
Abstract
Background
Aloe vera leaf gel has proven efficacious roles in the amelioration of several human diseases and illness-conditions. Specific purified gel-derived bio-constituents as well as the naturally harvested unprocessed A. vera gel have shown promise in modifying systemic inflammation. However, the synergistic role of natural herbal remedies, a mainstay of traditional Indian Ayurveda, has not been evaluated rigorously in this plant. In this study, the prevention of membrane lysis and protein denaturation in the presence of A. vera gel homogenate up to the concentration of 1000 μg/ml of gel has been assessed in vitro. Also, regulation of expression of inflammation-mediator genes (TNF-α and Cox-2) has been investigated in vivo in Freund’s complete adjuvant (FCA)-induced inflammatory arthritic Wistar albino rats in a 28-day long study following the daily oral supplementation of Aloe vera gel homogenate doses up to 0.40 and 0.80 g/kg body weight (low-dose and high-dose groups respectively).
Results
Our results indicated that A. vera gel homogenate inhibits hypotonicity-induced (74.89 ± 1.26%) and heat-induced (20.86 ± 0.77%) RBC membrane lyses respectively at a concentration of 1000 μg/ml, compared to indomethacin standard (80.52 ± 0.65% and 43.98 ± 1.52% respectively at 200 μg/ml concentration). The similar concentration of gel also showed 39.35 ± 4.25% inhibition of protein denaturation compared to standard diclofenac sodium (46.74 ± 1.84% at 100 μg/ml concentration) in vitro. When assessed in vivo, TNF-α expression was found to be decreased by 35.88% and 38.52%, and Cox-2 expression was found to be decreased by 31.65% and 34.96%, in low-dose and high-dose groups respectively, when compared to the arthritic controls.
Conclusions
Our findings justify the role of unprocessed A. vera gel homogenate in preventing tissue damage and in the downregulation of TNF-α and Cox-2 gene expressions for the immune-modulation of inflammatory arthritis condition.
Similar content being viewed by others
Background
Inflammation is a common complex biological cascade of events that occurs in response to any kind of injuries, mechanical damage, infection, toxin exposure, or chemical irritation in the tissue. Inflammatory pathways lead to the protection of the body. However, prolonged inflammation can cause chronic disorders and extensive tissue damage. Depending on the duration of the inflammatory responses in the body, the inflammatory processes are commonly grouped into acute (short-duration) and chronic (long-duration) inflammation. During inflammation, immune cells release lysosomal contents that act as mediators of inflammation. These enzymes work non-specifically on the nearby cells and cause further damage by breaking their membranes. This process significantly increases the degree of inflammation [1]. The erythrocyte (RBC) membranes are considered as a model of the lysosomal membrane for their structural similarity [2]. It is expected that the drugs or products capable of preventing the rupture of the erythrocyte membrane experimentally would also protect the lysosomal membrane of the affected tissue in vivo resulting in the inhibition of inflammation. On the other hand, an increase in protein denaturation has been a hallmark of inflammation [3]. Drugs exhibiting the protein denaturation-inhibition in vitro are expected to elicit similar results in biological systems and would stabilize inflammation [4].
Rheumatoid arthritis (RA) is a complex systemic inflammatory disease of the bone joints which leads to disabilities of joint movements. Presently, about 0.5–2% of the world population is affected by the disease. The current treatment of RA focuses on the pain reduction and inhibition of disease manifestation through blockage of prime mediator molecules related to the disease [5]. Cytokines and Cox-2 are the prime regulators of inflammatory diseases. TNF-α is one of the important cytokines which regulates the progression of inflammatory RA by inducing different signaling pathways. These signaling pathways are instrumental in immune cell infiltration and increased production of other cytokines [6, 7]. Elevated COX-2 expression in synovial tissues of the arthritic joint is mediated chiefly by the pro-inflammatory cytokines TNF-α and IL-1. The effector product of Cox-2 is prostaglandin E2, which contributes extensively to the degree of inflammation [8, 9].
To treat the consequences of extensive inflammation and inflammatory diseases like RA, non-steroidal anti-inflammatory drugs (NSAIDs), or steroids are commonly used. But due to the different side effects of conventional NSAIDs and steroids, the use of herbal remedies is increasingly becoming a popular choice. However, many of these herbal remedies lack scientific exploration and thus experimental validation [10]. These products are generally classified as complementary and alternative medicines (CAMs). CAMs are generally inexpensive and with no or low side effects. In the Indian CAM system, Ayurveda emphasizes the use of plant and animal products, dietary supplements, minerals, and salts, largely in crude and unprocessed forms, in amelioration of disease conditions. World Health Organization (WHO) has documented that about 80% of the world population directly or indirectly depends on ethnic medications [10, 11].
Aloe vera (Family Xanthorrhoeaceae) is one of the pioneer plants which have been used historically to reduce inflammatory symptoms by different ethnic populations. Researchers have found potent wound healing, anti-inflammatory, anti-arthritic, and anti-nociceptive properties of the Aloe vera gel in appropriate model systems [5, 12,13,14]. The interaction of the plant gel constituents with inflammation modulators is a key area of interest at the experimental level [15]. There is no detailed work reported on such naturally occurring unprocessed A. vera gel in animal models and thus the presented data is expected to contribute to the existing body of knowledge on the medicinal properties of A. vera gel. In the present work, assessment of in vitro anti-inflammatory potentials of unprocessed naturally harvested A. vera plant gel homogenate is done and in vivo assessment of the expression of two apex biomolecules of inflammation namely TNF-α and Cox-2 has been done in FCA-induced inflammatory arthritic rat model following A. vera gel homogenate oral consumption.
Methods
Collection of plant materials and authentication
Naturally grown A. vera plants were collected from Siliguri and adjacent regions which is located in the sub-Himalayan Terai region of West Bengal, India. Collection of the plant material and experiments were done during the year 2016–2018. The plant was identified by competent authority, and a voucher specimen was deposited [Accession No. 09884].
Preparations of plant extract and dose determination
Crude Aloe gel was collected by peeling out the green outer dermal layer and by taking out the gel aseptically with the help of a scalpel. For the in vitro anti-inflammatory tests, the gel was weighed and properly homogenized with isotonic buffer solution or with distilled water (for hypotonicity-induced membrane stabilization test) to obtain final concentrations of 600, 800, and 1000 μg of gel/ml. The sample was freshly prepared every time before use. Authors postulate that the process of simple homogenization preserves all the ingredients of the crude gel in the same proportion as it is obtained naturally from the leaves. The use of homogenization as an extraction process has been supported by other works [16, 17]. For the in vivo experiments on arthritic rat models, doses of 0.40 and 0.80 g of Aloe vera gel/kg body weight (b.w.) (low dose or LD and high dose or HD groups respectively) were prepared by homogenizing Aloe vera gel with distilled water at a ratio of 1:3 (w/v). These doses were prepared based on our previous works [5, 18] considering the possible amount of daily consumption of the Aloe vera gel to be 25–50 g for a human weighing 60 kg. No post-harvesting extraction or processing was done to preserve the naturally available bioactive components of the gel.
Drugs and chemicals
All the chemicals used for the experiments were molecular biology grade chemicals procured from Merck, Sigma Aldrich and HiMedia (India). Freund’s complete adjuvant was procured from Sigma, USA. Reverse transcriptase enzyme was obtained from Thermo Fischer, USA; total RNA was prepared using Trizol (Invitrogen, USA); random hexamer, oligo dT, and dNTP were used from GCC Biotech, India, and SybrGreen reaction mixture was obtained from Roche, USA.
Experimental animal maintenance
Wistar albino male rats (Rattus norvegicus) (150 ± 15 g) were procured from Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA, India) registered animal vendor (M/s Chakraborty Enterprise, Kolkata, India) after the approval of the experimental protocols by the Institutional Animal Ethical Committee (IAEC, NBU) (Registration number 840/ac/04/CPCSEA; IAEC protocol number IAEC/NBU/2018/02). The animals were housed in the departmental animal house facility at 24 ± 2 °C with a 12 h/12 h day/night cycle and were fed with standard pellet and water was provided ad libitum. All animals were acclimatized for a period of at least 10 days before the initiation of the experiments. Four or fewer rats were kept per cage during the entire period.
In vitro anti-inflammatory tests
Collection of blood and preparation of erythrocyte suspension for in vitro membrane-stabilizing experiments
Three milliliters (3 ml) of fresh blood was collected in an EDTA vial from a healthy human volunteer who had not taken any NSAID at least 15 days prior to the collection. Erythrocyte suspension was prepared following the protocol described by Anosike et al. [2] and was stored at 4 °C prior to the experiments [2].
The membrane stabilizing activities of A. vera gel homogenate were assessed by a method following Shinde et al. [19] with some minor modifications.
Hypotonic solution-induced hemolysis test
Hypotonic solution-induced hemolysis test was undertaken following the methodology of Shinde et al. [19], with some modifications. Aloe vera gel homogenates were prepared to attain final concentrations of 600, 800, and 1000 μg of gel/ml either in hypotonic solution (homogenized in distilled water) or in isotonic solution (homogenized in isotonic buffer solution of pH 7.4). The RBC membrane breakage in hypotonic situation was calculated compared to that in the isotonic solution. In standard group, standard drug indomethacin was added to 5 ml isotonic buffer as well as to hypotonic distilled water in separate tubes to attain a final concentration of 200 μg/ml [19]. The absorbance values (OD) of the supernatants were measured at 540 nm. The percentage of inhibition of hemolysis was calculated considering the hemolysis occurring in the hypotonic solution or in distilled water of the control tubes to be 100%. Thus, the percentage of inhibition (PI) of hemolysis was calculated using the following equation:
where:
OD1 = absorbance of test sample in isotonic solution
OD2 = absorbance of test sample in hypotonic solution (distilled water)
OD3 = absorbance of control sample in hypotonic solution (distilled water)
Heat-induced hemolysis test
For this test, one untreated control group, three experimental plant extract-treated groups, and one standard drug group were considered following the methodology described by Shinde et al. [19] with some minor modifications. In the experimental groups, Aloe vera gel was homogenized in isotonic phosphate buffer solution (pH 7.4) at a final concentration of 600, 800, and 1000 μg/ml. The standard drug group contained indomethacin in 5 ml of isotonic buffer solution at a final concentration 200 μg/ml [19]. Absorbance values (OD) of supernatant was measured at 540 nm. The percent inhibition (PI) of hemolysis was calculated using the following equation [20]:
where:
OD1 = absorbance of heated test sample (isotonic buffer)
OD2 = absorbance of heated control sample (distilled water)
Protein denaturation-inhibition test
The protein denaturation-inhibition test was done following established protocol [21] with some minor modifications to investigate the protein denaturation-inhibition activity of Aloe gel homogenate. For the experiment, along with 0.2 ml of egg albumin and 2.8 ml of PBS (pH 6.4), 2 ml of various concentrations of Aloe vera gel homogenates were added to experimental groups which finally gave rise to 600, 800, and 1000 μg/ml concentration of gel in the groups. Diclofenac sodium was used as standard drug. In the standard drug group, the drug was added in the mixture to achieve a final concentration of 100 μg/ml in the 5 ml mixture which contained 0.2 ml of egg albumin and 2.8 ml of PBS (pH 6.4) [21]. The absorbance of the solutions (OD) was measured spectrophotometrically at 660 nm. The percentage of inhibition (PI) of protein denaturation was calculated using the following equation [22]:
where:
OD1 = absorbance of heated test sample
OD2 = absorbance of heated control sample
In vivo anti-arthritic tests
The experimental set up consisted of 24 rats distributed in four animal groups each containing 6 rats (n = 6). Animals were randomly distributed in one positive control (PC), one FCA or negative control group (FCA), and two experimental groups of LD and HD respectively, receiving daily oral supplement of A. vera gel homogenate at a dose of 0.40 g/kg b.w. and 0.80 g/kg b.w. respectively, once a day from the first day of the experiment till 28th day. Freund’s complete adjuvant (Sigma, USA) was administrated (0.1 ml) in the right hind paw of all the experimental rats except PC on the first day. Arthritic swelling was observed within 3–4 days and a booster dose of same amount was given on 14th day. The inflammatory paw swelling of all the groups was observed and was assessed through the measurement of paw circumference with the help of a vernier caliper up to 28th day at a regular interval of 3 days (data not shown). The animals were sacrificed on 28th day following complete anesthesia using diethyl ether. On the day of sacrifice, blood was collected separately from each rat. Total RNA was prepared using Trizol (Invitrogen, USA) following the instruction of the manufacturer. The total RNA prepared separately from each of the animals of each group was then pooled together group-wise. From each group, sufficient amount of cDNA was prepared using reverse-transcriptase enzyme (Thermo Fischer, USA), random hexamer, oligo dT, and dNTP (GCC biotech, India) following the protocol suggested by the manufacturer (Thermo Fischer, USA). The prepared cDNA was used to assess the relative expression of TNF-α and Cox-2 among different groups in Lightcycler 96 real-time quantitative PCR (Roche, Switzerland). The exon-specific primers of TNF-α and Cox-2 was used along with a housekeeping gene GAPDH as endogenous control (Table 1). In brief, 5 μl of cDNA was used and mixed with 10 μl of SybrGreen, 0.3 μl of each of the forward and reverse primers (10 mM) and 1 μl of dNTP. Reaction volume was adjusted up to 20 μl by adding nuclease-free water. Annealing temperature of 59 °C was standardized for all the three genes and 45 amplification cycles were implemented. The relative mRNA expressions of the selected genes were calculated using 2−∆∆Ct method. In this method, the expression of target genes were measured in fold change values which were analyzed with respect to the endogenous control gene expression.
Statistical analysis
The in vitro anti-inflammatory tests were carried out in triplicates for each of the groups. All statistical analyses for anti-inflammatory tests were done using GraphPad prism ver 6.01. All the data were represented as mean ± S.E.M. (standard error mean) and were analyzed with one-way ANOVA followed by Dunnett’s multiple comparisons test. The results were considered statistically significant at P < 0.05 compared to the control group. The *** denotes significance value at P < 0.001.
Results
In vitro anti-inflammatory tests
Effect of Aloe vera gel homogenate on hypotonicity-induced hemolysis of hRBCs
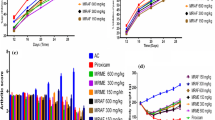
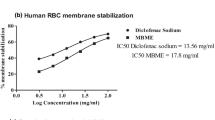
The standard NSAID drug indomethacin showed the highest protection (80.52 ± 0.65%) at the concentration of 200 μg/ml against hypotonicity-induced lysis of RBCs. A. vera gel homogenate protected the human RBCs in a concentration-dependent manner (Fig. 1a). Among the experimental groups, highest protective effect was seen in 1000 μg/ml A. vera gel homogenate dose group (74.89 ± 1.26%).

Results of in vitro anti-inflammatory tests in graph. Hypotonicity-induced hemolysis test of hRBC (a), heat-induced hemolysis test of hRBC (b), and protein denaturation-inhibition test of hen’s albumin (c) when treated with Aloe vera gel homogenate. All data represents % of inhibition ± SEM. Significance value at P < 0.001 is indicated as ***. Indomethacin and diclofenac sodium are used as standard drugs in experiments as indicated. Control group (not shown in the figures) is considered to exert 100% hemolysis
Effect of Aloe vera gel homogenate on heat-induced hemolysis of hRBCs
All the doses of A. vera crude gel homogenate (600, 800, 1000 μg/ml) showed a significant inhibition activity against heat-induced hemolysis of hRBCs (Fig. 1b). The PI of hemolysis within experimental groups was dose dependent and maximum inhibition of hemolysis was observed in 1000 μg/ml dose group (20.86 ± 0.77%). Standard NSAID drug indomethacin showed the maximum inhibition of 43.98 ± 1.52% at 200 μg/ml concentration.
Effect of Aloe vera gel homogenate on protein denaturation-inhibition test
All the dose groups of A. vera gel homogenate showed significant inhibition of protein denaturation in a dose dependent manner (Fig. 1c). In the experimental groups, Aloe dose groups of 600 μg/ml, 800 μg/ml, and 1000 μg/ml showed 34.27 ± 3.86%, 37.82 ± 5.30%, and 39.35 ± 4.25% inhibition of protein denaturation, respectively. The standard NSAID drug diclofenac sodium showed an inhibition of 46.74 ± 1.84% at a concentration of 100 μg/ml.
In vivo anti-arthritic test
The expression of the target and the internal control genes were assessed in the negative control (FCA) and experimental animals (LD and HD) relative to the normal animals (PC) (Figs. 2 and 3). For both the genes, inhibition of expression of target genes in the experimental groups was observed in dose-dependent manner. The TNF-α showed an elevated expression (fold change) in the FCA group (2.271 ± 0.85) compared to PC (1 fold) (Fig. 3a). TNF-α expression was reduced in both the Aloe gel-fed groups (1.456 ± 0.11 and 1.396 ± 0.10 folds in LD and HD groups respectively). Cox-2 increased in FCA-treated animals compared to positive control (1.842 ± 0.68) but decreased in the dose groups (1.259 ± 0.11 and 1.198 ± 0.083 folds for LD and HD groups respectively) (Fig. 3b). It is evident that the expression of TNF-α was decreased by 35.88% and 38.52% in LD and HD groups respectively compared to FCA group animals; Cox-2 expression decreased by 31.65% and 34.96% respectively in LD and HD compared to FCA group animals.

Amplification curves for all the three selected genes in different experimental groups (in duplicates) in reverse transcription quantitative PCR

Results of in vivo anti-inflammatory tests in graph. Relative expression fold changes in the TNF-α (a) and Cox-2 (b) genes assessed through reverse transcriptase real-time quantitative PCR. The significance value of low dose (LD) and high dose (HD) groups are compared with the experimental group (FCA). Positive control (PC). P value at P < 0.01 is considered significant
Discussion
Erythrocyte membrane stability test is a well-established study to screen the possible anti-inflammatory effect of synthetic drugs as well as of various traditional herbal extracts [19]. During inflammation, lysis of the membranes of the lysosomal vesicles occurs that releases their component enzymes which induce the inflammatory response. Thus, a stabilized membrane prevents the release of its contents as well as the progression of inflammation. Non-steroidal anti-inflammatory drugs (NSAIDs) exert their beneficial effects by either stabilizing the lysosomal membranes or by inhibiting the release of lysosomal enzymes [2]. Exposure of RBCs to hypotonic medium or high temperatures results in the lysis of the RBC membranes accompanied by hemolysis and oxidation of hemoglobin. In the hypotonic solution, the hemolytic effect is related to excessive accumulation of fluid within the cell resulting in the rupturing of its membrane. Increased body temperatures also cause the rupture of the RBC membrane resulting in hemolysis [23]. In our study, a dose-dependent relationship of Aloe gel homogenate against both hypotonicity and heat-induced hemolysis has been found. Therefore, both the doses of the plant extract may inhibit the release of lysosomal content during the inflammatory processes. A possible explanation for the membrane stabilizing activity of plant extracts could be an increase in surface area/volume ratio of the cell [24] or by a stabilization of the skeletal proteins such as tropomyosin [25, 26].
In the protein denaturation-inhibition test, the denaturation of egg albumin was induced by heat treatment. Heat exposure causes the breakage of the functional three-dimensional structure of protein. Protein denaturation is a well-documented feature in chronic inflammatory diseases like RA, especially in the severe stages of the disease [27]. Standard NSAID drugs can inhibit heat-induced protein denaturation [4, 21]. In this study, it has been found that Aloe gel homogenate can also inhibit heat-induced protein denaturation. This signifies the anti-inflammatory roles of the plant in vitro and further encourages exploration of its efficacy in inflammatory conditions in vivo.
Cytokines are the key mediators of inflammation. TNF-α is a pro-inflammatory cytokine released from macrophages and monocytes. TNF-α is responsible for the transportation of more immune cells and chemokines to the inflamed region. It also initiates the production of matrix metalloproteinases responsible for cartilage degradation [7]. On the other hand, Cox-2, an immune modulator, is responsible for the increased production of prostaglandin E2 which results in increased pain and swelling at the site of inflammation [9]. Cytokines along with Cox-2 are target bio-molecules of the host body which have a regulatory role on inflammation progression. NSAIDs primarily work on cyclooxygenase pathway [28]. Cytokine inhibitors have been introduced in the medication against inflammation as well.
In the present study, our results show that the A. vera unprocessed gel homogenate downregulates TNF-α as well as Cox-2 expressions in experimental animals in a dose-dependent manner. The crude unprocessed homogenized form of A. vera gel has been orally administered without any post-harvesting processing, a method which is in traditional use in different ethnic communities of Egypt, Rome, Africa, and Asia [29]. In our previous works, we have reported the ameliorative role of unprocessed A. vera gel homogenate in the regulation of inflammatory and arthritic symptoms in experimental rats. The paw circumference, serum biochemical parameters, and blood profile were restored to normal levels after oral feeding of unprocessed Aloe gel homogenate [5, 30]. The present study now further shows that the unprocessed A. vera gel homogenate concomitantly downregulates both TNF-α and Cox-2 in the experimental animal groups. This report can be considered as baseline data describing the efficacy of crude unprocessed plant products against inflammatory arthritis. By not extracting the gel in polar or non-polar solvents, we presume, the natural constituents of A. vera gel have been restored in its natural proportion and thus natural synergistic role of the gel has been monitored. Considering the previous works done in this regard, different bioactive compounds of the gel has already been mentioned by other workers which contribute to the efficacy of the plant as an anti-inflammatory resource. Davis et al. [12] has mentioned that the polysaccharides obtained from Aloe vera are potent anti-inflammatory agents; Hutter et al. [31] identified 8-[C-beta-D-[2-O-(E)-cinnamoyl]glucopyranosyl]-2-[(R)-2-hydroxypropyl]-7-methoxy-5-methylchromone from Aloe barbadensis (Synonym Aloe vera) as anti-arthritic agent. Apart from these findings, anthroquinones were identified as potent anti-inflammatory mediators in Aloe vera by Kashirsagar and co-workers [15]. The TNF-α downregulating activity of the plant has also been documented by Prabjone et al. [32] in helicobacter pylori-infected rats. It is highly expected that the anti-inflammatory properties of the subject plant in model systems will be equally attributed to the human system as well. Our data showed that the crude A. vera gel reduced the extent of lysosomal membrane lysis and protein denaturation in vitro and also regulated the expressions of major pro-inflammatory cytokine TNF-α and a key inflammation modulator Cox-2 in vivo following daily oral consumption in inflammatory RA rat models. To our knowledge, it is the first report on the effect of unprocessed Aloe vera gel oral treatment on TNF-α and Cox-2 gene expressions in arthritic animal models.
Conclusions
The study validates the efficacy of unprocessed aqueous Aloe gel homogenate in inflammatory disease condition. Our data indicate promising anti-inflammatory activity of Aloe gel in the inhibition of lysosomal membrane lysis, protein denaturation in vitro, and downregulation of TNF-α and Cox-2 expression in vivo. The scientific basis of use of raw unprocessed A. vera thus has been explored. However, an expression study on a broader spectrum of cytokines would further clarify the scenario.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- A. vera :
-
Aloe vera
- RA:
-
Rheumatoid arthritis
- NSAIDs:
-
Non-steroidal anti-inflammatory drugs
- CAMs:
-
Complementary and alternative medicines
- WHO:
-
World Health Organization
- LD:
-
Low dose
- HD:
-
High dose
- b.w.:
-
Body weight
- CPCSEA:
-
Committee for the Purpose of Control and Supervision of Experiments on Animals
- IAEC:
-
Institutional Animal Ethical Committee
- ANOVA:
-
One-way analysis of variance
- EDTA:
-
Ethylenediaminetetraacetic acid
References
Chayen J, Bitensky L (1971) Lysosomal enzymes and inflammation with partivular reference to rheumatoid diseases. Ann Rheum Dis 30:522–536
Anosike CA, Obidoa O, LUS E (2012) Membrane stabilization as a mechanism of the anti-inflammatory activity of methanol extract of garden egg (Solanum aethiopicum). DARU J Pharm Sci 20:76
Mizushima Y (1964) Inhibition of pretein denaturation by antirheumatic or antiphlogistic agents. Arch Int Pharmacodyn Ther 149:1–7
Saso L, Valentini G, Casini ML, Grippa E, Gatto MT, Leone MG, Silvestrini B (2001) Inhibition of heat-induced denaturation of albumin by nonsteroidal anti-inflammatory drugs (NSAIDs): Pharmacological implications. Arch Pharm Res 24(2):150–158
Paul S, Dutta T, Chaudhuri TK, Bhattacharjee S (2017) Curative and protective properties of crude gel of Aloe vera from sub-Himalayan West Bengal in chronic and acute inflammatory rat models. Indian J Tradit Knowl 16(1):121–127
Feldmann M, Brennan FM, Maini RN (1996) Role of cytokines in rheumatoid arthritis. Annu Rev Immunol 14:397–440
Saleem A, Saleem M, Akhtar MF, Shahzad M, Jahan S (2020) Moringa rivae leaf extract attenuate Complete Freund’s adjuvant induced arthritis in Wistar rats via modulation of inflammatory and oxidative stress biomarkers. Inflammopharmacol 28:139–151
Crofford LJ (1999) The role of COX-2 in rheumatoid arthritis synovial tissues. Arthritis Res 1(1):S1
Sano H (2011) The role of lipid mediators in the pathogenesis of rheumatoid arthritis. Inflamm Regen 31:151–156
Parasuraman S, Thung SG, Arumugam S (2014) Polyherbal formulation: concept of Ayurveda. Pharm Rev 8(16):73–80
World Health Organization (WHO). General guidelines for methodologies on research and evaluation of traditional medicine. WHO/EDM/TRM 2000.1. http://archives.who.int/tbs/tbs_trm.htmhttp://archives.who.int/tbs/tbs_trm.htm. Accessed 20 July 2019
Davis RH, Donato JJ, Hartman GM, Haas RC (1994) Anti-inflammatory and wound healing activity of a growth substance in Aloe vera. J Am Podiatr Med Assoc 84:77–81
Davis RH, Leitner MG, Russo JM, Byrne ME (1989) Wound healing: oral and topical activity of Aloe vera. J Am Podiatr Med Assoc 79:559–562
Surjushe A, Vasani R, Saple DG (2008) Aloe vera: a short review. Ind J Dermatol 53:163–166
Kashirsagar AD, Panchal PV, Harle UN, Nanda RK, Shaikh HM (2014) Anti-inflammatory and antiarthritic activity of anthroquinone derivatives in rodents. Int J Inf Secur 690596. https://doi.org/10.1155/2014/690596
Rashed AN, Afifi FU, Disi AM (2003) Simple evaluation of the wound healing activity of a crude extract of Portulaca oleracea L. (growing in Jordan) in Mus musculus JVI-1. J Ethnopharmacol 88:131–136
Afolayan AJ (2003) Extracts from the shots of Arctotis arctotoides inhibit the growth of bacteria and fungi. Pharm Biol 41(1):22–25
Paul S, Chakraborty AK, Modak D, Sen A, Bhattacharjee S (2018) In vivo and in silico investigations of the toxicological and analgesic properties of unprocessed Aloe vera gel in experimental rat models. Arch Biol Sci 70(4):727–735
Shinde UA, Phadke AS, Nair AM, Mungantiwar AA, Dikshit VJ, Saraf MN (1999) Membrane stabilizing activity - a possible mechanism of action for the anti-inflammatory activity of Cedrusdeodara wood oil. Fitoterapia 70(3):251–257
Tatiya AU, Saluja AK (2011) Further studies on membrane stabilizing, anti-inflammatory and FCA induced arthritic activity of various fractions of bark of Machilusmacrantha in rats. Rev Bras 21(6):1052–1064
Mizushima Y, Kobayashi M (1968) Interaction of antiinflammatory drugs with serum proteins, especially with some biologically active proteins. J Pharm Pharmacol 20(3):169–173
Gupta AK, Parasar D, Sagar A, Choudhary V, Chopra BS, Garg R, Khatri N (2015) Analgesic and anti-inflammatory properties of gelsolin in acetic acid induced writhing, tail immersion and carrageenan induced paw edema in mice. PLoS One 10(8):e0135558
Aloni B, Eitan A, Livne A (1977) The erythrocyte membrane site for the effect of temperature on osmotic fragility. Biochim Biophys Acta-Biomem 465(1):46–53
Abe H, Katada K, Orita M, Nishikibe M (1991) Effects of calcium antagonists on the erythrocyte membrane. J Pharm Pharmacol 43(1):22–26
Chasis JA, Mohandas N (1986) Erythrocyte membrane deformability and stability: two distinct membrane properties that are independently regulated by skeletal protein associations. J Cell Biol 103(2):343–350
An X, Salomao M, Guo X, Gratzer W, Mohandas N (2007) Tropomyosin modulates erythrocyte membrane stability. Blood 109(3):1284–1288
Rowley M, Tait B, Mackay IR, Cunningham T, Phillips B (1986) Collagen antibodies in rheumatoid arthritis.Significance of antibodies to denatured collagen and their association with HLADR4. Arthritis Rheum 29(2):174–184
Efthimiou P, Kukar M (2010) Complementary and alternative medicine use in rheumatoid arthritis: proposed mechanism of action and efficacy of commonly used modalities. Rheumatol Int 30:571–586
Shedoeva A, Leavesley D, Upton Z, Fan C (2019) Wound healing and the use of medicinal plants. eCAM 2684108:1–30
Guha P, Paul S, Das A, Halder B, Bhattacharjee S, Chaudhuri TK (2014) Analyses of human and rat clinical parameters in Rheumatoid Arthritis raise the possibility of use of crude Aloe vera gel in disease amelioration. Immunome Res 10:081
Hutter JA, Salman M, Stavinoha WB, Satsangi N, Williams RF, Streeper RT, Weintraub ST (1996) Antiinflammatory C-glucosyl chromone from Aloe barbadensis. J Nat Prod 59(5):541–543
Prabjone R, Thong-Ngam D, Wisedopas N, Chatsuwan T, Patumraj S (2006) Anti-inflammatory effects of Aloe vera on leukocyte-endothelium interaction in the gastric microcirculation of helicobacter pylori-infected rats. Clin Hemorheol Microcirc 35(3):359–366
Acknowledgements
The authors acknowledge the expertise of Prof. A. P. Das (Retd.), Department of Botany, University of North Bengal, presently associated with Department of Botany, Rajib Gandhi University, Itanagar, Arunachal Pradesh, for the maintenance of the plant and identification. The authors also thank the Department of Zoology for providing the animal house and the DST-FIST instrumental facilities. Authors also acknowledge the expertise of Dr. Pokhraj Guha (Assistant Professor, Garhbeta College, WB, India) for sharing expertise in molecular biological techniques.
Plant authentication
The identification of the plant species was authenticated by the Department of Botany, University of North Bengal, India, and a voucher specimen was deposited in the Departmental herbarium of the Department of Botany, University of North Bengal bearing the Accession No NBU-09884.
Funding
The research was partially funded by the University Grants Commission, India, under the Basic Scientific Research Fellowship Scheme for Meritorious Students in Science [UGC-BSR Fellowship Scheme File No. 7-134 (2007) awarded to Subhashis Paul].
Author information
Authors and Affiliations
Contributions
S.P, D.M, D.N, A.S, and J.R performed the in vitro experiments, analyzed the data, and prepared the tables and figures; S.P, D.M, and S.C performed the in vivo experiments and prepared and analyzed the data. S.P, D.M, and S.C also prepared the manuscript. T.K.C and S.B contributed to the concept and design of the experiments and data analyses, contributed to the critical revision of the manuscript, and gave the final approval of the manuscript for publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The work was approved by the Institutional Animal Ethical Committee of the University of North Bengal (IAEC, NBU) (Approval number 840/ac/04/CPCSEA).
Consent for publication
Not applicable.
Competing interests
Authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Paul, S., Modak, D., Chattaraj, S. et al. Aloe vera gel homogenate shows anti-inflammatory activity through lysosomal membrane stabilization and downregulation of TNF-α and Cox-2 gene expressions in inflammatory arthritic animals. Futur J Pharm Sci 7, 12 (2021). https://doi.org/10.1186/s43094-020-00163-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43094-020-00163-6