
Abstract
Background
Collagen extracted from fish body parts is a promising biological material. It has an important role in many pharmaceutical, medical applications and tissue engineering such as corneal regeneration and stromal replacement. The present work investigates a new trend to extract collagen from the fish cornea, as a prospected substituent of human corneal collagen by characterizing some biochemical and optical properties of the fish corneal collagen.
Results
Examination of the corneal tissue of Nile tilapia; Oreochromis niloticus was conducted using electron microscopy, Fourier transform infrared (FTIR) spectroscopy, UV–visible spectrophotometry, optical properties, and thermal properties. The fish were divided into 10 groups each of which consisted of 5 fish. 2 groups of fish were examined for each technique. Results indicated that the corneal layers of O. niloticus are thin at the center and thicker at the periphery with the stroma consisting of a triple helical structure collagen type I. The fish cornea showed very weak transmission at the UV regions (190 nm) and maximum transmission at the visible regions. The values of transmission (T), reflected light (R) and scattered light (S) were 2.685 mw, 100 × 10−3 mw at 45° and 40 × 10−3 mw, respectively. Consequently, the percentage of absorbed light is 21.76%. The denaturation temperature of the fish corneal stroma is 22.27 °C.
Conclusions
The method for obtaining fish collagen affects the specific properties of collagen and consequently its further uses as a potential biomedical substituent for mammalian collagen. Specification of the fish species and tissue type is crucial in identifying the quality as well as the physical and functional properties of the extracted collagen.
Similar content being viewed by others


Background
The link between aquaculture production and fish waste is direct. This problem represents a worldwide environmental challenge. Recycling fish waste and obtaining valuable biomaterials is an environmentally accepted solution for this problem. Several authors stressed the importance of isolating and converting collagen from fish waste into marketable products, as well as the positive influence of this process on the environment [1, 2]. Compensatory corneal tissues represent another added value that could be achieved from recycling fish collagenous byproducts. Human corneal damage and corneal diseases represent the main causes of vision loss [3]. Human corneal transplantation is a vision rehabilitation procedure, used when the corneal clarity is negatively affected. Regardless of the used technique, the scarcity of a donor for corneal tissue is the greatest limiting factor in developing countries, while globally this problem results in approximately 10 million untreated patients [4, 5].
Recently, corneal tissue could be obtained from several animal species [6]. However, when selecting an animal species for conducting a human corneal xenograft, the biochemical composition, and structural differences of corneal proteins between humans and other animal species must be considered to minimize the immune response and improve xenotransplantation outcomes [7]. Although several authors discussed the extraction of collagen from fish skin, scales, and bone, as far as known, no previous research has investigated the extraction of collagen from fish cornea.
Studies investigating the histological and biochemical structure of vertebrate corneas have identified several regions that constitute the corneal tissue [8, 9]. The outer peripheral non-keratinized epithelium is supported by a thin dense membrane called the basement membrane and followed by Bowman’s layer which is a thin dense acellular sheath in between the basement membrane and the anterior stroma of the cornea. The corneal stromal layers are collagenous, soft, and flexible. Stroma provides the thickness of the cornea. Differences in shape and structure between vertebrate corneas are related to the differences in stromal thickness and nature of the basement membrane. Beneath the stroma lies the Descemet's membrane, which is a dense and thick layer that supports a single layer of endothelial cells. The endothelial layer constitutes the innermost layer of the cornea.
The cornea is a transparent avascular tissue that has refractive power. Light passes through the cornea to the lens then reaches the retina inside the eyeball [10]. The cornea also protects the internal constituents of the eyeball. Consequently, clarity, tensile strength and proper shape are the main features of the corneal tissue. Fish have a wide range of corneal phenotypes depending on their habitats. Differences in biochemical and structural properties of vertebrate collagen fibers are the main reasons for the presence of different collagen types [11, 12].
The advantages of marine collagen draw attention to its use instead of mammalian collagen in the manufacture of medical dressings such as sponges and wound-treatment membranes. Marine collagen is fat-free, shows weak antigenicity, compatible with mammalian collagen and is easily absorbed by the human body [13, 14]. Marine collagen is used as a biomaterial for ophthalmic medical devices due to its low toxicity and low antigenicity [15, 16]. The low melting point, difficulties of extraction and low mechanical strength are the main disadvantages that restrict the use of marine collagen as an alternative to mammalian collagen [17,18,19,20].
The present study aims to investigate utilization of collagen obtained from the cornea of Nile tilapia fish; Oreochromis niloticus as a potential alternative to mammalian corneal collagen by examining its biochemical and optical properties.
Methods
Experimental fish
A total of 50 healthy specimens of unisex Nile tilapia O. niloticus, with an average body weight of 250.0 ± 10.0 g, an average body length of 20.0 ± 1.0 cm and an average corneal diameter of 12.0 ± 1.0 mm were used. Fish were transferred to the laboratory in well-aerated 150 L fiberglass tanks to avoid fish hyperactivity, physical injuries, and stress. Upon arrival, the fish were immersed in 5.0 g/L NaCl sterile solution, followed by 1.0% KMnO4 sterile solution to eliminate any possibility of the presence of ectoparasites. Fish were screened for any pathological symptoms and acclimatized under laboratory conditions for 2 weeks in a fiberglass tank (1.0 m3) supplied with de-chlorinated tap water under natural photoperiod and temperature. Renewal of 50% of water was done every week. Ammonia and ammonium concentrations were checked twice a week. Fish were fed twice daily on a conventional fish diet (30% crude protein) at a rate of 3% of the body weight. Water temperature, pH, alkalinity, total hardness and dissolved oxygen were measured daily and maintained at normal conditions during the whole study period. A total of 5 healthy fish were considered as a single group to conduct duplicate group treatments for each of the following techniques.
Collagen purification
Fish were anaesthetized with MS-222 (50 ppm), and then the whole cornea was examined using a slit lamp. After that, the eyes were enucleated and the corneas were isolated for scanning electron microscopy (SEM), UV–visible spectrophotometry and optical properties. Corneal epithelium, endothelium and basement membrane were removed by scratching and lamellae were collected and processed for the Fourier transform infrared (FTIR) spectroscopy and thermal properties investigations.
Scanning electron microscopy (SEM)
The morphological structure of the fish cornea and its quality in terms of collagen layers’ smoothness and regularity was observed under SEM. Samples were dehydrated in ascending concentrations of ethyl alcohol, dried, examined and photographed at different angles using a VEGA3 TESCAN electron microscope at accelerating voltages of 5 or 10 kV.
Fourier transform infrared (FTIR) spectroscopy
For the identification of collagen type, FTIR spectra were recorded (Thermo Scientific FTIR, model Nicolet 5s Madison, WI, USA). The equipment consists of an ATR ID3 accessory for germanium crystal reflection. The spectral resolution was 4 cm−1. 64 scans were obtained in the range of 400–4000 cm−1. Fresh corneal lamellae were cut manually into small pieces and stored in phosphate-buffered saline (PBS) solution. The C=O, NH2 and C–N groups were tested for absorption of amid I, II and III bands, respectively.
UV–visible spectrophotometry
The UV–visible absorption-transmittance spectrum of the O. niloticus corneal tissue samples were quantified using UV–visible spectrophotometry (Thermo Scientific, Genesys 10S UV–Vis, USA). The absorbance was quantified at different lengths in the scope of 180–1000 nm (5 nm intervals). The spectrophotometer was set up so the cornea and microscope cover glass could be placed perpendicular to the estimating beam. The cover glass was placed on the cuvette holder. The transmission percentage indicator was chosen. First, the base spectrum of the microscope cover glass was measured. Second, the corneal sample was placed on the cover glass with the epithelium directed downward. By placing the samples in this orientation, the estimated light beam of the spectrophotometer entered the cornea from the epithelial side, that is, from the identical direction as the cornea in situ. The cover glass and the cornea were replaced. Their position was adjusted so that the rectangular estimating beam was in the center of the cornea. After adjustments were made, the cornea samples were scanned.
Optical properties (transmission, reflection, scattering and absorption)
A laser-based bench-top optical system was used. A 650 nm red laser was applied to detect the percentage of transmission (T), reflection (R) of light at 45° and scattering (S) of incident light on fish cornea at incident light intensity (Po) of 3.61 mw. Where, T% = T/Po × 100, R% = R/Po × 100 and S% = S/Po × 100. The absorption percentage (A%) is calculated as 100-(T% + S% + R%). Fresh fish eyeballs were obtained, and the cornea was cut along with a part of the eyeball to maintain the tensile strength of the cornea. It was cleaned of pigments and then tested.
Thermal properties (denaturation temperature)
The thermal properties were determined using a differential scanning calorimetry thermogram method (DSC-60 detector). The stromal sample (weight 2.070 mg) was sealed in an Aluminum seal cell and was subjected to the test in a nitrogen gas atmosphere on a scale from − 50 to 50 °C and a temperature rate of 10 °C/min.
Results
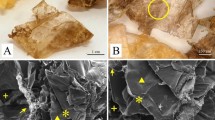
Figure 1A shows that the cornea is thin at the center and thicker at the periphery. The collagen fibers were amalgamated during the processing of the corneal tissue for SEM imaging (Fig. 1B). Figure 1C shows the junction between epithelium and stroma. Figure 1D shows the multiple patterns of the configuration of the collagen fibers at the central sections as well as the anterior and posterior parts of the stroma. The FTIR spectroscopy results (Fig. 2) showed a clear sharp peak at 3285.85 cm−1. The source of the signal is the N–H stretch coupled with a hydrogen bond indicating the presence of amide A. The extremely small peak at 2900 cm−1 may refer to amide B. The peak at 1633.41 cm−1 was related to amide I. Amide II was measured at 1553.09 cm−1. Amide III was measured at 1233 cm−1. The present findings confirm the presence of a triple helical structure characteristic of collagen type I. Figure 3 shows little transmittance of UV in the C region ranging from 100 to 280 nm. A 650 nm red laser beam was used to determine the optical properties of the cornea. The detected results showed that transmission (T), reflected light (R) and scattered light (S) were 2.685 mw, 100 × 10−3 mw at 45° and 40 × 10−3 mw, respectively. The T, R and S percentages were 74.37%, 2.77% and 1.1%, respectively. Consequently, the absorbed light (A) percentage is 21.76%. Results of the present study illustrated that the denaturation temperature of O. niloticus corneal stroma was 22.27 °C (Fig. 4).

Scanning electron microscopic (SEM) micrographs of the fish cornea. A corneal thickness; B amalgamated collagen fibers during processing for SEM imaging; C junction between epithelium and stroma; D patterns of the configuration of collagen fibers; CTP, Corneal thickness at the periphery; CTC, corneal thickness at the center; ACF, amalgamated collagen fibers; NE, nerve ending; CF, collagen fibers

Fourier transform infrared (FTIR) spectroscopy of the fish corneal stroma

UV–visible transmission spectrophotometry of the fish cornea

Thermal properties (denaturation temperature) of the fish corneal stroma
Discussion
The present study highlights the biochemical and optical properties of Nile tilapia; O. niloticus corneal tissue. The principal objective of the assessment of the fish cornea by SEM is to observe the quality (rough or smooth), thickness and to estimate the presence of burrs, curls, or tissue bridges in the stroma region. The cornea of freshwater teleost has one continuous stroma that consists of collagen fibers arranged in bundles with the keratocytes distributed in between [9, 21]. The stroma of fish and human corneas have thin collagen fibers that together appear to indicate an overall parallel collagen direction. The human cornea consists of collagen fibers of different types with unique stacked spacing and arrangement which is an important factor affecting corneal transparency [22].
FTIR spectroscopy analysis is used to detect the functional groups that appear to characterize a compound by estimating its absorption of infrared radiation over different wavelengths. FTIR spectroscopy is used to assess the structure and type of collagen material. In addition, it serves to distinguish between methods of collagen extraction [23,24,25]. Similar pattern of FTIR spectroscopy results was indicated by Wang et al. [26] who explained the connection with the asymmetric stretching of C–H. The peak at 1633.41 cm−1 was related to amide I as a sign of the peptide's secondary structure, and the source of the signal is the C=O [27]. Amide II was measured at 1553.09 cm−1, which is due to the stretching of the carbonyl group attached to a carboxyl group. Amide III was measured at 1233 cm−1, which was determined by combining the C–N stretching and N–H bending modes (Fig. 2). The detected triple helical structure agrees with results reported by Sun et al. [28] for collagen extracted from Nile tilapia; O. niloticus skin. Figure 2 also shows that the functional groups found in the triple helix were not damaged owing to the method used to obtain collagen directly by removing both the epithelium and endothelium cells. Variations in width and height of the corresponding signals in several graphs could be attributed to the different experimental conditions of each study such as different solvents with different concentrations [14].
The wavelength of the UV spectrum ranges from 100 to 400 nm. The range is divided into three regions: C, B and A ranging from 100 to 280 nm, 280–315 nm and 315–400 nm, respectively. Figure 3 shows little transmittance of UVC. This may be attributed to the low thickness of the fish cornea in general. The UV absorption of collagen could be attributed to the peptide bonds and side chains in the corneal collagen structure. The collagen isolated from tilapia skin showed maximum absorbance at 230 nm [27] which is consistent with the characteristic absorbance of collagen type I in the present study. Similar findings were reported by several authors for collagen extracted from different marine species [26,27,28,29]. The human cornea transmits radiation only at 295 nm and above. The transmission of the human cornea is highest in the long wavelength area of the visible spectrum from 600 to 700 nm. Meanwhile, transmission of O. niloticus cornea is highest in the long wavelength region of the visible spectrum ranging from 600 to 700 nm and lowest in the short wavelength region below 500 nm (Fig. 3).
Huibertus et al. [30] measured light scattering and light transmission in fish scale-derived collagen (FSDC) and they concluded that FSDC is proposed as an alternative for mammalian donor corneal tissue which supports the present findings. The refractive index of corneal tissues of human and O. niloticus fish are 1.363 ± 0.011 and 1.360 ± 0.009, respectively [9].
Regarding the thermal properties, it was noticed that one of the most important characteristics of biomaterials is their denaturation temperature because it affects their thermal stability and consequently their physical and mechanical properties [31]. The denaturation temperature is defined as the temperature degree at which the deformation of collagen helical structure occurs [32]. The denaturation temperature of skin collagen of O. niloticus ranged between 25 and 26 °C depending on the conducted extraction procedures [33]. Meanwhile it was detected at 34.85 °C for collagen extracted from O. niloticus scales [34]. The denaturation temperature of collagen depends on both the source tissue and the extraction procedures. Freshwater fish collagen generally show high thermal stability which could be attributed to its high content of proline and hydroxyproline [34,35,36,37]. In agreement with this finding, Yamamoto et al. [18] documented a higher denaturation temperature of Nile tilapia; O. niloticus collagen in comparison with other marine sources.
Conclusions
The present study confirms that the method for obtaining fish collagen (extraction, isolation, purification, etc.) may affect its specific properties and consequently the further uses of collagen as a potential biomedical substituent for mammalian collagen. Specification of the fish species and tissue type is crucial in identifying the quality of the extracted collagen as well as its physical and functional properties. Further studies are recommended to evaluate the potential uses of O. niloticus corneal collagen as a biological substituent for mammalian collagen.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- A:
-
Absorption
- DSC:
-
Differential scanning calorimeter
- FSDC:
-
Fish scale-derived collagen
- FTIR:
-
Fourier transform infrared
- O. niloticus :
-
Oreochromis niloticus
- P o :
-
The incident light intensity
- PBS:
-
Phosphate-buffered saline
- R :
-
Reflection
- SEM:
-
Scanning electron microscopy
- S :
-
Scattering
- T :
-
Transmission
- UV:
-
Ultraviolet
References
Zhang Q, Wang Q, Lv S, Lu J, Jiang S, Regenstein J, Lin L (2016) Comparison of collagen and gelatin extracted from the skins of Nile tilapia (Oreochromis niloticus) and channel catfish (Ictalurus punctatus). Food Biosci 13:41–48. https://doi.org/10.1016/j.fbio.2015.12.005
Ahmed M, Verma AK, Patel R (2020) Collagen extraction and recent biological activities of collagen peptides derived from sea-food waste: a review. J Sustain Chem Pharm 18:100315. https://doi.org/10.1016/j.scp.2020.100315
Tidke SC, Tidake P (2022) A review of corneal blindness: causes and management. Cureus 14(10):30097. https://doi.org/10.7759/cureus.30097
Kalter ESJ, de By TMMH (1997) Tissue banking programmes in Europe. Br Med Bull 53(4):798–816. https://doi.org/10.1093/oxfordjournals.bmb.a011649
Congdon NG, Friedman DS, Lietman T (2003) Important causes of visual impairment in the world today. J Am Med Assoc 290(15):2057–2060. https://doi.org/10.1001/jama.290.15.2057
Sharifi R, Yang Y, Adibnia Y, Dohlman CH, Chodosh H, Gonzalez-Andrades M (2019) Finding an optimal corneal xenograft using comparative analysis of corneal matrix proteins across species. Sci Rep 9:1876. https://doi.org/10.1038/s41598-018-38342-4
Singh RB, Marmalidou A, Amouzegar A, Chen Y, Dana R (2020) Animal models of high-risk corneal transplantation: a comprehensive review. Exp Eye Res 198:108152. https://doi.org/10.1016/j.exer.2020.108152
Hayashi S, Osawa T, Tohyama K (2002) Comparative observations on corneas, with special reference to Bowman’s layer and Descemet’s membrane in mammals and amphibians. J Morphol 254:247–258. https://doi.org/10.1002/jmor.10030
Lai T, Tang S (2014) Cornea characterization using a combined multiphoton microscopy and optical coherence tomography system. Biomed Opt Express 5(5):1494–1511. https://doi.org/10.1364/BOE.5.001494
Sridhar MS (2018) Anatomy of cornea and ocular surface. Indian J Ophthalmol 66(2):190–194. https://doi.org/10.4103/ijo.IJO_646_17
Copes F, Pien N, Van Vlierberghe S, Boccafoschi F, Mantovani D (2019) Collagen-based tissue engineering strategies for vascular medicine. J Front Bioeng Biotechnol 7:166. https://doi.org/10.3389/fbioe.2019.00166
Lin K, Zhang D, Macedo MH, Cui W, Sarmento B, Shen G (2019) Advanced collagen-based biomaterials for regenerative biomedicine. Adv Funct Mater 29:1804943. https://doi.org/10.1002/adfm.201804943
Chinh NT, Manh VQ, Trung VQ, Lam TD, Huynh MD, Tung NQ, Trinh ND, Hoang T (2019) Characterization of collagen derived from tropical freshwater carp fish scale wastes and its amino acid sequence. J Nat Commun 14(7):1–12. https://doi.org/10.1177/1934578X19866288
Jafari H, Lista A, Siekapen MM, Ghaffari-Bohlouli P, Nie L, Alimoradi H, Shavandi A (2020) Fish collagen: extraction, characterization, and applications for biomaterials engineering. J Polym 12(10):2230–2237. https://doi.org/10.3390/polym12102230
Ulery BD, Nair LS, Laurencin CT (2011) Biomedical applications of biodegradable polymers. J Polym Sci 49(B):832–864. https://doi.org/10.1002/polb.22259
Sionkowska A, Skrzyński S, Śmiechowski K, Kołodziejczak A (2017) The review of versatile application of collagen. Polym Adv Technol 28:4–9. https://doi.org/10.1002/pat.3842
Subhan F, Ikram M, Shehzad A, Ghafoor A (2014) Marine collagen: an emerging player in biomedical applications. J Food Sci Technol 52:4703–4707. https://doi.org/10.1007/s13197-014-1652-8
Yamamoto K, Igawa K, Sugimoto K, Yoshizawa Y, Yanagiguchi K, Ikeda T, Yamada S, Hayashi Y (2014) Biological safety of fish (Tilapia) collagen. J BioMed Res Int 9:630757. https://doi.org/10.1155/2014/630757
Carvalho AM, Marques AP, Silva TH, Reis RL (2018) Evaluation of the potential of collagen from codfish skin as a biomaterial for biomedical applications. J Mar Drugs 16(12):495. https://doi.org/10.3390/md16120495
El Blidi O, El Omari N, Balahbib A, Ghchime R, El Menyiy N, Ibrahimi A, Ben Kaddour K, Bouyahya A, Chokairi O, Barkiyou M (2021) Extraction methods, characterization and biomedical applications of collagen: a review. J Biointerface Res Appl Chem 11(5):13587–13613. https://doi.org/10.33263/BRIAC115.1358713613
Van Horn DL, Edelhauser HF, Schultz RO (1969) A comparative study of stromal swelling in sea lamprey spectacle and brook trout cornea. J Ultrastruct Res 28:452–461. https://doi.org/10.1016/S0022-5320(69)80033-X
Meek KM, Knupp C (2015) Corneal structure and transparency. J Prog Retin Eye Res 49:1–16. https://doi.org/10.1016/j.preteyeres.2015.07.001
Payne KJ, Veis A (1988) Fourier transforms IR spectroscopy of collagen and gelatin solutions: deconvolution of the amide I band for conformational studies. J Biopolym 27(11):1749–1760. https://doi.org/10.1002/bip.360271105
Sripriya R, Kumar R (2015) A novel enzymatic method for preparation and characterization of collagen film from swim bladder of fish Rohu (Labeo rohita). J Food Nutr Sci 6:1468–1478. https://doi.org/10.4236/fns.2015.615151
Sanden KW, Kohler A, Afseth NK, Böcker U, Rønning SB, Liland KH, Pedersen ME (2019) The use of fourier-transform infrared spectroscopy to characterize connective tissue components in skeletal muscle of Atlantic cod (Gadus morhua L.). J Biophotonics 12(9):201800436. https://doi.org/10.1002/jbio.201800436
Wang J, Pei X, Liu H, Zhou D (2018) Extraction and characterization of acid-soluble and pepsin-soluble collagen from skin of loach (Misgurnus anguillicaudatus). Int J Biol Macromol 106:544–550. https://doi.org/10.1016/j.ijbiomac.2017.08.046
Sampath N, Nazeer R (2013) Characterization of acid and pepsin soluble collagen from the skin of horse mackerels (Magalaspis cordyla) and croaker (Otolithes ruber). Int J Food Prop 16(3):613–621. https://doi.org/10.1080/10942912.2011.557796
Sun L, Hou H, Li B, Zhang Y (2017) Characterization of acid- and pepsin-soluble collagen extracted from the skin of Nile tilapia (Oreochromis niloticus). Int J Biol Macromol 99:8–14. https://doi.org/10.1016/j.ijbiomac.2017.02.057
Zhang J, Duan R, Tian Y, Konno K (2009) Characterisation of acid-soluble collagen from skin of silver carp (Hypophthalmichthys molitrix). J Food Chem 116:318–322. https://doi.org/10.1016/j.foodchem.2009.02.053
Huibertus TE, Lin CC, Hussain AK, Maas S, Lai HJ, Linnartz H, van den Berg TJTP, Salvatori DCF, Luyten GPM, Jager MJ (2013) A fish scale-derived collagen matrix as artificial cornea in rats: properties and potential. Invest J Ophthalmol Vis Sci 54(5):3224–3233. https://doi.org/10.1167/iovs.13-11799
Gauza-Włodarczyk M, Kubisz L, Mielcarek S, Włodarczyk D (2017) Comparison of thermal properties of fish collagen and bovine collagen in the temperature range 298–670 K. Mater Sci Eng C 80(2):468–471. https://doi.org/10.1016/j.msec.2017.06.012
Song WK, Liu D, Sun LL, Li BF, Hou H (2019) Physicochemical and biocompatibility properties of Type I collagen from the skin of Nile tilapia (Oreochromis niloticus) for biomedical applications. Mar Drugs 17(3):137. https://doi.org/10.3390/md17030137
Abdelaal H, Mohamed MAH, Salah S, Elhosany R (2021) Characteristics and functional properties of collagen extracted from Nile tilapia (Oreochromis niloticus) skin. J Mod Res 3:36–43. https://doi.org/10.21608/jmr.2020.37541.1034
Ikoma T, Kobayashi H, Tanaka J, Walsh D, Mann S (2003) Physical properties of type I collagen extracted from fish scales of Pagrus major and Oreochromis niloticus. Int J Biol Macromol 32(3–5):199–204. https://doi.org/10.1016/s0141-8130(03)00054-0
Nagai T, Suzuki N, Nagashima T (2008) Collagen from common minke whale (Balaenoptera acutorostrata) unesu. J Food Chem 111(2):296–301. https://doi.org/10.1016/j.foodchem.2008.03.087
Montero P, Jimennez-Colmenero F, Borderias J (1991) Effect of pH and the presence of NaCl on some hydration properties of collagenous material from trout (Salmo irideus Gibb) muscle and skin. J Sci Food Agric 54:137–146. https://doi.org/10.1002/jsfa.2740540115
Duan R, Zhang J, Dua X, Yao X, Konno K (2009) Properties of collagen from skin, scale and bone of carp (Cyprinus carpio). J Food Chem 112:702–706. https://doi.org/10.1016/j.foodchem.2008.06.020
Acknowledgements
The authors would like to acknowledge Deanship of Scientific Research, Taif University, KSA, Giza Memorial Institute for Ophthalmic Research (MIOR), Giza, Egypt, and National Institute of Oceanography and Fisheries, Cairo, Egypt.
Funding
This work was self-funded.
Author information
Authors and Affiliations
Contributions
HHA-E: idea formulation, primary data collection, generation of database, writing of the manuscript, development of data analysis methodology, interpretation of results, editing of the manuscript; WAO and HAE: data analysis, preparation and editing of the manuscript. All the authors participated in the discussion of the results.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The present experimental investigation was not conducted with humans. Consent to participate is not applicable. All international, national, and/or institutional standards for the use of fish in experiments and conditions of fish care have been followed. The study received approval of the National Institute of Oceanography and Fisheries-Committee for ethical care and use of animals/aquatic animals (NIOF-IACUC) under ethical clearance reference number NIQF-FW3-F-23-R-043.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abd-Elrahman, H.H., Omar, W.AM. & Elnashar, H.A. Characterization of biochemical and optical properties of Nile tilapia (Oreochromis niloticus) corneal collagen. Futur J Pharm Sci 10, 39 (2024). https://doi.org/10.1186/s43094-024-00597-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43094-024-00597-2