
Abstract
Background
Proprotein convertase subtilisin/kexin type-9 (PCSK9), an enzyme produced mainly by hepatocytes and breaks low-density lipoprotein receptor (LDL-R), inflammatory markers [toll like receptor 4 (TLR4), high mobility group box 1 (HMGB1), tumor necrosis factor alpha (TNFα), c-reactive protein (CRP)], and monocyte subtypes are associated with coronary artery disease (CAD) pathogenesis. The circulating microRNA-218 (miR-218) can relieve CAD through the suppression of HMGB1 in monocyte-derived inflammatory cytokines. Herein, we explored the association between circulatory miR-218 expression and serum levels of PCSK9, inflammatory markers, and monocyte subtypes in statin and non-statin CAD patients. This study involved 91 healthy (control) and 91 stable CAD participants which were subdivided into no-statin (NS, n = 25), low-statin (LS, n = 25), and high-statin (HS, n = 41) groups. low-density lipoprotein cholesterol (LDL-C) and CRP serum levels were calorimetrically determined. Serum levels of PCSK9, TLR4, HMGB1, and TNFα were detected by ELISA, while monocyte subsets [classical (CM), intermediate (IM), non-classical (NC)] were calculated by flow cytometry. Circulatory miR-218 expression was detected by real-time PCR.
Results
The CAD group had significantly lower miR-218 expression and significantly higher levels of PCSK9, inflammatory markers (HMGB1, CRP, TLR4, and TNFα), and IM% than the control group. Among CAD patients, LS and HS groups had significantly lower miR-218 expression, LDL-C levels, and inflammatory markers and significantly higher levels of PCSK9 than the NS group. The HS group exhibited the lowest miR-218 expression and inflammatory markers and the highest PCSK9 levels. However, there were no significant changes in IM% among statin and non-statin groups. In the three CAD groups, miR-218 showed a significantly negative correlation with PCSK9 and inflammatory markers (HMGB1, CRP, TLR4, and TNFα), while this expression exhibited a significantly negative correlation with CM%, IM%, and NCM% only in the NS group. Results of multivariable linear regression indicated a correlation between miR-218 and five independent variables (PCSK9, HMGB1, CRP, TLR4, and TNFα) in the total statin (LS + HS) group, and eight independent variables (PCSK9, HMGB1, CRP, TLR4, and TNFα, CM%, IM%, NCM%) in the NS group. Provided that all other independent variables are constant, miR-218 expression was significantly correlated to CRP (Beta = 0.234) and PCSK9 (Beta = − 0.875) in the total statin group; TLR4 (Beta = − 0.554) in the LS group; HMGB1 (Beta = − 0.507) in the HS group; and CRP (Beta = − 0.745) in the NS group.
Conclusions
Statin-treated CAD patients have a unique negative correlation between miR-218 and PCSK9, HMGB1, and TLR4, and subsequently with CAD progress. Therefore, it could be recommended to combine activators of miR-218 and inhibitors of PCSK9, HMGB1, and TLR4 with statin to efficiently treat CAD.
Similar content being viewed by others
1 Background
Among cardiovascular diseases (CVDs), CAD leads the pack in terms of global death rate [1] with fatalities accounting for 78,897 (21.73%) of all deaths in Egypt in 2013 [2]. Stable CAD applies to patients with diagnosed atherosclerotic disease of the coronary arteries who haven’t had a recent heart attack, stroke, or other major cardiovascular event [3]. The atherosclerotic plaque is considered a hallmark of the pathophysiology of CAD [4]. Fueled by oxidative stress and higher LDL-cholesterol (LDL-C) levels, endothelial damage can also induce plaque formation [5]. When endothelial cells are damaged, LDL-C can be oxidized into Ox-LDL within the artery wall [6]. Ox-LDL accumulation leads to macrophages becoming foam cells, which deposit fat in the artery wall and release inflammatory cytokines [7]. Statins are drugs that lower LDL-C through the blocking of hydroxymethyl-glutaryl-coenzyme A (HMG-CoA) reductase, an enzyme that makes cholesterol in the liver, and overexpressed LDL-R on hepatocytes [8].
In addition to dyslipidemia, many other risk factors associated with atherosclerosis and CAD were recently discovered [9]. One of these risk factors is PCSK9 which is formed by liver cells, where it unites to LDL-R on the same cell surface and regulates its activity [10]. After binding to LDL-R, PCSK9 is taken up by liver cells and broken down in lysosomes. This causes an increase in blood levels of LDL-C while decreasing the quantity of LDL-R on cell surfaces [11]. The greatest risk of cardiovascular disease is linked to hypercholesterolemia caused by PCSK9 gain-of-function mutations [12]. PCSK9 levels in the blood are higher in CVD patients, especially those who are newly diagnosed at the time of acute myocardial infarction [13]. The inflammatory response is positively impacted by PCSK9 because of its role in regulating TLR4 and NF-кB [14]. Statins can stimulate the production of PCSK9 in liver cells [15].
The inflammatory regulator HMGB1 is also linked to atherosclerosis [16]. HMGB1 levels are high in plaque, and blood vessel walls [17] and can trigger the over-release of cytokines, which helps the inflammation continue and makes the plaque grow bigger [18]. When blood vessels are damaged, HMGB1 expression increases in the plaque, thereby recruiting inflammatory cells and leading to the progression of CVD [19]. Toll-like receptors (TLRs) can identify endogenous ligands, including ox-LDL and HMGB1, and so they also engage in atherosclerosis and CVD [20]. TLR4 agonists can induce atherosclerosis in mice [21]. Ox-LDL can elevate TLR4 expression and then activate NF-kB, which triggers TNFα formation [22, 23]. TNFα knockdown mice administered atherogenic food were less susceptible to atherosclerosis [24] and high TNFα blood levels were positively linked to CAD patients [25]. The CRP is a pentraxins protein that is released by endothelial and macrophage cells and seems to be a useful marker and predictor of CVD as it contributes to all stages of atherosclerosis [26]. TNFα, IL-1β, and IL-6 are the primary inductors of the CRP gene [27].
Monocytes are key players in atherosclerosis, as they can stick to and cross the endothelial cells into the artery wall, take up altered lipids, and become foam cells [28]. Depending on their surface markers and as measured by flow cytometry, human monocytes are divided into three groups: classical monocytes (CD14++CD16−; CM), intermediate monocytes (CD14++CD16+; IM), and non-classical monocytes (CD14+CD16++; NCM) [29]. CM have a typical monocyte function (they rise during clotting), IM has a high inflammatory potential (they rise during a heart attack and stroke), and NCM has a special role in patrolling, especially near damaged artery walls (they rise during heart disease) [30]. PCSK9 can cause inflammation in plaque by attracting monocytes [31]. Monocytes have receptors that can bind to ox-LDL to induce plaque formation [32].
MicroRNAs (miRNAs) modulate gene expression by influencing post-transcriptional RNA degradation or translation. More miRNAs in the blood, especially those related to the heart, have been found as new indicators of CAD [33]. MiR-218 expression was low in mice with aortic constriction and CAD patients [34, 35]. MiR-218 could protect against myocardial damage and isoproterenol-induced cardiac hypertrophy in rodents [36]. Bioinformatics research revealed that the miR-218 gene can target TLR4 [37]. MiR-218 may also reduce HMGB1 expression and inflammatory cytokines that come from monocytes, or endothelial cells, thereby diminishing inflammation [38]. All these findings show that miR-218 could be involved in CVD. Nevertheless, the exact mechanism of action by which miR-218 could affect atherosclerosis and CAD remains mostly unknown [37, 39].
As shown above, miR-218 expression, PCSK9 levels, monocyte subtypes, and inflammatory markers (HMGB1, CRP, TLR4, and TNFα) are closely related to CVD/CAD development and outcome, but they vary among patients. However, the correlation between miR-218 and the other CVD/CAD factors is poorly understood, especially in statin and non-statin patients. Therefore, this study aimed to explore how circulatory miR-218 and levels of PCSK9, monocyte subtypes, and inflammatory markers are correlated in statin and non-statin CAD patients.
2 Methods
2.1 Ethics statement and informed consent
The ethics committee of Kafrelsheikh University gave their approval for the study (Approval Code: MKSU 50-3-5). The study complied with the ethical standards of the Helsinki Declaration of the World Medical Association in 2013. All the patients who participated in the study gave their written consent.
2.2 Subjects and study design
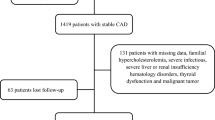
The study was a single-center case–control study conducted at the Cardiology Department of Kafrelsheikh University Hospitals. It involved 182 volunteers, half of whom had atherosclerosis and the other half were healthy controls matched by age and sex. The controls had no cardiovascular disease history, signs, or symptoms and underwent regular health check-ups. The atherosclerosis group consisted of patients older than 45 years who had stable CAD confirmed by elective coronary angiography. Exclusive criteria included (1) acute coronary syndrome (ACS) patients; (2) patients who had percutaneous coronary intervention (PCI) in the past 90 days; (3) patients with cardiac failure and severe valve stenosis; and (4) patients with inflammatory diseases. The atherosclerosis group was further subdivided into three groups based on statin therapy and ESC/EAS lipid guidelines: NS group (no statin prescription by the physician); LS group [low statin (simvastatin 10 mg, pravastatin 5–20 mg, lovastatin 20 mg, fluvastatin 20–40 mg, or pitavastatin 1 mg)]; and HS group [high statin (atorvastatin 40 to 80 mg daily or rosuvastatin 20 to 40 mg daily).
2.3 Blood sampling and serum collection
Before coronary angiography in the morning, blood was collected from the antecubital vein in the elbow area of fasting participants in tubes coated with EDTA and plain tubes. Flow cytometry was performed on whole blood samples right away. Sera were separated by spinning the blood samples from the vacutainer tubes at 3000 rpm at 4 °C and were kept at − 80 °C for future analysis.
2.4 Serum biochemical assays
Serum levels of the lipid profile parameters [cholesterol, triglycerides, LDL-C, and high-density lipoprotein-cholesterol (HDL-C)] and CRP were measured using a chemistry autoanalyzer (Indiko, ThermoFisher Scientific, USA).
2.5 ELISA
A specific ELISA kit (Bioassay Technology Laboratory, Shanghai, China, Cat # E2320Hu) was used to measure PCSK9 serum levels following the manufacturer’s instructions. The kit had an assay range of 1 to 300 ng/mL with inter-assay and intra-assay coefficients of variance of CV < 12% and CV < 10%, respectively. The serum levels of the inflammatory markers TLR4 (MyBioSource, San Diego, CA, USA, Cat # MBS2702401; CV of intra-assay and inter-assay precision = 10% and 12%), TNFα (MyBioSource, Cat # MBS2502004; CV of intra-assay and inter-assay precision = 5.5% and 4.5%), and HMGB1 (MyBioSource, Cat # MBS701378; CV of intra-assay and inter-assay precision = 9% and 8%) were quantified using specific human ELISA kits as detailed in the manufacturer’s guidelines. The ODs of all samples were determined by EL800, Bioelisa ELISA Reader (Biokit, USA).
2.6 Molecular analysis by real-time PCR
The total RNA from serum samples was extracted using Trizol (Invitrogen) according to the manufacturer’s protocol. A Nanodrop (Quawell, Q3000) was used to measure the concentration and purity of the total RNA. The total RNA was converted to cDNA using reverse transcriptase (RevertAid H Minus, Thermo Scientific, Cat # EP0451, USA). The qPCR was performed in a real-time thermal cycler (Step OnePlus, Applied Biosystems) with cDNA, primers, and syber green master mix (QuantiTect SYBR Green qPCR, Thermo Scientific) following the reaction cycles described previously [40]. The relative expression was calculated using the 2−∆∆Ct equation. U6 was the internal control for the fold change of gene expression. The sequence of primers was as follow: F: 5′‐CGCCGCCTGTGCGTGAGGAT‐3′; R: 5′‐GCCTAGTTTGACGAGAGGT‐3′ for miR-218 and F: 5′‐CTCGCTTCGGCAGCACA‐3′; F: 5′‐CTCGCTTCGGCAGCACA‐3′ for U6 [39].
2.7 Flow cytometry analysis
Monocyte subsets were calculated by flow cytometry as previously detailed [41]. In brief, 100 μl whole blood containing up to 106 nucleated cells were mixed gently with 10 μl of each monoclonal Abs then 2 ml of the lysis buffer (BD FACS, Biosciences) were added followed by incubation for 10 min. After centrifugation (2500 rpm/6 min), the cells were fixed in 1% paraformaldehyde in PBS. The mAbs used were allophycocyanin (APC)-H7-labeled mAb for CD14 (Elabscience, Houston, Texas, USA, Cat# E-AB-F1209E), and fluorescein isothiocyanate (FITC)-labeled mAb for CD16 (Elabscience, Cat# E-AB-F1005C). We used a FACSCantoII flow cytometer equipped with BD FACSDiva Software 9.0 (Becton Dickinson, Biosciences, San Jose, CA, USA) to count labeled cells. Scatter characteristics were used to identify and gate monocytes as previously detailed [41].
2.8 Statistical analysis
Data were analyzed by SPSS 21.0 software (IBM SPSS Inc., Chicago, IL) and GraphPad Prism 8 Software (GraphPad Inc., LaJolla, CA, USA). According to the Kolmogorov–Smirnov test, the data followed a normal distribution. The χ2 test was used to assess the enumeration data. A one-way analysis of variance (ANOVA) followed by Tukey's Honestly Significant Difference (Tukey’s HSD) was used when comparing three or more groups, whereas the t-test was used when comparing two groups. The correlation between groups was determined by Pearson’s coefficient. Multivariable linear regression models were calculated for the association of miR-218 expression, PCSK9 levels, monocyte subtypes, and inflammatory markers (HMGB1, CRP, TLR4, and TNFα). The data were presented as mean ± standard deviation (SD) or standard error of the mean (SEM) and significance was declared at p ≤ 0.05.
3 Results
3.1 Baseline characteristics of participants
Table 1 presents an overview of the fundamental information and clinical characteristics of both the healthy population and CAD patients. Among the 91 patients diagnosed with stable CAD, there were 59 males and 32 females, with a mean age of 57.59 ± 10.41 years. Comparatively, in the group of 91 healthy controls, there were 54 males and 37 females, with a mean age of 56.10 ± 8.25 years. Notably, there were no significant (p > 0.05) differences observed in baseline characteristics, including age, gender, hypertension, BMI, diabetes mellitus, smoking, and blood lipid profile (triglycerides, total cholesterol, and HDL-C), between the two groups. These results provide more evidence that the baseline features of the control and CAD groups were similar.
3.2 Comparison of molecular, biochemical parameters and monocyte subsets in healthy controls and CAD patients
We first compared the molecular (miR-218 expression), biochemical (LDL-C, CRP, PCSK9, TLR4, HMGB1, and TNFα), and percentage of monocytes subsets [classical (CM), intermediate (IM), non-classical (NCM)] in the health controls and the CAD group. The results exhibited that the CAD group had significantly (p ≤ 0.0001) lower miR-218 expression and significantly higher levels of PCSK9 (p ≤ 0.0001), inflammatory markers [HMGB1 (p ≤ 0.05), CRP (p ≤ 0.05), TLR4 (p ≤ 0.0001), and TNFα (p ≤ 0.0001)], and IM% (p < 0.0001) than the control group. However, no significant (p > 0.05) difference was found in CM%, NCM%, and LDL-C between the two groups (Fig. 1).

Comparison of molecular (miR-218 expression), biochemical parameters (LDL-C, CRP, PCSK9, TLR4, HMGB1, and TNFα) and monocytes subsets (CM%, IM%, and NCM%) in controls and CAD patients as analyzed by t-test. Relative expression of miR‐218 was determined in peripheral blood by real-time PCR. Serum levels of LDL-C, CRP, PCSK9, TLR4, HMGB1, and TNFα were detected by ELISA. CM%, IM%, and NCM% were quantified using flow cytometry as will be detailed in Fig. 3. Data were expressed as mean ± SD, n = 91/group. Significant levels were set at *p ≤ 0.05, and ****p ≤ 0.0001. CAD Coronary artery disease, Cnt Control
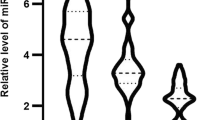
Next, we compared these parameters in healthy controls and the three subgroups of the CAD groups: no-statin (NS, n = 25), low-statin (LS, n = 25), and high-statin (HS, n = 41) groups. As presented in Fig. 2, LS and HS groups had significantly (p ≤ 0.0001) lower miR-218 expression, LDL-C levels, and inflammatory markers (HMGB1, CRP, TLR4, and TNFα) and significantly higher levels of PCSK9 than NS. The HS group exhibited significantly lower miR-218 expression (p ≤ 0.0001), LDL-C levels (p ≤ 0.05), inflammatory markers [HMGB1 (p ≤ 0.0001), CRP (p ≤ 0.05), TLR4 (p ≤ 0.0001), and TNFα (p ≤ 0.0001)] and significantly higher PCSK9 levels (p ≤ 0.0001) than the LS group. However, no significant (p > 0.05) changes in IM% among statin (LS or HS) and NS groups. Figure 3 shows representative flow cytometry scattered plots distinguishing % of monocyte subset in each group.

Comparison of molecular (miR-218 expression), biochemical parameters (LDL-C, CRP, PCSK9, TLR4, HMGB1, and TNFα) and monocytes subsets (CM%, IM%, and NCM%) in controls and the three subgroups of CAD patients (NS, LS, and HS) as analyzed by one way ANOVA. Data were presented as mean ± SD. Number of samples were 91, 25, 25, and 41 in the control, NS, LS, and HS groups, respectively. Values with different superscript letters, with “a” and “d” referring to the highest and lowest values, respectively, were significantly different at p ≤ 0.05. All groups were compared to each other. Cnt Control group, HS High stain group, LS Low stain group, HS High stain group

Representative flow cytometry scattered plots distinguishing % of monocyte subset in each group. In each group, the left image refers to the forward (FSC-A) and sideward (SSC-A) scatter profile while the right image displays monocyte subsets based on CD14 and CD16 surface markers. HS High statin group, LS Low statin group, NS No-statin group
3.3 Correlation between miR-218 expression and other CAD-related parameters in statin and non-statin CAD patients
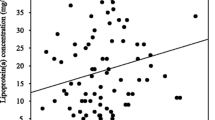
Figures 4 and 5 depict the results of the correlation between miR-218 expression and CAD-related biochemical parameters, including LDL-C, CRP, PCSK9, TLR4, HMGB1, TNFα, and monocytes subsets, using the Pearson correlation coefficient. In general, miR-218 expression significantly and negatively correlated with levels of PCSK9, HMGB1, CRP, TLR4, and TNFα in the three CAD groups, while this expression exhibited a significantly negative correlation with CM%, IM%, and NCM% only in the NS group. As detailed in Fig. 4, miR-218 expression significantly and negatively correlated with levels of PCSK9 (r = − 0.679, p ≤ 0.001 in the NS group; r = − 0.5048, p ≤ 0.05 in the LS group; r = − 0.3934, p ≤ 0.05 in the HS group), levels of HMGB1 (r = − 0.6452, p ≤ 0.001 in the NS group; r = − 0.4095, p ≤ 0.05 in the LS group; r = − 0.5072, p ≤ 0.001 in the HS group), levels of CRP (r = − 0.6568, p ≤ 0.001 in the NS group; r = − 0.4535, p ≤ 0.05 in the LS group; r = − 0.4524, p ≤ 0.01 in the HS group), and levels of TLR4 (r = − 0.6632, p ≤ 0.001 in the NS group; r = − 0.5539, p ≤ 0.01 in the LS group; r = − 0.4760, p ≤ 0.01 in the HS group). However, no significant correlation was noticed between miR-218 expression and LDL-C (r = − 0.4095, p ≥ 0.05 in the NS group; r = − 0.04923, p ≥ 0.05 in the LS group; and r = 0.2358, p ≥ 0.05 in the HS group).

Correlation between miR-218 expression and CAD-related parameters (LDL-C, CRP, PCSK9, TLR4, HMGB1) in NS, LS, and HS subgroups of CAD patients as analyzed by Person correlation. The number of samples was 91, 25, 25, and 41 in the control, NS, LS, and HS groups, respectively. Cnt Control group, HS High stain group, LS Low stain group, HS High stain group

Correlation between miR-218 expression and CAD-related parameters (TNFα, CM%, IM%, and NCM%) in NS, LS, and HS subgroups of CAD patients as analyzed by Person correlation. The number of samples was 91, 25, 25, and 41 in the control, NS, LS, and HS groups, respectively. Cnt Control group, HS High stain group, LS Low stain group, HS High stain group
As depicted in Fig. 5, miR-218 expression also significantly and negatively correlated with levels of TNFα (r = − 0.7379, p ≤ 0.0001 in the NS group; r = − 0.4060, p ≤ 0.05 in the LS group; r = − 0.4473, p ≤ 0.01 in the HS group). Regarding the correlation between miR-218 expression and monocytes subsets (Fig. 5), only the NS group exhibited significantly negative correlations between miR-218 expression and CM% (r = − 0.4993, p ≤ 0.05), IM% (r = − 0.5835, p ≤ 0.01), and NCM% (r = − 0.5860, p ≤ 0.01). However, in the statin groups (LS and HS) miR-218 expression did not significantly correlate with CM% (r = − 0.05696, p ≥ 0.05 in the LS group; r = − 0.1954, p ≥ 0.05 in the HS group), IM% (r = 0.03971, p ≥ 0.05 in the LS group; r = 0.2334, p ≥ 0.05 in the HS group) or NCM% (r = 0.2808, p ≥ 0.05 in the LS group; and r = − 0.01412, p ≥ 0.05 in the HS group).
3.4 Association between miR-218 expression and other CAD-related parameters in statin and non-statin CAD patients
The results of multivariable linear regression between miR-218 expression and the serum levels of the five independent variables (PCSK9, HMGB1, CRP, TLR4, and TNFα) in the total statin (LS + HS) group were presented in Table 2. These data revealed the existence of a multiple correlation between the miR-218 and the five independent variables (R = 0.884, R2 = 0.782, F = 42.979, p ≤ 0.01). Moreover, there was a significant association between miR-218 and PCSK9 levels (Beta = − 0.875, p ≤ 0.01) suggesting that PCSK9 serum levels could affect miR-218 expression in the total statin (LS + HS) group provided that all other independent variables (HMGB1, CRP, TLR4, and TNFα were constant. Additionally, we used progressive multiple regression analysis (simplified model) to find out the most influential independent variables on the miR-218 expression in the total statin group and found that serum levels of significantly PCSK9 and CRP correlated with miR-218 expression (R = 0.883, F = 111.281, p ≤ 0.01). The value of the determination coefficient (R2) was 0.779 which means that these two variables alone explain 77.9% of the miR-218 expression variance (72.4% due to PCSK9 and 5.5% due to CRP) where PCSK9 had a significant negative effect (Beta = − 0.854, p ≤ 0.01), and CRP had a significant positive effect (Beta = 0.234, p ≤ 0.01) on the miR-218 expression (Table 2).
In the NS group (Table 3), we found a multiple correlation between miR-218 expression and the serum levels of eight independent variables, PCSK9, HMGB1, CRP, TLR4, TNFα, CM%, IM%, and NCM%, (R = 0.817, R2 = 0.668, F = 4.022, p ≤ 0.01) with CRP being the most influential independent variable according to progressive multiple regression analysis (R = − 0.745, R2 = 0.556, F = 28.747**, p ≤ 0.01). These results infer that CRP alone explains 55.6% of the miR-218 expression variance and it had a negative effect on this expression.
In the HS group (Table 4), miR-218 expression was associated with the serum levels of five independent variables, PCSK9, HMGB1, CRP, TLR4, and TNFα (R = 0.533, R2 = 0.306, F = 3.086, p ≤ 0.05) with HMGB1 being the most influential independent variable according to progressive multiple regression analysis (R = 0.507, R2 = 0.257, F = 13.506, p ≤ 0.01) which means that HMGB1 alone explains 25.7% of the miR-218 expression variance and it had a significant negative effect on this expression (Beta = − 0.507, p ≤ 0.01). However, in the LS group (Table 5), TLR4 was the most influential independent variable (R = 0.554, R2 = 0.307, F = 10.179, p ≤ 0.01) suggesting that TLR4 alone could explain 30.7% of the miR-218 expression variance and it had a negative effect on this expression (Beta = − 0.554, p ≤ 0.01).
4 Discussion
It has been shown that miR-218 has positive effects on the cardiovascular system such as promoting endothelial cell renewal and blood vessel formation [42], and reducing inflammation [39]. Other factors such as PCSK9, LDL-C, monocyte subtypes, and inflammatory markers (HMGB1, CRP, TLR4, and TNFα) are also linked to CAD development, and they vary depending on whether the patients take statins or not [41, 43, 44]. The relationship between miR-218 and these other CAD factors is not well studied, particularly in patients who use or do not use statins. Thus, this study aimed to investigate the relationship between circulatory miR-218 expression and serum levels of PCSK9, monocyte subtypes, and inflammatory markers in statins and non-statins CAD patients. Overall, our findings showed that CAD patients who received statins had a distinct negative relationship between miR-218 expression and levels of PCSK9, HMGB1, and TLR4, and thus with the advancement of atherosclerosis. Figure 6 summarizes inflammatory mechanisms involved in atherosclerosis and miR-218 effect.

Inflammatory mechanisms involved in atherosclerosis and miR-218 effect. In the presence of circulating PCSK9, the LDL-R-LDL-C complex is internalized into endothelial cells (ECs) and undergoes lysosomal degeneration. PCSK9, HMGB1, and Ox-LDL stimulate and bind to TLR4, which induces the nuclear translocation of NF-κB in ECs thus increasing monocyte infiltration and the expression and secretion of proinflammatory cytokines such as TNFα. TNFα influences the production and secretion of CRP and PCSK9 in the liver creating an endless loop of atherosclerotic inflammation. LDL-C undergoes oxidation in the intima (Ox-LDL) and endothelial activation differentiates monocytes into macrophages with scavenger receptors (SRs) on their surface. PCSK9 upregulates the expression of SRs. Ox-LDL is internalized into macrophages through SRs under the influence of proinflammatory cytokines such as TNFα. Ox-LDL accumulation transforms macrophages into foam cells, releasing more proinflammatory cytokines (TNFα) further augmenting atherosclerosis and inflammation. MiRNA-218 can relieve CAD symptoms and atherosclerosis via suppressing HMGB1 and TLR4 and our study illustrated a novel negative correlation between miRNA 218 and PCSK9 in statin-treated CAD patients. ECs Endothelial cells, LDL-C Low-density lipoprotein cholesterol, LDL-R Low-density lipoprotein receptor, PCSK9 Protein proprotein convertase subtilisin/kexin 9, Ox-LDL Oxidized low-density lipoprotein, TNFα Tumor necrosis factor α, NF-κB Nuclear factor-kappa B, CRP C reactive protein, TLR4 Toll-like receptor 4, SRs Scavenger receptors, HMGB1 High mobility group box 1, miR-218 MicroRNA-218
We first compared the levels of CAD-related parameters miR-218, PCSK9, LDL-C, CRP, monocyte subtypes (CM%, IM%, and NCM%), TLR4, HMGB1, and TNFα between healthy controls and CAD patients. CAD patients exhibited significantly higher serum levels of PCSK9, CRP, IM%, TLR4, HMGB1, and TNFα and significantly lower expression of miR-218 than the healthy controls. However, no significant difference was noticed in LDL-C levels, CM%, and NCM%. Many other previous studies support these findings [38, 39, 41, 43, 44].
We then split the CAD group into three subgroups according to statin treatment: non-statin (NS), low statin (LS), and high statin (HS) groups. We wanted to see if statin could affect the results of these 10 parameters. Indeed, we found some differences between the NS and statin (LS and HS) groups. LS and HS groups of CAD patients showed significantly reduced miR-218 expression, LDL-C levels, and inflammatory markers (CRP, TLR4, HMGB1, and TNFα), and significantly increased PCSK9 levels compared to the NS group. The HS group had the least miR-218 expression and inflammatory markers, and the most PCSK9 levels. The IM% results, however, did not differentiate statin users from non-statin users. Similarly, Krychtiuk, et al. [41] reported that CAD patients receiving statins possessed higher PCSK9, and lower LDL-C levels, with similar IM% compared to those who did not receive statins. Despite this, they failed to notice a noteworthy variation in CRP or TNFα. Moreover, various research demonstrated how statins affect miRNAs [45, 46]. Three microRNAs were overexpressed after hypercholesterolemic patients received atorvastatin [47]. However, Saavedra, et al. [48] and Zambrano, et al. [49] have found downregulated expression of miR20a-5p, miR-17-3p, and miR-590-5p following atorvastatin treatment in high-cholesterol patients. We and Gao, et al. [39] also found a downregulation of miR-218 in CAD individuals. We also found that miR-218 is more reduced in statin-treated patients which is a new novel finding.
Previous studies explored the relationship between LDL-C, inflammatory markers (CRP, TLR4, HMGB1, and TNFα), PCSK9, and CAD development in statins and non-statins patients. However, the correlation between miR-218 and each of these factors in statin and non-statin patients is poorly understood. In all three CAD groups, we observed a negative association between miR-218 expression and PCSK9 and inflammatory marker levels (CRP, TLR4, HMGB1, and TNFα). Keeping all other independent variables fixed, miR-218 expression was significantly associated with levels of PCSK9 in the total statin (LS + HS) group. Progressive multiple regression analysis revealed one significant variable (TLR4) in the LS group and one significant variable (HMGB1) in the HS group. These data infer that statins may influence miR-218 and its relationship with other CAD-associated factors in CAD patients. This could be attributed to statin's anti-inflammatory potential [50]. Statins can repress NF-κB and its downstream inflammatory cytokines such as IL-1β, IL-6, and TNFα [51]. Statins can also inhibit the formation of the inflammatory marker CRP [52].
In the total statin group, the multivariable linear regression analysis showed that Beta for PCSK9 was significant which means that PCSK9 serum levels could affect the expression of circulatory miRNA-218 provided that all other independent variables are constant. According to the progressive multiple regression analysis (stepwise) in the total statin group, PCSK9 only accounted for 72.4% of miR-218 variance, and PCSK9 was the most important negative independent factor that influenced miR-218 in statin-treated patients among other CAD-related factors. This strong negative relationship might result from statin intake, as more statins raise PCSK9 and lower miR-218, or due to the contrasting effects of miR-218 and PCSK9 on the target inflammatory pathway (CRP, TLR4, HMGB1, and TNFα), which they inhibited and activated, respectively. One mechanism by which statins lower LDL-C levels is by raising hepatocyte surface LDL-R expression; however, this action is accompanied by a rise in SREBP-2 activity, which in turn increases circulating PCSK9 protein [53]. Our study showed that statins not only increase PCSK9 but may also decrease miR-218 expression which may be due to the negative effect that PCSK9 has on miR-218 expression. A remaining question is how PCSK9 causes an inflammatory response. People with systemic inflammatory disorders have been found to have elevated PCSK9 levels [54]. PCSK9 triggers the production of proinflammatory cytokines by activating TLR4, which then switches on and moves NF-кB, a transcription factor for many inflammatory cytokines, to the nucleus [55]. All of which may further explain the negative effect that PCSK9 has on miR-218 expression as both of them work on the same inflammatory pathways one inhibiting (miR-218) via blocking TLR4 [37] and the other one (PCSK9) by activating TLR4 and its downstream target NF-кB [14, 43]. PCSK9 can also induce inflammation through the activation of scavenger receptors in monocytes, which in turn increases the absorption of the inflammatory marker ox-LDL [43]. PCSK9-induced inflammation affects vascular stability and activates atherosclerotic plaques which attract and infiltrate more monocytes from the blood (Punch et al., 2022; Ruscica et al., 2019). PCSK9’s inflammatory effects on CAD development are not related to LDL-R regulation only [44].
Interestingly, we also found that TLR4 is the most influential negative cofactor on miR-218 expression in the LS group as evidenced by progressive multiple regression analysis which revealed that TLR4 alone explains 30.7% of miR-218 variance. In support, miR-218 was downregulated in CAD patients, and it has the potential to decrease plaque through inhibition of TLR4 [37]. On the other hand, statins can down-regulate TLR4 and NF-ĸB activity, resulting in less secretion of inflammatory cytokines such as TNFα, IL-6, and IL-1β [56]. Remarkably, low-intensity statin did not completely diminish the relationship between TLR4 and miR-218 but high-intensity statin did. In support, atorvastatin (high-intensity statin) affects atherosclerotic plaque by regulating lipid levels and inhibiting the TLR4/NF-kΒ signaling pathway [57].
The HS group’s progressive multiple regression analysis revealed that only HMGB1 accounted for 25.7% of miR-218 variance and had a negative impact on it. In support, Gao et al. [39] suggested that miR-218 may block HMGB1 thus reduce cardiac microvascular endothelial cells injury induced by coronary artery disease. HMGB1 is an inflammatory mediator that is involved in many cardiovascular diseases including atherosclerosis [58,59,60]. The release of pro-inflammatory mediators, such as TNFα, IL-8, and CRP can be induced by HMGB1 [17, 61]. HMGB1 inflammatory effect is mediated through its receptors such as TLR4 [62]. The anti-inflammatory impact of statins against the heart and vascular damage is mediated by inhibiting HMGB1 release [63, 64]. Statins can inhibit the formation of NLRP3 inflammasome by blocking HMGB1 release [65]. An explanation for HMGB1 being more dominant than TLR4 in the HS group is that a high statin dose suppresses TLR4. In support, Shen et al. [57] reported that atorvastatin affects the TLR4/NF-kB signaling pathway, lowers TLR4 production, controls lipid levels, and impacts atherosclerotic plaque. Yang et al. [56] also found that atorvastatin can suppress TLR4 at both the transcriptional and protein level resulting in declined inflammatory cytokines from LPS-treated monocytes. We also observed a declined TNFα concentration in the HS group. On the other hand, progressive multiple regression analysis in the NS group showed that the inflammatory marker CRP alone explains 55.6% of the miR-218 effect and it had a negative effect on it. These outcomes might be linked to the anti-inflammatory potential of miR-218 as supported by Gao, et al. [39] who discovered that miR-218 expression reduced inflammation in CMEC cells damaged by CAD by suppressing HMGB1 expression.
There is substantial evidence that statin medication can reduce cholesterol levels, but there are some disadvantages when used as a sole therapy [66]. Long-lasting statin doses could cause some side effects such as muscle pains in some patients [67, 68]. Some other patients had a lower response and a higher cardiovascular risk with significant atheroma growth [69]. Because of these side effects, finding novel methods to reduce lipids is essential. PCSK9 reduces statin effectiveness by inducing lysosomal degradation of LDL-R [70]. Consequently, lowering lipids may be best accomplished with a combination of statins and PCKS9 inhibitors [41]. We found that miR-218 in statin-treated CAD individuals have a unique negative correlation with PCSK9, HMGB1, and TLR4, and subsequently with CAD progress. Therefore, it could be recommended to combine activators of miR-218 and inhibitors of PCSK9, HMGB1, and TLR4 with statin to efficiently treat CAD.
5 Conclusions
Statin-treated CAD patients have a unique negative correlation between miR-218 expression and levels of TLR4, HMGB1, and PCSK9, and subsequently with atherosclerosis progress. This result implies that miR-218 agonists and inhibitors of PCSK9, TLR4, and HMGB1 could be a promising treatment approach for CAD patients.
Availability of data and materials
The data supporting the present findings are contained within the manuscript.
Abbreviations
- CAD:
-
Coronary artery disease
- CM:
-
Classical monocytes
- CRP:
-
C-reactive protein
- CVDs:
-
Cardiovascular diseases
- HMGB1:
-
High mobility group box one
- HS:
-
High statin
- IM:
-
Intermediate monocytes
- LDL-R:
-
Low-density lipoprotein receptors
- LS:
-
Low statin
- miR-218:
-
MicroRNA-218
- NC:
-
Non-classical monocytes
- NS:
-
No-statin
- Ox-LDL:
-
Oxidized LDL
- PCSK9:
-
Proprotein convertase subtilisin/kexin type 9
- qPCR:
-
Real-time PCR
- SRs:
-
Scavenger receptors
- TLR4:
-
Toll-like receptor 4
- TNFα:
-
Tumor necrosis factor-alpha
References
Benjamin EJ, Blaha MJ, Chiuve SE, Cushman M, Das SR, Deo R, Ferranti SDD, Floyd J, Fornage M, Gillespie C et al (2017) Heart disease and stroke statistics—2017 update: a report from the american heart association. Circulation 135:e146–e603
El-Moselhy EA, Mohammed A, El-Aziz AAA, Sadek IM, Hagrass SAA, Farag GA (2018) Coronary artery disease among elderly Egyptian patients: I. Socio-demographic, lifestyle, psychosocial, medical, and biochemical risk factors. Am J Gerentol Geriatr 1(2):1006
Fox KAA, Metra M, Morais J, Atar D (2020) The myth of “stable” coronary artery disease. Nat Rev Cardiol 17:9–21
Tibaut M, Mekis D, Petrovic D (2017) Pathophysiology of myocardial infarction and acute management strategies. Cardiovasc Hematol Agents Med Chem 14:150–159
Rueckschloss U, Galle J, Holtz J, Zerkowski H-R, Morawietz H (2001) Induction of NAD(P)H oxidase by oxidized low-density lipoprotein in human endothelial cells. Circulation 104:1767–1772
Tavafi M (2013) Complexity of diabetic nephropathy pathogenesis and design of investigations. J Renal Inj Prev 2:59–62
Ross R (1999) Atherosclerosis—an inflammatory disease. N Engl J Med 340:115–126
Tobert JA (2003) Lovastatin and beyond: the history of the HMG-CoA reductase inhibitors. Nat Rev Drug Discov 2:517–526
Takahashi M (2022) NLRP3 inflammasome as a key driver of vascular disease. Cardiovasc Res 118:372–385
Seidah NG, Awan Z, Chrétien M, Mbikay M (2014) PCSK9: a key modulator of cardiovascular health. Circ Res 114:1022–1036
Shapiro MD, Fazio S (2017) PCSK9 and atherosclerosis—lipids and beyond. J Atheroscler Thromb 24:462–472
Humphries SE, Whittall RA, Hubbart CS, Maplebeck S, Cooper JA, Soutar AK, Naoumova R, Thompson GR, Seed M, Durrington PN et al (2006) Genetic causes of familial hypercholesterolaemia in patients in the UK: relation to plasma lipid levels and coronary heart disease risk. J Med Genet 43:943–949
Almontashiri NAM, Vilmundarson RO, Ghasemzadeh N, Dandona S, Roberts R, Quyyumi AA, Chen H-H, Stewart AFR (2014) Plasma PCSK9 levels are elevated with acute myocardial infarction in two independent retrospective angiographic studies. PLoS ONE 9:e106294
Punch E, Klein J, Diaba-Nuhoho P, Morawietz H, Garelnabi M (2022) Effects of PCSK9 targeting: alleviating oxidation, inflammation, and atherosclerosis. J Am Heart Assoc 11:e023328
Dubuc G, Chamberland A, Wassef H, Davignon J, Seidah NG, Bernier L, Prat A (2004) Statins upregulate PCSK9, the gene encoding the proprotein convertase neural apoptosis-regulated convertase-1 implicated in familial hypercholesterolemia. Arterioscler Thromb Vasc Biol 24:1454–1459
Li W, Sama AE, Wang H (2006) Role of HMGB1 in cardiovascular diseases. Curr Opin Pharmacol 6:130–135
Inoue K, Kawahara K, Biswas KK, Ando K, Mitsudo K, Nobuyoshi M, Maruyama I (2007) HMGB1 expression by activated vascular smooth muscle cells in advanced human atherosclerosis plaques. Cardiovasc Pathol 16:136–143
Kalinina N, Agrotis A, Antropova Y, DiVitto G, Kanellakis P, Kostolias G, Ilyinskaya O, Tararak E, Bobik A (2004) Increased expression of the DNA-binding cytokine HMGB1 in human atherosclerotic lesions: role of activated macrophages and cytokines. Arterioscler Thromb Vasc Biol 24:2320–2325
de Souza AW, Westra J, Limburg PC, Bijl M, Kallenberg CG (2012) HMGB1 in vascular diseases: its role in vascular inflammation and atherosclerosis. Autoimmun Rev 11:909–917
Andreakos E, Foxwell B, Feldmann M (2004) Is targeting Toll-like receptors and their signaling pathway a useful therapeutic approach to modulating cytokine-driven inflammation? Immunol Rev 202:250–265
Mullick AE, Tobias PS, Curtiss LK (2005) Modulation of atherosclerosis in mice by Toll-like receptor 2. J Clin Investig 115:3149–3156
Oikonomou E, Tsaplaris P, Anastasiou A, Xenou M, Lampsas S, Siasos G, Pantelidis P, Theofilis P, Tsatsaragkou A, Katsarou O et al (2022) Interleukin-1 in coronary artery disease. Curr Top Med Chem 22:2368–2389
Xu XH, Shah PK, Faure E, Equils O, Thomas L, Fishbein MC, Luthringer D, Xu XP, Rajavashisth TB, Yano J et al (2001) Toll-like receptor-4 is expressed by macrophages in murine and human lipid-rich atherosclerotic plaques and upregulated by oxidized LDL. Circulation 104:3103–3108
Canault M, Peiretti F, Mueller C, Kopp F, Morange P, Rihs S, Portugal H, Juhan-Vague I, Nalbone G (2004) Exclusive expression of transmembrane TNF-alpha in mice reduces the inflammatory response in early lipid lesions of aortic sinus. Atherosclerosis 172:211–218
Kaptoge S, Seshasai SRK, Gao P, Freitag DF, Butterworth AS, Borglykke A, Di Angelantonio E, Gudnason V, Rumley A, Lowe GDO et al (2013) Inflammatory cytokines and risk of coronary heart disease: new prospective study and updated meta-analysis. Eur Heart J 35:578–589
Zhang W, Speiser JL, Ye F, Tsai MY, Cainzos-Achirica M, Nasir K, Herrington DM, Shapiro MD (2021) High-sensitivity c-reactive protein modifies the cardiovascular risk of lipoprotein(a): multi-ethnic study of atherosclerosis. J Am Coll Cardiol 78:1083–1094
Devaraj S, Jialal I (2011) C-reactive protein polarizes human macrophages to an M1 phenotype and inhibits transformation to the M2 phenotype. Arterioscler Thromb Vasc Biol 31:1397–1402
Ginhoux F, Jung S (2014) Monocytes and macrophages: developmental pathways and tissue homeostasis. Nat Rev Immunol 14:392–404
Ziegler-Heitbrock L, Ancuta P, Crowe S, Dalod M, Grau V, Hart DN, Leenen PJM, Liu Y-J, MacPherson G, Randolph GJ et al (2010) Nomenclature of monocytes and dendritic cells in blood. Blood 116:e74–e80
Kapellos TS, Bonaguro L, Gemünd I, Reusch N, Saglam A, Hinkley ER, Schultze JL (2019) Human monocyte subsets and phenotypes in major chronic inflammatory diseases. Front Immunol 10:2035
Giunzioni I, Tavori H, Covarrubias R, Major AS, Ding L, Zhang Y, DeVay RM, Hong L, Fan D, Predazzi IM et al (2016) Local effects of human PCSK9 on the atherosclerotic lesion. J Pathol 238:52–62
Steinberg D (1997) Low density lipoprotein oxidation and its pathobiological significance. J Biol Chem 272:20963–20966
Labbaf A, Ghaedi H, Alipoor B, Omrani MD, Kazerouni F, Shanaki M, Ghaffarzadeh M, Pashaiefar H, Rahimipour A (2017) The pre-mir-499 variant rs3746444 may contribute to coronary artery disease susceptibility: a case-control and meta-analysis study. Clin Lab 63:587–595
Sayed D, Hong C, Chen IY, Lypowy J, Abdellatif M (2007) MicroRNAs play an essential role in the development of cardiac hypertrophy. Circ Res 100:416–424
Thum T, Galuppo P, Wolf C, Fiedler J, Kneitz S, van Laake LW, Doevendans PA, Mummery CL, Borlak J, Haverich A et al (2007) MicroRNAs in the human heart: a clue to fetal gene reprogramming in heart failure. Circulation 116:258–267
Ke X, Yang R, Wu F, Wang X, Liang J, Hu X, Hu C (2021) Exosomal miR-218-5p/miR-363-3p from endothelial progenitor cells ameliorate myocardial infarction by targeting the p53/JMY signaling pathway. Oxid Med Cell Longev 2021:5529430
Chen J, Tang Z, Chen Z, Wei Y, Liang H, Zhang X, Gao Z, Zhu H (2023) MicroRNA-218-5p regulates inflammation response via targeting TLR4 in atherosclerosis. BMC Cardiovasc Disord 23:122
Wu H, Zhang J (2018) miR-126 in peripheral blood mononuclear cells negatively correlates with risk and severity and is associated with inflammatory cytokines as well as intercellular adhesion molecule-1 in patients with coronary artery disease. Cardiology 139:110–118
Gao W, Cui H, Li Q, Zhong H, Yu J, Li P, He X (2020) Upregulation of microRNA-218 reduces cardiac microvascular endothelial cells injury induced by coronary artery disease through the inhibition of HMGB1. J Cell Physiol 235:3079–3095
Elsied MA, Sharawi ZW, Al-Amrah H, Hegazy RA, Mohamed AE, Saleh RM, El-kholy SS, Farrag FA, Fayed MH, El-Magd MA (2023) Walnut kernel oil and defatted extracts enhance mesenchymal stem cell stemness and delay senescence. Molecules 28:6281
Krychtiuk KA, Lenz M, Hohensinner P, Distelmaier K, Schrutka L, Kastl SP, Huber K, Dostal E, Oravec S, Hengstenberg C et al (2021) Circulating levels of proprotein convertase subtilisin/kexin type 9 (PCSK9) are associated with monocyte subsets in patients with stable coronary artery disease. J Clin Lipidol 15:512–521
Small EM, Sutherland LB, Rajagopalan KN, Wang S, Olson EN (2010) MicroRNA-218 regulates vascular patterning by modulation of Slit-Robo signaling. Circ Res 107:1336–1344
Tang Y, Li SL, Hu JH, Sun KJ, Liu LL, Xu DY (2020) Research progress on alternative non-classical mechanisms of PCSK9 in atherosclerosis in patients with and without diabetes. Cardiovasc Diabetol 19:33
Wu N-Q, Shi H-W, Li J-J (2022) Proprotein convertase subtilisin/kexin type 9 and inflammation: an updated review. Front Cardiovasc Med 9:763516
Cerda A, Bortolin RH, Manriquez V, Salazar L, Zambrano T, Fajardo CM, Hirata MH, Hirata RDC (2021) Effect of statins on lipid metabolism-related microRNA expression in HepG2 cells. Pharmacol Rep 73:868–880
Zambrano T, Hirata RDC, Hirata MH, Cerda Á, Salazar LA (2015) Altered microRNome profiling in statin-induced HepG2 cells: a pilot study identifying potential new biomarkers involved in lipid-lowering treatment. Cardiovasc Drugs Ther 29:509–518
Ubilla CG, Prado Y, Angulo J, Obreque I, Paez I, Saavedra N, Saavedra K, Zambrano T, Salazar LA (2021) MicroRNA-33b is a potential non-invasive biomarker for response to atorvastatin treatment in chilean subjects with hypercholesterolemia: a pilot study. Front Pharmacol 12:674252
Saavedra K, Leal K, Saavedra N, Prado Y, Paez I, Ubilla CG, Rojas G, Salazar LA (2022) MicroRNA-20a-5p downregulation by atorvastatin: a potential mechanism involved in lipid-lowering therapy. Int J Mol Sci 23:5022
Zambrano T, Hirata RDC, Hirata MH, Cerda Á, Salazar LA (2018) Statins differentially modulate microRNAs expression in peripheral cells of hyperlipidemic subjects: a pilot study. Eur J Pharm Sci 117:55–61
Davaro F, Forde SD, Garfield M, Jiang Z, Halmen K, Tamburro ND, Kurt-Jones E, Fitzgerald KA, Golenbock DT, Wang D (2014) 3-Hydroxyl-3-methylglutaryl coenzyme A (HMG-CoA) reductase inhibitor (statin)-induced 28-kDa interleukin-1β interferes with mature IL-1β signaling. J Biol Chem 289:16214–16222
Schönbeck U, Libby P (2004) Inflammation, immunity, and HMG-CoA reductase inhibitors: statins as antiinflammatory agents? Circulation 109:Ii18-26
Nissen SE, Tuzcu EM, Schoenhagen P, Crowe T, Sasiela WJ, Tsai J, Orazem J, Magorien RD, O’Shaughnessy C, Ganz P (2005) Statin therapy, LDL cholesterol, C-reactive protein, and coronary artery disease. N Engl J Med 352:29–38
Careskey HE, Davis RA, Alborn WE, Troutt JS, Cao G, Konrad RJ (2008) Atorvastatin increases human serum levels of proprotein convertase subtilisin/kexin type 9. J Lipid Res 49:394–398
Boyd JH, Fjell CD, Russell JA, Sirounis D, Cirstea MS, Walley KR (2016) Increased plasma PCSK9 levels are associated with reduced endotoxin clearance and the development of acute organ failures during sepsis. J Innate Immun 8:211–220
Tang ZH, Peng J, Ren Z, Yang J, Li TT, Li TH, Wang Z, Wei DH, Liu LS, Zheng XL et al (2017) New role of PCSK9 in atherosclerotic inflammation promotion involving the TLR4/NF-κB pathway. Atherosclerosis 262:113–122
Yang SS, Li R, Qu X, Fang W, Quan Z (2012) Atorvastatin decreases toll-like receptor 4 expression and downstream signaling in human monocytic leukemia cells. Cell Immunol 279:96–102
Shen DZ, Xin SL, Chen C, Liu T (2013) Effect of atorvastatin on expression of TLR4 and NF-κB p65 in atherosclerotic rabbits. Asian Pac J Trop Med 6:493–496
Kang R, Chen R, Zhang Q, Hou W, Wu S, Cao L, Huang J, Yu Y, Fan X-G, Yan Z et al (2014) HMGB1 in health and disease. Mol Aspects Med 40:1–116
Funayama A, Shishido T, Netsu S, Narumi T, Kadowaki S, Takahashi H, Miyamoto T, Watanabe T, Woo C-H, Abe J-I et al (2013) Cardiac nuclear high mobility group box 1 prevents the development of cardiac hypertrophy and heart failure. Cardiovasc Res 99:657–664
Andersson U, Wang H, Palmblad K, Aveberger AC, Bloom O, Erlandsson-Harris H, Janson A, Kokkola R, Zhang M, Yang H et al (2000) High mobility group 1 protein (HMG-1) stimulates proinflammatory cytokine synthesis in human monocytes. J Exp Med 192:565–570
Kalinina N, Agrotis A, Antropova Y, DiVitto G, Kanellakis P, Kostolias G, Ilyinskaya O, Tararak E, Bobik A (2004) Increased expression of the DNA-binding cytokine HMGB1 in human atherosclerotic lesions. Arterioscler Thromb Vasc Biol 24:2320–2325
Wahid A, Chen W, Wang X, Tang X (2021) High-mobility group box 1 serves as an inflammation driver of cardiovascular disease. Biomed Pharmacother 139:111555
Koushki K, Shahbaz SK, Mashayekhi K, Sadeghi M, Zayeri ZD, Taba MY, Banach M, Al-Rasadi K, Johnston TP, Sahebkar A (2021) Anti-inflammatory action of statins in cardiovascular disease: the role of inflammasome and toll-like receptor pathways. Clin Rev Allergy Immunol 60:175–199
Han QF, Wu L, Zhou YH, Wang LH, Zhang DY, Liu T, Yao HC (2015) Simvastatin protects the heart against ischemia reperfusion injury via inhibiting HMGB1 expression through PI3K/Akt signal pathways. Int J Cardiol 201:568–569
Lv ZH, Phuong TA, Jin SJ, Li XX, Xu M (2017) Protection by simvastatin on hyperglycemia-induced endothelial dysfunction through inhibiting NLRP3 inflammasomes. Oncotarget 8:91291–91305
Karlson BW, Wiklund O, Palmer MK, Nicholls SJ, Lundman P, Barter PJ (2016) Variability of low-density lipoprotein cholesterol response with different doses of atorvastatin, rosuvastatin, and simvastatin: results from VOYAGER. Eur Heart J Cardiovasc Pharmacother 2:212–217
Ahmad Z (2014) Statin intolerance. Am J Cardiol 113:1765–1771
Guan ZW, Wu KR, Li R, Yin Y, Li XL, Zhang SF, Li Y (2019) Pharmacogenetics of statins treatment: efficacy and safety. J Clin Pharm Ther 44:858–867
Ridker PM, Mora S, Rose L (2016) Percent reduction in LDL cholesterol following high-intensity statin therapy: potential implications for guidelines and for the prescription of emerging lipid-lowering agents. Eur Heart J 37:1373–1379
Khera A (2012) Statins, plasma proprotein convertase subtilisin/kexin type 9 concentrations, and LDL lowering. Clin Chem 58:6–7
Acknowledgements
Not applicable.
Funding
Authors did not receive any external fund.
Author information
Authors and Affiliations
Contributions
Conceptualization: NS & MS, methodology: DD & ME, Formal analysis and investigation: DD, DS, MS, writing—original draft preparation: DD, NS, DS, MS Writing—review and editing: NS, MS, supervision, validation and final editing: NS, DS, ME, MS, all authors commented on previous versions of the manuscript, All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The ethics committee of Kafrelsheikh University gave their approval for the study (Approval Code: MKSU 50-3-5). The study complied with the ethical standards of the Helsinki Declaration of the World Medical Association in 2013. All the patients who participated in the study gave their written consent.
Consent for publication
Not applicable.
Competing interests
Dr Mohammed A. El‑Magd is a co-author of this study and an associate editor of the journal. He has not been involved in handling this manuscript during the review process. The rest of the authors have no conflict of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Desouky, D.A., Nosair, N.A., Sherif, D.E. et al. Association between circulatory microRNA-218 expression, serum PCSK9 levels, inflammatory markers, and monocyte subsets in coronary artery disease patients: impact of statin therapy. Beni-Suef Univ J Basic Appl Sci 13, 52 (2024). https://doi.org/10.1186/s43088-024-00515-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43088-024-00515-8