found. Across all six experiments, only the intention to remember or to forget influenced later recognition of study words: Whether a study word appeared in an attended or in an unattended location had no obvious bearing. This was true despite the fact that our cues proved effective for allocating attentional resources ahead of study word onset—at least when participants had incentive to use those cues to anticipate the study word location (Experiments 2–6). Indeed, so long as the cue was given predictive validity for the study word location, we obtained evidence of cueing effects for both peripheral onset cues (Experiment 2) and central symbolic cues (Experiments 3–5). And these cueing effects occurred whether they were measured by RTs to study items that appeared alone in a peripheral location (Experiments 2 and 3); to study items that appeared simultaneously with equal-length distractor strings in the opposite location (Experiments 4 and 6); or to study items that appeared simultaneously with equal-length distractor words in the opposite location (Experiment 5). Moreover, these cueing effects occurred whether they were measured using a perceptual colour discrimination task (Experiments 2–5) or a lexical decision task (Experiment 6). There was thus ample evidence that participants used the cues to direct attention to the cued location and that they did so in advance of the study word onset. Nevertheless, the evidence suggested that intentional encoding control was unaffected for both TBR and TBF items (Experiments 2–6); incidental encoding of distractor words (Experiment 5) was also unaffected.
Attention versus intention
There are two principal outcomes of our study. First, as far as we are aware, our findings are the first to show that the successful allocation of attention to a study item does not, in and of itself, increase the likelihood of memory formation (either intentional or incidental) and/or decrease the likelihood of forgetting. This finding is quite interesting because a tight link between attentional processing and item memorability is often assumed.
Arguably, we might have observed an effect of attention on memory if we had greatly shortened the presentation time of our study items to bring them nearer the sensory-perceptual threshold. The cueing effect was 70 ms when pooled across Experiments 2–5 and was 75 ms in Experiment 6: With a much shorter word presentation time, it is possible that this 70–75-ms advantage to cued items would be sufficient to mediate subsequent efforts to control encoding, whereas it had little impact in the context of the 400 ms available to process the study word before presentation of the memory instruction in the current study. Indeed, when the study word presentation time is doubled from 1 s to 2 s (Hockley et al., 2016) and as a Forget instruction is increasingly delayed from 0–5 s after a default instruction to encode the item (e.g., Hourihan & Taylor, 2006), participants become less likely to successfully exclude unwanted TBF items from long-term memory. Although such changes in processing are unrelated to the prior allocation of attention, they nevertheless suggest that increased processing time can impair efforts to intentionally forget under some conditions.
That said, however, greatly reducing our study item presentation times in an effort to discern an effect of increased processing time due to the prior allocation of attention would have made our results less generalizable to the wider directed forgetting literature. And, more importantly, doing so would have made it impossible to know whether recognition memory is vulnerable to the effects of top-down attentional allocation per se or merely sensitive to the quality of the bottom-up input (see also Taylor & Ivanoff, 2020). Accordingly, the second principal outcome of our study is that it establishes the predominant direction of the relation between memory intentions and attentional allocation in an item-method directed forgetting task. To wit: The intention to remember and forget appears to influence the allocation of attentional resources (Fawcett & Taylor, 2010; Taylor, 2005, 2018; Taylor & Fawcett, 2011; Thompson et al., 2014) and can interact with available processing time to influence the likelihood of successful forgetting (e.g., Hockley et al., 2016; Hourihan & Taylor, 2006); however, it appears that the prior allocation of attentional resources is not sufficient—in and of itself—to influence directed forgetting of supra-threshold stimuli.
The “knew-it-all-along” effect (Slovic & Fischhoff, 1977; Wood, 1978) might lead one to conclude that our current findings are self-evident. We would argue that they are not. Attention operates in the service of memory: As an upstream cognitive function, the formation of a memory representation necessarily depends on the quality of initial perceptual processing. With no other explicit instructions, it seems likely that the probability that an item is encoded into memory, and the quality of the long-term representation that is formed would both be influenced by whether the item appeared in an attended or in an unattended location. Our results, however, demonstrate that any processing advantage for items in attended compared with unattended locations (e.g., as revealed by faster colour discrimination and lexical decision RTs) does not afford an additional memorial advantage. When participants need to exert control over long-term memory encoding, they appear to marshal attentional resources to engage rehearsal strategies in the case of a remember instruction or to eschew further processing in the case of a forget instruction; however, allocating these attentional resources in advance of the study word does not appear to mitigate the control processes that occur after the memory instruction.
We do not claim that attention never matters for memory formation. Indeed, there is convincing evidence that the distraction of attentional resources during encoding interferes with explicit memory retrieval (e.g., Craik, Govoni, Naveh-Benjamin, & Anderson, 1996; Fernandes & Moscovitch, 2000; Mulligan, 1998). It turns out that initial item processing and registration is particularly vulnerable to the effects of distraction (Naveh-Benjamin, Guez, & Sorek, 2007); explicit recognition requires at least some attention to be paid to studied items during presentation (e.g., Szymanski & MacLeod, 1996). We claim only that for supra-threshold items that receive sufficient attention to be consciously apprehended, the intention to remember or forget in an item-method directed forgetting task is likely a stronger determinant of subsequent recognition performance than is the availability of additional attentional resources for initial item processing.
Notes
We extinguished the fixation stimulus during cue presentation in order to equate—as far as possible—the stimulus events across experiments (the central cues used in Experiments 3–6 required removal of the fixation stimulus).
We recruited a larger sample size in Experiment 5 than in Experiment 4 because only half as many TBR and TBF study items were tested in Experiment 5 (see Method). Our goal was to collect data from a sample of 60 participants. However, uncertainty over how many end-of-term participants might fail to show up (a common occurrence) meant that more participants were scheduled to participate than were actually needed. As it happened, the majority of participants who signed up showed up, resulting in a larger sample size than targeted. No data were analysed before the end of the last scheduled participant; all collected data were included in the reported sample.
We contemplated assessing recognition of studied nonwords as a test for incidental memory formation on trials with Remember and Forget instructions. However, we were concerned that forcing recognition of nonword letter strings would be so difficult that it would risk frustrating our participants and leading them to “give up” on trying to give considered responses to each recognition test item. A reliable measure of intentional forgetting and remembering seemed more critical than also obtaining a measure of incidental memory for nonword letter strings.
References
Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55(5), 485–496.
Bancroft, T. D., Hockley, W. E., & Farquhar, R. (2013). The longer we have to forget the more we remember: The ironic effect of postcue duration in item-based directed forgetting. Journal of Experimental Psychology: Learning, Memory, and Cognition, 39(3), 691–699.
Basden, B. H. (1996). Directed forgetting: Further comparisons of the item and list methods. Memory, 4(6), 633–654.
Basden, B. H., Basden, D. R., & Gargano, G. J. (1993). Directed forgetting in implicit and explicit memory tests: A comparison of methods. Journal of Experimental Psychology: Learning, Memory, and Cognition, 19(3), 603–616.
Berger, A., Henik, A., & Rafal, R. (2005). Competition between endogenous and exogenous orienting of visual attention. Journal of Experimental Psychology: General, 134(2), 207–221.
Bjork, R. A. (1989). Retrieval inhibition as an adaptive mechanism in human memory. In H. L. Roediger III & F. I. M. Craik (Eds.), Varieties of memory and consciousness (pp. 309–330). Hillsdale, NJ: Erlbaum.
Carrasco, M. (2011). Visual attention: The past 25 years. Vision Research, 51(13), 1484–1525.
Cheal, M., & Lyon, D. R. (1991). Central and peripheral precuing of forced-choice discrimination. The Quarterly Journal of Experimental Psychology, 43(4), 859–880.
Cheng, S. K., Liu, I. C., Lee, J. R., Hung, D. L., & Tzeng, O. J. L. (2012). Intentional forgetting might be more effortful than remembering: An ERP study of item-method directed forgetting. Biological Psychology, 89(2), 283–292. doi:https://doi.org/10.1016/j.biopsycho.2011.10.019
Chica, A. B., Lupiánez, J., & Bartolomeo, P. (2006). Dissociating inhibition of return from endogenous orienting of spatial attention: Evidence from detection and discrimination tasks. Cognitive Neuropsychology, 23(7), 1015–1034.
Chica, A. B., Bartolomeo, P., & Lupiáñez, J. (2013). Two cognitive and neural systems for endogenous and exogenous spatial attention. Behavioural Brain Research, 237, 107–123.
Cohen, J., & Cohen, P. (1983). Applied multiple regression/correlation analysis for the behavioral sciences (2nd ed.). Hillsdale: Erlbaum.
Cohen J. D., MacWhinney B., Flatt M., & Provost J. (1993). PsyScope: A new graphic interactive environment for designing psychology experiments. Behavioral Research Methods, Instruments, and Computers, 25, 257–271.
Coltheart, M. (1981) The MRC Psycholinguistic Database. Quarterly Journal of Experimental Psychology, 33A, 497–505.
Craik, F. I., Govoni, R., Naveh-Benjamin, M., & Anderson, N. D. (1996). The effects of divided attention on encoding and retrieval processes in human memory. Journal of Experimental Psychology: General, 125(2), 159.
Dewhurst, S. A., Anderson, R. J., Howe, D., & Clough, P. J. (2019). The relationship between mental toughness and cognitive control: Evidence from the item-method directed forgetting task. Applied Cognitive Psychology, 33(5), 943–951.
Fawcett, J. M., & Taylor, T. L. (2008). Forgetting is effortful: Evidence from reaction time probes in an item-method directed forgetting task. Memory & Cognition, 36(6), 1168–1181.
Fawcett, J. M., & Taylor, T. L. (2010). Directed forgetting shares mechanisms with attentional withdrawal but not with stop-signal inhibition. Memory & Cognition, 38(6), 797–808.
Fawcett, J. M., & Taylor, T. L. (2012). The control of working memory resources in intentional forgetting: Evidence from incidental probe word recognition. Acta Psychologica, 139(1), 84–90.
Fawcett, J. M., Taylor, T. L., & Nadel, L. (2013). Event-method directed forgetting: Forgetting a video segment is more effortful than remembering it. Acta Psychologica, 144(2), 332–343.
Fellner, M.-C., Waldhauser, G. T., & Axmacher, N. (2019). Tracking selective rehearsal and active inhibition of memory traces in directed forgetting. bioRxiv, 864819. https://doi.org/10.1101/864819
Fernandes, M. A., & Moscovitch, M. (2000). Divided attention and memory: Evidence of substantial interference effects at retrieval and encoding. Journal of Experimental Psychology: General, 129(2), 155–176.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18(4), 1030–1044.
Gardiner, J. M., Gawlik, B., & Richardson-Klavehn, A. (1994). Maintenance rehearsal affects knowing, not remembering: Elaborative rehearsal affects remembering, not knowing. Psychonomic Bulletin & Review, 1(1), 107–110.
Gottsdanker, R., & Shragg, G. P. (1985). Verification of Donders’ subtraction method. Journal of Experimental Psychology: Human Perception and Performance, 11(6), 765–776.
Hawkins, H. L., Hillyard, S. A., Luck, S. J., Mouloua, M., Downing, C. J., & Woodward, D. P. (1990). Visual attention modulates signal detectability. Journal of Experimental Psychology: Human Perception and Performance, 16(4), 802–811.
Hockley, W. E., Ahmad, F. N., & Nicholson, R. (2016). Intentional and incidental encoding of item and associative information in the directed forgetting procedure. Memory & Cognition, 44(2), 220–228.
Hourihan, K. L., Ozubko, J. D., & MacLeod, C. M. (2009). Directed forgetting of visual symbols: Evidence for nonverbal selective rehearsal. Memory & Cognition, 37(8), 1059–1068.
Hourihan, K. L., & Taylor, T. L. (2006). Cease remembering: control processes in directed forgetting. Journal of Experimental Psychology: Human Perception and Performance, 32(6), 1354–1365.
Hsieh, L. T., Hung, D. L., Tzeng, O. J. L., Lee, J. R., & Cheng, S. K. (2009). An event-related potential investigation of the processing of remember/forget cues and item encoding in item-method directed forgetting. Brain Research, 1250, 190–201.
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eye’s movement. Attention and Performance IX (pp. 187–203). Hillsdale: Erlbaum.
Jonides, J., & Yantis, S. (1988). Uniqueness of abrupt visual onset in capturing attention. Perception & Psychophysics, 43(4), 346–354.
Lawrence, M. A. (2016). ez: Easy analysis and visualization of factorial experiments (R package Version 4.4-0) [Computer software]. Retrieved from https://CRAN.R-project.org/package=ez
Leber, A. B., & Egeth, H. E. (2006). It’s under control: Top-down search strategies can override attentional capture. Psychonomic Bulletin & Review, 13(1), 132–138.
Lee, Y. S. (2012). Cognitive load hypothesis of item-method directed forgetting. The Quarterly Journal of Experimental Psychology, 65(6), 1110–1122.
Lee, Y. S. (2018). Withdrawal of spatial overt attention following intentional forgetting: evidence from eye movements. Memory, 26(4), 503–513.
Lee, Y. S., & Lee, H. M. (2011). Divided attention facilitates intentional forgetting: Evidence from item-method directed forgetting. Consciousness and Cognition, 20(3), 618–626.
Luck, S. J. (1995). Multiple mechanisms of visual-spatial attention: Recent evidence from human electrophysiology. Behavioural Brain Research, 71(1/2), 113–123.
MacLeod, C. M. (1989). Directed forgetting affects both direct and indirect tests of memory. Learning, Memory, & Cognition, 15(1), 13–21.
MacLeod, C. M. (1998). Directed forgetting. In J. M. Golding & C. M. MacLeod (Eds.), Intentional forgetting: Interdisciplinary approaches (pp. 1–57). Mahwah: Erlbaum.
Mangun, G. R. (1995). Neural mechanisms of visual selective attention. Psychophysiology, 32(1), 4–18.
Masson, M. E. (2011). A tutorial on a practical Bayesian alternative to null-hypothesis significance testing. Behavior Research Methods, 43(3), 679–690.
Montagliani, A., & Hockley, W. E. (2019). Item-based directed forgetting for categorized lists: Forgetting of words that were not presented. Canadian Journal of Experimental Psychology, 73(3), 135–143.
Müller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary orienting of visual attention: time course of activation and resistance to interruption. Journal of Experimental Psychology: Human Perception and Performance, 15(2), 315–330.
Mulligan, N. W. (1998). The role of attention during encoding in implicit and explicit memory. Journal of Experimental Psychology: Learning, Memory, and Cognition, 24(1), 27–47.
Naveh-Benjamin, M., Guez, J., & Sorek, S. (2007). The effects of divided attention on encoding processes in memory: Mapping the locus of interference. Canadian Journal of Experimental Psychology, 61(1), 1–12.
Peirce, J., Gray, J. R., Simpson, S., MacAskill, M.., Höchenberger, R., Sogo, H., Kastman, E., & Lindeløv, J. K. (2019). PsychoPy2: Experiments in behavior made easy. Behavior Research Methods, 51, 195–203.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology, 32(1), 3–25.
Posner, M.I. (1988). Structures and functions of selective attention. In T. Boll & B. Bryant (Eds.), Clinical neuropsychology and brain function: Research, measurement and practice (pp. 169–202). Washington: American Psychological Association.
Posner, M. I., Walker, J. A., Friedrich, F. J., & Rafal, R. D. (1984). Effects of parietal injury on covert orienting of attention. Journal of Neuroscience, 4, 1863–1874.
Quinlan, C. K., Taylor, T. L., & Fawcett, J. M. (2010). Directed forgetting: Comparing pictures and words. Canadian Journal of Experimental Psychology, 64(1), 41–46.
R Core Team. (2019). R: A language and environment for statistical computing [Computer software]. Vienna: R Foundation for Statistical Computing. Retrieved from https://www.R-project.org/
Raftery, A. E. (1995). Bayesian model selection in social research. Sociological Methodology, 25, 111–164.
Rizio, A. A., & Dennis, N. A. (2013). The neural correlates of cognitive control: Successful remembering and intentional forgetting. Journal of Cognitive Neuroscience, 25(2), 297–312.
Rubinfeld, L. M., Taylor, T. L., & Hamm, J. P. (2019). Selection for encoding: No evidence of better endogenous orienting following forget than following remember instructions. Attention, Perception, & Psychophysics, 81(1), 237–252.
Slovic, P., & Fischhoff, B. (1977). On the psychology of experimental surprises. Journal of Experimental Psychology: Human Perception and Performance, 3(4), 544–551.
Szymanski, K. F., & MacLeod, C. M. (1996). Manipulation of attention at study affects an explicit but not an implicit test of memory. Consciousness and Cognition, 5(1/2), 165–175.
Tan, P., Ensor, T. M., Hockley, W. E., Harrison, G. W., & Wilson, D. E. (2020). In support of selective rehearsal: Double-item presentation in item-method directed forgetting. Psychonomic Bulletin & Review, 27, 529–535.
Taylor, T. L. (2005). Inhibition of return following instructions to remember and forget. The Quarterly Journal of Experimental Psychology, 58(4), 613–629.
Taylor, T. L. (2018). Remember to blink: Reduced attentional blink following instructions to forget. Attention, Perception, & Psychophysics, 80(6), 1489–1503.
Taylor, T. L., & Fawcett, J. M. (2011). Larger IOR effects following forget than following remember instructions depend on exogenous attentional withdrawal and target localization. Attention, Perception, & Psychophysics, 73(6), 1790–1814.
Taylor, T. L., & Hamm, J. P. (2016). Selection for encoding: No evidence of greater attentional capture following forget than remember instructions. Attention, Perception, & Psychophysics, 78(1), 168–186.
Taylor, T. L., & Ivanoff, J. (2020). Forgetting under difficult conditions: Item-method directed forgetting following data-limited processing. Manuscript submitted for publication.
Theeuwes, J. (1994). Stimulus-driven capture and attentional set: Selective search for color and visual abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 20(4), 799–806.
Theeuwes, J. (2004). Top-down search strategies cannot override attentional capture. Psychonomic Bulletin & Review, 11, 65–70.
Thompson, K. M., Hamm, J. P., & Taylor, T. L. (2014). Effects of memory instruction on attention and information processing: Further investigation of inhibition of return in item-method directed forgetting. Attention, Perception, & Psychophysics, 76(2), 322–334.
Timmins, W. K. (1974). Varying processing time in directed forgetting. Journal of Verbal Learning and Verbal Behavior, 13(5), 539–544.
van Hoof, J. C., & Ford, R. M. (2011). Remember to forget: ERP evidence for inhibition in an item-method directed forgetting paradigm. Brain Research, 1392, 80–92.
Wickham, H. (2011). The split-apply-combine strategy for data analysis. Journal of Statistical Software, 40(1), 1–29. Retrieved from http://www.jstatsoft.org/v40/i01/
Wickham, H. (2016). ggplot2: Elegant graphics for data analysis. New York: Springer. Retrieved from https://ggplot2.tidyverse.org
Wickham, H. (2019). stringr: Simple, consistent wrappers for common string operations (R Package Version 1.4.0) [Computer software]. Retrieved from https://CRAN.R-project.org/package=stringr
Wickham, H. & Henry, L. (2019). tidyr: Tidy messy data. R package version 1.0.0. https://CRAN.R-project.org/package=tidyr
Wickham, H., Francois, R., Henry, H., & Müller, K. (2019). dplyr: A grammar of data manipulation (R Package Version 0.8.3) [Computer software]. Retrieved from https://CRAN.R-project.org/package=dplyr
Wilson, M. D. (1988). The MRC Psycholinguistic Database: Machine readable dictionary, Version 2. Behavioural Research Methods, Instruments and Computers, 20, 6–11.
Wood, G. (1978). The knew-it-all-along effect. Journal of Experimental Psychology: Human Perception and Performance, 4(2), 345–353.
Wylie, G. R., Foxe, J. J., & Taylor, T. L. (2008). Forgetting as an active process: An fMRI investigation of item-method–directed forgetting. Cerebral Cortex, 18(3), 670–682.
Yantis, S., & Jonides, J. (1984). Abrupt visual onsets and selective attention: Evidence from visual search. Journal of Experimental Psychology: Human Perception and Performance, 10, 601–621.
Acknowledgement
Thanks to Dr. Jonathan Fawcett for providing the custom software that was used to randomize words to study lists in Experiments 1–5, and to Caitlin Joudrey for collecting the data for Experiments 1–5. Thanks also to participants for volunteering their time to contribute to this research. Project funding was provided by a Discovery Grant awarded to T.L.T. by the Natural Sciences and Engineering Research Council of Canada.
Open practices statement
The data for these experiments are available from the first author upon request. None of the experiments was preregistered.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Taylor, T.L., Hamm, J.P. Intention matters more than attention: Item-method directed forgetting of items at attended and unattended locations. Atten Percept Psychophys 83, 1629–1651 (2021). https://doi.org/10.3758/s13414-020-02220-x
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-020-02220-x