
Abstract
Many studies have shown that covert visual attention precedes saccadic eye movements to locations in space. The present research investigated whether the allocation of attention is similarly affected by eye blinks. Subjects completed a partial-report task under blink and no-blink conditions. Experiment 1 showed that blinking facilitated report of the bottom row of the stimulus array: Accuracy for the bottom row increased and mislocation errors decreased under blink, as compared with no-blink, conditions, indicating that blinking influenced the allocation of visual attention. Experiment 2 showed that this was true even when subjects were biased to attend elsewhere. These results indicate that attention moves downward before a blink in an involuntary fashion. The eyes also move downward during blinks, so attention may precede blink-induced eye movements just as it precedes saccades and other types of eye movements.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Eye blinks occur about 15 times a minute, for a variety of reasons: Some are reflexive, generated in response to environmental stimuli such as a loud noise; others are endogenous, reflecting a person's emotional or cognitive state; and blinks can also be made voluntarily—in response to a command, for example (Stern, Walrath, & Goldstein, 1984).
The physical characteristics of these different kinds of eye blinks differ somewhat, with reflexive and voluntary blinks having shorter and less variable durations than do endogenous eye blinks (VanderWerf, Brassinga, Reits, Aramideh, & Ongerboer de Visser, 2003). All eye blinks are produced in the same way, however, by antagonistic interactions between two skeletal eyelid muscles, the levator palpebrae superioris (LPS) and the orbicularis oculi (OO) muscles (Evinger, Manning, & Sibony, 1991). Immediately before a blink, LPS motoneurons briefly stop firing while OO motoneurons fire briefly, causing a rapid lowering of the upper eyelid. LPS motoneurons resume firing, and OO motoneurons stop firing to return the upper eyelid to its original position (Bour, Aramideh, & Ongerboer de Visser, 2000). The eyes themselves also move somewhat during an eye blink (e.g., Bour et al., 2000; Collewijn, Van Der Steen, & Steinman, 1985; Evinger, Shaw, Peck, Manning, & Baker, 1984). Collewijn et al. reported that during voluntary and reflexive eye blinks, both eyes consistently move down and toward the nose, with amplitudes in the range of 1°–5°. The duration and velocity of these eye movements indicate that they are not saccadic eye movements; rather, they seem to occur as a result of the eyeballs being retracted during the eye blink to ease the passage of the eyelids (Bour et al., 2000; Collewijn et al., 1985; Evinger et al., 1984).
Although the physical characteristics of different kinds of eye blinks vary somewhat, in all cases vision is almost completely blocked by the closed eyelids for approximately 100–150 ms (Riggs, Volkmann, & Moore, 1981). Although dimming the lights in a room for this duration is very noticeable, the visual interruptions caused by eye blinks are hardly perceptible (Volkmann, Riggs, & Moore, 1980). Volkmann et al. (1980) demonstrated that this is because vision is suppressed during a blink (and, to a lesser extent, before and after it as well), so that the visual interruption is not perceived. This suppression occurs for reflexive eye blinks as well as for voluntary ones (Manning, Riggs, & Komenda, 1983).
The purpose of the present research was to investigate what happens to visual attention when one programs and executes an eye blink. Since vision is disrupted, does visual attention simply disengage from where it was located before the blink and await reallocation after the eyes reopen? Alternatively, does it stay located wherever it was before the eye blink, ensuring continuity of attentional focus across the eye blink? Or might attention actually move somewhere specific in response to the eye blink’s being programmed and executed, in much the same way that attention moves to the saccade target location before a saccadic eye movement (Deubel & Schneider, 1996; Henderson, Pollatsek, & Rayner, 1989; Hoffman & Subramaniam, 1995; Irwin & Gordon, 1998; Klein, 1980; Klein & Pontefract, 1994; Kowler, Anderson, Dosher, & Blaser, 1995; Rayner, McConkie, & Ehrlich, 1978; Shepherd, Findlay, & Hockey, 1986)?
Casual observation suggests that attention does not stay focused on the same spatial location across an eye blink. If one happens to blink while a computer display is updated during a screen refresh, for example, the fact that the postblink visual display is different from the preblink visual display is not always immediately apparent. O’Regan, Deubel, Clark, and Rensink (2000) demonstrated this experimentally by using a change detection procedure in which a picture displayed on a computer screen changed in some fashion (e.g., an object appeared or disappeared, changed color, or changed position) during an eye blink. Subjects were instructed to look for changes but were not told that the changes occurred during eye blinks. O’Regan et al. found that changes were frequently undetected, even when they occurred at the exact location that the participant was fixating before and after the blink. O’Regan et al. called this “looking without seeing” and suggested that the failure to detect changes that occur during a blink might be due to failures of either memory or attention. Higgins, Irwin, Wang, and Thomas (2009) similarly found that eye blinks interfered with the perception of object displacement, suggesting that the locus of attention is not fixed across a blink. Interestingly, they found that blinks interfered with the perception of object displacement even when the displacement took place before the eye blink was initiated, indicating that blink planning and programming alone is sufficient to cause visual attention to disengage from its locus.
Thomas and Irwin (2006) also examined the effects of eye blinks on visual memory and attention. They had subjects perform a Sperling (1960) partial-report task under blink and no-blink conditions to determine whether eye blinks might interfere with early sensory memory and/or visual short-term memory. Subjects were presented with a 3 × 3 array of letters for approximately 100 ms. One of three tones was presented 50, 150, or 750 ms after array offset, signaling report of one of the three rows in the array. In some blocks of trials, subjects performed this task while keeping their eyes open, whereas in others, they were instructed to blink as soon as the letters were presented. People do not blink their eyes instantaneously, of course. The average latency for blinks in this experiment was 225 ms, while the duration of the letter array was 106 ms, so, on average, the blink was initiated 119 ms after the offset of the letter display. In other words, blinking did not interfere with perception of the letters per se. Blinks did interfere with memory for the letters, however, but only at the shortest cue delay. Accuracy was significantly lower at the 50-ms cue delay under blink, as compared with no-blink, conditions, indicating that eye blinks interfere with iconic memory. The decrease in accuracy caused by an eye blink was accompanied by an increase in mislocation errors; that is, when subjects made a mistake, they reported a letter that had appeared somewhere in the array, but not in the indicated position. It is generally believed that attention is required to bind identity and position information together (e.g., Treisman & Gelade, 1980), so the fact that blinks increased mislocation errors suggests that blinks interfere with the allocation of attention. To elaborate, in order to perform a partial-report task accurately, one must direct attention to the cued row to read out the information from iconic memory. If blinks affect the allocation of visual attention, it may be the case that attention cannot be directed to the cued row, leading to a decrease in accuracy and an increase in mislocation errors.
The results of the studies summarized above indicate that eye blinks influence the allocation of visual attention. It seems clear that attention does not stay focused at the same spatial location across an eye blink. What is less clear (because it has not been investigated) is where attention goes when a blink is programmed and executed. Does it simply disengage, or might it move somewhere specific when a blink is programmed, just as it does when a saccade is programmed? A recent reanalysis of the data from Thomas and Irwin (2006) suggests that attention does not simply disengage before a blink is executed but, rather, moves in a downward direction. That is, although Thomas and Irwin found that blinks lowered accuracy and increased mislocation errors (relative to a no-blink condition) at a short cue delay in a Sperling partial-report task, an additional analysis conducted after their study was published revealed that this was true only when the top or middle row of the array was cued. When the bottom row was cued, accuracy was equally high under blink and no-blink conditions. This is consistent with attention moving downward before a blink, thereby facilitating the encoding of the letters in the bottom row of the array. Although suggestive, this analysis is post hoc, and the 3 × 3 format of the displays centered at fixation used by Thomas and Irwin is not ideal for investigating possible movements of visual attention. Thus, the present study investigated directly the hypothesis that attention moves downward before a blink. Accuracy and error patterns in a partial-report task were measured under blink and no-blink conditions, using a more spatially distributed array than that used by Thomas and Irwin.
Experiment 1
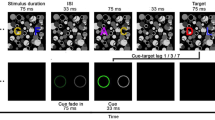
In Experiment 1, subjects were briefly shown a display containing four groups of letters, three letters per group. The letter groups were arranged around the center of the display in a diamond pattern, centered 3° above, below, to the left, and to the right of center (see Fig. 1). This letter display was presented for 106 ms and then was followed immediately by an arrow in the center of the display that pointed up, down, left, or right. Subjects were instructed to report the group of letters indicated by the arrow. On some trials, subjects were told to blink as soon as they saw the letter display (blink condition), while on other trials, they were told to keep their eyes open and fixated (no-blink condition). The latency of voluntary blinks exceeded the duration of the letter display by over 100 ms, so the blink did not prevent subjects from seeing the letters; rather, the blink was being programmed while the letters were displayed on the screen. If programming an eye blink causes a general disengagement of visual attention, accuracy should be higher for no-blink than for blink conditions, regardless of which letter group is probed. In contrast, if programming an eye blink causes attention to move in a downward direction, subjects should be more accurate in reporting the bottom group of the letter array under blink than under no-blink conditions.

Procedure for Experiment 1. Following drift correction, a blank screen was presented for 500 ms. Then the letter display was presented for 106 ms, followed immediately by an arrow cue with a duration of 47 ms. A blank screen was then presented for 1,000 ms before a response prompt appeared. On blink trials, the blink was executed during this blank period (on average, 114 ms after arrow cue offset), whereas on no-blink trials, the eyes remained open
Subjects
Thirty-nine students from the University of Illinois community were recruited for this experiment. The data from 3 subjects were deleted from analysis because they failed to follow the blink instructions. Subjects reported normal or corrected-to-normal vision and were naïve as to the purpose of the experiment. They received payment for participating in a single 50-min session.
Stimuli
The stimuli consisted of randomly generated arrays of capital letters (consonants only). Each letter display contained four groups of letters, three letters per group. Each letter subtended 1° × 1°, as did the partial-report cue (the arrow). The letter groups were arranged around the center of the display in a diamond pattern, centered 3° above, below, to the left, and to the right of center.
Apparatus
Stimuli were presented on a 21-in. monitor with a resolution of 800 × 600 pixels and a refresh rate of 85 Hz. Eye movements and blinks were recorded with an Eyelink II video-based eyetracker (SR Research Ltd., Mississauga, Ontario, Canada) with a temporal resolution of 500 Hz, a spatial resolution of 0.1°, and a pupil-size resolution of 0.1% of pupil diameter. The output of the eyetracker was parsed online by Eyelink II analysis software. Each data sample from the eyetracker contained a time stamp in milliseconds, the velocity and the position of the eye, and the area of the pupil. The parser computed instantaneous velocity and acceleration for each sample and compared these with velocity and acceleration thresholds for saccade detection. A change in eye position was classified as a saccade when its distance exceeded 0.2° and its velocity reached 30°/s or when its distance exceeded 0.2° and its acceleration reached 9,500°/s2.
The eyetracker software defined an eye blink as a period of missing pupil for at least 6 consecutive milliseconds that was preceded and followed by a period of artifactual saccade caused by the motion of the eyelids across the pupil. Blink onset and blink offset were defined to correspond to the beginning and ending of this artifactual saccade. Fixations were defined as any period that was not a blink or a saccade. Custom C code was written to display stimuli and collect responses. Subjects’ heads were stabilized with a chinrest, fixed at 48 cm from the computer monitor. The height of the chair that subjects sat in was adjusted for each individual so that their eyes were centered with respect to the display monitor. The display background was light gray (luminance = 86.3 cd/m2), and all stimuli were black (luminance = 6.5 cd/m2). Subjects entered their partial-report responses using a standard computer keyboard.
Each experimental block of trials began with a nine-position calibration procedure in which the edges and center of the screen were fixated; a drift correction procedure was completed at the beginning of each experimental trial. The circular calibration/drift correction dot subtended 0.6°.
Procedure
Each trial began with a drift correction procedure in which subjects fixated a dot in the center of the display and pressed the return key. Following successful drift correction, a blank screen was presented for 500 ms in order to prevent visual masking by the drift correction dot. After this delay, the letter display was presented for 106 ms, followed 0 ms later by the arrow cue with a duration of 47 ms. One second after the arrow cue disappeared, a response prompt appeared, and subjects were asked to type the three letters that had appeared in the cued group. There was no time limit on responding. Subjects pressed the return key to enter their response, and feedback was provided in the center of the display for 1,000 ms, indicating correct responses with a “+” and incorrect responses with a “−.” The correct letter had to be reported in the correct position in order for the response to be considered correct. The next trial began after this feedback was provided.
In the no-blink condition, subjects kept their eyes open and steady until the response prompt appeared. Trials in the blink condition were identical to trials in the no-blink condition, with one exception: Subjects were instructed to perform a voluntary eye blink as soon as the letter display appeared. Trials on which the participant blinked before the partial-report cue was extinguished (i.e., blinks with latencies of less than 154 ms) were deleted from analysis; this occurred on 7% of the blink trials. The average latency of voluntary blinks in this experiment was 267 ms (SE = 15 ms), and the average blink duration was 263 ms (SE = 33 ms). Because the duration of the letter array was 106 ms, the blink was initiated 161 ms, on average, after the offset of the letter display.
Design
Subjects completed two blocks of 36 practice partial-report trials before beginning the experimental blocks. The subjects did not wear the eyetracker during the practice trials (for reasons of comfort), and no instructions about blinking were provided. After completing these practice trials, subjects were fitted with the eyetracker, and a calibration procedure was run. They then completed three blocks of 36 trials in the no-blink partial-report condition and three blocks of 36 trials in the blink condition. The ordering of conditions was counterbalanced, with half of the subjects running through the no-blink condition first and the other half running through the blink condition first. The direction of the arrow cue was sequenced randomly across trials, with each letter group being indicated 9 times in each block. The interstimulus interval (ISI) between letter offset and arrow cue onset was 0 ms. A written prompt appeared for 2,000 ms at the beginning of each block that instructed subjects whether or not to blink during the trials in that block.
Results and discussion
The primary question under investigation was whether partial-report performance would differ in the blink versus no-blink condition and whether this would interact with the location of the cued letter group. Three measures of performance on the partial-report task were examined. First, the percentages of correct responses were calculated for both the blink and no-blink conditions. A response was classified as correct if the subject reported the correct letter in the correct position in the array. Next, errors were separated into two groups. A mislocation error occurred when the participant reported a letter that had been present in the array, but not in the position that he or she indicated. An intrusion error occurred when the participant reported a letter that was not present in the array. Intrusion errors are typically thought to arise from a lack of item identity information in memory, whereas mislocation errors arise when location information is not bound to item identity information (e.g., Dick, 1969; Irwin & Yeomans, 1986; Mewhort, Campbell, Marchetti, & Campbell, 1981; Townsend, 1973). Because attention is required for feature binding, if attention movements precede eye blinks, accuracy and error patterns should vary across letter group locations. Specifically, if attention moves downward before a blink, accuracy should be higher and mislocation errors should be lower for the bottom letter-group position under blink than under no-blink conditions.
Figure 2 shows the mean percentage of correct responses, mislocation errors, and intrusion errors in the blink and no-blink conditions as a function of the position of the letter group (top, left, right, or bottom). Separate two-way ANOVAs with factors of condition (blink vs. no blink) and position were conducted on each dependent measure. The results for correct responses are considered first. Overall accuracy on blink and no-blink trials was virtually identical, yielding a nonsignificant main effect of condition, F(1, 35) = 0.004, MSE = 0.016, p > .9. The main effect of position was significant, however, F(3, 105) = 26.0, MSE = 0.018, p < .001. The error term for the position main effect was used to construct a 95% confidence interval for comparing two means; differences greater than 3.6% were significant, indicating that accuracy was significantly higher for the top position (71.5%) than for all the other positions, and accuracy for the bottom position (67.5%) was significantly higher than accuracy for the left (56.0%) and right (56.0%) positions. Of most importance is the fact that there was a significant interaction between condition and position, F(3, 105) = 2.7, MSE = 0.004, p < .05. The error term for this interaction was used to construct a 95% confidence interval for comparing two means; differences greater than 2.5% were significant. Thus, the interaction occurred because there was a significant difference in accuracy between the no-blink and blink conditions only for the bottom position, with accuracy higher under blink (69.2%) than under no-blink (65.7%) conditions. In contrast, mean accuracy for the other three array positions (top, left, and right) was lower under blink (60.6%) than under no-blink (61.7%) conditions, but this difference was not significant. In sum, the accuracy data indicate that blinking does not have an overall deleterious effect on performance, as it would if attention simply disengaged before a blink, but rather that attention moves downward before a blink, actually facilitating partial-report performance for information presented at the bottom of the stimulus display.

Mean percentages of correct responses, mislocation errors, and intrusion errors as a function of position in the stimulus display for no-blink (NB) and blink (B) trials in Experiment 1. Error bars represent standard errors of the means
In order to provide additional support for this conclusion, the types of errors that occurred in the blink and no-blink conditions were examined. Separate two-way ANOVAs with factors of condition and position were conducted on the mean percentages of mislocation errors and intrusion errors. Examination of Fig. 2 shows that subjects were much more likely to make mislocation errors than intrusion errors in either condition, consistent with many previous studies of partial-report performance (e.g., Dick, 1969; Irwin & Yeomans, 1986; Mewhort et al., 1981; Townsend, 1973). There were no differences in the percentage of intrusion errors between blink and no-blink conditions, F(1, 35) = 1.48, MSE = 0.002, p > .2, nor did condition and position interact, F(3, 105) = 0.83, MSE = 0.001, p > .4. The main effect of position was significant, however, F(3, 105) = 11.21, MSE = 0.002, p < .001. A 95% confidence interval based on the error term for this main effect showed that differences greater than 1.2% were significant. Thus, there were fewer intrusion errors for the top (7.3%) and bottom (8.1%) positions than for the left (11.1%) and right (10.1%) positions.
Turning to the mislocation errors, the main effect of condition was not significant, F(1, 35) = 0.019, MSE = 0.010, p > .6. The main effect of position was significant, however, F(3, 105) = 24.7, MSE = 0.012, p < .001. A 95% confidence interval based on the error term for this effect showed that differences greater than 2.9% were significant. Thus, there were significantly fewer mislocation errors for the top position (21.2%) than for all the other positions and significantly fewer mislocation errors for the bottom position (24.5%) than for the left (33.3%) and right (33.7%) positions. Most important, there was a significant interaction between condition and position, F(3, 105) = 2.84, MSE = 0.004, p < .05. The error term for this interaction was used to construct a 95% confidence interval for comparing two means; differences greater than 2.3% were significant. Thus, the interaction occurred because there was a significant difference in mislocation errrors between the no-blink and blink conditions only for the bottom position, with fewer errors under blink (23.1%) than under no-blink (25.9%) conditions. The fact that mislocation errors for the bottom position were lower in the blink condition than in the no-blink condition suggests that attention moves downward before a blink, thereby facilitating the binding of object identity with object position in iconic memory.
An additional analysis of interest concerns eye position following the eye blink. After the eye blink, the horizontal position of the eye was 0.7° left of center when the top array location was cued, 1.4° left of center when the left location was cued, at the center when the right location was cued, and 0.6° left of center when the bottom location was cued. The effect of array location was significant in a one-way ANOVA, F(3, 105) = 20.5, MSE = 178.2, p < .001. The error term for this effect was used to construct a 95% confidence interval for comparing two means; differences greater than 0.29° were significant, indicating that the eyes deviated to the left significantly more when the left location was cued than when the top or bottom locations were cued and significantly more to the right (relatively speaking) when the right location was cued. Similar results were found for vertical eye position. After the eye blink, the vertical position of the eye was 0.1° below center when the top location was cued, 0.4° below center when the left location was cued, 0.4° below center when the right location was cued, and 0.9° below center when the bottom location was cued. The effect of array location was significant in a one-way ANOVA, F(3, 105) = 10.9, MSE = 136.2, p < .001. The error term for this effect was used to construct a 95% confidence interval for comparing two means; differences greater than 0.25° were significant, indicating that the eyes ended up significantly higher when the top location was cued than when the left or right locations were cued and significantly lower when the bottom location was cued.
Could these variations in the position of the eyes after the eye blink account for the accuracy and error data reported above? For example, is it possible that accuracy was higher for the bottom row of the letter array under blink conditions because the eyes always ended up being close to the bottom of the array after the eye blink? The answer is no. Recall that the average blink latency was 267 ms, the average blink duration was 263 ms, and the duration of the letter array was 106 ms. This means that, on average, the letter display was extinguished more than 400 ms before the eyes opened following the eye blink. That is, after the eye blink, the eyes opened to a blank screen. There was nothing to see, so there was no way for eye position after the eye blink to influence accuracy or errors. Furthermore, it was not the case that the eyes always ended up near the bottom of the array following an eye blink. Rather, horizontal and vertical eye position were influenced by the partial-report cue, as summarized above. One possible explanation for this is that following the eye blink, attention began to be reallocated toward the position indicated by the partial report cue. This might have the effect of influencing eye position (e.g., Laubrock, Kliegl, Rolfs, & Engbert, 2010) but would have no effect on accuracy or errors, because there was no letter information available on the display.
In sum, the results of Experiment 1 indicate that blinking does not simply interfere with performance in general, as it would if attention were simply withdrawn from the display during blink programming, but rather that attention moves in a downward direction before the blink is executed. Attention is required to bind object identity information with object position information, so accuracy increased and mislocation errors decreased for the bottom group of letters on blink trials, as compared with no-blink trials. The offset of the letter array occurred, on average, 161 ms before the blink was actually initiated, indicating that the downward attention movement took place during blink programming, rather than during blink execution. Following blink execution, analysis of the eye position data suggests that task instructions may have resumed control over attentional allocation.
Experiment 2
Previous research investigating the relationship between saccadic eye movements and attention has shown that attention movements precede saccades in an obligatory fashion; that is, attention moves to the saccade target location even when subjects are instructed to attend elsewhere (e.g. Deubel & Schneider, 1996; Hoffman & Subramaniam, 1995; Irwin & Gordon, 1998; Kowler et al., 1995; Shepherd et al., 1986). Experiment 2 investigated whether this is true for blinks as well.
As in Experiment 1, subjects were briefly shown a display containing four groups of letters, three letters per group, arranged around the center of the display in a diamond pattern. Subjects were instructed to attend to one group position (top, left, or right) more than the others because this position would be cued 3 times as often as any other position. As in Experiment 1, on some trials subjects were told to blink as soon as they saw the letter display (blink condition), while on other trials they were told to keep their eyes open and fixated (no-blink condition). If attention moves downward before a blink in an obligatory fashion, report of the bottom letter group should be more accurate under blink than under no-blink conditions even when subjects are trying to attend to some other array position.
Subjects
Eighteen students from the University of Illinois community were recruited for this experiment. Subjects reported normal or corrected-to-normal vision and were naïve as to the purpose of the experiment. They received payment for participating in a single 50-min session.
Procedure
The stimuli and apparatus were identical to those used in Experiment 1. The procedure was also the same as in Experiment 1, except that one position (the high-probability position; top, left, or right) was cued 3 times as often as any other position during the course of the experiment. Subjects were informed of this manipulation and were instructed to attend to the high-probability position because it was most likely to be cued for report on any given trial. The top position was the high-probability position for one group of 6 subjects, the left position for a second group of 6 subjects, and the right position for a third group of 6 subjects.
The sequence of events on each trial was identical to that in Experiment 1. Following a drift correction procedure, a blank screen was presented for 500 ms, followed by presentation of the letter display for 106 ms. Immediately after display offset, the arrow cue was presented for 47 ms. One second after the arrow cue disappeared, a response prompt appeared, and subjects were asked to type the three letters that had appeared in the cued group. There was no time limit on responding. Subjects pressed the return key to enter their response, and feedback was provided. The correct letter had to be reported in the correct position in order for the response to be considered correct. The next trial began after this feedback was provided.
As in Experiment 1, each participant completed blocks of trials under blink and no-blink conditions. Trials on which the participant blinked before the partial-report cue was extinguished (i.e., blinks with latencies of less than 154 ms) were deleted from analysis; this occurred on 2% of the blink trials. The average latency of voluntary blinks in this experiment was 362 ms (SE = 48 ms), and the average blink duration was 197 ms (SE = 16 ms). Because the duration of the letter array was 106 ms, the blink was initiated 256 ms, on average, after the offset of the letter display.
Design
Subjects completed two blocks of 36 practice partial-report trials before beginning the experimental blocks. Subjects were instructed to attend to one group position (top, left, or right) more than to the others because this position would be cued 3 times as often as any other position. The subjects did not wear the eyetracker during the practice trials, and no instructions about blinking were provided. After completing these practice trials, subjects were fitted with the eyetracker, and a calibration procedure was run. They then completed three blocks of 36 trials in the no-blink partial-report condition and three blocks of 36 trials in the blink condition. The ordering of conditions was counterbalanced, with half of the subjects running through the no-blink condition first and the other half running through the blink condition first. The direction of the arrow cue was sequenced randomly across trials, with the high-probability letter group being indicated 18 times and the three other letter groups being indicated 6 times each in each block. The ISI between letter offset and arrow cue onset was 0 ms. A written prompt appeared for 2,000 ms at the beginning of each block, which instructed subjects whether or not to blink during the trials in that block.
Results and discussion
As in Experiment 1, the primary question under investigation was whether partial-report performance would differ in the blink versus no-blink condition and whether this would interact with the position of the cued letter group. To increase power and to make the data easier to interpret, three positions were defined for analysis purposes. The biased position refers to the position that a subject was instructed to attend to (i.e., top, left, or right); the bottom position refers to the bottom letter-group in the array, which was never the focus of the attentional bias instruction; and the unbiased position refers to the average of the two remaining position. This would be the left and right positions if a subject was instructed to attend to the top position, the top and right positions if a subject was instructed to attend to the left position, and the top and left positions if a subject was instructed to attend to the right position. Correct responses, mislocation errors, and intrusion errors were analyzed as a function of position and blink condition (blink vs. no-blink) to determine whether blinks would induce attention to move downward or whether subjects could maintain their attention at the instructed position.
Figure 3 shows the mean percentage of correct responses, mislocation errors, and intrusion errors in the blink and no-blink conditions as a function of the position of the letter group (biased, unbiased, or bottom). Separate two-way ANOVAs with factors of condition (blink vs. no-blink) and position were conducted on each dependent measure. The results for correct responses are considered first. Subjects paid attention to the attention instruction, as shown by the fact that accuracy for the biased position was the highest (68%), while accuracy for the unbiased (44%) and bottom (40%) positions was low; the main effect of position was significant, F(2, 34) = 19.4, MSE = 0.044, p < .001. There was no significant difference in overall accuracy between blink (51.7%) and no-blink (49.0%) trials, F(1, 17) = 2.3, MSE = 0.006, p > .14. Most important, there was a significant interaction between condition and position, F(2, 34) = 3.6, MSE = 0.004, p < .04. The error term for this interaction was used to construct a 95% confidence interval for comparing two means; differences greater than 3.9% were significant. Thus, the interaction occurred because there was a significant difference in accuracy between the no-blink and blink conditions only for the bottom position, with accuracy higher under blink (43.1%) than under no-blink (36.3%) conditions. Blinking had no effect on the biased and unbiased positions. These data indicate that attention moves downward before a blink, facilitating partial-report performance for information presented at the bottom of the stimulus display, even when subjects are trying to attend to another position. Lack of voluntary control and performance benefits without costs are two of the signatures of an automatic attentional process (Posner & Snyder, 1975).

Mean percentages of correct responses, mislocation errors, and intrusion errors as a function of position in the stimulus display for no-blink (NB) and blink (B) trials in Experiment 2. Error bars represent standard errors of the means
Mislocation errors were much lower for the biased position (25%) than for the unbiased (42%) and bottom (43%) positions, F(2, 34) = 16.0, MSE = 0.024, p < .001, indicating that attention facilitated the binding of identity and position information. There was no significant difference in the percentages of mislocation errors for the blink (36%) and no-blink (37%) conditions, F(1, 17) = 1.0, MSE = 0.006, p > .3. Although there were fewer mislocation errors for the bottom position under blink (41.2%) than under no-blink (45.2%) conditions, the interaction between condition and position was not significant, F(2, 34) = 1.6, MSE = 0.004, p > .2. Intrusion errors were lower for the biased position (7%) than for the unbiased (15%) and bottom (17%) positions, F(2, 34) = 16.1, MSE = 0.006, p < .001, indicating that attention facilitated letter identification (cf. Eriksen & Collins, 1969; Eriksen & Hoffman, 1972). There was no significant difference in the percentages of intrusion errors for the blink (13%) and no-blink (14%) conditions, F(1, 17) = 1.6, MSE = 0.001, p > .2. There were fewer intrusion errors for the bottom position under blink (15.7%) than under no-blink (18.5%) conditions, but the interaction between condition and position was not significant, F(2, 34) = 2.2, MSE = 0.001, p > .12.
The horizontal and vertical eye positions following the eye blink were analyzed in ANOVAs with array location as a within-subjects factor and the attentional bias instruction as a between-subjects factor. The bias instruction did not interact with array location in either analysis (for the horizontal eye position, F(6, 45) = 2.2, MSE = 160.2, p > .05; for the vertical eye position, F(6, 45) = 0.5, MSE = 193.7, p > .7), so the results described next are averaged across groups. After the eye blink, the horizontal position of the eye was 0.3° left of center when the top location was cued, 0.9° left of center when the left location was cued, 0.7° right of center when the right location was cued, and 0.1° left of center when the bottom location was cued. The effect of array location was significant, F(3, 45) = 15.9, MSE = 160.2, p < .001. The error term for this effect was used to construct a 95% confidence interval for comparing two means; differences greater than 0.39° were significant, indicating that the eyes deviated to the left significantly more when the left position was cued than when the top or bottom position was cued, and significantly more to the right when the right position was cued. Similar results were found for the vertical eye position. After the eye blink, the vertical position of the eye was 0.1° above center when the top location was cued, 0.4° below center when the left location was cued, 0.4° below center when the right location was cued, and 1.1° below center when the bottom location was cued. The effect of array location was significant, F(3, 45) = 7.3, MSE = 193.7, p < .02. The error term for this effect was used to construct a 95% confidence interval for comparing two means; differences greater than 0.43° were significant, indicating that the eyes ended up significantly higher when the top location was cued than when the left or right location was cued and significantly lower when the bottom location was cued. These results replicate those of Experiment 1.
Given that the average blink latency in this experiment was 362 ms, the average blink duration was 197 ms, and the duration of the letter array was 106 ms, the letter display was extinguished more than 450 ms, on average, before the eyes opened following the eye blink. Thus, there was no way for eye position after the eye blink to influence accuracy or errors. The fact that horizontal and vertical eye positions after the eye blink were influenced by the partial-report cue suggests that following the eye blink, attention begins to be reallocated toward the position indicated by the partial-report cue. This influences eye position but has no effect on accuracy or errors, because there is no letter information available on the display.
In sum, the results of Experiment 2 indicate that attention moves in a downward direction before a blink is executed, even when subjects are trying to attend to another location. Subjects were never instructed to attend to the bottom letter group, and it was cued for report on only one sixth of all trials, but accuracy for this position was significantly higher under blink than under no-blink conditions. Both kinds of errors were reduced under blink conditions, since the blink-induced attention shift appeared to facilitate both identification and binding of items in this low-probability position. Accuracy was higher for the high-probability position (the position that subjects were instructed to attend to) than for all the other positions, indicating that subjects did follow the attention instructions. It is worth noting that blink latency was substantially longer in this experiment (362 ms) than in Experiment 1 (267 ms). It is possible that subjects attempted to encode the letters in the high-probability position before programming their eye blink, thereby delaying blink latency. Similar effects have been found on saccade latency in experiments that have instructed subjects to attend to one location while making a saccade to a different location (e.g., Hoffman & Subramaniam, 1995; Irwin & Gordon, 1998; Kowler et al., 1995). Delaying blink programming would also delay the downward shift of attention, which would explain why accuracy for the bottom letter group, although higher on blink than on no-blink trials, was not as high as accuracy for the position that subjects were instructed to attend to. That is, because the blink occurred well after display offset (on average, 256 ms after display offset in Experiment 2), iconic memory for the display would have decayed substantially before the blink was programmed and attention was moved downward in the display. This would lower overall accuracy for the bottom group of letters in the array; despite this, the most important finding is that blinking still facilitated report of the information contained there.
General discussion
The purpose of the present research was to investigate how the allocation of visual attention is affected by eye blinks. Subjects completed a partial-report task under blink and no-blink conditions. Experiment 1 showed that blinking facilitated report of the bottom row of the stimulus array: Accuracy for the bottom row increased and mislocation errors decreased under blink, as compared with no-blink, conditions, indicating that attention moves downward before a blink. Experiment 2 showed that this was true even when subjects were biased to attend elsewhere, indicating that the blink-induced attention movement is involuntary. Initiation of the eye blink occurred well after the stimulus display was extinguished, indicating that it was blink programming, rather than blink execution, that caused the attention movement. Task instructions presumably drove attention after the blink was executed, as suggested by the eye position analyses that showed that the eyes drifted toward the cued location after the eye blink.
Our results show that the relationship between eye blinks and visual attention is similar to the relationship between saccadic eye movements and visual attention. That is, previous research has shown that attention precedes the eyes to the saccade target location even when subjects are instructed to attend elsewhere (e.g., Deubel & Schneider, 1996; Hoffman & Subramaniam, 1995; Irwin & Gordon, 1998; Kowler et al., 1995; Shepherd et al., 1986), and the present results show that attention moves downward before an eye blink as well. As was noted in the introduction, the eyes move downward before an eye blink, which raises the question of whether the blink-induced attention shifts that we observed were actually due to saccades, rather than to eye blinks. We think not, because the characteristics of blink-induced eye movements (e.g., duration and velocity) are different from those of saccades (Bour et al., 2000; Collewijn et al., 1985; Evinger et al., 1984). Rather, blink-induced eye movements seem to occur as a result of the eyeballs being retracted during the eye blink to ease the passage of the eyelids (Bour et al., 2000; Collewijn et al., 1985; Evinger et al., 1984). Nonetheless, it is the case that attention precedes many kinds of eye movements (see Hoffman, 1998, for a review), so perhaps blink-induced eye movements can be added to the list.
Because accuracy for the bottom row of the stimulus display was higher under blink than under no-blink conditions, it is natural to wonder whether this might have been due to the bottom row being the last thing seen before the eyes blinked. Strictly speaking, this is not true, because the stimulus display was erased from the screen well before eye blink initiation; that is, there was nothing on the display to be seen when the eyes started to move downward. It does seem likely that subjects encoded the stimulus display from top to bottom under both the blink and no-blink conditions, on the basis of the fact that accuracy was highest for the top row under both conditions. This does not explain why accuracy for the bottom row was higher under the blink than under the no-blink condition, however. In a metaphorical sense, one could say that the bottom row was the last thing seen by the “mind’s eye,” however, as attention moved downward during eye blink programming. In this sense, saying that the bottom row was the last thing “seen” is just a restatement of the conclusion that attention moves downward before an eye blink.
One might also wonder whether our results are specific to eye blinks, or whether any secondary motor task might cause attention to move in a downward direction. Although not directly investigated here, the results of Thomas and Irwin (2006) suggest not. As was described above in the introduction to the present article, Thomas and Irwin found that eye blinks interfered with partial-report performance in general, but not when the bottom row of the letter array was cued for report. Thomas and Irwin found, in addition, that buttonpresses and eye closing (as opposed to eye blinks) had no deleterious effects on partial-report performance, indicating that not all motor movements influence partial-report performance. Furthermore, reanalysis of their buttonpress and eye-closing experiments showed that these movements had no differential effect on performance as a function of array position, as would be the case if attention moved downward before these movements.
A question that requires further investigation is whether attention moves downward before reflexive eye blinks, as well as before the voluntary eye blinks that were investigated here. Reflexive eye blinks occur involuntarily, in response to some external stimulus (such as a loud noise or a puff of air directed at the cornea), whereas voluntary eye blinks are made in response to experimental instructions or other endogenous factors. Reflexive and voluntary eye blinks affect visual sensitivity in the same way (Manning et al., 1983; Volkmann, 1986; Volkmann, Riggs, Ellicott, & Moore, 1982; Volkmann et al., 1980), so it is possible that reflexive eye blinks would cause attention to move downward just as do voluntary eye blinks. If it is the eye movement per se that is important, the same results should be found for both reflexive and voluntary eye movements. On the other hand, the neural circuits for reflexive and voluntary eye movements are different, so if it is elements of the motor program that are important, it is possible that attention would not move downward before reflexive eye blinks. Either result would be interesting and informative, so we plan to investigate this in future research.
In conclusion, the purpose of this research was to examine the following question: Where does attention go when you blink? The results of the present experiments indicate that attention does not simply disengage from where it was located before the blink and await reallocation after the eyes reopen; nor does it stay located wherever it was before the eye blink, ensuring continuity of attentional focus across the eye blink. Rather, attention moves somewhere specific in response to the eye blink’s being programmed and executed, in much the same way that attention moves somewhere specific before a saccadic eye movement. In particular, attention moves downward before a blink, even when you are trying to attend somewhere else.
References
Bour, L. J., Aramideh, M., & Ongerboer de Visser, B. W. (2000). Neurophysiological aspects of eye and eyelid movements during blinking in humans. Journal of Neurophysiology, 83, 166–176.
Collewijn, H., Van Der Steen, J., & Steinman, R. M. (1985). Human eye movements associated with blinks and prolonged eyelid closure. Journal of Neurophysiology, 54, 11–27.
Deubel, H., & Schneider, W. X. (1996). Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research, 36, 1993–1997.
Dick, A. (1969). Relations between the sensory register and short-term storage in tachistoscopic recognition. Journal of Experimental Psychology, 82, 279–284.
Eriksen, C. W., & Collins, J. F. (1969). Temporal course of selective attention. Journal of Experimental Psychology, 80, 254–261.
Eriksen, C. W., & Hoffman, J. E. (1972). Temporal and spatial characteristics of selective encoding from visual displays. Perception & Psychophysics, 12, 201–204.
Evinger, C., Manning, K. A., & Sibony, P. A. (1991). Eyelid movements: Mechanisms and normal data. Investigative Ophthalmology & Visual Science, 32, 387–400.
Evinger, C., Shaw, M. D., Peck, C. K., Manning, K. A., & Baker, K. (1984). Blinking and associated eye movements in human, guinea pigs and rabbits. Journal of Neurophysiology, 52, 323–339.
Henderson, J. M., Pollatsek, A., & Rayner, K. (1989). Covert visual attention and extrafoveal information use during object identification. Perception & Psychophysics, 45, 196–208.
Higgins, J. S., Irwin, D. E., Wang, R. F., & Thomas, L. E. (2009). Visual direction constancy across eye blinks. Attention, Perception, & Psychophysics, 71, 1607–1617.
Hoffman, J. E. (1998). Visual attention and eye movements. In H. Pashler (Ed.), Attention (pp. 119–154). London: University College London Press.
Hoffman, J. E., & Subramaniam, B. (1995). The role of visual attention in saccadic eye movements. Perception & Psychophysics, 57, 787–795.
Irwin, D. E., & Gordon, R. D. (1998). Eye movements, attention, and transsaccadic memory. Visual Cognition, 5, 127–155.
Irwin, D. E., & Yeomans, J. M. (1986). Sensory registration and informational persistence. Journal of Experimental Psychology: Human Perception and Performance, 12, 343–360.
Klein, R. (1980). Does oculomotor readiness mediate cognitive control of visual attention? In R. S. Nickerson (Ed.), Attention and performance VIII (pp. 259–276). Hillsdale: Erlbaum.
Klein, R., & Pontefract, A. (1994). Does oculomotor readiness mediate cognitive control of visual attention? Revisited! In C. Umiltà & M. Moskovitch (Eds.), Attention and performance XV: Conscious and nonconscious information processing (pp. 333–350). Cambridge, MA: MIT Press.
Kowler, E., Anderson, E., Dosher, B., & Blaser, E. (1995). The role of attention in the programming of saccades. Vision Research, 35, 1897–1916.
Laubrock, J., Kliegl, R., Rolfs, M., & Engbert, R. (2010). When do microsaccades follow spatial attention? Attention, Perception, & Psychophysics, 72, 683–694.
Manning, K. A., Riggs, L. A., & Komenda, J. K. (1983). Reflex eyeblinks and visual suppression. Perception & Psychophysics, 34, 250–256.
Mewhort, D., Campbell, A., Marchetti, F., & Campbell, J. (1981). Identification, localization, and “iconic memory”: An evaluation of the bar-probe task. Memory & Cognition, 9, 50–67.
O’Regan, K., Deubel, H., Clark, J. J., & Rensink, R. A. (2000). Picture changes during blinks: Looking without seeing and seeing without looking. Visual Cognition, 7, 191–211.
Posner, M., & Snyder, C. (1975). Attention and cognitive control. In R. L. Solso (Ed.), Information processing and cognition: The Loyola symposium (pp. 55–85). Hillsdale: Erlbaum.
Rayner, K., McConkie, G., & Ehrlich, S. (1978). Eye movements and integrating information across fixations. Journal of Experimental Psychology: Human Perception and Performance, 4, 529–544.
Riggs, L. A., Volkmann, F. C., & Moore, R. K. (1981). Suppression of the blackout due to blinks. Vision Research, 21, 1075–1079.
Shepherd, M., Findlay, J., & Hockey, R. (1986). The relationship between eye movements and spatial attention. The Quarterly Journal of Experimental Psychology, 38A, 475–491.
Sperling, G. (1960). The information available in brief visual presentations. Psychological Monographs, 74, 1–29.
Stern, J. A., Walrath, L. C., & Goldstein, R. (1984). The endogenous eyeblink. Psychophysiology, 21, 22–33.
Thomas, L. E., & Irwin, D. E. (2006). Voluntary eyeblinks disrupt iconic memory. Perception & Psychophysics, 68, 475–488.
Townsend, V. (1973). Loss of spatial and identity information following a tachistoscopic exposure. Journal of Experimental Psychology, 98, 113–118.
Treisman, A., & Gelade, G. (1980). A feature integration theory of attention. Cognitive Psychology, 12, 97–136.
VanderWerf, F., Brassinga, P., Reits, D., Aramideh, M., & Ongerboer de Visser, B. (2003). Eyelid movements: Behavioral studies of blinking in humans under different stimulus conditions. Journal of Neurophysiology, 89, 2784–2796.
Volkmann, F. C. (1986). Human visual suppression. Vision Research, 26, 1401–1416.
Volkmann, F. C., Riggs, L. A., Ellicott, A. G., & Moore, R. K. (1982). Measurements of visual suppression during opening, closing and blinking of the eyes. Vision Research, 22, 991–996.
Volkmann, F. C., Riggs, L. A., & Moore, R. K. (1980). Eyeblinks and visual suppression. Science, 207, 900–902.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Irwin, D.E. Where does attention go when you blink?. Atten Percept Psychophys 73, 1374–1384 (2011). https://doi.org/10.3758/s13414-011-0111-0
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-011-0111-0