
Abstract
Background
Adipose tissue secretes adiponectin (ADIPOQ), a hormone related to fat oxidation, glucose metabolism, and reproduction. The polymorphism of adiponectin is associated with productive traits in domestic animals. Thus, this study investigated the association of adiponectin gene polymorphism with lipid profile and reproductive hormones in Awassi ewe. In this study, 200 ewes between the ages of 2.5 and 5 years, neither pregnant nor lactating, were included. To determine the lipid profile and reproductive hormones, sera were separated from the blood. DNA extraction, genotyping, and sequencing reactions were used to verify the variants in the amplified fragments (exon 1).
Results
Three genotypes, CC, CA, and AA, were identified from 368 bp amplicons (exon 1). A sequencing reaction revealed a novel mutation, c.198473337C > A, in the CA genotype. The results revealed significant differences (P ≤ 0.05) in cholesterol and HDL levels in the AA genotype than CC and CA genotypes. The AA genotype had higher estradiol and progesterone levels (50.52 ± 0.64) (pg/ml) and (7.10 ± 0.04) (ng/ml), respectively, than those with the CC and CA genotypes.
Conclusions
These results conclude that the ADIPOQ gene affects lipid profiles and sex hormone levels in Awassi sheep. Choosing sheep that are polymorphic for the ADIPOQ gene should be a future study, as this gene could be linked to high prolificacy.
Similar content being viewed by others


1 Background
Adiponectin gene (ADIPOQ) is positioned on chromosome 1 (BTA) in cattle [1], and chromosome 1q27 in sheep with three exons and two introns (NCBI Reference Sequence NC_019480.2). This gene-encoded adiponectin protein mainly controls the endocrine function of adipose tissues and plays a central role in energy homeostasis [2, 3]. Moreover, Hadley et al. [4] report that it stimulates the production of ovarian steroidogenesis in granulosa and theca cells and has been found in ovarian cells in various species, including dairy cows [5]. Furthermore, adiponectin reduces lipid accumulation and is negatively correlated with triglycerides, while good cholesterol (HDL-c) is positively related [6, 7]. Adiponectin performs these functions by mediating several tissue-specific signaling pathways. Through its effects on the AMP-activated protein kinase (AMPK), adiponectin enhanced fatty acid oxidation and glucose uptake in skeletal muscles, thus contributing to carcass traits [8]. Moreover, adiponectin activates the AMPK pathway to affect oocyte nutrition and reproductive traits [9]. Polymorphisms of adiponectin have been documented in domestic animals with economic traits. Adiponectin gene polymorphism has been confirmed to affect ribeye muscle area, fat thickness, and marbling in Angus cattle [10]. In addition, the adiponectin gene is found to be highly associated with carcass and meat quality traits in Qinchuan cattle with CD genotypes exhibiting higher slaughter weight, subcutaneous fat thickness, and back fat thickness [11]. Genetic variants (g.81966235CNT, g.81966377 TNC, and g.81966364DNI) of the ADIPOQ significantly affected marbling score and carcass traits in Hanwoo cattle [12]. A novel BC140488:m.832T > A polymorphism in the 3'UTR within the goat adiponectin gene has been associated with growth traits [13]. Moreover, in New Zealand Romney lambs, haplotypes of the adiponectin gene have been associated with growth and carcass traits [14], suggesting that this gene is a potential candidate for animal productive traits. A limited amount of research has been conducted on the genetic polymorphism of the ADIPOQ and its association with livestock reproductive traits. One study examined polymorphisms of the ADIPOQ concerning reproductive performance and litter size in domestic porcine. A novel SNP (c. 1138G > A) is associated with litter size in the Wannan Black pig, a breed of pig found in China [15]. Further, the ADIPOQ/TasI genotypes revealed that TT genotype showed a shorter calving interval, a longer lactation period, and a higher milk yield in comparison with CC and CT genotypes [1].
In light of the aforementioned studies, only a few studies have investigated the relationship between genetic polymorphisms of the ADIPOQ gene and productive traits in sheep, and no studies have investigated their correlation with lipid profile and reproductive hormones in Awassi sheep. Therefore, this study evaluated the genetic diversity and polymorphism of the ADIPOQ gene and examined how ADIPOQ polymorphism influences the levels of lipids and reproductive hormones in Awassi ewes.
2 Methods
2.1 Animals
The study was conducted Al-Qasim Green University between July 2020 and March 2021 according to international guidelines for animal care and use, with approval number Agri, No. 020,7,18. A total of 200 Awassi ewes that were sexually mature, not pregnant or lactating, between the ages of 2.5 and 5 years were included in this study. A random sampling of sheep stations was conducted in Babylon and Karbala. Animals were fed seasonal grass and concentrate food (2.5% of their live body weight) daily, which consisted of barley (59%), bran (40%), salt (1%), and freshwater. Blood was collected from the jugular vein of the sheep before it was fed in the morning. Serum was stored at − 20 °C and then used in biochemical analysis.
2.2 Biochemical analysis
A serum sample was separated at 2000 ×g for 15 min at room temperature to measure the lipid profile and hormonal levels. RANDOX Laboratories’ kits were used to measure triglycerides, HDL-c, and total cholesterol in serum. LDL-c was calculated using Friedewald et al.’s equation [16]. ELISA kits were used to measure reproductive hormones (catalog numbers E0047Sh, E0015Sh, E0105Sh, and E0106Sh) from Bioassay Technology Laboratory.
2.3 DNA extraction and PCR amplification
A rapid salting-out technique was used to extract genomic DNA from whole blood [17]. DNA extracts were analyzed with a Nanodrop (Biodrop, UK) for DNA amplification. An amplification reaction contains 10 pmols of each primer, 50 ng of genomic DNA, 50 mM Tris-HCl (pH 9), 30 mM KCl, 1.5 mM MgCl2, and one unit of top DNA polymerase. Based on the GenBank ovine sequence (NC_019480.2), primers were designed for the ADIPOQ gene (exon 1, 368 bp). The sequences of the forward and reverse primers were 5′-CCTGTATCTCTCCCCACCCT-3′ and 5′-GTGTGATGCCTGCAGCTCTA-3′, respectively. An initial denaturation of the PCR product was carried out at 94 °C for 4 min, an annealing step at 57 °C for 30 s, an extension step at 72 °C for 30 s, and finally a final extension step at 72 °C for 10 min [18]. PCR products were visualized using a 2% agarose gel and UV light [19].
2.4 Genotyping and sequencing reaction
The genotyping step was performed following PCR using PCR-SSCP [20]. Denaturing-loading buffer SSCP was applied equally to each amplification product. The denaturated sample was loaded onto neutral polyacrylamide gels after seven minutes of denaturation and 10 min of cooling on wet ice using a 0.5 TBE buffer. A constant current and voltage of 200 mA and 100 V were used during electrophoresis for 4 h at room temperature. The gels were stained using the method described by Byun et al. [21]. Using Macrogen Geumcheon (Korea) and BioEdit 7.1 (DNASTAR, Madison), polymorphisms in each genotype were sequenced, edited, and visualized using SnapGene Viewer 4.0.4 (http://www.snapgene.com). Additionally, the Ensemble genome browser 96 checked the ADIPOQ gene for new variants.
2.5 Statistical analysis
PopGen32, version 1.31, was used for genetic analysis [22]. The following model was used to analyze the relationships between ADIPOQ genotypes and traits of interest using SPSS (version 23.0), which was compared with a Tukey–Kramer test:
where Yijk = traits studied, μ = overall mean, Gi = fixed effect of ith genotypes (i = CC,CA,AA), Pj = fixed effect of jth parity (j = 1, 2, 3, 4), Ak = fixed effect of kth age group (2.5–3.5, > 3.5–5), and eijk = random error. A preliminary statistical analysis showed no significant impact of interactions between variables, season, and nutrition on the studied traits.
3 Results
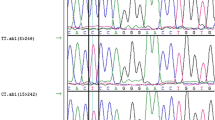
All 200 samples were investigated with PCR (Fig. 1). Based on PCR-SSCP banding patterns, three distinct genotypes CC, CA, and AA were identified (Fig. 2). Sequence analysis revealed that the c.198473337 C > A SNP was present in only one SSCP variant, indicating heterogeneity in exon 1. ADIPOQ genetic diversity indicated the most common genotype was CC with a total frequency of 0.48 (n = 97). The CA genotype was detected with a frequency of 0.33 (n = 66), followed by the AA genotype with a frequency of 0.19 (n = 37) (Fig. 3).

Agarose gel electrophoresis for PCR products amplification of the ADIPOQ gene. Lane M: 100 bp DNA ladder; lanes 1–25 indicate samples of ewes. Electrophoresis conditions: 1.5% agarose concentration, electrophoresis time 25 min. The buffer used TBE pH 8.3

SSCP non-denaturing polyacrylamide gel electrophoresis of ovine ADIPOQ gene (exon 1) PCR fragments, which appeared three PCR-SSCP banding patterns, CC, CA, and AA in Awassi ewes

Genetic diversity of the ADIPOQ gene exhibited the most preponderant genotype was CC genotype
Concerning the association analysis, the results refer to the significant differences (P ≤ 0.05) in cholesterol and HDL levels in the AA genotype than CC and CA genotypes, as shown in Table 1. Regarding the effects of ADIPOQ genotypes on reproductive hormones, the three genotypes showed significant differences (P ≤ 0.05) in sex hormone levels (Table 2). The AA genotype had higher estradiol and progesterone levels (50.52 ± 0.64) (pg/ml) and (7.10 ± 0.04) (ng/ml), respectively, than CC and CA genotypes.
4 Discussion
Varieties of regions of the ADIPOQ gene have been reported to contain genetic polymorphisms. Genetic polymorphism has been observed in the promoter region of ADIPOQ in Angus cattle, as reported by Morsci et al. [10]. Shin and Chung [23] identified three genotypes of the ADIPOQ gene promoter in the Hanwoo (Korean) cattle population using PCR–RFLP: CC, CT, and TT. Lan et al. [12] identified 13 SNPs within exon 1 of the ADIPOQ gene in New Zealand Romney lambs. In light of the above studies, there have been no studies conducted on the genotyping of the ADIPOQ gene in Awassi sheep. Marker-assisted selection programs can benefit from these genotypic data and associations.
The AA genotype was significantly (P ≤ 0.05) associated with higher cholesterol and HDL levels as compared to the CC and CA genotypes. This indicates that this genotype is related to lipid metabolism. It has been shown that adiponectin has several functions, such as controlling energy metabolism, insulin sensitivity, and regulating lipid levels [24]. Moreover, ADIPOQ levels are associated with lipoprotein metabolism, in particular, triglyceride and HDL metabolism [7]. Adiponectin increases HDL levels and lowers TG levels [25]. This hormone may raise HDL levels by increasing lipoprotein lipase (LPL) and the transporter of ATP while lowering hepatic lipase. By increasing LPL activity and VLDL receptor, as well as decreasing apo-CIII, VLDL catabolism might be increased, resulting in a reduction in serum TG [7]. Furthermore, adiponectin exhibits distinct anti-atherosclerosis properties, including reducing macrophage scavenger receptors and increasing cholesterol clearance [26]. According to Tang et al. [27], the ADIPOQ gene is implicated in lipid metabolism by inhibiting lipid synthesis and promoting fatty acid oxidation. Most research on the ADIPOQ gene has focused on humans and rodents, but fewer studies have examined livestock and poultry [28, 29]. Liu et al. [29] found that ADIPOQ gene expression levels correlated positively with intramuscular fat content during early fattening and negatively with late fattening in Shandong black cattle and Luxi cattle. This suggests that ADIPOQ appears to be an effective candidate gene for intramuscular fat accumulation and adipogenesis.
As it relates to reproductive hormones, adiponectin regulates the hypothalamus–pituitary–gonadal axis [30]. By its role in regulating hypothalamic–pituitary axis activity, adiponectin has been identified as important for FSH and LH secretion [31]. The release of progesterone (P4) and estradiol (E2) by luteal and follicular cells has been observed in response to adiponectin [32]. Moreover, the expression of adiponectin and its receptors has been demonstrated in the reproductive organs of many animals, which suggests that this hormone could affect follicular development and reproductive [5, 33]. Ovarian function is disrupted by adiponectin gene mutations that affect GnRH immunoreactive neurons [34]. Three SNPs (c. 178G > A, c. 1165A > G, and c. 1138G > A) in the porcine ADIPOQ gene have been shown to influence litter size [15]. A recent study by [1] reported that SNP 1431C > T in the ADIPOQ gene promoter has also been associated with reproductive traits in Indian dairy cattle.
Although the Awassi breed is known for its hardiness in unfavorable conditions [35], its reproduction rates are low compared to nearby breeds similar to Karakul and Assaf in the Middle East [36, 37]. Based on these findings, genetic variation within the ADIPOQ gene could contribute to breeding improvements in Awassi sheep. It is possible that the c.198473337 C > A SNP is responsible for the higher levels of reproductive hormones in the AA genotype. As the AA genotype exhibited higher levels of reproductive hormones, it tended to be more prolific than the AC or CC genotypes.
5 Conclusions
A polymorphism in the ADIPOQ gene affects the lipid profile and levels of sex hormones in Awassi sheep. The AA genotype showed a higher lipid profile and sex hormone levels, which made them more productive. This suggests that ADIPOQ is a potential candidate gene for traits associated with fat deposition in livestock. Future studies should investigate ADIPOQ gene polymorphisms and their relationship with high prolificacy in sheep.
Availability of data and materials
Data and materials are available.
Abbreviations
- ADIPOQ:
-
Adiponectin
- AMPK:
-
AMP-activated protein kinase
- HDL-c:
-
High-density lipoprotein cholesterol
- LDL-c:
-
Low-density lipoprotein cholesterol
- PCR:
-
Polymerase chain reactions
- SSCP:
-
Single-strand conformation polymorphism
- LPL:
-
Lipoprotein lipase
- FSH:
-
Follicle-stimulating hormone
- LH:
-
Luteinizing hormone
- P4:
-
Progesterone
- E2:
-
Estradiol
- GnRH:
-
Gonadotropin-releasing hormone
References
Pandey V, Nigam R, Singh SP, Sharma D, Tiwari M, Garg SK (2020) Genetic polymorphism in the adiponectin (ADIPOQ) gene and its association with the production and reproduction traits of Indian dairy cattle. Vet arhiv 90(1):16–26. https://doi.org/10.24099/vet.arhiv.0411
Kurowska P, Mlyczyńska E, Barbe A, Mellouk N, Dupont J, Rak A (2018) The adipokines in domestic animal reproduction: expression and role in the regulation of ovarian function. New Insights Theriogenol. https://doi.org/10.5772/intechopen.81035
Choi HM, Doss HM, Kim KS (2020) Multifaceted physiological roles of adiponectin in inflammation and diseases. Int J Mol Sci 21(4):1219. https://doi.org/10.3390/ijms21041219
Hadley JA, Ocón-Grove O, Ramachandran R (2020) Adiponectin is secreted by theca layer cells isolated from chicken ovarian follicles. Reproduction 159(3):275–288. https://doi.org/10.1530/REP-19-0505
Campos DB, Albornoz M, Papa PC, Palin MF, Bordignon V, Murphy BD (2015) Relationship between adiponectin and fertility in the female pig. Reprod Fertil Dev 27(3):458–470. https://doi.org/10.1071/RD13201
Natah TM (2014) Serum adiponectin levels in pre-postmenopausal metabolic syndrome women and the correlation with some physio-biochemical parameters. Aust J Basic Appl Sci 8(17):360–366
Yanai H, Yoshida H (2019) Beneficial effects of adiponectin on glucose and lipid metabolism and atherosclerotic progression: mechanisms and perspectives. Int J Mol Sci 20(5):1190. https://doi.org/10.3390/ijms20051190
Frankenberg ADV, Reis AF, Gerchman F (2017) Relationships between adiponectin levels, the metabolic syndrome, and type 2 diabetes: a literature review. Arch Endocrinol Metabol 61:614–622. https://doi.org/10.1590/2359-3997000000316
Yang W, Wang L, Wang F, Yuan S (2020) Roles of AMP-activated protein kinase (AMPK) in mammalian reproduction. Front Cell Dev Biol 8:593005. https://doi.org/10.3389/fcell.2020.593005
Morsci NS, Schnabel RD, Taylor JF (2006) Association analysis of adiponectin and somatostatin polymorphisms on BTA1 with growth and carcass traits in Angus cattle. Anim Genet 37(6):554–562. https://doi.org/10.1111/j.1365-2052.2006.01528.x
Yang YJ (2009) SNPs detection of adiponectin gene and its relationship with carcass and meat quality traits in Qinchuan cattle. Yi Chuan= Hereditas 31(10):1006–1012. https://doi.org/10.3724/sp.j.1005.2009.01006
Lan X, Wei T, Zhang L, Chen H, Hu S, Lei C, Fang XT (2009) Novel Polymorphism at the 3′ UTR of the Caprine Adiponectin Gene. Biochem Genet 47(3):251–256. https://doi.org/10.1007/s10528-009-9223-9
Choi Y, Davis ME, Chung H (2015) Effects of genetic variants in the promoter region of the bovine adiponectin (ADIPOQ) gene on marbling of Hanwoo beef cattle. Meat Sci 105:57–62. https://doi.org/10.1016/j.meatsci.2015.02.014
An Q, Zhou H, Hu J, Luo Y, Hickford JG (2017) Haplotypes of the ovine Adiponectin gene and their association with growth and carcass traits in New Zealand Romney lambs. Genes 8(6):160. https://doi.org/10.3390/genes8060160
Zhang X, Zhou N, Zhou J, Wang N, Qin X, Dong H, Ding Y, Yin Z (2016) Characterization of porcine adiponectin gene (ADIPOQ) polymorphisms and their association with litter size. Turk J Vet Anim Sci 40(2):150–156
Friedewald WT, Levy RI, Fredrickson DS (1972) Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem 18(6):499–502
Al-Shuhaib MBSA (2017) A universal, rapid, and inexpensive method for genomic DNA isolation from the whole blood of mammals and birds. J Genet 96(1):171–176. https://doi.org/10.1007/s12041-017-0750-6
Ajafar MH, Al-Thuwaini TM, Dakhel HH (2022) Association of OLR1 gene polymorphism with live body weight and body morphometric traits in Awassi ewes. Mol Biol Rep. https://doi.org/10.1007/s11033-022-07481-3
Al-Thuwaini T (2020) Association between polymorphism in BMP15 and GDF9 genes and impairing female fecundity in diabetes type 2. Middle East Fertil Soc J 25(1):1–10. https://doi.org/10.1186/s43043-020-00032-5
Imran FS, Al-Thuwaini TM, Al-Shuhaib MBS, Lepretre F (2020) A novel missense single nucleotide polymorphism in the GREM1 gene is highly associated with higher reproductive traits in Awassi sheep. Biochem Genet 59(2):422–436. https://doi.org/10.1007/s10528-020-10006-x
Byun SO, Fang Q, Zhou H, Hickford JGH (2009) An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal Biochem 385(1):174–175. https://doi.org/10.1016/j.ab.2008.10.024
Yeh FC, Yang RC, Boyle T (1999) Microsoft window-based freeware for population genetic analysis (POPGENE), ver. 1.31. University of Alberta, Canada.
Shin S, Chung E (2013) Novel SNPs in the bovine ADIPOQ and PPARGC1A genes are associated with carcass traits in Hanwoo (Korean cattle). Mol Biol Rep 40(7):4651–4660. https://doi.org/10.1007/s11033-013-2560-0
Rak A, Mellouk N, Froment P, Dupont J (2017) Adiponectin and resistin: potential metabolic signals affecting hypothalamo-pituitary gonadal axis in females and males of different species. Reproduction 153(6):R215–R226. https://doi.org/10.1530/REP-17-0002
Vergès B (2015) Pathophysiology of diabetic dyslipidaemia: where are we? Diabetologia 58(5):886–899. https://doi.org/10.1007/s00125-015-3525-8
Hafiane A, Gasbarrino K, Daskalopoulou SS (2019) The role of adiponectin in cholesterol efflux and HDL biogenesis and metabolism. Metabolism 100:153953. https://doi.org/10.1016/j.metabol.2019.153953
Tang N, Wang S, Qi J, Wu Y, Li Z (2018) Research progress on adiponectin regulating lipid metabolism. Acta Vet Zootech Sin 49(12):2550–2557
Holland WL, Xia JY, Johnson JA, Sun K, Pearson MJ, Sharma AX, Quittner-Strom E, Tippetts TS, Gordillo R, Scherer PE (2017) Inducible overexpression of adiponectin receptors highlight the roles of adiponectin-induced ceramidase signaling in lipid and glucose homeostasis. Mol Metab 6(3):267–275. https://doi.org/10.1016/j.molmet.2017.01.002
Liu R, Liu X, Bai X, Xiao C, Dong Y (2020) Different expression of lipid metabolism-related genes in Shandong black cattle and Luxi cattle based on transcriptome analysis. Sci Rep 10(1):1–14. https://doi.org/10.1038/s41598-020-79086-4
Ranjan A (2017) Adipokines as a modulator of reproductive function. J Sci Res 61:131–140
AL-Thuwaini TM, (2022) Adiponectin and its physiological function in ruminant livestock. Rev Agric Sci 10:115–122. https://doi.org/10.7831/ras.10.0_115
Dobrzyn K, Smolinska N, Kiezun M, Szeszko K, Rytelewska E, Kisielewska K, Gudelska M, Kaminski T (2018) Adiponectin: a new regulator of female reproductive system. Int J Endocrinol. https://doi.org/10.1155/2018/7965071
Merhi Z, Bazzi AA, Bonney EA, Buyuk E (2019) Role of adiponectin in ovarian follicular development and ovarian reserve. Biomed Rep 10(6):337–342. https://doi.org/10.3892/br.2019.1213
Barbe A, Bongrani A, Mellouk N, Estienne A, Kurowska P, Grandhaye J, Elfassy Y, Levy R, Rak A, Froment P, Dupont J (2019) Mechanisms of adiponectin action in fertility: an overview from gametogenesis to gestation in humans and animal models in normal and pathological conditions. Int J Mol Sci 20(7):1526. https://doi.org/10.3390/ijms20071526
Al-Thuwaini TM (2021) The relationship of hematological parameters with adaptation and reproduction in sheep: a review study. Iraqi J Vet Sci 35(3):575–580. https://doi.org/10.33899/ijvs.2020.127253.1490
Al-Thuwaini TM (2021) Novel single nucleotide polymorphism in the prolactin gene of Awassi ewes and its role in the reproductive traits. Iraqi J Vet Sci 35(3):429–435. https://doi.org/10.33899/ijvs.2020.126973.1423
Ajafar MH, Kadhim AH, AL-Thuwaini TM, (2022) The reproductive traits of sheep and their influencing factors. Rev Agric Sci 10:82–89. https://doi.org/10.7831/ras.10.0_82
Acknowledgements
In acknowledging the sheep stations that provided the Awassi ewes, the authors express their gratitude.
Funding
This study was not supported by any specific grant from funding.
Author information
Authors and Affiliations
Contributions
All authors contributed equally. In addition, all authors reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approvals and consent to participate
The study was conducted by Al-Qasim Green University between July 2020 and March 2021 according to international guidelines for animal care and use, with approval number Agri, No. 020,7,18.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
AL-Jaryan, I.L., AL-Thuwaini, T.M. & Al-Jebory, H.H. Novel variants associated with adiponectin-related traits in Awassi ewes. Beni-Suef Univ J Basic Appl Sci 11, 148 (2022). https://doi.org/10.1186/s43088-022-00328-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43088-022-00328-7